
Extended Data Fig. 9 |. Antennal lobe response to human and animal odours during pan-neuronal imaging.
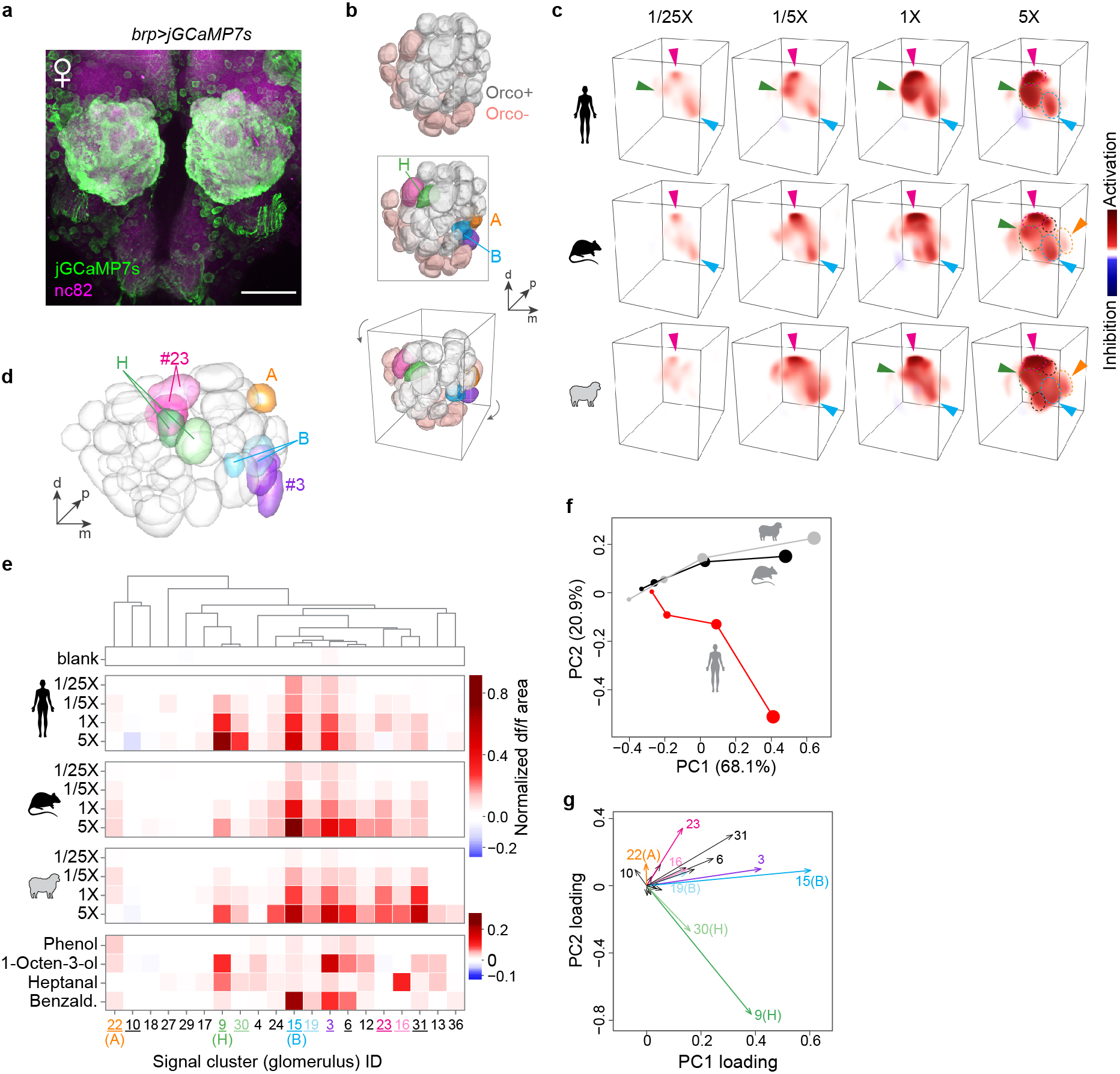
a, Antibody staining of mosquito antennal lobe in an animal expressing jGCaMP7s under the control of the brp pan-neuronal driver. All glomeruli are strongly labelled with jGCaMP7s. Scale bar 50 μm. b, AL reconstruction from confocal imaging highlighting Orco+ and Orco− glomeruli (top), five focal glomeruli discussed below (middle), and the viewing angle used in (c) (bottom). c, 3D renderings of the response of a single representative female mosquito to human, rat, and sheep odour. Dashed circles outline glomeruli that responded strongly at 5X. Arrowheads highlight key glomeruli, including an non-Orco glomerulus (magenta) adjacent to H that responded strongly to both human and animal odour in most replicate mosquitoes. d–g, Automated analysis of pan-neuronal imaging data, showing segmented antennal lobe of the reference mosquito (d), mean normalized response for all signal clusters that could be matched in the brains of at least 3 of 4 replicate mosquitoes (e), and principal components analysis of mean responses (f,g). As explained in Extended Data Fig. 2, the limited resolution of fast, volumetric imaging causes low-level bleed-through of signal from one glomerulus to adjacent regions of neighboring glomeruli, especially along the z (depth) axis. This can make the delineation of adjacent glomeruli during unsupervised segmentation imperfect, especially when all neurons express GCaMP as they do here. Some glomeruli are split into two or more initial segments (e.g. two green H segments in (d)), while others show a shadow of the response pattern of their neighbours. For example, signal cluster #30 shadows H, #19 shadows B, and #16 shadows #23 (magenta). To help readers more easily distinguish shadow signals from independent glomeruli, we have used dark and light shades of the same colour to highlight what we believe are source and shadow clusters, respectively, in (e) and (g). The overall pattern is consistent with the result from Orco+ imaging (Extended Data Fig. 2) in that four key Orco+ glomeruli contribute to host responses: B, H, A, and the 1-octen-3-ol-sensitive glomerulus posterior to B (signal cluster #3, purple). Importantly, H is still the only glomerulus that is selective for human odour. However, we now see two to three broadly host-sensitive signals coming from Orco− regions of the AL. Of these, the magenta glomerulus adjacent to H was the most consistent upon manual data inspection. It can be seen clearly in (c), but is less prominent in the automated analysis (signal cluster #23) due to the challenges of matching glomeruli across brains in this non-model species. Based on position, this glomerulus may be one of those that express the Ir8a co-receptor51,52.