
FIGURE 3.
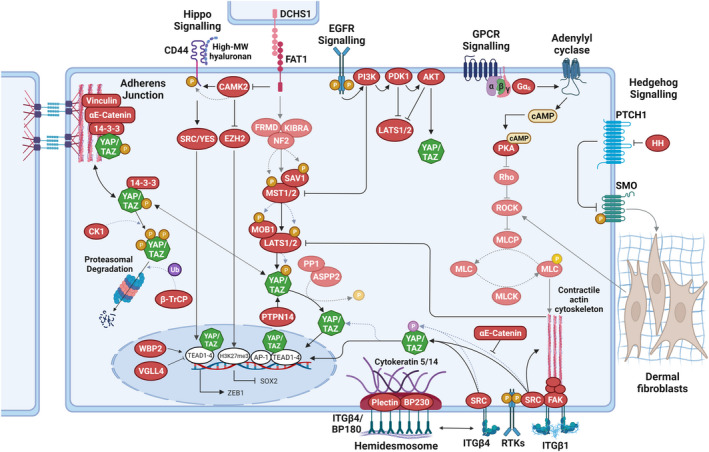
Regulation of YAP/TAZ in normal and neoplastic epidermal cells. Speculative aspects of signalling pathways that are not yet supported by experimental data are indicated by faded graphical elements. Hippo signalling via MOB1A/MOB1B and large tumour suppressor (LATS)1/LATS2 inhibits Yes‐associated protein (YAP) and transcriptional co‐activator with PDZ‐binding motif (TAZ) via serine phosphorylation (orange) to promote cytoplasmic retention and/or proteasomal degradation. Integrin (ITG)–SRC signalling in the basal epidermal cell compartment promotes YAP/TAZ nuclear localization and TEA domain (TEAD) binding. Direct phosphorylation of YAP/TAZ on tyrosine residues (pink) by SRC promotes increased nuclear localization. In the nucleus, vestigial such as family member 4 (VGLL4) competes with YAP/TAZ in binding to TEADs, while WW domain binding protein‐2 (WBP2) enhances the co‐activator functions of YAP/TAZ. The contractile F‐actin‐myosin cytoskeleton stabilizes ITGβ1 adhesions and thus contributes to SRC activation. ITGβ4 adhesions are part of hemidesmosomal complexes that are anchored to keratin 5/14 intermediate filaments. Nuclear localization of YAP in basal epidermal cells is also negatively regulated by PTPN14. At adherens junctions, α‐catenin controls YAP/TAZ activity and phosphorylation by modulating its interaction with 14–3‐3. α‐catenin can also inhibit activation of SRC by ITGβ4. EGFR signalling inactivates the Hippo pathway through the stimulation of PI3K–PDK1. G protein‐coupled receptor (GPCR) signalling, involving Gαs and PKA, suppresses LATS1/2 activation, presumably via decreasing F‐actin‐myosin cytoskeletal contractility downstream of Rho/ROCK. During cSCC progression, loss of function of the protocadherin FAT1 activates a CAMK2‐CD44‐SRC axis that promotes nuclear translocation of YAP and this drives the expression of zinc finger E‐box binding homeobox 1 (ZEB1) that stimulates the mesenchymal state. FAT1 loss of function also inactivates enhancer of zeste homologue 2 (EZH2), promoting SRY‐box transcription factor 2 (SOX2) expression, which sustains the epithelial state. Together, these molecular events promote a hybrid epithelial‐to‐mesenchymal transition (EMT) phenotype. If FAT1 can directly activate the Hippo pathway, is currently not known. In BCC, fibroblast activation and ECM remodelling in papillary dermis as a consequence of increased HH signalling in the epidermis may indirectly activate epidermal ROCK signalling through mechano‐reciprocity. AKT, Ak strain transforming; AP, activator protein; cAMP, 3′ 5′‐cyclic adenosine monophosphate; CAMK, Ca2+/calmodulin‐dependent protein kinase; CK1, casein kinase 1δ/1ε; DCHS, dachsous; EGFR, epidermal growth factor receptor; FAK, focal adhesion kinase; HH, hedgehog; MLC, myosin light chain; MLCK, myosin light chain kinase; MLCP, myosin light chain phosphatase; P, phosphorylation; PDK, pyruvate dehydrogenase kinase; PI3K, phosphoinositide 3‐kinase; PKA, protein kinase A; PP1, protein phosphatase 1; PTPN, protein tyrosine phosphatase non‐receptor type; RASSF, RAS association domain family; ROCK, Rho‐associated kinase; SFK, SRC‐family kinase; SMO, smoothened; Ub, ubiquitylation. Dotted lines indicate post‐translational modification events. Figure graphics were created with BioRender.com.