
Fig. 4.
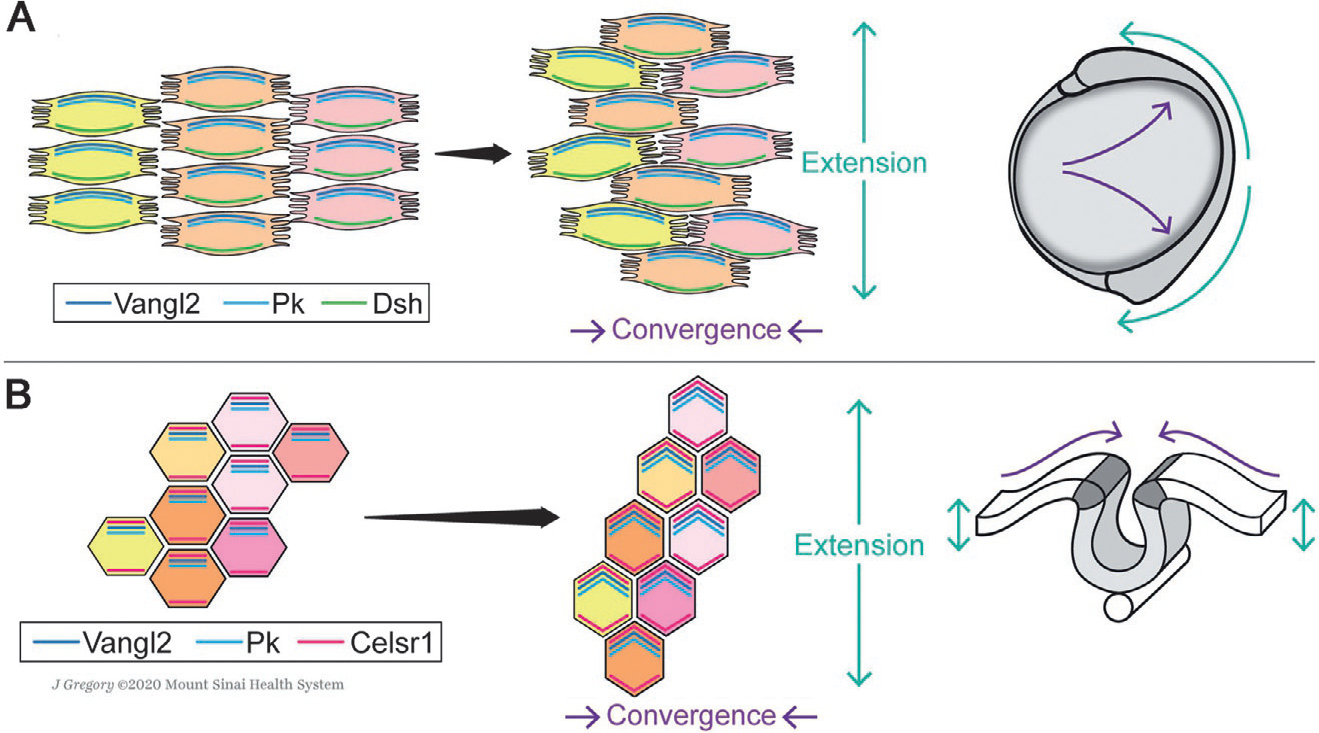
Schematic of planar cell polarity core factor localization during CE movements of gastrulation and neurulation. (A) During vertebrate gastrulation, mesendodermal cells move dorsally toward the midline of the embryo and intercalate between their neighbors along the A/P-axis (Keller et al., 2000; Tada & Heisenberg, 2012; Yin et al., 2009), narrowing the body along the medio-lateral (M/L) axis and elongate it along the anterior-posterior (A/P) axis. Bipolar protrusions emanating from mesendodermal cells at medial and lateral edges facilitate this process by allowing for stable contacts with the neighboring cells and generating traction (Keller et al., 2000; Tada & Heisenberg, 2012; Yin et al., 2009). The core PCP factors localize asymmetrically in the converging cells, as shown here schematically for Vangl2/Pk (note that there are multiple Pk factors) and Dsh (Dvl in mammals), and mediate their protrusive behavior (Roszko et al., 2015; Yin et al., 2008). (B) During neurulation, the neural folds that emerge from apical thickening of the neuroepithelium unite dorsally and close the neural tube via CE rearrangements toward the midline (Keller et al., 2000; Keller, Shih, & Sater, 1992; Nikolopoulou, Galea, Rolo, Greene, & Copp, 2017; Tada & Heisenberg, 2012). In this context, cell-cell intercalations were reported to be driven by polarized actomyosin contractions within the neuroepithelium, regulated by asymmetric core PCP factor localization. Vangl2, Pk and Celsr1/Fmi bridges the cells in the A/P axis, signaling in the apical domain (Butler & Wallingford, 2018; Ciruna et al., 2006; McGreevy, Vijayraghavan, Davidson, & Hildebrand, 2015; Nishimura et al., 2012; Ossipova, Kim, & Sokol, 2015). Anterior is up in all images. See main text for details.