

Abstract
Background
Frailty, defined as physical performance impairment, is a common condition in older adults and can anticipate the development of sarcopenia, a geriatric syndrome characterized by loss of muscle strength and mass. microRNAs (miRNAs) are short molecules of RNA endowed with the ability to modulate gene expression; miRNAs are present in serum and are considered potential biomarkers for several diseases. Serum concentration of miR-451a, miR-93-5p, miR-155-5p, miR-421-3p, miR-425-5p, miR-495-3p and miR-744-5p was recently shown to be altered in sarcopenic patients.
Methods
We verified if a particular miRNAs pattern could be detected in frailty as well by analyzing these molecules in 50 frail and 136 robust subjects. Additionally, a subgroup of these subjects (15 frail and 30 robust) underwent a 12-week program based on a multicomponent exercise protocol (VIVIFRAIL) consisting of resistance training, gait retraining, and balance training. After the program, serum miRNAs concentration was measured again, to verify whether the physical activity had an effect on their concentration. Moreover, clinical characteristics and indicators of physical performance of all subjects were compared before and after intervention to verify the effect of the VIVIFRAIL program.
Results
At the end of the multicomponent exercise program, Short Physical Performance Battery (SPPB) score as well right and left handgrip (p < 0.05) were significantly increased in frail subjects; right and left handgrip significantly were increased also in robust subjects (p < 0.05). Interestingly, the variation of SPPB was significantly higher in frail compared to robust subjects (p < 0.0001). Moreover, at the end of the program, in frail compared to robust subjects: miR-451a serum concentration was significantly increased (frail: 6.59 × 104; 1.12 × 104–2.5 × 105 c/ng; robust: 2.31 × 104; 1.94 × 103–2.01 × 105 c/ng) (p < 0.05); and 2) miR-93-5p and miR-495-3p serum concentration was reduced, whereas that of miR-155-5p was significantly increased (p < 0.05 in both cases). Serum concentration of miR-93-5p and miR-495-3p was decreased, and that of miR-155-5p was increased at the end of the program in robust subjects alone, statistical significance being reached for miR-93-5p alone (p = 0.02).
Conclusion
These results suggest that serum miR-451a should be investigated as a potential biomarker for frailty and show that the VIVIFRAIL multicomponent program modulates circulatory miRNAs expression, at least in older adults.
Keywords: microRNA, Sarcopenia, Frailty, Rehabilitation, Physical Activity, Biomarkers
Background
The higher clinical complexity that features older adults is well represented by the concept of frailty, a condition characterized by increased vulnerability to stressors and reduced homeostatic reserves [1]. Physical decline is considered the cardinal sign of frailty [2, 3]; indeed, aging is typically characterized by muscle wasting that progressively causes disability, loss of muscle function and of self-sufficiency in older adults. This phenomenon is called sarcopenia, a term that describes the age-related loss of muscle mass and muscle strength and/or function (i.e., dynapenia) [4–8]. The etiology of sarcopenia is multifactorial, involving many biological mechanisms [9, 10], like neuromuscular junction dysfunction, reduced satellite cell number/function, intramuscular adipose tissue infiltration [11] as well as chronic inflammation [12]. It has been proposed that sarcopenia may be the biological substrate for the development of physical frailty [13, 14]. However, the causal relationship between the two manifestations remains largely unknown.
Several proteins and biomolecules involved in inflammation and in oxidative stress are hypothesized to play a pathogenetic role in frailty and sarcopenia [15]. Among these biomolecules, microRNAs (miRNAs) are involved in various pathways and cellular mechanisms that are potentially associated with sarcopenia, including satellite cell regulation [16, 17] and the functionality of muscle fibers and mitochondria [18]. Notably, miRNAs have also repeatedly been associated with chronic conditions in which sarcopenia is more frequently observed, such as neurodegenerative pathologies [reviewed by 19].
miRNAs are short single-stranded RNA molecules (about 22 nucleotides) that are usually coded within introns. miRNAs recognize complementary patterns mainly in the 3′ untranslated region (3′-UTR) of target mRNAs and regulate their expression by degradation or translation blockage, acting as critical post-transcriptional mediators of cell function [20–22]. miRNAs also play a crucial role in diseases, as their ability to regulate the expression of specific genes results in the control and modulation of the activity of multiple biological pathways [reviewed by 22].
We have recently shown that serum miR-451a concentration is significantly increased in severe sarcopenia; we also observed that serum concentration of this miRNA, as well as that of miR-93-5p, miR-155-5p, miR-421-3p, miR-425-5p, miR-495-3p and miR-744-5p, is modulated by rehabilitation in severe sarcopenic patients [23]. On the basis of that finding, we analyzed whether these miRNAs are deregulated in frail subjects as well in comparison with robust subjects, and verified if an intervention program based on a multicomponent exercise protocol could modulate their expression.
Material and methods
Patients and controls
One-hundred-eighty-six individuals were included in the study: 50 (16 males and 34 females) frail subjects and 136 (43 males and 93 females) robust subjects (controls). All subjects were recruited by the Rehabilitative Unit of the IRCCS Santa Maria Nascente, Fondazione Don Gnocchi ONLUS, and by the Fondazione IRCCS Ca’ Granda, Ospedale Maggiore Policlinico, both in Milan, Italy. SPPB score (Short Physical Performance Battery) of the frail individuals ranged between 3 and 9, and was > 9 in robust subjects [24, 25].
A subgroup of enrolled subjects (15 frail and 30 robust subjects) underwent a program of physical activity at home. This intervention program of 12 weeks was based on multicomponent exercise protocol (VIVIFRAIL©) [26] consisting of resistance training, gait retraining, and balance training. Different functional capacity levels were determined based on the scores obtained from SPPB and the 6 m gait velocity test, with each leading to the recommendation of a certain customized multicomponent physical exercise program. Programs include arm and leg strength and power exercises, balance and coordination to prevent falls, flexibility and cardiovascular endurance exercises [27]. Before as well after exercise protocols, all patients and controls were characterized for: SPPB [28], Activity of Daily Living (ADL) [29], Mini-Mental State Examination (MMSE) [30], Clock Drawing test (CDT) [31] and Geriatric Depression Scale (GDS) [32]. Inclusion criteria were: age > 60 years, sufficient cognitive abilities to both provide informed consent to the study and to participate and follow the multicomponent exercise (MMSE ≥ 24), and ability to walk in autonomy. Exclusion criteria were: localized loss of strength and aphasia due to severe stroke, severe impairment of motor skills, steroid therapy, and concomitant diagnosis of neoplastic or neurodegenerative diseases.
The study was performed in accordance with the Declaration of Helsinki; an informed consent was signed by all the participants according to a protocol approved by the ethics committee of the IRCCS Fondazione Don Carlo Gnocchi ONLUS (n#9_04/04/2018).
Serum miRNAs extraction and cDNA reverse transcription
Serum was obtained from blood by centrifugation (2000 g × 10’ at room temperature). Absence of hemolysis was evaluated by visual inspection and by spectrophotometric measurement of hemoglobin absorbance at 414 nm [33]. miRNAs were semi-automatically extracted from 200 µl of serum using a column-based kit (miRNeasy serum/plasma kit, Qiagen GmbH, Hilden, Germany) by robotic workstation (Qiacube, Qiagen), according to manufacturer’s instruction. miRNAs were quantified by Qubit microRNA assay kit (Thermo Fisher, Foster City, CA, US) with a Qubit 3.0 Fluorometer (Thermo Fisher), according to manufacturer’s recommendation. Equal concentration of miRNAs was retro-transcribed in cDNA (miRCURY LNA RT kit, Qiagen) with the following protocol: 60’ at 42 °C, heat-inactivation of reverse transcriptase enzyme for 5’ at 95 °C, and a hold at 4 °C. All the variables involved in the procedure were kept consistent throughout the study to avoid variations due to sample differences and handling.
miRNAs quantification by droplet digital PCR (ddPCR)
miR-93-5p, miR-155-5p, miR-421-3p, miR-425-5p, miR-451a, miR-495-3p and miR-744-5p were quantified by droplet digital PCR (ddPCR QX200, Bio-Rad, Hercules, CA, US). Briefly, 3 µl cDNA (1:25), mixed with specific LNA™-primers (Qiagen) and ddPCR EvaGreen Supermix (Bio-Rad), were emulsified with droplet generator oil (Bio-Rad) using a QX200 droplet generator, according to manufacturer’s instruction. A no template control and a negative control for each reverse transcription reaction were included in every assay to check for lack on non-specific amplification. Droplets were transferred into a plate, heat-sealed with a piercable sealing foil sheet (PX1, PCR plate sealer, Bio-Rad), and followed by end-point amplification (10’ at 95 °C; 40 cycles: 94 °C for 30’’, 58 °C for 60’’, 10’ at 98 °C; hold at 4 °C). Plates were then loaded into the Droplet Reader (QX200 droplet reader, Bio-Rad). Each well was queried for fluorescence to determine the quantity of positive events (droplets), and the results were displayed as dot plots. Samples resulted in less than two positive droplets are considered negative [34]. The miRNAs concentration was expressed as copies/ng (c/ng) of extracted RNA. QuantaSoft software, version 1.7.4.0917 (BioRad) and QX software, version 1.2 (Bio-Rad) were used to quantified copies of miRNAs. Thresholds were determined manually for each experiment, according to the negative controls, which included a no template control.
Statistical analysis
Normally distributed data were summarized as mean ± standard deviation, whereas not-normally distributed data were summarized as median and interquartile range (IQR). Categorical data were compared using Pearson’s Chi squared test. Demographic and clinical data—MMSE excluded—were normally distributed data and the comparison between frail and robust subjects was analyzed using student’s t-test, whereas the comparison before and after the multicomponent exercise activity inside each group was analyzed using paired sample t-test. MMSE and biological data (miRNAs) were not-normally distributed and the comparison between frail and robust subjects was analyzed using Mann–Whitney U test, whereas the comparison before and after the multicomponent exercise activity inside each group was analyzed using Wilcoxon rank-sum test. Correlations were tested with Spearmen’s correlation coefficient. The p-values corresponding to ≤ 0.05 were described as statistically significant in the text. Statistical analyses were performed using commercial software (MedCalc Statistical Software, version 14.10.2, Ostend, Belgium).
Results
Clinical characteristics
Fifty frail subjects and 136 robust subjects were enrolled in the study. According to the original study plan, all 50 frail subjects and 50 robust controls were supposed to undergo the VIVIFRAIL multicomponent exercise program, but, due to COVID pandemic, this had to be modified. Thus, 15 frail and 30 robust subjects were able to complete the VIVIFRAIL program. Demographic and clinical characteristics of the study population are summarized in Table 1. Age, gender and MMSE were similar in the two groups. As per definition, SPPB was significantly lower in frail subjects compared to robust ones, before (8; 7–9 vs. 11; 11–12) as well after intervention (10; 9–11 vs. 12; 12–12) (p < 0.0001 for both comparisons). Similarly, right and left handgrip was significantly lower in frail subjects compared to robust before (right: p = 0.015; left: p = 0.03) and after intervention (right: p = 0.03; left p = 0.01). Importantly, SPPB as well as right and left handgrip significantly improved after intervention both in frail subjects (SPPB: p < 0.0001; right handgrip: p = 0.03; left handgrip: p = 0.007) and in robust subjects (p < 0.0005 for all the parameters). Interestingly, the variation of SPPB was significantly higher in frail subjects (2.1 ± 0.99) compared to robust subjects (0.55 ± 0.76) (p < 0.0001), whereas no statistical differences were observed between the two groups regarding the variation of left and/or right handgrip.
Table 1.
Demographic and clinical characteristics of frail subjects and of age-and-sex-matched robust subjects
| Frail subjects | Robust subjects |
|
|---|---|---|
| N | 50 | 136 |
| Gender (F:M) | 34:16 | 93:43 |
| Age, years | 82.42 ± 5.32 | 81.93 ± 6.15 |
|
MMSE pre-intervention |
28.00 ± 1.64 | 28.86 ± 1.31 |
|
MMSE post-intervention |
28.85 ± 1.53 | 29.40 ± 0.54 |
|
SPPB pre-intervention |
8 7–9 |
11* 11–12 |
|
SPPB post- intervention |
10 9–11¶ |
12 12–12*# |
|
ADL pre-intervention |
5.7 ± 0.4 | 5.6 ± 0.5** |
|
ADL post-intervention |
5.7 ± 0.4 | 5.6 ± 0.5** |
|
Right Handgrip pre- intervention |
19.39 ± 7.20 | 23.18 ± 8.45** |
|
Right Handgrip post-rehabilitation |
20.68 ± 5.97¶ | 26.80 ± 9.45**# |
|
Left Handgrip pre- intervention |
17.90 ± 7.65¶# | 21.12 ± 7.46** |
|
Left Handgrip post- intervention |
19.41 ± 5.72¶ | 25.36 ± 7.63**# |
|
CDT pre-intervention |
2.95 ± 1.91 | 3.86 ± 1.50* |
|
CDT post-intervention |
3.22 ± 2.22 | 4.31 ± 1.28 |
|
GDS pre-intervention |
5.47 ± 5.06 | 3.93 ± 3.77 |
|
GDS post-intervention |
4.44 ± 4.56 | 2.66 ± 2.50 |
Age, MMSE, handgrip, ADL, CDT and GDS: mean ± standard deviation; SPPB: median and Interquartile range. ADL Activity of daily living, CDT Clock Drawing Test, GDS Geriatric Depression Scale, MMSE mini mental state evaluation, SPPB Short Physical Performance Battery
*p < 0.005 Frail subjects vs. robust subjects
**p < 0.05 Frail subjects vs. robust subjects
¶p < 0.05 Frail subjects before vs. after intervention
#p < 0.05 Robust subjects before vs. after intervention
Serum concentration of the miRNAs of interest by ddPCR
Analysis of miRNAs serum concentration by ddPCR in frail and in robust subjects before intervention showed that miR-451a was significantly augmented in frail subjects (6.59 × 104; 1.12 × 104–2.5 × 105 c/ng) compared to robust (2.31 × 104; 1.94 × 103–2.01 × 105 c/ng; p = 0.04) (Fig. 1A). Concentration of miR-425-5p was higher as well in serum of frail subjects (5.25 × 104; 0.00–2.54 × 105 c/ng) compared to robust (6.33 × 103; 0.00–2.01 × 105 c/ng), even if this difference approached but did not reach statistical significance (Fig. 1B). In robust subjects, miR-425-5p serum concentration appeared to cluster into two different zones: high and low. Analyses performed on demographic and clinical (i.e. SPPB value, handgrip, sex, age) parameters did not reveal any differences between robust subjects in whom miR-425-5p concentration was either high or low.
Fig. 1.

miR-451a and miR-425-5p serum concentration in frail and in age-and sex-matched robust subjects. miR-451a (Panel A) and miR-425-5p (Panel B) serum concentration in frail subjects and in robust subjects. Horizontal bars indicate median
Serum concentration of miR-93-5p was also increased, although not significantly, in frail subjects (Fig. 2A); in these individuals alone miR-93-5p concentration was negatively correlated with SPPB scores (p = 0.04, Fig. 2B). In the frail subjects alone miR-93-5p was also significantly augmented in females (1.98 × 103; 2.33 × 102–9.17 × 104 c/ng) compared to males (4.64 × 102; 0.00–9.74 × 103 c/ng; p = 0.01) (Fig. 3), and was negatively correlated with right handgrip (p = 0.016).
Fig. 2.

miR-93-5p serum concentration in frail and in age-and sex-matched robust subjects. Panel A miR-93-5p serum concentration in frail subjects and in robust subjects. Horizontal bars indicate median. Panel B Correlation between miR-93-5p serum concentration and SPPB score in frail subjects. Regression line is represented in the figure
Fig. 3.
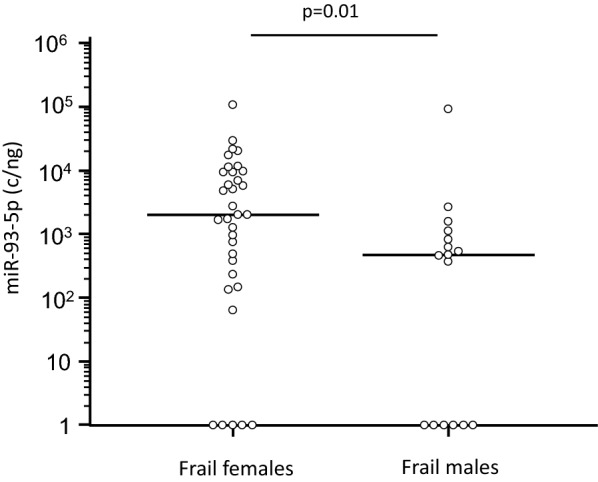
miR-93-5p serum concentration in frail subjects classified based on gender. Horizontal bars indicate median
No differences were observed regarding the other miRNAs (miR-155-5p, miR-421-3p, miR-495-3p and miR-744-5p) (Table 2) we analyzed.
Table 2.
Serum miRNAs expression in frail subjects and in robust subjects before and after intervention.
| Frail subjects before intervention | Frail subjects after intervention | Robust subjects before intervention | Robust subjects after intervention | |
|---|---|---|---|---|
|
miR-93-5p (copies/ng) |
1.04 × 103; 1.33 × 102–5.83 × 103 |
2.67 × 102; 0.00–6.23 × 102 |
6.17 × 102; 3.25 × 101–6.79 × 103 |
3.25 × 102; 0.00–9.00 × 102* |
|
miR-155-5p (copies/ng) |
0.00; 0.00–1.39 × 102 |
6.41 × 101; 0.00–1.90 × 102 |
0.00; 0.00–1.56 × 102 |
4.48 × 101; 0.00–1.50 × 102^ |
|
miR-421-3p (copies/ng) |
0.00; 0.00–3.03 × 102 |
0.00; 0.00–2.31 × 102 |
0.00; 0.00–2.15 × 102 |
0.00; 0.00–5.58 × 102 |
|
miR-425-5p (copies/ng) |
5.25 × 104; 0.00–2.54 × 105 |
2.67 × 102; 0.00–6.68 × 102 |
6.33 × 102; 0.00–2.01 × 105 |
0.00; 0.00–4.50 × 102 |
|
miR-451a (copies/ng) |
6.59 × 104; 1.12 × 104–2.54 × 105 |
6.56 × 103; 3.03 × 103–1.78 × 104 |
2.31 × 104; 1.94 × 103–2.01 × 105 |
1.20 × 104; 3.23 × 103-3.72 × 104 |
|
miR-495-3p (copies/ng) |
4.39 × 102; 2.20 × 102–1.58 × 103 |
1.17 × 102; 0.00–4.65 × 102# |
3.20 × 102; 1.83 × 102–1.25 × 103 |
3.07 × 102; 0.00–1.44 × 103* |
|
miR-744-5p (copies/ng) |
0.00; 0.00–1.08 × 102 |
0.00; 0.00–1.00 × 102 |
0.00; 0.00–1.56 × 102 |
0.00; 0.00–1.17 × 102 |
Data are reported as median and interquartile range.
*p = 0.02 Robust subjects before vs. after intervention.
#p = 0.001 Frail subjects before vs. after intervention.
^p = 0.05 Robust subjects before vs. after intervention.
Effect of intervention on serum miRNAs
We finally verified whether an intervention program of physical activity based on a multicomponent exercise protocol (VIVIFRAIL) consisting of resistance training, gait retraining, and balance training could have an effect on serum concentration of miRNAs.
miR-93-5p and miR-495-3p serum decreased at the end of the program both in frail and in robust subjects; the variation was statistically significant for both miRNAs in robust subjects (p = 0.02 for both), whereas it was statistically significant for miR-495-3p alone (p = 0.02) in frail subjects.
An opposite effect was observed for miR-155-5p, whose serum concentration increased at the end of the intervention program; the difference reached statistical significance in robust subjects (p = 0.04). Intervention did not modulate serum concentration of the other miRNAs (miR-421-3p, miR-425-5p, miR-451a and miR-744-5p) either in frail or in robust individuals (Table 2).
Discussion
In the present work we analyzed the serum miRNA concentration in frail and robust subjects who underwent a multicomponent exercise program; results showed that: (1) miR-451a is more expressed in frail compared to robust subjects; and (2) the multicomponent exercise program results on a differential effect on miRNAs expression, as it increased miR-155-5p and reduced miR-93-5p and miR-495-3p.
The age-related decline of muscle activity (loss of mass and strength) is an overlapping aspect between sarcopenia (a complex disease in which several factors—genetics, environmental, inflammation, hormones, neuromuscular degeneration) and frailty, a state of increased vulnerability to stress due decreased physiological reserve in multiple functions [35, 36]. Because in western countries life expectancy constantly increases, age-related diseases, are a major concern for public health. For this reason, the definition of biomolecules that could be used as biomarkers of physical frailty and/or possible risk of developing sarcopenia, would be extremely important.
In our previous work, serum concentration of miR-451a was found to be significantly increased in severe sarcopenic patients compared to robust subjects, suggesting this miRNA as a possible biomarker of the disease [23]. In the present work we found that serum concentration of this same miRNA is increased as well in physical frail subjects: individuals with a SPPB score lower than robust subjects but not (yet) suffering from sarcopenia.
miR-451a is coded by a gene localized at chromosome 17 (17q11.2) [37], is mainly expressed in skeletal muscle, and is known to play a key role in several human tumors [38–41]. Interestingly, miR-451a expression was observed to be increased in muscles of powerlifter [42] and of people that are low-responders to resistance exercise training [43]. miR-451a concentration in skeletal muscle was also shown to be inversely correlated to age in rhesus macaques [44], whereas its plasma concentration was inversely correlated with whole body muscle mass in middle aged men [42]. Notably, both findings indicate a possible role played by this miRNA in the age-associated decline of muscle mass and function characteristic of frailty and sarcopenia. miR-451a is also present in serum, where it can be found either inside vesicles or as non-vesicle associated protein (miRNA-protein, or miRNA-lipid/lipoprotein complexes) [40, 45–48]. Importantly, when inside vesicles miR-451a miRNA regulates innate immune response, as it was shown to suppress the production of pro-inflammatory cytokines after influenza vaccine [45] and, at least in vitro, even in the course of influenza A infection [49]. To the best of our knowledge, though, the only results suggesting an involvement of miR-451a in sarcopenia are the ones we have recently published [23], and no data are available on the possible relationship between this miRNA and frailty. Results herein indicating that plasma concentration of miR-451a is significantly increased in physical frailty suggests that this miRNA is a precocious and sensible biomarker of age-related muscle alterations that can be analyzed in easy-to-collect samples obtained with a standard venipuncture.
miR-425-3p and miR-93-5p were more expressed in serum of frail subjects as well, although without reaching statistical significance. To note, in these individuals the concentration of miR-93-5p negatively correlated with SPPB score. Thus, miR-93-5p serum concentration in physical frail subjects is very similar to that of robust individuals, and its concentration increases with the worsening of the impairment. Alteration in serum concentration of miR-425-3p (3p21.31) and miR-93-5p (7q22.1) were observed to be associated with multiple human tumors [50–53]. The mouse mmu-miR-425-3p was also found to be expressed in animal muscle tissue and to be a possible biomarker for viral myocarditis, suggesting a role for this miRNA in muscle-related pathologies. Plasma concentration of miR-93-5p, on the other hand, was recently shown to be reduced in sarcopenic patients [54]. These data are in apparent contrast with ours; this apparent discrepancy can be explained as follows: (1) we studied physical frail individuals whereas He analyzed sarcopenic patients; and (2) we analyzed miR-93-5p serum concentration by ddPCR, which quantifies in an absolute way (copies/ng) miRNA concentration, whereas He measured miR-93-5p in plasma using traditional qPCR, a method in which miRNA concentration is relative to a synthetic reference miRNA. Notably, miR-93-5p was shown to be augmented also in serum, bone tissue and bone cells of patients suffering from osteoporosis, in whom miR-93-5p concentration was correlated with bone mass density [55]. Nevertheless, studies in larger groups, analyzing miRNAs expression in serum and in plasma, and possibly including different ethnicity, are necessary to clarify this discrepancy.
All the individuals enrolled in the study underwent an intervention program based on physical activity at home. Importantly, this activity has an important positive and beneficial effects in both groups, with a significant improvement of physical condition as measured by SPPB and handgrip, in particular for frail subjects, in accordance with previous papers [reviewed by 56], indicating that this multicomponent program is a useful strategy to improve physical performance, at least in frailty.
Physical activity modulated serum expression of miR-93-5p, miR-495-3p and miR-155-5p both in frail and robust subjects. In particular: miR-495-3p and miR-93-5p were robustly down-regulated whereas miR-155-5p was up-regulated by intervention in in both groups. A very limited amount of data is available on the effect of program of intervention on circulatory miRNAs expression, and these data usually stem from analyses performed in athletes or in young people [i.e. 57, 58]. Scarce results are available on the effect of acute or intense exercise on circulatory miRNAs expression in older adults (i.e. 57, 59]. We have recently shown that rehabilitation has an effect on the expression of miRNAs that are related to SNAP-25 and IL-17 expression in severe sarcopenic patients [23, 60]. To the best of our knowledge, these are the first results showing the intervention has a significant effect on the expression of miRNA detectable in serum and possibly involved in muscle metabolism in physical frail subjects. Notably, results herein suggest that miRNAs expression is modulated by physical activity in older adults. As these proteins target several biological processes, modulation of their expression might regulate diverse mechanisms that contribute to the pathophysiology of the frailty. Although the multicomponent exercise program was similar for frail and robust subjects, and although the demographic variables were comparable between the two groups, we cannot exclude that other factors, including drugs, diet, comorbidities, etc., could have influenced the miRNAs expression.
Conclusions
To summarize, our results indicate that miR-451a expression is altered not only in sarcopenia, but in frailty as well, and that miR-93-5p, miR-155-5p and miR-495-3p can be modulated by a program of physical intervention. These data suggest that miRNAs serum concentration might be used as a biomarker in the diagnosis, the prognosis and the evaluation of physical and rehabilitative protocols in age-related muscle degenerative pathologies.
Acknowledgements
The authors thanks all the subjects enrolled in the study and all the nurses and MD for taking care of the patients who participated in the study.
Abbreviations
- ADL
Activity of daily living
- CDT
Clock Drawing test
- ddPCR
Droplet digital polymerase chain reaction
- GDS
Geriatric Depression Scale
- IADL
Instrument activity of daily living
- IL-17
Interleukin-17
- IQR
Interquartile range
- miRNA
MicroRNA
- MMSE
Mini-Mental State Examination
- PCR
Polymerase chain reaction
- SNAP25
SNARE synaptosomal-associated protein of 25 kDa
- SPPB
Short Physical Performance Battery
Author contributions
SA and RM conceived and designed the project. GAM and BA assisted with biological samples and data collection. SA, and LAC performed the experiments. SA, RM, and LAC performed the data analysis. MC supervised the project. SA and RM wrote the manuscript draft. All authors reviewed and edited the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported and funded by Fondazione Cariplo (Grant Number 2017-0622) and also by the Italian Ministry of Health—Ricerca Corrente 2021–2023. It was partially supported by grants from Fondazione Alessandro e Vincenzo Negroni Prati Morosini and Fondazione Romeo ed Enrica Invernizzi.
Availability of data and materials
The dataset generated and/or analyzed during the current study are not publicly available due to privacy or ethical restriction but are available from the corresponding author on reasonable request.
Declarations
Ethics approval and consent to participate
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of IRCCS Fondazione Don Carlo Gnocchi ONLUS (n#9_04/04/2018, date of approval: 2018/04/04). All subjects or their care-givers gave informed and written consent.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Clegg A, Young J, Iliffe S, Rikkert MO, Rockwood K. Frailty in elderly people. Lancet. 2013;381:752–762. doi: 10.1016/S0140-6736(12)62167-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fried LP, Tangen CM, Walston J, Newman AB, Hirsch C, Gottdiener J, Seeman T, Tracy R, Kop WJ, Burke G, McBurnie MA, Cardiovascular Health Study Collaborative Research Group Frailty in older adults: evidence for a phenotype. J Gerontol A Biol Sci Med Sci. 2001;56:M146–M156. doi: 10.1093/gerona/56.3.m146. [DOI] [PubMed] [Google Scholar]
- 3.Hoogendijk EO, Afilalo J, Ensrud KE, Kowal P, Onder G, Fried LP. Frailty: implications for clinical practice and public health. Lancet. 2019;394:1365–1375. doi: 10.1016/S0140-6736(19)31786-6. [DOI] [PubMed] [Google Scholar]
- 4.Clark BC, Manini TM. Sarcopenia=/= dynapenia. J Gerontol A Biol Sci Med Sci. 2008;63:829–834. doi: 10.1093/gerona/63.8.829. [DOI] [PubMed] [Google Scholar]
- 5.Clark BC, Manini TM. What is dynapenia? Nutrition. 2012;28:495–503. doi: 10.1016/j.nut.2011.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Morley JE, Abbatecola AM, Argiels JM, Baracos V, Bauer J, Bhasin S, Cederholm T, Stewart Coats AJ, Cummings SR, Evans WJ, Fearon K, Ferrucci L, Fielding RA, Guralnik JM, Harris TB, Inui A, Kalantar-Zadeh K, Kirwan BA, Mantovani G, Muscaritoli M, Newman AB, Rossi-Fanelli F, Rosano GM, Sokol GH, Storer TW, Vellas B, von Haehling S, Yeh SS, D’Anker S, Society on Sarcopenia, Cachexia and Wasting Disorders Trialis Workshop Sarcopenia with limited mobility: an international consensus. J Am Med Dir Assoc. 2011;12:403–409. doi: 10.1016/j.jamda.2011.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Baker BA. Efficacy of age-specific high-intensity stretch-shortening contractions in reversing dynapenia, sarcopenia, and loss of skeletal muscle quality. J Funct Morphol Kinesiol. 2018;3:36. doi: 10.3390/jfmk3020036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cruz-Jentoft AJ, Bahat G, Bauer J, Boirie Y, Bruyere O, Cederholm T, Cooper C, Landi F, Rolland Y, Sayer AA, Schneider SM, Sieber CC, Topinkova E, Vanderwoude M, Visser M, Zamboni M, Writing Group for the European Working Group on Sarcopenia in Older People 2 (EWGSOP2), and the Extended Group for EWGSOPS2 Sarcopenia: revised European consensus on definition and diagnosis. Age Ageing. 2019;48:16–31. doi: 10.1093/ageing/afy169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Malafrina V, Uriz-Otano F, Iniesta R, Gil-Guerrero L. Sarcopenia in the elderly: diagnosis, physiopathology and treatment. Maturitas. 2012;71:109–114. doi: 10.1016/j.maturitas.2011.11.012. [DOI] [PubMed] [Google Scholar]
- 10.Pedersen L, Hojman P. Muscle-to-organ cross talk mediated by myokines. Adipocyte. 2012;1:164–167. doi: 10.4161/adip.20344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pagano AF, Brioche T, Arc-Chagnaud C, Demangel R, Chopard A, Py G. Short-term disuse promotes fatty acid infiltration into skeletal muscle. J Cachexia Sarcopenia Muscle. 2018;9:335–347. doi: 10.1002/jcsm.12259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fulop T, Larbi A, Dupuis G, Le Page A, Frost EH, Cohen AA, Witkowski JM, Franceschi C. Immunosenescence and inflamm-aging as two side of the same coin: friends or foes? Front Immunol. 2018;8:1960. doi: 10.3389/fimmu.2017.01960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Landi F, Calvani R, Cesari M, Tosato M, Martone AM, Bernabei R. Sarcopenia as the biological substrate of physical frailty. Clin Geriatr Med. 2015;31:367–374. doi: 10.1016/j.cger.2015.04.005. [DOI] [PubMed] [Google Scholar]
- 14.Landi F, Calvani R, Cesari R, Tosato M, Martone AM, Ortolani E, Savera G, Salini S, Sisto A, Picca A, Marzetti E. Sarcopenia: an overview on current definitions, diagnosis and treatment. Curr Protein Pept Sci. 2018;19:633–638. doi: 10.2174/1389203718666170607113459. [DOI] [PubMed] [Google Scholar]
- 15.Picca A, Coelho-Junior HJ, Calvani R, Marzetti E, Vetrano DL. Biomarkers shared by frailty and sarcopenia in older adults: a systematic review and meta-analysis. Agein Res Rev. 2022;73:101530. doi: 10.1016/j.arr.2021.101530. [DOI] [PubMed] [Google Scholar]
- 16.Williams AH, Liu N, van Rooij E, Olson EN. microRNA control of muscle development and disease. Curr Opin Cell Biol. 2009;21:461–469. doi: 10.1016/j.ceb.2009.01.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Xu M, Chen X, Chen D, Yu B, Li M, He J, Huang Z. Regulation of skeletal myogenesis by microRNAs. J Cell Phyisol. 2020;235:87–104. doi: 10.1002/jcp.28986. [DOI] [PubMed] [Google Scholar]
- 18.Yin J, Qian Z, Chen Y, Li Y, Zhou X. microRNA regulatory network in the pathogenesis of sarcopenia. J Cell Mol Med. 2020;24:4900–4912. doi: 10.1111/jcmm.15197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Roy B, Lee E, Li T, Rampersaud M. Role of miRNAs in neurodegeneration: from disease cause to tools of biomarkers discovery and therapeutics. Genes (Basel) 2022;13:425. doi: 10.3390/genes13030425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843–854. doi: 10.1016/0092-8674(93)90529-y. [DOI] [PubMed] [Google Scholar]
- 21.Abdellatif M. Differential expression of microRNAs in different disease states. Circ Res. 2012;110:638–650. doi: 10.1161/CIRCRESAHA.111.247437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lu TX, Rothenberg ME. microRNA. J Allergy Clin Immunol. 2018;141:1202–1207. doi: 10.1016/j.jaci.2017.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Agostini S, Mancuso R, Costa AS, Guerini FR, Trecate F, Miglioli R, Menna E, Arosio B, Clerici M, on behalf of the SA.M.B.A. project Sarcopenia associates with SNAP-25 SNPs and a miRNAs profile which is modulated by structured rehabilitation treatment. J Transl Med. 2021;19:315. doi: 10.1186/s12967-021-02989-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cesari M, Landi F, Calvani R, Cherubini A, Di Bari M, Kortebein P, Del Signore S, Le Lain R, Vellas B, Pahor M, Roubenoff R, Bernabei R, Marzetti E, SPRINTT Consortium Rationale for a preliminary operational definition of physical frailty and sarcopenia in the SPRINTT trial. Aging Clin Exp Res. 2017;29:81–88. doi: 10.1007/s40520-016-0716-1. [DOI] [PubMed] [Google Scholar]
- 25.Marzetti E, Cesari M, Calvani R, Msihid J, Tosato M, Rodriguez-Manas L, Lattanzio F, Cherubini A, Bejuit R, Di Bari M, Maggio M, Vellas B, Dantoine T, Cruz-Jentoft AJ, Sieber CC, Freiberger E, Skalska A, Grodzicki T, Sinclair AJ, Topinkova E, Ryznarova I, Strandberg T, Schols AM, Schols JM, Roller-Wirnsberg R, Jonsson PV, Ramel A, Del Signore S, Pahor M, Roubenoff R, Bernabei R, Landi F, SPRINTT Consortium The, “Sarcopenia and Physical fRailty IN older people: multi-componenT Treatment strategies” (SPRINTT) randomized controlled trial: case finding, screening and characteristics of eligible participants. Exp Gerontol. 2018;113:48–57. doi: 10.1016/j.exger.2018.09.017. [DOI] [PubMed] [Google Scholar]
- 26.Petrella M, Aprahamian I, Mamoni RL, de Vasconcellos Romanini CF, Lima NA, de Cássio RE, da Costa DL, An VN, Aguirre BN, Galdeano JR, Fernandes IC, Soleman Hernandez SS, Cesari M, Morley JE, Izquierdo M, Oude Voshaar RC. The effect of a multicomponent exercise protocol (VIVIFRAIL©) on inflammatory profile and physical performance of older adults with different frailty status: study protocol for a randomized controlled trial. BMC Geriatr. 2021;21:83. doi: 10.1186/s12877-021-02030-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Casas-Herrero A, Anton-Rodrigo I, Zambom-Ferraresi F, Saez de Asteasu ML, Martinez-Velilla N, Elexpuru-Estomba J, Marin-Epelde I, Ramon-Espinoza F, Petidier-Torregrosa R, Sanchez-Sanchez JL, Ibanez B, Izquierdo M. Effect of multicomponent exercise programme (VIVIFRAIL) on functional capacity in frail community elders with cognitive decline: study protocol for a randomized multicenter control trial. Trials. 2019;20:362. doi: 10.1186/s13063-019-3426-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Guralnik JM, Simonsick EM, Ferrucci L, Glynn RJ, Berkman LF, Blazer DG, Scherr PA, Wallace RB. A short physical performance battery assessing lower extremity function: association with self-reported disability and prediction of mortality and nursing home admission. J Gerontol. 1994;49:M85–94. doi: 10.1093/geronj/49.2.m85. [DOI] [PubMed] [Google Scholar]
- 29.Lawton MP, Brody EM. Assessment of older people: self-maintaining and instrumental activities of daily living. Gerontologist. 1969;9:179–186. [PubMed] [Google Scholar]
- 30.Petersen RC. Mild cognitive impairment. N Engl J Med. 2011;364:2227–2234. doi: 10.1056/NEJMcp0910237. [DOI] [PubMed] [Google Scholar]
- 31.Shulman KI, Shedletsky R, Silver IL. The challenge of time: clock-drawing and cognitive function in the elderly. Int J Geriatr Psychiatry. 1986;1:135–140. [Google Scholar]
- 32.Yesavage JA, Sheikh JI. Geriatric Depression Scale (GDS) recent evidence and development of a shorter version. Clin Gerontologist. 2008;5:165–173. [Google Scholar]
- 33.Shah JS, Soon PS, Marsh DJ. Comparison of methodologies to detect low levels of hemolysis in serum for accurate assessment of serum microRNAs. PLoS ONE. 2016;11:e0153200. doi: 10.1371/journal.pone.0153200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Agostini S, Mancuso R, Costa AS, Guerini FR, Clerici M. COS-7 cells are a cellular model to monitor polyomavirus JC miR-J1-5p expression. Mol Biol Rep. 2020;47:9201–9205. doi: 10.1007/s11033-020-05862-0. [DOI] [PubMed] [Google Scholar]
- 35.Curcio F, Ferro G, Basile C, Liguori I, Parrella P, Pirozzi F, Della-Morte D, Gargiulo G, Testa G, Tocchetti CG, Bonaduce D, Abete P. Biomarkers in sarcopenia: a multifactorial approach. Exp Gerontol. 2016;85:1–8. doi: 10.1016/j.exger.2016.09.007. [DOI] [PubMed] [Google Scholar]
- 36.Liguori I, Russo G, Aran L, Bulli G, Curcio F, Della-Morte D, Gargiulo G, Testa G, Cacciatore F, Bonaduce D, Abete P. Sarcopenia: assessment of disease burden and strategies to improve outcomes. Clin Interv Aging. 2018;13:913–927. doi: 10.2147/CIA.S149232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kozomara A, Griffiths-Jones S. miRbase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014;42:D68–73. doi: 10.1093/nar/gkt1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Shen YY, Cui JY, Yuan J, Wang X. MiR-451a suppressed cell migration and invasion in non-small cell lung cancer through targeting ATF2. Eur Rev Med Pharmacol Sci. 2018;22:5554–5561. doi: 10.26355/eurrev_201809_15818. [DOI] [PubMed] [Google Scholar]
- 39.Van de Worp WRP, Schols AMW, Dingemans AMC, Op den Kamp CMH, Degens JHR, Kelders MC, Coort S, Woodruff HC, Kratassiouk G, Harel-Bellan A, Theys J, van Helvoort A, Langen RC. Identification of microRNAs in skeletal muscle associated with lung cancer cachexia. J Cachexia Sarcopenia Muscle. 2020;11:452–463. doi: 10.1002/jcsm.12512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chen J, Yao D, Chen W, Li Z, Guo Y, Zhu F, Hu X. Serum exosomal miR-451a acts as a candidate marker for pancreatic cancer. Int J Biol Markers. 2022;37:74–80. doi: 10.1177/17246008211070018. [DOI] [PubMed] [Google Scholar]
- 41.Zavesky L, Jandakova E, Weinberger V, Minar L, Hanzikova V, Duskova D, Faridova AT, Turyna R, Slanar O, Horinek A, Kohoutova M. Small non-coding RNA profiling in breast cancer: plasma U6 snRNA, miR-451a and miR-548b-5p as novel diagnostic and prognostic biomarkers. Mol Biol Rep. 2022;49:1955–1971. doi: 10.1007/s11033-021-07010-8. [DOI] [PubMed] [Google Scholar]
- 42.D’Souza RF, Bjornsen T, Zeng N, Aasen KMM, Raastad T, Cameron-Smith D, Mitchell CJ. microRNAs in muscle: characterizing the powerlifter phenotype. Front Physiol. 2017;8:383. doi: 10.3389/fphys.2017.00383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Davidsen PK, Gallagher IJ, Hartman JW, Tarnopolosky MA, Dela F, Helge JW, Timmons JA, Phillips SM. High responders to resistance exercise training demonstrate differential regulation of skeletal muscle microRNA expression. J Appl Physiol. 1985;2011(18):165–178. doi: 10.1152/japplphysiol.00901.2010. [DOI] [PubMed] [Google Scholar]
- 44.Mercken EM, Majounie E, Ding J, Guo R, Kim J, Bernier M, Mattison J, Cookson MR, Gorospe M, de Cabo R, Abdelmohsen K. Age-associated miRNA alterations in skeletal muscle form rhesus monkeys reversed by caloric restriction. Aging (Albany NY) 2013;5:692–702. doi: 10.18632/aging.100598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Okamoto M, Fukushima Y, Kouwaki T, Daito T, Kohara M, Kida H, Oshiumi H. microRNA-451a in extracellular, blood-resident vesicles attenuates macrophage and dendritic cell responses to influenza whole-virus vaccine. J Biol Chem. 2018;293:18585–18600. doi: 10.1074/jbc.RA118.003862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zhang Z, Zhang D, Cui Y, Qiu Y, Miao C, Lu X. Identification of microRNA-451a as a novel circulating biomarker for colorectal cancer. Biomed Res Int. 2020;2020:5236326. doi: 10.1155/2020/5236236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Oshiumi H. Circulating extracellular vesicles carry immune regulatory miRNAs and regulate vaccine efficacy and local inflammatory response after vaccination. Front Immunol. 2021;12:685344. doi: 10.3389/fimmu.2021.685344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zou X, Xia T, Li M, Wang T, Liu P, Zhou X, Huang Z, Zhu W. microRNA profiling in serum: potential signatures for breast cancer diagnosis. Cancer Biomark. 2021;30:41–53. doi: 10.3233/CBM-201547. [DOI] [PubMed] [Google Scholar]
- 49.Rosenberger CM, Podyminogin RL, Navarro G, Zhao GW, Askovich PS, Weiss MJ, Aderem A. miR-451 regulates dendritic cell cytokine responses to influenza infection. J Immunol. 2012;189:5965–5975. doi: 10.4049/jimmunol.1201437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Vaira V, Roncalli M, Carnaghi C, Faversani A, Maggioni M, Augello C, Rimassa L, Pressiani T, Spagnuolo G, Di Tommaso L, Fagiuoli S, Rots Caremoli E, Barberis M, Labianca R, Santoro A, Bosari S. microRNA-425-3p predicts response to sorafenib therapy in patients with hepatocellular carcinoma. Liver Int. 2015;5:1077–1086. doi: 10.1111/liv.12636. [DOI] [PubMed] [Google Scholar]
- 51.Rana S, Valbuena GN, Curry E, Bevan CL, Keun HC. MicroRNAs as biomarkers for prostate cancer prognosis: a systematic review and a systematic reanalysis of public data. Br J Cancer. 2022;126:502–513. doi: 10.1038/s41416-021-01677-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yang L, Yang T, Wang H, Dou T, Fang X, Shi L, Li X, Feng M. DNMBP-AS1 regulates NHLRC3 expression by sponging miR-93-5p/17-5p to inhibit colon cancer progression. Front Oncol. 2022;12:765163. doi: 10.3389/fonc.2022.765163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yang M, Xiao R, Wang X, Xiong Y, Duan Z, Li D, Kan Q. miR-93-5p regulates tumorigenesis and tumor immunity by targeting PD-L1/CCND1 in breast cancer. Ann Transl Med. 2022;10:203. doi: 10.21037/atm-22-97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.He N, Zhang Y, Zhang Y, Feng B, Zheng Z, Wang D, Zhang S, Ye H. Increasing fracture risk associates with plasma circulating microRNAs in aging people’s sarcopenia. Front Physiol. 2021;12:678610. doi: 10.3389/fphys.2021.678610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kelch S, Balmayor ER, Seeliger C, Verster H, Kirschke JS, van Griensven M. miRNAs in bone tissue correlate to bone mineral density and circulating miRNAs are gender independent in osteoporotic patients. Sci Rep. 2017;7:15861. doi: 10.1038/s41598-017-16113-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Nascimento CM, Ingles M, Salvador-Pascual A, Cominetti MR, Gomez-Cabrera MC, Vina J. Sarcopenia, frailty and their prevention by exercise. Free Radical Biol Med. 2019;132:42–49. doi: 10.1016/j.freeradbiomed.2018.08.035. [DOI] [PubMed] [Google Scholar]
- 57.Sapp RM, Shill DD, Roth SM, Hagberg JM. Circulating microRNAs in acute and chronic exercise: more than mere biomarkers. J Appl Physiol. 1985;2017(122):702–717. doi: 10.1152/japplphysiol.00982.2016. [DOI] [PubMed] [Google Scholar]
- 58.Barber JL, Zellars KN, Barringhaus KG, Bouchard C, Spinale FG, Sarzynski MA. The effects of regular exercise on circulating cardiovascular-related microRNAs. Sci Rep. 2019;9:7527. doi: 10.1038/s41598-019-43978-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Margolis LM, Lessard SJ, Ezzyat Y, Fielding RA, Rivas DA. Circulating microRNA are predictive of aging and acute adaptive response to resistence exercise in men. J Gerontol A Biol Sci Med Sci. 2017;72:1319–1326. doi: 10.1093/gerona/glw243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.La Rosa F, Agostini S, Saresella M, Costa AS, Piancone F, Miglioli R, Trecate F, Clerici M. Deregulation of IL-37 and its miRNAs modulators in sarcopenic patients after rehabilitation. J Transl Med. 2021;19:172. doi: 10.1186/s12967-021-02830-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The dataset generated and/or analyzed during the current study are not publicly available due to privacy or ethical restriction but are available from the corresponding author on reasonable request.