

Abstract
Lipid nanoparticles (LNPs) have delivered siRNA and mRNA drugs in humans, underscoring the potential impact of improving the therapeutic window of next-generation LNPs. To increase the LNP therapeutic window, we applied lessons from small-molecule chemistry to ionizable lipid design. Specifically, given that stereochemistry often influences small-molecule safety and pharmacokinetics, we hypothesized that the stereochemistry of lipids within an LNP would influence mRNA delivery. We tested this hypothesis in vivo using 128 novel LNPs that included stereopure derivatives of C12-200, an ionizable lipid that when formulated into LNPs delivers RNA in mice and non-human primates but is not used clinically due to its poor tolerability. We found that a novel C12-200-S LNP delivered up to 2.8-fold and 6.1-fold more mRNA in vivo than its racemic and C12-200-R controls, respectively. To identify the potential causes leading to increased delivery, we quantified LNP biophysical traits and concluded that these did not change with stereochemistry. Instead, we found that stereopure LNPs were better tolerated than racemic LNPs in vivo. These data suggest that LNP-mediated mRNA delivery can be improved by designing LNPs to include stereopure ionizable lipids.
Graphical Abstract
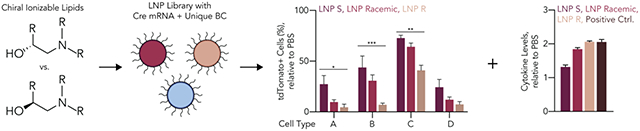
Introduction
In humans, lipid nanoparticles (LNPs) have delivered therapeutic siRNA and mRNA to the liver after systemic administration1, 2 and mRNA to antigen-presenting cells after intramuscular administration3, 4. LNPs are typically created by mixing5 an aqueous solution containing the nucleic acid with an organic solution containing four components: an ionizable or cationic lipid, cholesterol, a poly(ethylene glycol) (PEG)-lipid, and a “helper” lipid, which in the three Food and Drug Administration (FDA)-approved LNPs1, 3, 4 is 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC)6. Using this approach, scientists have varied the structure of the cationic or ionizable lipid using rational design7 or combinatorial chemistry8. These efforts revealed that ionizable lipid tolerability could be increased by including hydrolysable ester bonds, which increase LNP biodegradability9. This design rule has been incorporated into all three FDA- or EUA-approved LNP RNA drugs, which contain lipids with ester bonds. However, several lipids that have delivered RNA in non-human primates10, 11 contain nondegradable bonds and may lose activity if their chemical composition is changed to include esters. In addition, the chemical diversity of future lipids could be expanded if ester bonds were not required to achieve tolerability.
We therefore sought an ester-independent design rule to improve ionizable lipid tolerability. We focused on stereochemistry, since small-molecule enantiomers can possess different pharmacokinetic, pharmacodynamic, and safety properties12, 13. However, while the role of stereochemistry in small molecules is commonly investigated before drugs are FDA approved, the question of whether the stereochemistry of lipids influences LNP delivery remains understudied, with leading ionizable lipids containing stereocenters (Supplementary Figure 1).
Results and Discussion
An ideal lipid to test the hypothesis that stereopurity could improve LNP tolerability would be efficacious but limited by toxicity. We therefore focused on the lipid C12-200, which delivered siRNA to hepatocytes in mice at doses as low as 0.01 mg kg−1 11, delivered siRNA in NHPs11, 14, and has delivered siRNA or mRNA in pre-clinical disease models14–17. However, C12-200 LNPs also led to liver necrosis in mice18. We first synthesized, purified, and characterized stereopure C12-200 (Supplementary Figure 2). Specifically, we reacted N1-(2-(4-(2-aminoethyl)piperazin-1-yl)ethyl)ethane-1,2-diamine with chiral epoxides to create two distinct C12-200 lipids: one with five R stereocenters, termed C12-200-R, and one with five S stereocenters, termed C12-200-S (Supplementary Figure 1). We purified the products using a silica gel, achieving yields between 53 and 62%. To control for the possibility that results would be specific to only one LNP, we created a systematic panel of related lipids using two approaches. First, we synthesized six additional stereopure ionizable lipids by varying lipid tail length; thus, we synthesized C9R, C10R, C12R, and C13R as well as C9S, C10S, C12S, and C13S, creating a total of eight stereopure ionizable lipids (Figure 1A,B and Supplementary Figure 2). Second, since LNPs are formulated with a mixture of ionizable lipid, cholesterol, PEG-lipid, and helper lipid, we formulated each ionizable lipid 16 ways, creating 128 distinct LNPs (Figure 1C,D). Specifically, we mixed (i) one of the eight stereopure ionizable lipids with (ii) either cholesterol or 20-alpha hydroxycholesterol (20α-OH), (iii) the PEG-lipid C14PEG2000, and (iv) 1,2-dioleoyl-snglycero-3-phosphoethanolamine (DOPE) at eight different molar ratios (Figure 1C,D). We chose these constituents since cholesterol, C14PEG2000, and DOPE were used in the optimized formulation of racemic C12-200 for mRNA delivery19, and because these components are used in all three FDA- or EUA-approved LNP–RNA drugs1, 3, 4. In all cases, the LNP: nucleic acid mass ratio was 10:1.
Figure 1.
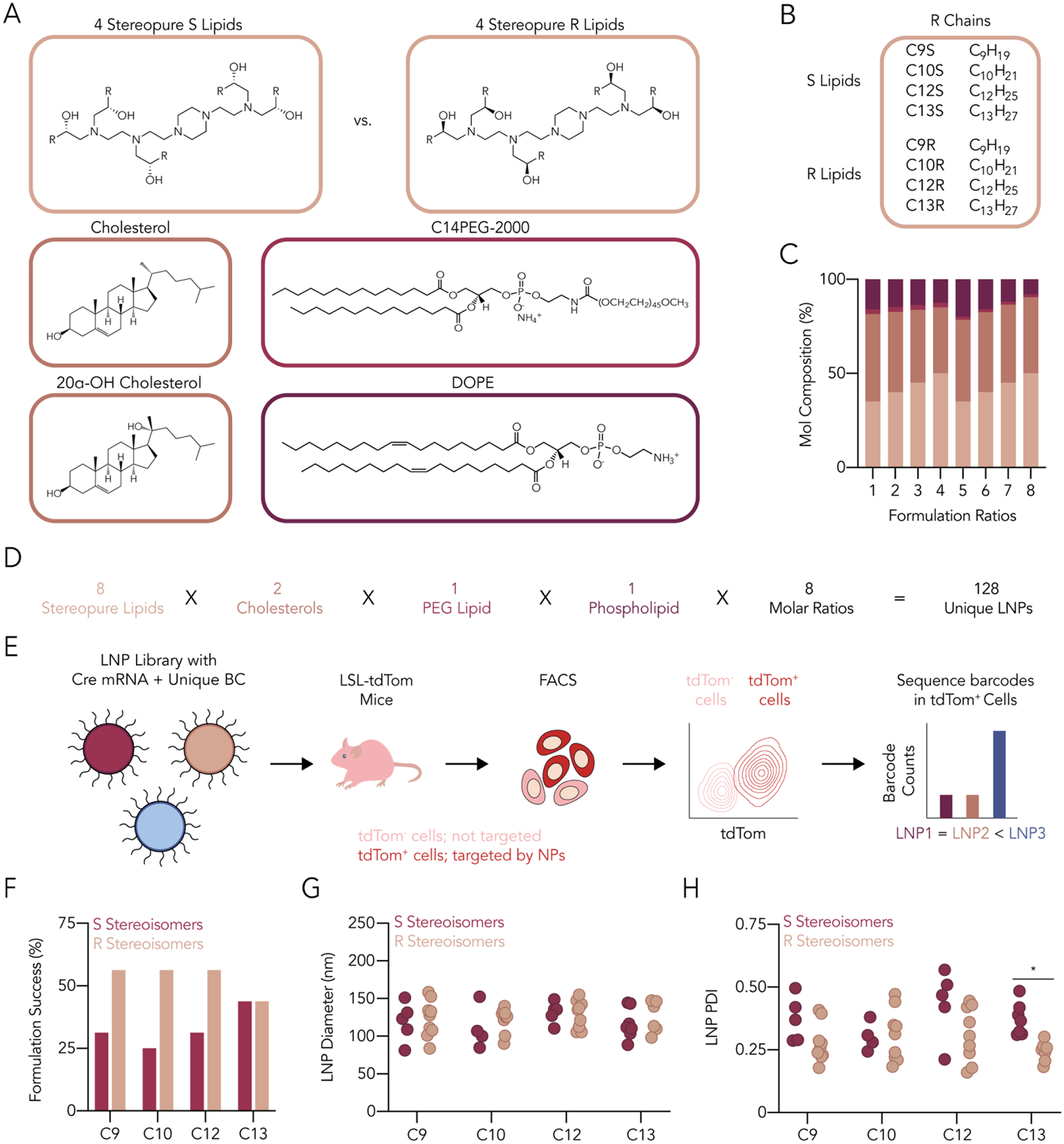
Designing and characterizing a library of LNPs containing stereopure ionizable lipids. (A) Lipid nanoparticle constituents consisted of a combination of stereopure R and S ionizable lipids, cholesterol and an oxidized cholesterol, a PEG-lipid, and a phospholipid. (B) We used eight stereopure ionizable lipids, four for each enantiomer having different hydrocarbon tail lengths. (C) LNP chemical constituents were then mixed at eight different molar ratios. (D) The resulting LNP library contained 128 nanoparticles. (E) Stable nanoparticles formulated with Cre mRNA and a unique DNA barcode were then pooled and intravenously administered to Ai14 mice at a total nucleic acid dose of 1.5 mg kg−1. Seventy-two hours later, tdTomato+ cells were identified and sorted using FACS and barcode representation quantified using NGS. (F) More nanoparticles containing R ionizable lipid enantiomers met our formulation criteria compared to the S conformation, but we found limited statistical differences in (G) LNP diameter and (H) polydispersity index. Statistical analyses were conducted using a two-way ANOVA between LNP diameters or PDIs of LNPs with different chiral ionizable lipids with Šidák’s multiple comparison test at every hydrocarbon tail length of the ionizable lipid. ns (p > 0.05, not shown), * (p < 0.05).
We then evaluated the LNPs in vivo, since in vitro nanoparticle delivery can poorly predict in vivo nanoparticle delivery20. Given that traditional one-by-one in vivo analysis would require several hundred mice, we used Fast Identification of Nanoparticle Delivery (FIND), a DNA barcoding system that evaluates how dozens of LNPs deliver mRNA in vivo with cellular resolution21–23 (Figure 1E). In this system, LNP-1, with chemical structure 1, is formulated to carry Cre mRNA and DNA barcode 1; this process is repeated N times so that LNP-N, with chemical structure N, is formulated to carry Cre mRNA and DNA barcode N. After the hydrodynamic diameter and stability of each LNP was characterized with dynamic light scattering (DLS), LNPs with monodisperse DLS spectra and a hydrodynamic diameter between 30 and 200 nm were pooled and administered to Ai14 mice; other LNPs were discarded. Since Ai14 mice are genetically engineered with a CAG-Lox-Stop-Lox-tdTomato construct, cells in which Cre mRNA is delivered and translated into functional Cre protein become tdTomato+. After isolating tdTomato+ cells using fluorescence-activated cell sorting (FACS), we used next-generation sequencing24 to identify the barcodes colocalized with cells in which functional mRNA delivery occurred.
Of the 128 LNPs we formulated, 55 passed our quality control criteria (Supplementary Figure 3A). Specifically, these LNPs had diameters between 30 nm and 200 nm as well as monodisperse DLS spectra. We noted that for C9, C10, and C12 lipids, more R stereoisomers met our quality inclusion criteria than S stereoisomers (Figure 1F). We then analyzed all the LNPs that met our inclusion criteria. We did not find statistically significant differences in hydrodynamic diameter (Figure 1G) and only found one statistically significant difference in polydispersity index (PDI) (Figure 1H). We then pooled the 55 LNPs and intravenously injected them into mice at a total nucleic acid dose of 1.5 mg kg−1 (i.e., 0.027 mg/kg/LNP, on average, for 55 LNPs). To minimize the effect of the barcodes on delivery, we used rationally designed, sensitive barcodes25, 26. To evaluate whether the LNPs were stable after pooling, we analyzed the hydrodynamic diameter and PDI of the pooled LNPs. Both the hydrodynamic diameter and PDI of the pool were within the range of the diameters and the PDI of the LNPs composing the pool (Supplementary Figure 3B–E), suggesting that the pooled LNPs did not aggregate. Seventy-two hours after LNP administration, which is a sufficiently long time for functional Cre mRNA delivery to generate tdTomato protein27, 28, we isolated the liver, lung, spleen, kidney, and bone marrow from the injected mice and digested them into single-cell suspensions. We then quantified the percentage of tdTomato+ cell types, a measure of functional Cre mRNA delivery, across 20 cell types in five organs (Figure 2A). Consistent with previous data demonstrating that C12-200 efficiently delivers RNA to hepatocytes, we observed the highest percentage of tdTomato+ cell types in the liver, including 87% of hepatocytes (Figure 2B). We observed less robust but statistically significant delivery in the splenic macrophages and endothelial cells (Figure 2C). The remaining organs had negligible tdTomato+ cells in all analyzed types (Supplementary Figure 4A–C). These data suggest that the pooled LNPs had tropism to hepatocytes and other liver cells.
Figure 2.
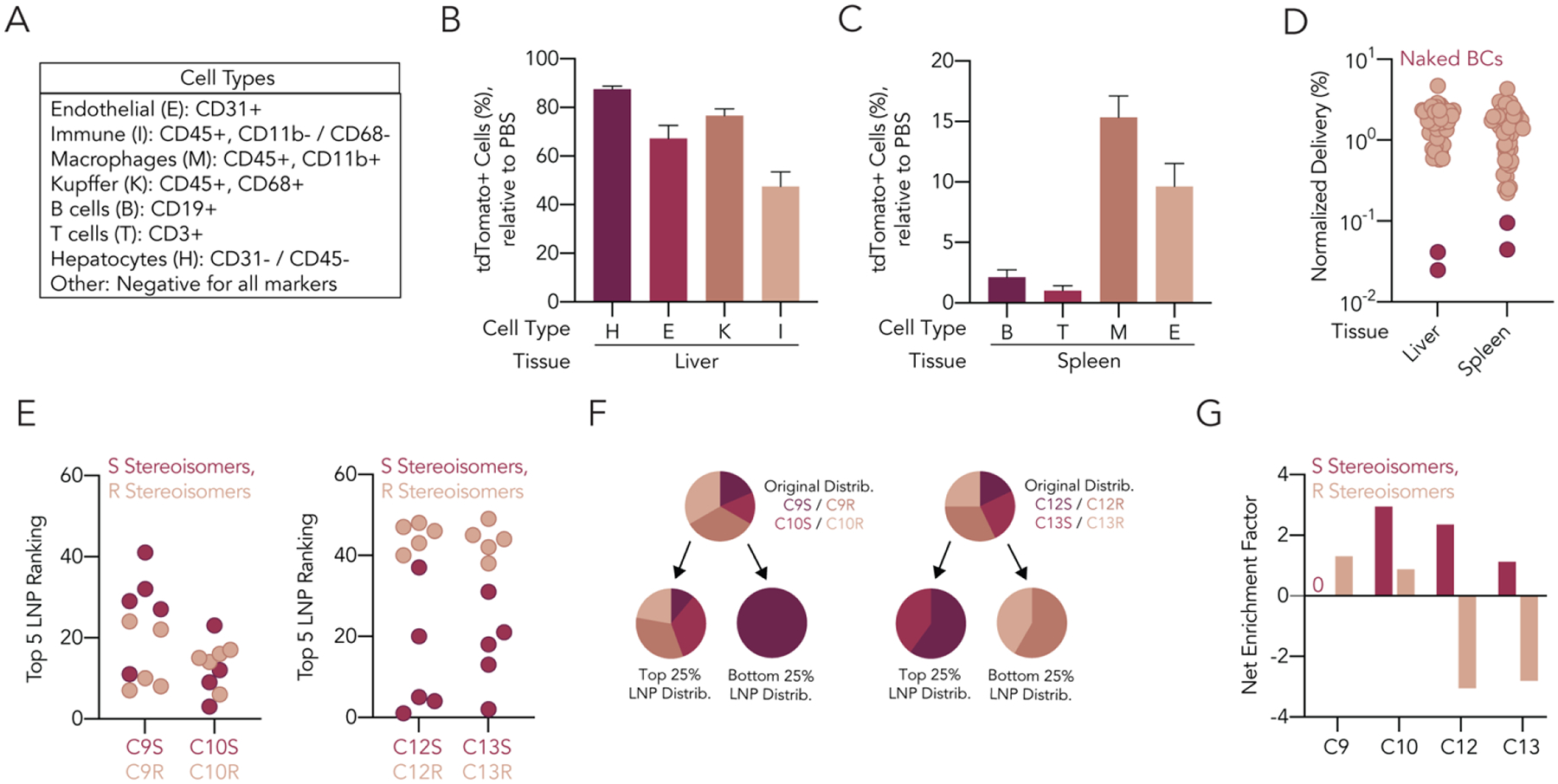
Quantifying functional mRNA delivery of 55 LNPs to multiple tissues in vivo. (A) We isolated eight different cell types from liver, lung, spleen, kidney, and/or bone marrow for analysis. Functional Cre mRNA delivery, quantified as tdTomato+ cells, was detected at significant levels for cell types in the (B) liver and (C) spleen. (D) Normalized delivery of all LNPs for the liver and the lung was calculated following DNA sequencing of barcodes within tdTomato+ cells in both organs. The naked barcodes fell out in both tissues, as expected. (E) We found no differences in the ranking of the top five LNPs for nanoparticles containing different stereoisomers of C9 and C10 ionizable lipids, but a complete separation in ranking for C12 and C13 ionizable lipids, where the S enantiomers outperformed the R. (F) Similarly, the original distribution of C9 and C10 ionizable lipids in the pool seemed equally represented in the top 25% of LNPs from the screen, but only S stereoisomers of C12 and C13 were represented in the top 25% of LNPs from the screen, with the R stereoisomers represented only in the bottom 25% of LNPs from the screen. (G) Fold net enrichment factor of all ionizable lipids, calculated by subtracting enrichment in the bottom 25% of LNPs from enrichment in the top 25% of LNPs.
We then sorted tdTomato+ hepatocytes, liver and spleen endothelial cells, liver Kupffer cells, and spleen macrophages; isolated DNA barcodes from each cell type; and sequenced the barcodes using NGS24. Next, we calculated the amount of delivery from each LNP by measuring the normalized delivery for each barcode; normalized delivery is similar to counts per million in RNA-seq28. We also analyzed two unencapsulated DNA barcodes, which act as negative sequencing controls. Since unencapsulated DNA does not readily enter cells, these barcodes should be delivered less efficiently than barcodes encapsulated in LNPs20; this was the case (Figure 2D).
The stability of the pooled LNPs, the poor performance of the naked barcodes, and the high percentages of tdTomato+ cells in the liver relative to other organs led us to conclude that our data were reliable enough to test the hypothesis that the stereochemistry of the ionizable lipid affects LNP delivery. We analyzed normalized delivery as a function of LNP stereochemistry and noted that stereopure C12S and C13S lipids ranked higher than the same LNPs containing C12R and C13R lipids (Figure 2E). Based on these data, we then analyzed whether R or S lipids were overrepresented in either the top 25% or bottom 25% of LNPs. For C12 and C13 lipids, only S stereoisomers were represented in the top 25% of LNPs and only R stereoisomers were represented in the bottom 25% of LNPs (Figure 2F,G, Supplementary Figure 4D)27, 29. In contrast, we did not find any meaningful differences between cholesterol and 20α-OH cholesterol (Supplementary Figure 4E). Notably, while stereochemistry did influence delivery, there was no relationship between delivery and LNP hydrodynamic diameter (Supplementary Figure 4F).
Like all high-throughput screens, FIND data are not conclusive; instead, they generate hypotheses that should be verified using individual LNPs. The screen data suggested that for lipids C12 and C13, the S stereoisomers performed significantly better than R stereoisomers (Figure 2E–G). We therefore formulated two LNPs: one LNP, termed LNP-42 S, contained the lipid C12S, while the other, termed LNP-60 S, contained C13S to confirm such hypothesis (Figure 3A and Supplementary Figure 3A). After formulating them to carry Cre mRNA, we verified that they formed stable nanoparticles meeting our formulation criteria from the screen (Figure 3B) and administered them to Ai14 mice at a dose of 1.0 mg kg−1. Importantly, both LNPs resulted in high percentages of tdTomato+ cells in the liver and spleen, recapitulating the results from the screen. In addition, LNP-42 S led to statistically significant increases in tdTomato+ cells, relative to LNP-60 S, in six of the eight cell types we measured (Figure 3C,D). We compared the LNPs in a dose response, injecting mice with 0.5, 0.25, or 0.1 mg kg−1. We again observed significantly more tdTomato+ cells in mice treated with LNP-42 S, relative to LNP-60 S, both in the liver (Figure 3E–H) and the spleen (Supplementary Figure 5A–D). We noted that delivery mediated by LNP-42 S was robust even at 0.1 mg kg−1: in hepatocytes we observed 34.1 ± 11.8% tdTomato+ cells, in endothelial cells we observed 25.8 ± 7.3% tdTomato+ cells, in Kupffer cells we observed 76.0 ± 5.2% tdTomato+ cells, and in other immune cells we observed 29.4 ± 11.4% tdTomato+ cells (Figure 3E–H), with 95% confidence intervals around the mean. To complement these flow cytometry data, we imaged livers isolated from mice treated with 0.1 mg kg−1 LNP-42 S or LNP-60 S and once again found higher levels of delivery with LNP-42 S (Figure 3I and Supplementary Figure 5E).
Figure 3.
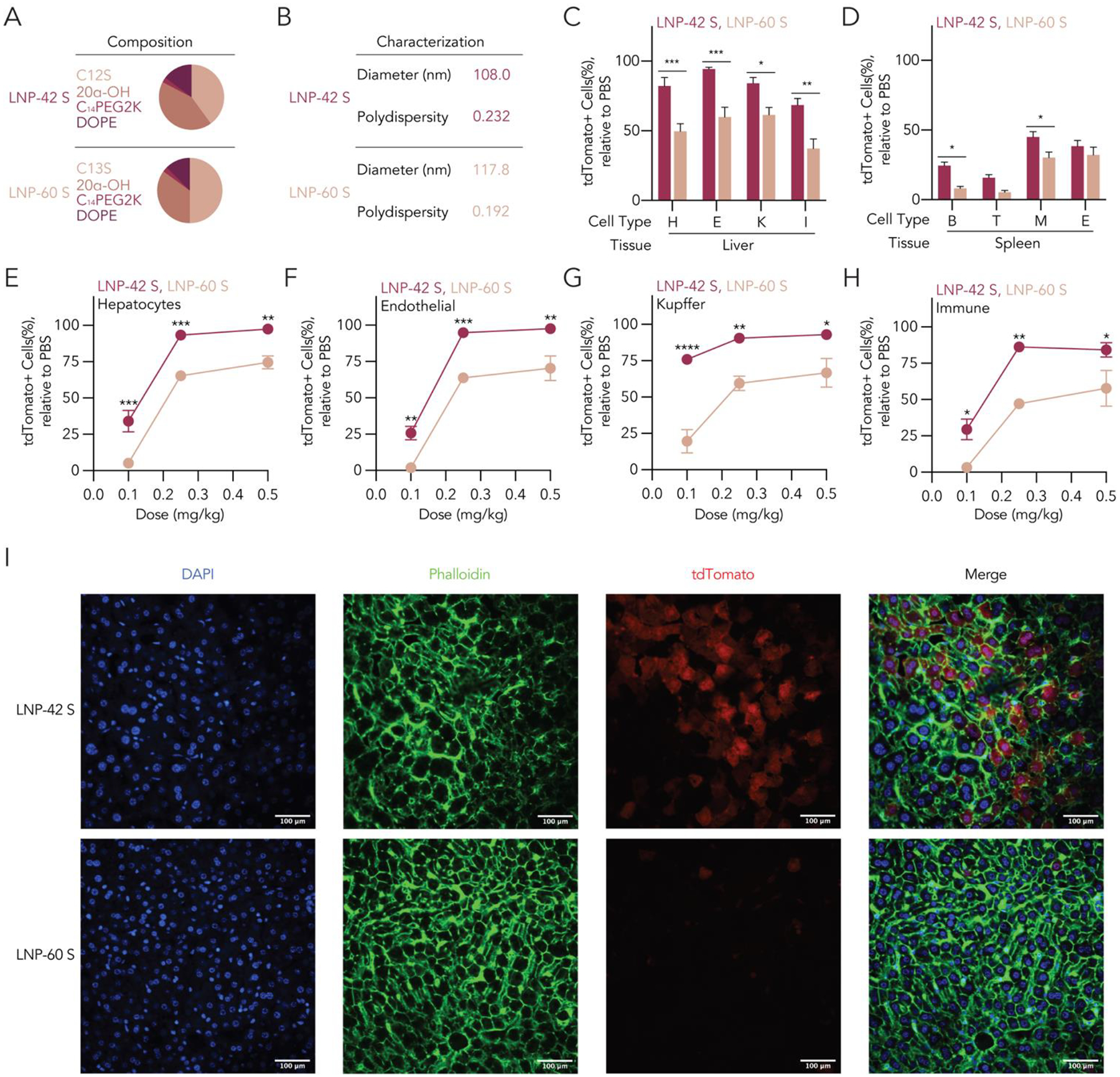
Individual LNPs recapitulate the potent delivery observed in the screen. (A) We formulated LNP-42 S and LNP-60 S, two nanoparticles containing the S enantiomer of C12 and C13, with Cre mRNA and (B) characterized their diameter and polydispersity index. We injected Ai14 mice with these LNPs at 1 mg kg−1 and observed strong functional delivery quantified by tdTomato+ cells relative to the control in the (C) liver and (D) spleen. We also observed statistically significant differences across the performance of both LNPs, where the percentage of tdTomato+ cells of LNP-42 S were higher than LNP-60 S. We then injected both LNPs at a dose of 0.5, 0.25, and 0.1 mg kg−1 and detected similar statistically significant differences in the percentage of tdTomato+ cells in liver (E) hepatocytes, (F) endothelial cells, (G) Kupffer cells, and (H) other immune cells. (I) Confocal microscopy of liver sections of LNP-42 S- and LNP-60 S-injected mice at a dose of 0.1 mg kg−1 showed clear differences in the levels of tdTomato+ cells present. Statistical analyses were conducted using a two-way ANOVA between tdTomato+ cells from LNP-42 S and LNP-60 S at different doses with Šidák’s multiple comparison test at every dose. ns (p > 0.05, not shown), * (p < 0.05), ** (p < 0.01), *** (p < 0.001), and **** (p < 0.001).
After confirming the activity of LNP-42 S, we used it to test the hypothesis, suggested by the DNA barcoding data, that stereochemistry affected mRNA delivery. We formulated S, racemic, and R versions of LNP-42 S, termed LNP-42 S, LNP-42 Racemic, and LNP-42 R, respectively, to carry Cre mRNA and intravenously injected them into Ai14 mice at dose of 0.05 mg kg−1. We chose this dose for two reasons: (i) we wanted our dose to be as clinically relevant (i.e., as low) as possible and (ii) we were concerned about saturating tdTomato+ cells in the Kupffer cell population (Figure 3C,G).We observed significantly higher percentages of tdTomato+ cells in hepatocytes, endothelial cells, and Kupffer cells in mice treated with LNP-42 S than with LNP-42 R (Figure 4A). Furthermore, for all cell types tested, LNP-42 Racemic achieved percentages of tdTomato+ cells between LNP-42 S and LNP-42 R. We repeated this experiment by substituting 20α-OH cholesterol with regular cholesterol and observed similar results (Supplementary Figure 6A). Finally, we wanted to confirm that these results could be replicated with a different payload. Therefore, we injected LNP-42 S, LNP-42 R, and LNP-42 Racemic encapsulating aVHH mRNA30, 31 to C57BL/6J mice at a dose of 0.1 mg kg−1. Similar to the results with Cre mRNA, we observed higher levels of aVHH protein in mice receiving LNP-42 S compared to LNP-42 Racemic and LNP-42 R (Supplementary Figure 6B).
Figure 4.
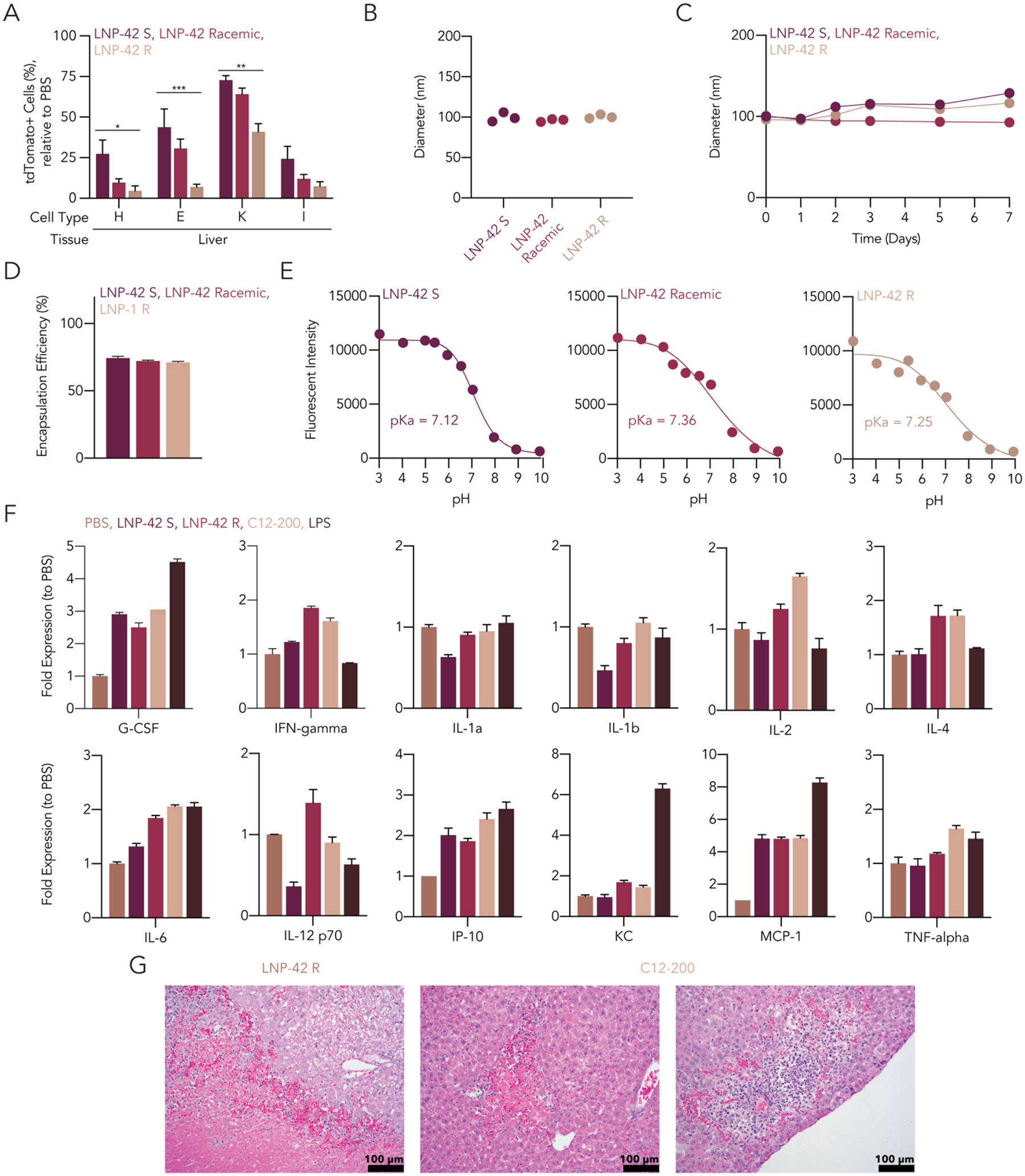
LNP-42 exhibits different functional delivery when formulated with ionizable lipids of different stereochemistry. (A) LNP-42 formulated with the S enantiomer, R enantiomer, or the racemic mixture of the ionizable lipid C12 achieved significantly different levels of tdTomato protein expression in the liver when administered intravenously to Ai14 mice at a dose of 0.05 mg kg−1. LNP-42 S, LNP-42 Racemic, and LNP-42 R showed (B) minimum changes in diameter after formulation and (C) stable diameters over a week of incubation at 4 °C. (D) There were no significant differences in the encapsulation efficiency of LNP-42 S, LNP-42 Racemic, and LNP-42 R containing Cre mRNA. (E) LNP-42 S, LNP-42 Racemic, and LNP42-R exhibited similar pKa. (F) Selection of cytokines and chemokines levels relative to PBS mice. LNP-42 S was shown to only have significantly higher levels of three cytokines when compared to PBS, whereas LNP-42 R, C12-200 Control, and LPS had seven, nine, and six, respectively. (G) Representative 20X images of liver sections stained for hematoxylin and eosin (H&E). Areas of hepatocellular necrosis were found in the LNP-42 R group and both hepatocellular necrosis (left) and inflammation (right) in the C12-200 group. Statistical analyses were conducted using a two-way ANOVA between tdTomato+ cells from LNP-42 containing different ionizable lipids with Tukey’s multiple comparison test at every cell type. ns (p > 0.05, not shown), * (p < 0.05), ** (p < 0.01), and *** (p < 0.001).
We reasoned that these stereochemistry-dependent differences in LNP-42 could be driven by overt changes in the LNP biophysical structure. However, when we measured the LNP diameter (Figure 4B), LNP stability over time (Figure 4C), encapsulation efficiency of the mRNA drug (Figure 4D), and pKa (Figure 4E), we found only slight differences between LNPs that did not explain the significant differences in LNP performance (Figure 4A and Supplementary Figure 6A,B). After excluding these variables, we hypothesized that increased delivery mediated by LNP-42 S could be caused by improved tolerability. This hypothesis is supported by evidence that even mild increases in inflammation can reduce LNP-mediated mRNA delivery32. Specifically, previous research has shown that cationic lipids can interact with pattern recognition receptors (PRR), including toll-like receptor 4 (TLR4), toll-like receptor 2 (TLR2), and NOD-like receptor family, pyrin domain containing 3 (NLRP3), mediate cytokine and chemokine release, and trigger an inflammatory response33–35. It follows that a reduced inflammatory response following administration of an LNP could achieve greater delivery efficiencies of the nanoparticle’s nucleic acid payload. To test this hypothesis, we formulated LNP-42 S and LNP-42 R with Cre mRNA and administered them intravenously to mice at a dose of 3 mg kg−1, a dose 60-fold higher than the dose needed for statistical mRNA delivery mediated by LNP-42 S. As a positive control, we also injected mice with racemic C12-200 formulated with a historic ratio of constituents (termed Control-C12-200) (Supplementary Figure 6C); this compound is an appropriate positive control given its reported toxicity at high RNA doses18. At this dose, all mice lost weight, compared to the PBS-treated controls (Supplementary Figure 6D). We then evaluated the high dose tolerability by quantifying serum cytokine levels, this time adding an LPS administered at a dose of 0.5 mg kg−1 as an additional positive control. Five hours after administration, we isolated blood from the tail vein in mice receiving PBS, the different LNPs, or LPS, and quantified serum cytokine levels (Figure 4F). We chose this timepoint based on the FDA approval document for Onpattro, a clinically approved LNP-siRNA formulation36 as well as data demonstrating that early cytokine levels are higher than 24-hour timepoints35, 37. Whereas only three cytokines were significantly upregulated in LNP-42 S mice (G-CSF, IP-10, and MCP-1), relative to PBS-treated mice, seven and nine cytokines were upregulated in LNP-42 R (G-CSF, IFN-gamma, IL-4, IL-6, IP-10, KC, and MCP-1) and C12-200-Control (G-CSF, IFN-gamma, IL-2, IL-4, IL-6, IP-10, KC, MCP-1, and TNF-alpha), respectively. Of particular concern were the significantly upregulated cytokines IFN-gamma (in the LNP1-R and C12-200 group), IL-6 (in the LNP1-R, C12-200, and LPS group), and TNF-alpha (in the C12-200 and LPS groups). IFN-gamma is upregulated after pathogen infections, activating macrophages and the production of MHC-II molecules38. IL-6 regulates immune and inflammatory responses and play an important role in the body’s response to injury39. TNF-alpha mediates acute and chronic systematic inflammatory reactions and has been implicated on the host’s defense against viral and bacterial infections40. We also found statistically significant differences in seven cytokine levels between the LNP-42 S group and the C12-200 control (Supplementary Figure 6E). It should be noted that the chemokine and cytokine assay measures relative levels of individual analytes; these levels are quantified by the pixel intensity of the analyte on the nitrocellulose membrane and compared to the pixel intensity of control groups, and do not generate the same order of magnitude changes as concentration-based serum cytokine assays. To complement these cytokine readouts, we had a blinded, independent group analyze histology using hematoxylin and eosin. Consistent with previous reports18, C12-200 Control led to multifocal areas of hepatocellular necrosis and mild focal areas of hepatocellular inflammation (Figure 4G and Supplementary Figure 6F,G); one caveat is that previous reports only observed necrosis from a single LNP administration at a dose of 7.5 mg kg−1. In addition, LNP-42 R led to severe focal areas of hepatocellular necrosis. However, LNP-42 S led to no observed changes in liver histology (Supplementary Figure 6G).
We then repeated the in vivo dose responses using LNP-60 S. However, we did not observe statistically significant differences between LNP-60 S, LNP-60 Racemic, and LNP-60 R (Supplementary Figure 7). As a result, we cannot exclude the possibility that stereochemistry-dependent differences in delivery are limited to a subset of LNPs.
Conclusions
We found that LNPs containing stereopure ionizable lipids may play a role in improving the therapeutic profile of mRNA therapies. Specifically, in the context of our experiments, we discovered that the S enantiomer of the ionizable lipid C12 both improves the effectiveness at low doses and tolerability at high doses of Cre mRNA compared to the same nanoparticle containing the R enantiomer or LNPs containing racemic mixtures of this family of ionizable lipids. Notably, there are several lines of evidence that suggest this effect may be lipid-dependent. For example, researchers found that different enantiomers of the lipid DLin-KC2-DMA and the lipid polymer 7C1 had the same activity when formulated with siRNA41, 42, where in another example, the R enantiomer of DOTAP showed better siRNA transfection, relative to its S enantiomer and racemic mixture43. We nevertheless believe that these results may inform researchers who seek to improve the therapeutic index of nucleic acid drugs. Moving forward, it may be important to determine whether the therapeutic profile of these nanoparticles changes when using stereopure lipids instead of their racemic mixtures.
It is also necessary to acknowledge the limitations of this study. First, even though we reported improvements both in the efficacy and tolerability of LNP-42 S, we did not determine the biological mechanism by which this happens. We find it feasible that this ionizable lipid may interact with extracellular or intracellular receptors, facilitating uptake and the release of the nanoparticles’ cargo and/or toning down an innate immune response, although future work will be needed to substantiate these hypotheses. Second, these studies were performed with an mRNA drug. It will be necessary to repeat these experiments with other nucleic acid cargos before extending these observations to other genetic therapies. Third, we only evaluated the effect of chirality in C12-200 analogs. Future studies should evaluate whether enantiomers of other ionizable lipids affect delivery or toxicity of LNPs. Nonetheless, we believe these data are important for two key reasons. First, hundreds of millions of LNPs have been administered to humans. Second, the FDA anticipates that by 2025, 10 to 20 gene therapies will be approved each year44; currently over 900 IND applications were filed for ongoing clinical trials in the field45. Given that some of these emerging drugs will likely use ionizable lipids, it will be important to understand how chemical structure influences LNP activity and tolerability.
Supplementary Material
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Credit Author Statement
Alejandro J. Da Silva Sanchez: Conceptualization, Methodology, Formal Analysis, Investigation, Writing (Original Draft)
Kun Zhao: Conceptualization, Investigation, Writing (Review & Editing)
Sebastian G. Huayamares: Investigation
Marine Z.C. Hatit: Investigation
Melissa P. Lokugamage: Investigation
David Loughrey: Investigation, Writing (Review & Editing)
Curtis Dobrowolski: Investigation
Shuaishuai Wang: Investigation
Hyejin Kim: Investigation
Kalina Paunovska: Writing (Review & Editing)
Yanina Kuzminich: Investigation
James E. Dahlman: Conceptualization, Writing (Review & Editing), Funding Acquisition
References
- 1.Adams D; Gonzalez-Duarte A; O’Riordan WD; Yang CC; Ueda M; Kristen AV; Tournev I; Schmidt HH; Coelho T; Berk JL; Lin KP; Vita G; Attarian S; Plante-Bordeneuve V; Mezei MM; Campistol JM; Buades J; Brannagan TH 3rd; Kim BJ; Oh J; Parman Y; Sekijima Y; Hawkins PN; Solomon SD; Polydefkis M; Dyck PJ; Gandhi PJ; Goyal S; Chen J; Strahs AL; Nochur SV; Sweetser MT; Garg PP; Vaishnaw AK; Gollob JA; Suhr OB, Patisiran, an RNAi Therapeutic, for Hereditary Transthyretin Amyloidosis. The New England journal of medicine 2018, 379 (1), 11–21. [DOI] [PubMed] [Google Scholar]
- 2.Gillmore JD; Gane E; Taubel J; Kao J; Fontana M; Maitland ML; Seitzer J; O’Connell D; Walsh KR; Wood K; Phillips J; Xu Y; Amaral A; Boyd AP; Cehelsky JE; McKee MD; Schiermeier A; Harari O; Murphy A; Kyratsous CA; Zambrowicz B; Soltys R; Gutstein DE; Leonard J; Sepp-Lorenzino L; Lebwohl D, CRISPR-Cas9 In Vivo Gene Editing for Transthyretin Amyloidosis. New England Journal of Medicine 2021. [DOI] [PubMed] [Google Scholar]
- 3.Baden LR; El Sahly HM; Essink B; Kotloff K; Frey S; Novak R; Diemert D; Spector SA; Rouphael N; Creech CB; McGettigan J; Khetan S; Segall N; Solis J; Brosz A; Fierro C; Schwartz H; Neuzil K; Corey L; Gilbert P; Janes H; Follmann D; Marovich M; Mascola J; Polakowski L; Ledgerwood J; Graham BS; Bennett H; Pajon R; Knightly C; Leav B; Deng W; Zhou H; Han S; Ivarsson M; Miller J; Zaks T, Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. New England Journal of Medicine 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Polack FP; Thomas SJ; Kitchin N; Absalon J; Gurtman A; Lockhart S; Perez JL; Pérez Marc G; Moreira ED; Zerbini C; Bailey R; Swanson KA; Roychoudhury S; Koury K; Li P; Kalina WV; Cooper D; Frenck RW; Hammitt LL; Türeci Ö; Nell H; Schaefer A; Ünal S; Tresnan DB; Mather S; Dormitzer PR; Şahin U; Jansen KU; Gruber WC, Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. New England Journal of Medicine 2020, 383 (27), 2603–2615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cullis PR; Hope MJ, Lipid Nanoparticle Systems for Enabling Gene Therapies. Mol Ther 2017, 25 (7), 1467–1475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Paunovska K; Loughrey D; Dahlman JE, Drug delivery systems for RNA therapeutics. Nature Reviews Genetics 2022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Semple SC; Akinc A; Chen J; Sandhu AP; Mui BL; Cho CK; Sah DW; Stebbing D; Crosley EJ; Yaworski E; Hafez IM; Dorkin JR; Qin J; Lam K; Rajeev KG; Wong KF; Jeffs LB; Nechev L; Eisenhardt ML; Jayaraman M; Kazem M; Maier MA; Srinivasulu M; Weinstein MJ; Chen Q; Alvarez R; Barros SA; De S; Klimuk SK; Borland T; Kosovrasti V; Cantley WL; Tam YK; Manoharan M; Ciufolini MA; Tracy MA; de Fougerolles A; MacLachlan I; Cullis PR; Madden TD; Hope MJ, Rational design of cationic lipids for siRNA delivery. Nat Biotechnol 2010, 28 (2), 172–6. [DOI] [PubMed] [Google Scholar]
- 8.Lokugamage MP; Sago CD; Dahlman JE, Testing thousands of nanoparticles in vivo using DNA barcodes. Current Opinion in Biomedical Engineering 2018, 7, 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Maier MA; Jayaraman M; Matsuda S; Liu J; Barros S; Querbes W; Tam YK; Ansell SM; Kumar V; Qin J; Zhang X; Wang Q; Panesar S; Hutabarat R; Carioto M; Hettinger J; Kandasamy P; Butler D; Rajeev KG; Pang B; Charisse K; Fitzgerald K; Mui BL; Du X; Cullis P; Madden TD; Hope MJ; Manoharan M; Akinc A, Biodegradable lipids enabling rapidly eliminated lipid nanoparticles for systemic delivery of RNAi therapeutics. Molecular therapy : the journal of the American Society of Gene Therapy 2013, 21 (8), 1570–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Khan OF; Kowalski PS; Doloff JC; Tsosie JK; Bakthavatchalu V; Winn CB; Haupt J; Jamiel M; Langer R; Anderson DG, Endothelial siRNA delivery in nonhuman primates using ionizable low-molecular weight polymeric nanoparticles. Science advances 2018, 4 (6), eaar8409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Love KT; Mahon KP; Levins CG; Whitehead KA; Querbes W; Dorkin JR; Qin J; Cantley W; Qin LL; Racie T; Frank-Kamenetsky M; Yip KN; Alvarez R; Sah DW; de Fougerolles A; Fitzgerald K; Koteliansky V; Akinc A; Langer R; Anderson DG, Lipid-like materials for low-dose, in vivo gene silencing. Proc Natl Acad Sci U S A 2010, 107 (5), 1864–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hutt AJ; O’Grady J, Drug chirality: a consideration of the significance of the stereochemistry of antimicrobial agents. J Antimicrob Chemother 1996, 37 (1), 7–32. [DOI] [PubMed] [Google Scholar]
- 13.Vargesson N, The teratogenic effects of thalidomide on limbs. J Hand Surg Eur Vol 2019, 44 (1), 88–95. [DOI] [PubMed] [Google Scholar]
- 14.Novobrantseva TI; Borodovsky A; Wong J; Klebanov B; Zafari M; Yucius K; Querbes W; Ge P; Ruda VM; Milstein S; Speciner L; Duncan R; Barros S; Basha G; Cullis P; Akinc A; Donahoe JS; Narayanannair Jayaprakash K; Jayaraman M; Bogorad RL; Love K; Whitehead K; Levins C; Manoharan M; Swirski FK; Weissleder R; Langer R; Anderson DG; de Fougerolles A; Nahrendorf M; Koteliansky V, Systemic RNAi-mediated Gene Silencing in Nonhuman Primate and Rodent Myeloid Cells. Molecular therapy. Nucleic acids 2012, 1, e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Miao L; Lin J; Huang Y; Li L; Delcassian D; Ge Y; Shi Y; Anderson DG, Synergistic lipid compositions for albumin receptor mediated delivery of mRNA to the liver. Nature communications 2020, 11 (1), 2424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Koga J; Nakano T; Dahlman JE; Figueiredo JL; Zhang H; Decano J; Khan OF; Niida T; Iwata H; Aster JC; Yagita H; Anderson DG; Ozaki CK; Aikawa M, Macrophage Notch Ligand Delta-Like 4 Promotes Vein Graft Lesion Development: Implications for the Treatment of Vein Graft Failure. Arteriosclerosis, thrombosis, and vascular biology 2015, 35 (11), 2343–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Leuschner F; Dutta P; Gorbatov R; Novobrantseva TI; Donahoe JS; Courties G; Lee KM; Kim JI; Markmann JF; Marinelli B; Panizzi P; Lee WW; Iwamoto Y; Milstein S; Epstein-Barash H; Cantley W; Wong J; Cortez-Retamozo V; Newton A; Love K; Libby P; Pittet MJ; Swirski FK; Koteliansky V; Langer R; Weissleder R; Anderson DG; Nahrendorf M, Therapeutic siRNA silencing in inflammatory monocytes in mice. Nat Biotechnol 2011, 29 (11), 1005–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Whitehead KA; Dorkin JR; Vegas AJ; Chang PH; Veiseh O; Matthews J; Fenton OS; Zhang Y; Olejnik KT; Yesilyurt V; Chen D; Barros S; Klebanov B; Novobrantseva T; Langer R; Anderson DG, Degradable lipid nanoparticles with predictable in vivo siRNA delivery activity. Nature communications 2014, 5, 4277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kauffman KJ; Dorkin JR; Yang JH; Heartlein MW; DeRosa F; Mir FF; Fenton OS; Anderson DG, Optimization of Lipid Nanoparticle Formulations for mRNA Delivery in Vivo with Fractional Factorial and Definitive Screening Designs. Nano Lett 2015, 15 (11), 7300–6. [DOI] [PubMed] [Google Scholar]
- 20.Paunovska K; Sago CD; Monaco CM; Hudson WH; Castro MG; Rudoltz TG; Kalathoor S; Vanover DA; Santangelo PJ; Ahmed R; Bryksin AV; Dahlman JE, A Direct Comparison of in Vitro and in Vivo Nucleic Acid Delivery Mediated by Hundreds of Nanoparticles Reveals a Weak Correlation. Nano letters 2018, 18 (3), 2148–2157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Paunovska K; Da Silva Sanchez AJ; Sago CD; Gan Z; Lokugamage MP; Islam FZ; Kalathoor S; Krupczak BR; Dahlman JE, Nanoparticles Containing Oxidized Cholesterol Deliver mRNA to the Liver Microenvironment at Clinically Relevant Doses. Advanced materials 2019, e1807748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lokugamage MP; Gan Z; Zurla C; Levin J; Islam FZ; Kalathoor S; Sato M; Sago CD; Santangelo PJ; Dahlman JE, Mild Innate Immune Activation Overrides Efficient Nanoparticle-Mediated RNA Delivery. Advanced materials 2019, e1904905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sago CD; Lokugamage MP; Paunovska K; Vanover DA; Monaco CM; Shah NN; Gamboa Castro M; Anderson SE; Rudoltz TG; Lando GN; Mummilal Tiwari P; Kirschman JL; Willett N; Jang YC; Santangelo PJ; Bryksin AV; Dahlman JE, High-throughput in vivo screen of functional mRNA delivery identifies nanoparticles for endothelial cell gene editing. Proceedings of the National Academy of Sciences 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Guimaraes PPG; Zhang R; Spektor R; Tan M; Chung A; Billingsley MM; El-Mayta R; Riley RS; Wang L; Wilson JM; Mitchell MJ, Ionizable lipid nanoparticles encapsulating barcoded mRNA for accelerated in vivo delivery screening. J Control Release 2019, 316, 404–417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sago CD; Kalathoor S; Fitzgerald JP; Lando GN; Djeddar N; Bryksin AV; Dahlman JE, Barcoding chemical modifications into nucleic acids improves drug stability in vivo. Journal of Materials Chemistry B 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sago CD; Lokugamage MP; Lando GN; Djeddar N; Shah NN; Syed C; Bryksin AV; Dahlman JE, Modifying a Commonly Expressed Endocytic Receptor Retargets Nanoparticles in Vivo. Nano letters 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Paunovska K; Da Silva Sanchez AJ; Sago CD; Gan Z; Lokugamage MP; Islam FZ; Kalathoor S; Krupczak BR; Dahlman JE, Nanoparticles Containing Oxidized Cholesterol Deliver mRNA to the Liver Microenvironment at Clinically Relevant Doses. Adv Mater 2019, 31 (14), e1807748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sago CD; Lokugamage MP; Paunovska K; Vanover DA; Monaco CM; Shah NN; Gamboa Castro M; Anderson SE; Rudoltz TG; Lando GN; Munnilal Tiwari P; Kirschman JL; Willett N; Jang YC; Santangelo PJ; Bryksin AV; Dahlman JE, High-throughput in vivo screen of functional mRNA delivery identifies nanoparticles for endothelial cell gene editing. Proc Natl Acad Sci U S A 2018, 115 (42), E9944–E9952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Paunovska K; Gil CJ; Lokugamage MP; Sago CD; Sato M; Lando GN; Gamboa Castro M; Bryksin AV; Dahlman JE, Analyzing 2000 in Vivo Drug Delivery Data Points Reveals Cholesterol Structure Impacts Nanoparticle Delivery. ACS Nano 2018, 12 (8), 8341–8349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hatit MZC; Lokugamage MP; Dobrowolski CN; Paunovska K; Ni H; Zhao K; Vanover D; Beyersdorf J; Peck HE; Loughrey D; Sato M; Cristian A; Santangelo PJ; Dahlman JE, Species-dependent in vivo mRNA delivery and cellular responses to nanoparticles. Nat Nanotechnol 2022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dobrowolski C; Paunovska K; Schrader Echeverri E; Loughrey D; Da Silva Sanchez AJ; Ni H; Hatit MZC; Lokugamage MP; Kuzminich Y; Peck HE; Santangelo PJ; Dahlman JE, Nanoparticle single-cell multiomic readouts reveal that cell heterogeneity influences lipid nanoparticle-mediated messenger RNA delivery. Nat Nanotechnol 2022, 17 (8), 871–879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lokugamage MP; Gan Z; Zurla C; Levin J; Islam FZ; Kalathoor S; Sato M; Sago CD; Santangelo PJ; Dahlman JE, Mild Innate Immune Activation Overrides Efficient Nanoparticle-Mediated RNA Delivery. Adv Mater 2020, 32 (1), e1904905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lonez C; Irvine KL; Pizzuto M; Schmidt BI; Gay NJ; Ruysschaert JM; Gangloff M; Bryant CE, Critical residues involved in Toll-like receptor 4 activation by cationic lipid nanocarriers are not located at the lipopolysaccharide-binding interface. Cell Mol Life Sci 2015, 72 (20), 3971–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lonez C; Bessodes M; Scherman D; Vandenbranden M; Escriou V; Ruysschaert JM, Cationic lipid nanocarriers activate Toll-like receptor 2 and NLRP3 inflammasome pathways. Nanomedicine 2014, 10 (4), 775–82. [DOI] [PubMed] [Google Scholar]
- 35.Abrams MT; Koser ML; Seitzer J; Williams SC; DiPietro MA; Wang W; Shaw AW; Mao X; Jadhav V; Davide JP; Burke PA; Sachs AB; Stirdivant SM; Sepp-Lorenzino L, Evaluation of efficacy, biodistribution, and inflammation for a potent siRNA nanoparticle: effect of dexamethasone co-treatment. Mol Ther 2010, 18 (1), 171–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.NDA 210922 – Patisiran – Cross-Discipline Team Leader Review; 2018.
- 37.Tao W; Mao X; Davide JP; Ng B; Cai M; Burke PA; Sachs AB; Sepp-Lorenzino L, Mechanistically probing lipid-siRNA nanoparticle-associated toxicities identifies Jak inhibitors effective in mitigating multifaceted toxic responses. Mol Ther 2011, 19 (3), 567–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Schroder K; Hertzog PJ; Ravasi T; Hume DA, Interferon-gamma: an overview of signals, mechanisms and functions. J Leukoc Biol 2004, 75 (2), 163–89. [DOI] [PubMed] [Google Scholar]
- 39.Lotz M, Interleukin-6: a comprehensive review. Cancer Treat Res 1995, 80, 209–33. [DOI] [PubMed] [Google Scholar]
- 40.Chu WM, Tumor necrosis factor. Cancer Lett 2013, 328 (2), 222–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jayaraman M; Ansell SM; Mui BL; Tam YK; Chen J; Du X; Butler D; Eltepu L; Matsuda S; Narayanannair JK; Rajeev KG; Hafez IM; Akinc A; Maier MA; Tracy MA; Cullis PR; Madden TD; Manoharan M; Hope MJ, Maximizing the potency of siRNA lipid nanoparticles for hepatic gene silencing in vivo. Angew Chem Int Ed Engl 2012, 51 (34), 8529–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Da Silva Sanchez AJ; Dobrowolski C; Cristian A; Echeverri ES; Zhao K; Hatit MZC; Loughrey D; Paunovska K; Dahlman JE, Universal Barcoding Predicts In Vivo ApoE-Independent Lipid Nanoparticle Delivery. Nano Letters 2022, 22 (12), 4822–4830. [DOI] [PubMed] [Google Scholar]
- 43.Terp MC; Bauer F; Sugimoto Y; Yu B; Brueggemeier RW; Lee LJ; Lee RJ, Differential efficacy of DOTAP enantiomers for siRNA delivery in vitro. Int J Pharm 2012, 430 (1–2), 328–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gottlieb S, Statement from FDA Commissioner Scott Gottlieb, M.D. and Peter Marks, M.D., Ph.D., Director of the Center for Biologics Evaluation and Research on new policies to advance development of safe and effective cell and gene therapies. 2019.
- 45.FDA, FDA Continues Strong Support of Innovation in Development of Gene Therapy Products. 2020.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.