

Abstract
Breast cancer (BC) is the second most common cancer and cause of death in women. In recent years many studies investigated the association of long non-coding RNAs (lncRNAs), as novel genetic factors, on BC risk, survival, clinical and pathological features. Recent studies also investigated the roles of metformin treatment as the firstline treatment for type 2 diabetes (T2D) played in lncRNAs expression/regulation or BC incidence, outcome, mortality and survival, separately. This comprehensive study aimed to review lncRNAs associated with BC features and identify metformin-regulated lncRNAs and their mechanisms of action on BC or other types of cancers. Finally, metformin affects BC by regulating five BC-associated lncRNAs including GAS5, HOTAIR, MALAT1, and H19, by several molecular mechanisms have been described in this review. In addition, metformin action on other types of cancers by regulating ten lncRNAs including AC006160.1, Loc100506691, lncRNA-AF085935, SNHG7, HULC, UCA1, H19, MALAT1, AFAP1-AS1, AC026904.1 is described.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12967-023-03909-x.
Introduction
Breast cancer (BC) is one of the most important cancers in women [1]. Different types of genetics and epigenetics factors are associated with BC initiation, development, aggressiveness, and treatment. The long non-coding RNA (lncRNA) is the latest significant genetic factor affecting BC. Many studies investigated the oncogenic or tumor suppressor effects of different lncRNAs on BC features, for example, H19 is an oncogenic lncRNA that dysregulates in BC and affects different BC features including proliferation, invasion, migration, cell cycle arrest, apoptosis, metastasis, tumor values, steroid receptor status, tumor size, nodal status, disease-free survival, prognosis, stemness, mesenchymal-to-epithelial transition (MET), epithelial-to-mesenchymal transition (EMT) (Table 1 and Additional file 1: Table S1). However, while there are several studies on the role of lncRNA in BC, they investigated some limited number of BC-associated lncRNAs and there is no comprehensive review on the roles of BC-associated lncRNAs. In addition, the role of metformin (1, 1-dimethyl biguanide hydrochloride), as an anti-hyperglycemic drug, on the risk of cancers was investigated in previous studies. The protective effect of metformin on cancers and its potential use in cancer treatment or in combination with chemotherapy and radiation therapy were investigated in previous studies [2–4]. It is associated with apoptosis, cell cycle, incidence and growth of tumors [5]. Several inhibitory mechanisms of metformin are associated with BC [6, 7]. Thus, this comprehensive review examines the relationship and effects of metformin, lncRNAs, and BC on each other as shown in Fig. 1. In the first section, prevalence, mortality, types, risk factors and molecular markers of BC will be described. In the second section, history, medical uses side effects, and antitumor mechanisms of metformin will be stated. In the third section, lncRNAs history, conservation, and functions will be discussed. In the fourth section, the effects of metformin on BC incidence, mortality, survival, and outcomes will be assessed. In the fifth section, the role of lncRNAs on BC will be assessed. In the final section the association of metformin, lncRNAs, and BC is investigated and discussed.
Table 1.
lncRNAs associated with BC
| LncRNA/ID (ensemble/NCBI) | Dysregulation of lncRNA (promote/inhibit) | Oncogene/tumor suppressor | Effect on BC | References |
|---|---|---|---|---|
| AC026904.1 (LINC02599)/ENSG00000233858 | Upregulation/promote | Oncogene | Metastasis, EMT | [111] |
| AFAP1-AS1/ENSG00000272620 | Upregulation/promote | Oncogene | Chemoresistance, proliferation, migration, invasion | [112] |
| ANCR/ENSG00000226950 | Upregulation/inhibit | Tumor suppressor | Invasion, metastasis, migration, EMT | [113] |
| DANCR/ENSG00000226950 | Upregulation/promote | Oncogene | Proliferation, invasion, tumor growth | [114] |
| ARNILA/ENSG00000235072 | Upregulation/promote | Oncogene | Progression-free survival, EMT, invasion, metastasis | [115] |
| ATB (LNCRNA-ATB)/Gene ID: 114004396 | Upregulation/promote | Oncogene | EMT, drug resistance, anti-apoptosis, proliferation, invasion, metastasis | [116] |
| BC200 (BCYRN1)/ENSG00000236824 | Upregulation/promote | Oncogene | Proliferation, migration, invasion, tumor size | [117] |
| BCAR4/ENSG00000262117 | Upregulation/promote | Oncogene | Migration, invasion, metastasis | [118] |
| BDNF-AS/ENSG00000245573 | Upregulation/promote | Oncogene | Prognostic predictor for poor survival, proliferation, endocrine resistance, progression | [119] |
| BLACAT1/ENSG00000281406 | Upregulation/promote | Oncogene | Migration, invasion, metastasis, cell survival, proliferation | [120] |
| BORG | Upregulation/promote | Oncogene | Invasion, metastasis, disease recurrence | [121] |
| CASC2/ENSG00000177640 | Upregulation/inhibit | Tumor suppressor | Proliferation, metastasis, cell cycle arrest, apoptosis, migration | [122] |
| CCAT1/Gene ID: 100507056 | Upregulation/promote | Oncogene | Cell proliferation, migration, invasion, tumor growth, progression | [123] |
| CCAT2/ENSG00000280997 | Upregulation/promote | Oncogene | Migration, invasion, metastasis, cancer growth, cell cycle, apoptosis, proliferation | [124] |
| EPIC1/ENSG00000224271 | Upregulation/promote | Oncogene | Cell cycle, progression, prognosis, tumor growth | [125] |
| ES1 (LINC01108)/ENSG00000226673 | Upregulation/promote | Oncogene | Stemness, proliferation, cell cycle progression, apoptosis, migration, EMT | [126] |
| EZR-AS1/ENSG00000233893 | Upregulation/promote | Oncogene | Tumor growth, metastasis, prognosis, proliferation, cell cycle progression, apoptosis, migration, invasion | [127] |
| FEZF1-AS1/ENSG00000230316 | Upregulation/promote | Oncogene | Stemness, tumorigenesis, proliferation, migration, invasion, cells growth | [128] |
| FGF13-AS1/ENSG00000226031 | Upregulation/inhibit | Tumor suppressor | Stemness, proliferation, migration, invasion, prognosis | [129] |
| GACAT3/ENSG00000236289 | Upregulation/promote | Oncogene | Prognosis, preoperative MRI perfusion-related diffusion reduction and elevated perfusion fraction | [130] |
| GAS5/ENSG00000234741 | Upregulation/inhibit | Tumor suppressor | Metastasis, proliferation, drug resistance | [131] |
| GHET1/ENSG00000281189 | Upregulation/promote | Oncogene | Proliferation, invasion, migration, apoptosis, EMT | [132] |
| H19/ENSG00000130600 | Upregulation/promote | Oncogene | Proliferation, invasion, migration, cell cycle arrest, apoptosis, metastasis | [133] |
| HIF1A-AS2/Gene ID: 100750247 | Upregulation/promote | Oncogene | Prognosis, migration, invasion, overall survival | [134] |
| HOST2 (CERNA2)/ENSG00000285972 | Upregulation/promote | Oncogene | Cell motility, migration, invasion | [135] |
| HOTAIR/ENSG00000228630 | Upregulation/promote | Oncogene | Proliferation, invasion, EMT, metastasis, drug resistance | [136] |
| ITGB2-AS1/ENSG00000227039 | Upregulation/promote | Oncogene | Migration, invasion | [137] |
| LET | Upregulation/inhibit | Tumor suppressor | Proliferation, invasion, migration, apoptosis | [138] |
| LIMT (LINC01089)/ENSG00000212694 | Downregulation/promote | Tumor suppressor | Migration, invasion, metastasis, prognosis | [139] |
| LINC00115/ENSG00000225880 | Upregulation/promote | Oncogene | Metastasis, migration, invasion | [140] |
| LINC00152 (CYTOR)/ENSG00000222041 | Upregulation/promote | Oncogene | Invasion, migration, tumorigenesis, colony growth, tumor growth, apoptosis | [141] |
| LINC00461/ENSG00000245526 | Upregulation/promote | Oncogene | Invasion, migration, differentiation | [142] |
| LINC00511/Gene ID: 400619 | Upregulation/promote | Oncogene | Proliferation, invasion, stemness, tumourigenesis | [143] |
| LINC00628/ENSG00000280924 | Upregulation/inhibit | Tumor suppressor | Proliferation, invasion, migration, cell growth, metastasis, apoptosis | [144] |
| LINC00673/Gene ID: 100499467 | Upregulation/promote | Oncogene | Proliferation, metastasis | [145] |
| LINC00899/ENSG00000231711 | Upregulation/inhibit | Tumor suppressor | Proliferation, migration, invasion, progression | [146] |
| LINC01133/ENSG00000224259 | Upregulation/inhibit | Tumor suppressor | Migration, invasion, metastasis | [147] |
| LINC01296 (DUXAP9)/ENSG00000225210 | Upregulation/promote | Oncogene | Proliferation, prognosis, metastasis, apoptosis | [148] |
| LINC01787/ENSG00000231987 | Upregulation/promote | Oncogene | Proliferation, migration | [149] |
| LINC01857/ENSG00000224137 | Upregulation/promote | Oncogene | Invasion, migration | [150] |
| LincIN | Downregulation/inhibit | Oncogene | Migration, invasion, metastasis | [151] |
| Linc-ITGB1 (IATPR)/ENSG00000233387 | Upregulation/promote | Oncogene | Invasion, migration, proliferation | [152] |
| Linc-ROR/ENSG00000258609 | Upregulation/promote | Oncogene | Tumorigenesis, migration, invasion, metastasis, growth | [153] |
| LINP1/ENSG00000223784 | Upregulation/promote | Oncogene | Apoptosis, mobility, EMT, drug resistance, migration, invasion, tumor growth | [154] |
| Lnc015192 | Upregulation/promote | Oncogene | Migration, invasion, EMT, metastasis | [155] |
| Lnc01638/ENSG00000233521 | Upregulation/promote | Oncogene | Proliferation, tumor growth, invasion, colonization, metastasis | [156] |
| Lnc-BM | Upregulation/promote | Oncogene | Metastasis | [157] |
| LncKLHDC7B/ENSG00000226738 | Downregulation/promote | Tumor suppressor | Invasion, migration, apoptosis | [158] |
| LncRNA 91H | Upregulation/promote | Oncogene | Cell growth, migration, invasion, tumor growth | [159] |
| lncRNA HIT | Upregulation/promote | Oncogene | Migration, invasion, tumor growth, metastasis, EMT | [160] |
| LncRNA RP1/ENST00000420172 | Upregulation/promote | Oncogene | Prognosis, proliferation, metastasis, EMT, stemness | [161] |
| LncRNA-ATB/Gene ID: 114004396 | Upregulation/promote | Oncogene | Drug resistance, invasion-metastasis cascade | [162] |
| lncRNA-Hh (GAS1RR)/ENSG00000226237 | Upregulation/promote | Oncogene | Tumorigenesis | [163] |
| Lnc-SLC4A1-1 | Downregulation/inhibit | Oncogene | Apoptosis, proliferation, migration, invasion | [164] |
| MAGI2-AS3/ENSG00000234456 | Upregulation/inhibit | Tumor suppressor | Cell growth, proliferation, cell viability, colony formation, apoptosis | [165] |
| MALAT1/ENSG00000251562 | Upregulation/inhibit | Tumor suppressor | Migration, invasion | [166] |
| Upregulation/promote | Oncogene | |||
| Migration, progression, proliferation, differentiation, metastasis | [167] | |||
| MAYA (MNX1-AS1)/ENSG00000243479 | Upregulation/promote | Oncogene | EMT, proliferation, migration, invasion | [168] |
| MEG3/ENSG00000214548 | Upregulation/inhibit | Tumor suppressor | Proliferation, Migration, Invasion, Apoptosis | [169] |
| MIR100HG/ENSG00000255248 | Upregulation/promote | Oncogene | Proliferation, cell arrest in the G1 | [170] |
| MT1JP/ENSG00000255986 | Upregulation/inhibit | Tumor suppressor | Proliferation, invasion, drug sensitivity | [171] |
| NBAT1/ENSG00000260455 | Upregulation/inhibit | Tumor suppressor | Migration, invasion, metastasis, prognosis | [172] |
| NEAT1/ENSG00000245532 | Upregulation/promote | Oncogene | Tumor size, prognosis, proliferation, metastasis, EMT | [173] |
| LncRNA NEF (LNCNEF)/ENSG00000237396 | Upregulation/inhibit | Tumor suppressor | Invasion, migration | [174] |
| NKILA/ENSG00000278709 | Upregulation/inhibit | Tumor suppressor | EMT, metastasis | [175] |
| NLIPMT/ENSG00000278709 | Upregulation/inhibit | Tumor suppressor | Proliferation, motility, progression | [176] |
| NNT-AS1/ENSG00000248092 | Upregulation/promote | Oncogene | Migration, invasion, progression, EMT, proliferation | [177] |
| NORAD/ENSG00000260032 | Downregulation/promote | Tumor suppressor | Migration, invasion, metastasis | [178] |
| P10247 (lncRNA-BCHE)/ENSG00000114200 | Upregulation/promote | Oncogene | Metastasis, growth, migration, invasion | [179] |
| PDCD4-AS1/ENSG00000203497 | Upregulation/inhibit | Tumor suppressor | Progression | [180] |
| PRLB (SIRLNT)/ENSG00000253802 | Upregulation/promote | Oncogene | Proliferation, chemoresistance, metastasis, survival, invasion | [181] |
| PTENP1/ENSG00000237984 | Upregulation/inhibit | Tumor suppressor | Proliferation, invasion, tumorigenesis, tumor growth, metastasis, apoptosis, chemoresistance | [182] |
| PVT1/ENSG00000249859 | Upregulation/promote | Oncogene | Proliferation, colony formation, tumor growth | [183] |
| PITPNA-AS1/ENSG00000236618 | Downregulation/inhibit | Oncogene | Cell viability, proliferation, migration, invasion | [184] |
| SPRY4-IT1/GeneID:100642175 | Upregulation/promote | Oncogene | Proliferation, migration, invasion, cell cycle, apoptosis | [185] |
| SUMO1P3/ENSG00000235082 | Downregulation/inhibit | Oncogene | Progression, survival, proliferation, migration, invasion | [186] |
| TFAP2A-AS1/ENSG00000229950 | Upregulation/inhibit | Tumor suppressor | Invasion, migration, proliferation, cell cycle arrest, apoptosis, ability, tumor growth | [187] |
| TINCR/ENSG00000223573 | Upregulation/promote | Oncogene | Migration, invasion, tumor growth, proliferation, apoptosis | [188] |
| TUG1/ENSG00000253352 | Downregulation/promote | Tumor suppressor | Apoptosis, proliferation, metastasis, invasion | [189] |
| Downregulation/inhibit | Oncogene | Proliferation, metastasis, tumor size, TNM staging, migration, invasion, apoptosis | [190] | |
| TUNAR/ENSG00000250366 | Upregulation/promote | Oncogene | Stemness, motility, invasion, EMT | [191] |
| UCA1/ENSG00000214049 | Upregulation/promote | Oncogene | Apoptosis, drug resistance | [192] |
| XIST/ENSG00000229807 | Downregulation/promote | Tumor suppressor | Cell growth, migration, invasion | [193] |
| LINC02095(ROCR)/ENSG00000228639 | Upregulation/promote | Oncogene | Proliferation | [194] |
| WT1-AS/ENSG00000183242 | Upregulation/inhibit | Tumor suppressor | Clinical stages, migration, invasion | [195] |
| LINC00096(TP53TG1)/ENSG00000182165 | Upregulation/promote | Oncogene | Proliferation, invasion, metastasis | [196] |
| HEIH/ENSG00000278970 | Downregulation/inhibit | Oncogene | Proliferation, apoptosis | [197] |
| LUCAT1/ENSG00000248323 | Upregulation/promote | Oncogene | Proliferation, cell cycle progression, metastasis, apoptosis | [198] |
| ASRPS(LINC00908)/ENSG00000266256 | Downregulation/promote | Tumor suppressor | Angiogenesis, tumor growth | [199] |
| HAND2-AS1/ENSG00000237125 | Upregulation/inhibit | Tumor suppressor | Proliferation | [200] |
| LINC01096/ENSG00000246095 | Downregulation/inhibit | Oncogene | Proliferation, migration, invasion; apoptosis, cell viability | [201] |
| PANDA(PANDAR)/ENSG00000281450 | Upregulation/promote | Oncogene | Apoptosis | [202] |
| TP73-AS1/ENSG00000227372 | Upregulation/promote | Oncogene | Proliferation, invasion, migration | [203] |
| CRNDE/ENSG00000245694 | Upregulation/promote | Oncogene | Proliferation, migration, invasion | [204] |
| HCP5/ENSG00000206337 | Downregulation/promote | Tumor suppressor | Drug resistance | [205] |
| ADAMTS9-AS2/ENSG00000241684 | Downregulation/promote | Tumor suppressor | Drug resistance, apoptosis, viability | [206] |
| TMPO-AS1/ENSG00000257167 | Upregulation/promote | Oncogene | Proliferation, viability, apoptosis, drug resistance | [207] |
| DSCAM-AS1/ENSG00000235123 | Downregulation/inhibit | Oncogene | Proliferation, colony formation | [208] |
| MAFG-AS1(MILIP)/ENSG00000265688 | Downregulation/inhibit | Oncogene | Proliferation, apoptosis, drug resistance | [209] |
| DILA1(MIR99AHG)/ENSG00000215386 | Upregulation/promote | Oncogene | Drug resistance, proliferation, prognosis, tumor growth | [210] |
| DLX6-AS1/ENSG00000231764 | Upregulation/promote | Oncogene | Apoptosis, migration, drug resistance, EMT | [211] |
| SNHG7/ENSG00000233016 | Upregulation/promote | Oncogene | Viability, drug resistance | [212] |
| DCST1-AS1/ENSG00000232093 | Upregulation/promote | Oncogene | Drug resistance, EMT, chemoresistance | [213] |
| LINC00472/ENSG00000233237 | Downregulation/promote | Tumor suppressor | Growth, aggressiveness | [214] |
| AGAP2-AS1/ENSG00000255737 | Upregulation/promote | Oncogene | Tumor growth, apoptosis, chemoresistance, drug resistance | [215] |
| SNHG14/ENSG00000224078 | Upregulation/promote | Oncogene | Proliferation, invasion, drug resistance | [216] |
| MAPT-AS1/ENSG00000264589 | Downregulation/inhibit | Oncogene | Drug resistance, migration, invasion, proliferation | [217] |
| Linc00518/ENSG00000183674 | Upregulation/promote | Oncogene | Drug resistance | [218] |
| FTH1P3/ENSG00000213453 | Downregulation/inhibit | Oncogene | Drug resistance | [219] |
| FGF14-AS2/ENSG00000272143 | Downregulation/promote | Tumor suppressor | Progression, prognosis, tumor size, lymph node metastasis, clinical stage, overall survival | [220] |
| PAPAS | Upregulation/promote | Oncogene | Migration, invasion | [221] |
| lncMat2B | Upregulation/promote | Oncogene | Drug resistance | [222] |
| LOL | Downregulation/inhibit | Oncogene | Apoptosis, proliferation, drug resistance | [223] |
| BC032585 | Downregulation/promote | Tumor suppressor | Drug resistance | [224] |
| NONHSAT101069 | Upregulation/promote | Oncogene | Drug sensitivity, metastasis, migration, invasion | [225] |
| DRHC | Downregulation/promote | Tumor suppressor | Proliferation | [226] |
Fig. 1.

Study process
BC
The first evidence of BC dates back to 2500–3000 BC. There is also evidence of BC in Hippocrates' manuscripts. It is the second most common type of cancer in women in the United States [1]. The lifetime risk of developing BC is one out of every eight women [8]. About 264,000 new cases of BC in women are diagnosed in the United States every year [1]. After lung cancer, it is the second cause of death from cancers in women. The number of deaths from this cancer in women is about 42,000 each year [1], and 15% of women deaths between the age of 20 to 59 years occur due to BC [9]. Male BC is not common and makes up only one percent of all patients with BC. Although the incidence of BC is lower in men, the prognosis is worse [10].
Based on the cell origin, BC is carcinoma or sarcoma. Carcinomas arise from the epithelial components including the cells that line the lobules and ducts responsible for milk production. Sarcomas, a rarer form, arise from the stromal components including myofibroblasts and blood vessel cells [11]. Based on the criteria of pathological characteristics and aggressiveness, BC is graded as non-invasive/preinvasive (intraductal carcinoma, most common type), invasive, and metastatic. There are two ductal and lobular types of BC. The ductal form involves milk ducts. But in the lobular form, the milk-producing glands are involved. These types are also divided into invasive (or infiltrating) and in-situ forms based on their spread pattern. Unlike the in-situ form, the invasive form invades the surrounding tissues [12]. The invasive ductal carcinoma (80% of BC) includes tubular, medullary, mucinous, papillary, and cribriform carcinoma subtypes, while invasive lobular carcinoma (10–15% BC) is reported more in women with increasing age [11]. Also, there are two special invasive types of BC including triple-negative and inflammatory. In triple-negative BC (TNBC), cancerous cells do not have estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor (EGFR) which leads to poor survival outcomes [13]. TNBC common treatments are lumpectomy, mastectomy, radiation therapy, and chemotherapy. The human epidermal growth factor receptor 2 (HER2) is not present in these cells [10]. In the inflammatory form, lymphatic vessels are also blocked, which is a very aggressive form [14].
The breast completes its development in puberty and pregnancy through alterations in breast differentiation and proliferation with more ductal branching and lobuloalveolar growth. The balance between differentiation, proliferation, and apoptosis is critical for the normal development and homeostasis of breast cells. The premalignant stages change this balance and the regulatory biomarkers of the cell cycle. Change in this may lead to the accumulation of mutations in BC [15]. Uncontrolled cell proliferation in tumors is associated with lower apoptosis. Apoptosis is a marker of prognosis, the low level of apoptosis is a poor prognostic feature while high levels of apoptosis in tumors have been correlated with the absence of estrogen receptors and worse survival [15]. The apoptosis-associated proteins are involved in BC and its treatment. The Bad, Bak, Bax, and Bcl-xs are pro-apoptotic proteins and Bcl-2 and Bcl-xL are anti-apoptotic proteins. Tumor Protein 53 (P53) is associated with a poor prognosis of BC and poor response to chemotherapy. The XIAP, NIAP, cIAP1, cIAP2, and survivin proteins prevent apoptosis by directly inhibiting caspases [15].
Metastasis is associated with most cases of cancer mortality which genetically leads migration of cancerous cells to other organs by the blood and lymphatic vessels [9, 16]. It is associated with processes including MET, EMT, cancer cell migration, invasion, proliferation, stemness, angiogenesis, anoikis [16], endothelial cells, macrophages, extracellular matrix, stem cells [17] and cancer-associated fibroblasts [18]. The microvascular production increase in tumor cells and is associated with poor prognosis of BC. The hypoxia and genetic changes in the tumor cells are related to an increase in angiogenic factors [19]. The hypoxic tumor HIF-1α induces the expression of proangiogenic mediators [20]. Different factors such as Hypoxia, HIF-1α, HER2, and matrix metalloproteinase 9 (MMP-9) increase the expression of VEGF in BC, and VEGF is associated with angiogenesis in breast tumors. VEGF is involved in the initial BC tumor growth while bFGF is increased during further tumor growth. The FGF/FGFR signaling as an angiogenic driver is associated with BC therapeutic perspectives [21]. In general, different tissues are different in the angiogenic response.
Several risk factors have been identified for BC. However, more than 50% of affected people had no risk factors except for increasing age and female gender [10]. In general, these risk factors can be classified into two categories: changeable and non-changeable. The most important non-changeable risk factors are genetics, increasing age, female gender, race, taller height, first-degree family history of BC, benign BC, early menarche less than 12 years old, or late menopause older than 55 years old. The most important changeable risk factors include risk factors related to lifestyle and personal behavior, long-term hormone replacement therapy, chest wall irradiation, obesity, high-fat diet, and environmental chemicals. Although a variety of risk factors have been discovered, the exact causes of breast cancer are not yet entirely understood.
The “tumor biomarker” is a possible indicator of the onset, development, and progression of cancer. We must examine these biomarkers deeper and work to understand the underlying processes of tumor formation to improve treatment and personalized medicine [22]. The predictive and prognostic biomarkers, such as circulating tumor cells (CTCs), DNA, RNA, and miRNA, have been connected to probable clinical outcomes and treatment effectiveness of BC. Intact cells and larger molecules such as nucleic acids, genetic changes, and protein molecules are used as molecular, histological, radiographical, or physiological biomarkers in the detection of cancer [23]. Investigating molecular markers leads to the improvement of BC screening, diagnosis, and treatment processes. BC biomarkers are classified into two tumors or blood categories. Different regulatory noncoding RNAs such as miRNAs, lncRNAs, and piRNAs play non-invasive biomarkers roles in BC development, diagnosis, and prognosis. Inappropriate circulating mRNAs expression is associated with BC as a tumor marker, for example 5T4 circulating RNA may potentially be used to identify patients who can benefit from a 5T4 therapy utility of circulating RNA [24], or circulating circular RNA hsa_circ_0001785 upregulation in BC which could be a biomarker for BC diagnosis and progress [25]. The circulating cell-free miRNAs in the serum of BC patients emerged as a promising new noninvasive biomarker for the early detection of tumors and for predicting their molecular classifications. These miRNAs target mRNA that encodes proteins involved in different molecular pathways such as, proliferation, cell adhesion, and migration [26]. miRNA:mRNA interactions are associated with the invasiveness of BC [27]. miRNAs are suggested to be associated with BC therapeutic approaches [28], prognosis [29], progression, and metastasis [30]. P-element-induced wimpy testis (PIWI) interacting RNAs (piRNAs) are a novel type of non-coding RNAs, with act on both transcription and post-transcription. Upregulation of piR-021285, piRNA-823, piRNA-932, piR-016658, piR-651, piR-4987, piR-20365, piR-20485, and piR-20582 were associated with EMT, invasiveness, metastasis, lymph node metastasis, while downregulation of piR-36712, piR-016975, piR-FTH1 in BC tissue were related to EMT, chemosensitivity, and chemoresistance [31]. PiRNA-mediated epigenetic mechanism and altered DNA methylation are involved in BC tumorigenesis [32]. The function and mechanism of many piRNAs in cancers is unclear. The role of lncRNAs as one of the most important noncoding RNAs in BC will be fully discussed in “lncRNAs in BC” section.
Antitumor mechanisms of metformin in BC
Metformin is a synthetic derivative of galegine and/or guanidine and belongs to a group of compounds called biguanides. galegine synthesize from Gallega officinalis, a plant that has been used for centuries in Europe to treat diabetes [33]. Metformin works by improving insulin sensitivity in patients with type 2 diabetes (T2D) due to its biguanide properties [34]. For the first time, metformin was used to treat T2D in the 1950s, but from 1995 it became widely used in the United States as a first-line treatment [35]. It is an anti-hyperglycemic drug (lowering blood sugar level) and the first line of treatment for T2D. The reduction of blood sugar level is done by inhibiting hepatic glucose production, increasing glucose absorption, and its consumption by skeletal muscles [36]. In addition, Metformin reduces insulin resistance in the surrounding tissue and suppresses gluconeogenesis in the liver.
Metformin has pleiotropic effects and is beneficial in the treatment of diabetes in various diseases, including prediabetes, and type 1 diabetes mellitus [37]. Many previous studies, demonstrate the safety and well-tolerance of metformin, along with potential nephroprotective and cardioprotective effects [38]. There are several other indications for metformin that are not FDA-approved, including the treatment of gestational diabetes, the treatment of weight gain caused by antipsychotics, the prevention of type 2 diabetes, as well as the treatment and prevention of polycystic ovary syndrome [39]. However, in some specific populations, such as patients with renal or hepatic impairment, pregnant or breastfeeding women, and pediatric or geriatric patients, metformin usage may not be commonly safe. The primary potential of lactic acidosis production made it unsafe in some circumstances [40]. It is important to understand the benefit-risk balance of metformin treatment when it comes to elderly patients who are highly likely to have stable renal impairment, congestive heart failure, and/or coronary artery disease due to the high prevalence of these conditions.
Although no serious complications of metformin have been observed, this drug has some side effects like any other drug. There may be symptoms such as dizziness, severe drowsiness, muscle pain, tiredness, chills, blue/cold skin, rapid or difficult breathing, slow heartbeat, stomach pain associated with diarrhea, and a feeling of nausea and vomiting. In most cases, lactic acidosis is caused by an overdose of medication or is the result of some contraindications. Metformin rarely causes hypoglycemia, however, if it is used in conjunction with other anti-diabetic medications, low blood sugar may develop. In rare cases, it leads to a serious allergic reaction. The product contains inactive components that may cause reactions or other problems when administered to patients.
The risk of developing cancer in diabetic patients taking metformin is reduced compared to other people, and its protective effect increases with exposure to a higher dose of metformin [2]. Metformin reduces cell proliferation, induces apoptosis, stops the cell cycle in vitro, and also reduces the incidence and growth of tumors in vivo [3]. It can be used as a sensitizing agent or in combination with chemotherapy and radiation therapy to fight cancer [4]. There are several mechanisms by which metformin inhibits the proliferation of malignant cells, such as activation of AMP-activated protein kinase (AMPK) or Mitogen-activated protein kinases (MAPK), decrease in mechanistic target of rapamycin (mTOR) signaling, increase in p27 expression, protein synthesis, EGFR, Src, and expression of Cyclins. Controlling the expression levels of proteins that are necessary for the transition between the G1 and S cell cycle, including cyclin D1, cyclin E1, and E2F transcription factor 1 [6, 7]. Metformin inhibits Cyclin-dependent kinases and causes the cell cycle to stop in the G1 phase [41]. The ability to deactivate biosynthetic mitochondrial nodes in cells with the BRCA1 gene is a potential mechanism of action of metformin for suppressing the BC formation in cell types affected by BRCA1 [42]. It was also proven that BRCA1 haploinsufficiency leads to the activation of AKT/mTOR-mediated protein synthesis driven by hyperphosphorylation of the BRCA1 substrate rich in proline through AKT activity. Another interesting function of metformin is the disruption of AKT/mTOR-signaling network in BRCA1 haploinsufficient cells [43]. Decreasing insulin and insulin-like growth factor levels inhibit signaling that involve phosphoinositide 3-kinase, AKT, and mTOR, it may be related to the inhibition of mTOR signaling by activating AMPK [39]. A major target of metformin therapy is the regulation of AMPK through an AMP-dependent signaling pathway. AMPK activation inhibits cell mitosis and proliferation [44]. Metformin exerts its anti-neoplastic effects by stimulating AMPK through up-regulation of p53–p21 and down-regulation of Cyclin D1 levels. The activation of AMPK, through inhibition of the mTOR, activity, fatty acid synthesis signaling pathways, as well as stimulating the apoptotic pathway (p53/p21) is responsible for regulating tumor cell survival and tumor growth [5]. The neoadjuvant metformin administration in BC decreased insulin receptors, phosphorylation of protein kinase B (PKB)/Akt, AMPK, and extracellular signal-regulated kinase1/2. This insulin-dependent effect of metformin is consistent with its anticancer properties [45]. Metformin has numerous beneficial properties in both normal and cancerous cells, including reduced insulin levels, inhibition of insulin/IGF signaling pathways, as well as modulation of cellular metabolism [46]. There are several roles played by insulin/IGF-1 in the regulation of glucose uptake, as well as the regulation of carcinogenesis through the upregulation of signaling pathways associated with insulin/IGF receptors [47]. The signal through the insulin receptor substrate phosphorylates (but does not activate) mTORC1. Furthermore, through growth factor receptor-bound protein 2, insulin signals are transmitted to Ras/Raf/ERK pathway which is responsible for regulating cell growth [48]. Several studies have indicated that these pathways play a significant role in the changes that occur in the metabolism of cancer cells [49]. A significant decrease in IGF-1/insulin receptor activity, Akt, extracellular signal-regulated kinase (ERK) activity, and AMPK activity is observed with metformin use without activation of AMPK by this drug [46]. It has been shown that metformin inhibits mTOR signaling in Drosophila cells when AMPK is absent. There is an alternative pathway mediated by the RAG GTPase associated with tuberous sclerosis protein (TSC1/2-mTOR) which is AMPK independent [50]. On the other hand, as a result of decreasing the levels of HER2 in breast cancer cells, metformin can inhibit breast carcinoma cell growth, and inhibition of p70S6K1, an effector of the mTOR pathway, can mediate this effect [51].
Hyperglycemia induces oxidative stress both directly and indirectly in BC cells in part by increasing levels of insulin/IGF-1 as well as inflammatory cytokines [52]. Furthermore, the activation of nuclear factor kappa (NFκB), signal transducer activator of transcription 3 (STAT3), and the hypoxia-inducible factor 1-alpha (HIF1α) are involved [53]. Through anti-inflammatory effects in cell models, metformin inhibits the components of the NFκB pathway that are essential for the transformation of stem cells and the formation of cancer stem cells. Metformin also prevents phosphorylation of STAT3 in cancer stem cells [54, 55]. The suppression of chronic inflammatory response by metformin is related to the inhibition of TNF-α production in human monocytes and chronic inflammation provides a basis for cancer development [56]. Metformin decreases cellular-Myc (c-Myc) and increases double-stranded RNA specific endoribonuclease (DICER) in AMPK signaling [57]. Metformin leads to reprograming of lipid metabolism, as a hallmark of cancer, by increase in acetyl-CoA carboxylase (ACC) and fatty acid synthase (FASN), and miRNA regulation [58]. It also inhibits Complex I of the mitochondria and increases the AMP/ATP ratio which leads to further AMPK activation [59]. The anti-BC effects of metformin have been illustrated in Fig. 2.
Fig. 2.

Anti-BC effects of metformin
Autophagy is another way to prevent the growth of malignant cells. Autophagy initiates the formation of membrane structures, including autophagosomes, by recruiting a family of autophagy-related (ATG) proteins [60]. It has been suggested that metformin may have antitumor effects in part due to its ability to increase levels of ATG3, ATG5, and LC3-II in cells treated with metformin [61]. In order to repurpose metformin for the treatment of BC, epigenetic regulators would be modulated. Previous studies showed that metformin treatment altered the abundance of RBBP4, G9a, acH3K9, and acH3K18, suggesting that histone modifiers may play an important role in metformin cancer treatment [62].
Obesity, diabetes, and hyperinsulinemia are associated with an increased risk of BC [63]. Fat distribution in the body, BMI, and weight changes are among the factors affecting BC [64]. Several factors such as adipokines, IGFs (IGF-1 as mitogens), dyslipidemia, hyperglycemia, hyperinsulinemia, and inflammatory cytokines link BC with obesity and diabetes [63]. IGF-1 gene expression increases in people with breast cancer compared to healthy people, while the circulating levels of Insulin-like growth factor binding protein 3 (IGFBP-3) will decrease [65]. The level of circulating insulin decreases in diabetes while it increases in obesity and cancers [66]. Insulin resistance is involved in obesity and diabetes and contributes to the development of BC. Metformin decreases insulin resistance, while obesity and diabetes both induce insulin resistance which leads to hyperinsulinemia. Hyperinsulinemia increases IGF-1 and decreases IGFBP1/2 (that finally cause dyslipidemia or increased bioavailable estrogen), or induce β-cells failure and hyperglycemia to finally increase BC cell growth [63]. Hyperglycemia increases the production of free radicals, damage and mutations in oncogenes and tumor suppressor genes and finally proliferation of cancer cells [67]. In addition, obesity and diabetes increase BC cell growth by upregulation of inflammatory cytokines such as IL-6, IL-1β, TNF-α [63].
Despite the fact that BC is well known to be associated with metabolic characteristics of T2D, including hyperglycemia, hyperinsulinemia, inflammation, oxidative stress, and obesity, randomized controlled trials show opposite results for metformin as an insulin sensitizer [59]. Considering that glucose is a crucial cellular metabolic substrate and insulin signaling has mutagenic effects on BC, growing and spreading BC are intimately linked to glucose metabolism [68]. The growing evidence suggests that metformin may play a preventative role in BC. Metformin counteracts insulin-stimulatory effects and leads to anti-proliferative and anti-migratory effects in primary breast cancer cells [69]. The first evidence that metformin might have antitumor properties was discovered by Evanns et al. in 2005 after they found that T2D patients who were taking metformin had a lower risk of malignancy [70].
In the meta-analyses on the association between metformin use and BC incidence, the preventive effect of metformin was observed in Col 2012 and Zhang 2013 meta-analyses [71]. However, the protective effect of metformin on BC prevention was not confirmed in subsequent meta-analyses [72–75]. The results are shown in Fig. 3.
Fig. 3.
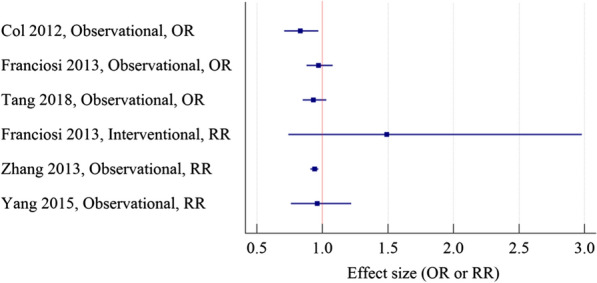
Meta-analyses on the association between metformin and BC incidence in diabetic patients. OR odds ratio, RR risk ratio
In a systematic review and meta-analysis conducted by Tang et al., the protective effect of metformin against BC mortality in patients with type 2 diabetes has been assessed. An approximate total of 3400 metformin users and 3000 non-metformin users were included. In the metformin user group, they found a 45% lower risk for all-cause mortality (HR = 0.55; 95% CI 0.44 to 0.70). However overall certainty of the evidence was very low and the included studies were only observational studies [73]. In a similar study conducted by Xu et al., the BC-specific survival time was better in the metformin user group (HR: 0.89; 95% CI 0.79 to 1.00). In diabetic patients who used metformin after BC diagnosis, a 36% risk reduction for cancer-specific mortality was reported (HR = 0.64; 95% CI 0.45 to 0.90). Also in BC patients who consumed metformin, a significant risk reduction of all-cause mortality compared with their non-diabetic parallel was observed (HR = 0.63; 95% CI 0.51 to 0.78) [42]. Overall, most studies demonstrate that metformin reduces BC mortality, especially in early-stage cancers. Nonetheless, caution is in order, as all of these studies were observational, and the results were heterogeneous. Despite observational systematic reviews and meta-analyses indicating better outcomes for metformin than placebo in BC patients (Fig. 4A) [42, 73–78], the adjuvant role of metformin was never confirmed in interventional systematic reviews and meta-analyses (Figs. 4B and 5) [78–82].
Fig. 4.
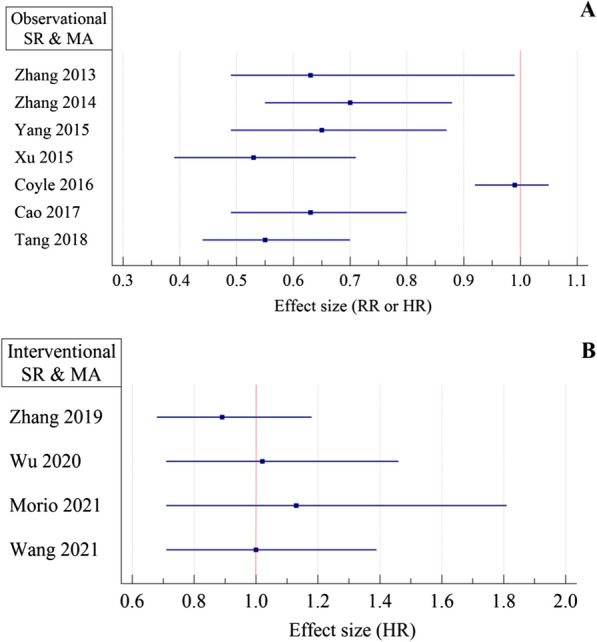
Observational meta-analyses on the association of metformin with all-cause mortality in BC patients. A metformin vs. non-metformin groups, B metformin vs. placebo, RR risk ratio, HR hazard ratio
Fig. 5.
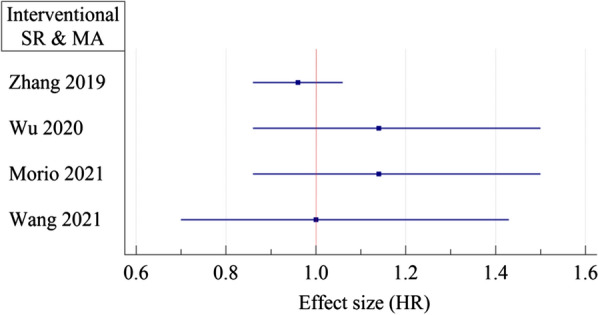
International meta-analyses on the association of metformin with progression-free survival of BC patients metformin vs. placebo. HR hazard ratio
In an interventional systematic review and meta-analysis conducted by Farkhondeh and colleagues for BC molecular markers, they discovered that metformin had no considerable effect on Ki-67 concentration (SMD = 0.08, 95% CI − 0.14 to 0.30) in the metformin group versus the placebo group [83]. In contrast, a similar study by Rahmani et al. showed a significant reduction in Ki-67 concentration (weighted MD = − 4.06, 95% CI − 7.59 to − 0.54) for the metformin group [84]. This inconsistency may relate to the different inclusion criteria of these studies.
Survival benefits of metformin in BC cohort studies have been reported in patients with positive ER or PR, HER2 overexpression, and high IGF-I receptor expression on the tumor cell surface [85]. According to a study conducted by Park et al. on 44,541 women, metformin was given to the majority of the diabetic population in the study (61%) as part of their treatment, they did not find any correlation between the use of metformin and the overall risk of BC after a median follow-up of 6–8 years. However, based on their findings, metformin therapy was associated with a reduced risk of estrogen receptor-positive (ER-positive) BC, and this inverse association was even more pronounced in a longer period (e.g., 10 years) treatment. The results of this study suggest that there is a T2D-dependent link between ER status and BC, and long-term metformin use might be able to reduce the association between T2D and ER-positive BC [86]. Also, an observational study from 23 Spanish hospitals found that metformin had a protective effect against ER-positive/HER2-negative BC when used as a daily treatment [87].
Finally, a large multicenter phase 3 RCT (the MA.32 RCT) for determining the adjuvant effect of metformin in patients with BC but without diabetes has been conducted recently. To the best of our knowledge, this study has the largest sample size for this topic even more than systematic reviews and meta-analyses (with 3649 enrolled patients and eight years of follow-up). The analysis demonstrated overall survival will not change with the addition of metformin vs placebo to the standard BC regimen (HR = 1.10; 95% CI 0.86 to 1.41; P = 0.47) [88].
LncRNAs in BC
The term lncRNAs refers to RNA molecules that contain over 200 nucleotides without encoding proteins. Many lncRNAs have a 5′ cap that makes their RNA structure more stable. The polyadenylation of the 3′ end of the lncRNA can also affect the stabilization of the structure of the lncRNA, but this can only be observed at certain points in the molecule [89]. The H19 and Xist genes were the first identified lncRNAs, classified as mRNAs at the time of their discovery [90]. It has been shown that lncRNA with long exons contains a number of exon regions, which allows the creation of different forms of this RNA family by splicing them together. In addition to their different functions, these forms may perform a variety of clinical roles as well [89]. The structure of lncRNAs allows them to escape the evolutionary limitations associated with poor interspecies conservation. Furthermore, the low conservation of the lncRNA sequence is likely to allow the structure of the lncRNA to be variable, to activate lncRNA functions and specializations as regulators within the cell [91]. The fact that lncRNAs have a high degree of specificity can be attributed to their key role in regulating organism function as well as in repairing disease processes in different conditions [92].
Originally, lncRNAs were thought to be genomic noise without any biological function. Recently, researchers have begun focusing their attention on the role lncRNAs. The interaction between RNA–protein, RNA–RNA, and DNA–RNA can be associated with lncRNA and form different functional complexes. As a result of their ability to regulate mRNA stability, translation, and cell signaling pathways, lncRNA can perform a variety of functions within the cell [93]. LncRNAs can be classified into five groups based on their closest protein-coding transcripts: sense, antisense, bidirectional, intronic, and intergenic [9]. In general, lncRNAs act in four distinct ways as signal, decoy, guide, and scaffold molecules [94]. lncRNAs are often thought to regulate downstream gene transcription as signal molecules, for example, lincRNA-p21 regulates JAK2/STAT3, Notch signaling. Occasionally, lncRNAs can also act as a decoy molecule to block certain molecular pathways. lncRNAs can inhibit the expression of mRNA by interacting with certain proteins. Decoying transcriptional factors or miRNAs, RNA interference, targeting of transcriptional factors, or chromatin modifier proteins to specific genomic loci, and transcriptional regulation in cis or trans [16]. In order to regulate gene transcription, lncRNAs can also act as a guide molecule that interacts with transcription factors on specific sequences of DNA and recruit chromatin-modifying enzymes for target genes, chromatin remodeling, and their epigenetic regulation [16, 95]. In addition, lncRNAs as scaffold molecules facilitate the assembly of various kinds of macromolecular complexes, promoting information, integration, and convergence [96]. Regulation of stabilized ribonucleoprotein complexes such as signaling molecules and nuclear structures is considered among the scaffolding activities of lncRNAs. The expression of genes can be regulated by lncRNAs at several levels, including epigenetics, transcription, and post-transcriptional regulation [97]. Contrary to mRNA, lncRNAs are found throughout the cell, not only in the nucleus but also in the cytoplasm and mitochondria [98]. There has been evidence that diseases linked to single-nucleotide polymorphisms within lncRNA genes and their promoters are associated with change in lncRNA expression which highlights their significance in the pathogenesis of the disease. The lncRNAs act as guides, scaffolds, or stabilizers that affect chromatin architecture and gene expression through interactions with epigenetic remodelers, transcription factors, and spliceosomes in the nucleus [99]. As lncRNA decoys, cytoplasmic lncRNAs regulate the stability of mRNA by directly regulating de-adenylation [93]. Xist is an important example of nuclear lncRNA which play an important role in females’ X chromosome by directing methyltransferases to the X chromosome. Xist facilitates macrophage polarization in breast and ovarian cancer. This function could be due to the regulation of C/EBPα and KLF6 by miR-101 [100]. There have been numerous studies on the association of lncRNAs and different types of cancers which make lncRNAs interesting targets for unique therapeutic and diagnostic pathways. LncRNAs play important role in cancer by regulating transcription and chromatin remodeling through interactions with chromatin remodelers like polycomb complex. For example, in BC, lncRNA PANDAR plays a key role in G1 to S phase regulation [101]. Regarding metastatic breast carcinomas, HOTAIR has changed the pattern of PRC2 occupancy, causing it to shift from breast epithelial cells to embryonic fibroblasts due to its ability to alter chromatin [102].
Normal development of breast stem cells is driven by some of the same signaling pathways, including estrogen receptors, HER2, and Wnt/b-catenin signaling pathways that control stem cell proliferation, cell death, cell differentiation, and cell motility. Regulation of BC heterogeneity and plasticity is one of the most urgent issues for treatment. Studies confirm that epigenetic regulation and non-coding RNAs may play an important role in BC development and may contribute to the heterogeneous and metastatic aspects of BC, especially triple-negative BC [11]. Many studies have investigated the association between lncRNAs and BC. In addition to basic research, the clinical application of lncRNAs is also an emerging research field [16]. lncRNAs expressed in cancers play essential roles in cancer-related biological processes and signaling pathways, regulating gene expression, post-transcriptional processes, chromatin changes, and regulation of protein function. Studies show that the distribution and activity of lncRNAs and their role in human cancers can be confirmed by transcription profiling studies [103]. Most of the activity of lncRNA is related to transcription. These activities include the formation of chromatin-modifying complexes, transcription activators, and chromatin ring regulators. Regulating the transcription of tumor suppressor and oncogene genes is one of the functions related to lncRNA oncology [103]. Some lncRNAs target DICER or pre-miRNA and hinder miRNA biogenesis that influences BC metastasis. For example overexpression of oncogenic lncRNA LINC01787 promotes BC cell proliferation, migration, and BC xenograft growth in vivo by repressing the maturation of miR-125b. The upregulation of LINC00899 represses the proliferation, migration, and invasion of BC cells by inhibiting miR-425 [9]. One of the main causes of death associated with BC is the metastasis of the primary tumor. lncRNAs are additional transcripts related to metastasis and cancer progression. Due to the necessary advances in transcriptome analysis technology, many articles confirm the expression of lncRNA in tumors and their correlation with metastatic conditions [104]. Since lncRNAs have a variable expression in cancer tissues compared to normal tissues, it raises the potential of these molecules as biomarkers for disease diagnosis. lncRNAs can alter multiple signaling pathways and regulate metastasis-related factors, alter the proteins, and transcription factors involved in metastasis. They can be used as an early diagnostic and therapeutic target for BC metastasis and therapy. For example, anti-metastatic lncRNAs can target oncogenes and inhibit metastasis whereas some prometastatic lncRNA reduce the expression of tumor suppressor genes and induce invasion and metastasis. Antisense oligonucleotides (ASOs) can reduce the expression of oncogenic lncRNAs and inhibit BC metastasis by degrading lncRNAs, cleaving endogenous RNaseH1, or regulating RNA–protein interactions [9]. Different novel technologies target lncRNAs in cancer therapy by small molecule inhibitors which are new directions in anti-tumor drug development, including targeting cytoplasmic and nuclear lncRNA by ASOs through RNaseH-dependent degradation to knock out lncRNAs, nanomedicine role of lncRNAs is associated with nano-carrier-absorbed on nano-drugs that target sub-nucleus lncRNAs to gain desired therapeutic effect and in cancer cell chemical resistance to different types of drugs. Other approaches include knock-out of lncRNA via specific gDNA by CRISPR/cas9 as a technology with extensive application in cancer treatment, and finally targeting lncRNAs by virus therapies including encapsulated shRNA to target tumor suppressor lncRNAs or natural antisense RNAs (NATs)-mediated technique [105, 106].
Generally, BC cells associated lncRNAs can be classified into two groups of enhance or attenuate the aggressiveness of BC cells, for example, DANCR, H19, HOTAIR, LINC00152, LINC00461, NEAT1, and LINC01857 cause invasion and migration of BC cells, H19, HOTAIR, HIF1A-AS2, RP1, and MALAT1 promote distant metastasis of BC cells, GAS5, MT1JP, NEF, NKILA, LET, TFAP2A-AS1, LncKLHDC7B prevent the invasion and migration of BC cells, MALAT1, MEG3, NLIPMT, and XIST inhibit distant metastasis of BC cells [16]. Also, in some studies, immune-related lncRNA signatures were associated with survival of BC [107–109], or the significant dysregulation of lncRNAs in BC was not associated with any of the clinical features [110]. Our study investigated the role of lncRNAs on BC in more than 40 general effects including metastasis, migration, invasion, progression, tumor growth, apoptosis, stemness, EMT, survival, histological grade, tumor size, tumor stage, proliferation, chemoresistance, radioresistance, drug resistance, endocrine resistance, disease recurrence, viability, differentiation, tumor initiation, aggressiveness, cell cycle, prognosis, diagnosis, tumor values, steroid receptor status, hormone negativity, tumourigenesis, colonization, invasion-metastasis cascade, colony formation, angiogenesis, motility, mobility, biological activities of BC cells, pathogenesis, clinicopathological features, clinical outcome, tumorigenic properties. Finally, 116 lncRNAs associated with these BC features were identified. In general most of lncRNAs were associated with invasion, migration, metastasis, and proliferation. The detailed results are presented in Table 1 and Additional file 1: Table S1.
Metformin action on lncRNAs in BC
Some of the lncRNAs listed in Table 1 including GAS5, HOTAIR, TUG1, MALAT1, and H19 are also associated with the effect of metformin on BC. As described in the antitumor mechanisms of metformin, it effects on mTOR signaling pathway. The inhibition of the mTOR is responsible for regulating tumor growth [48]. mTOR is related to liver kinase B1 (LKB1), a major downstream kinase of AMPK. There has been evidence that deletion of LKB1 function in tumor cells accelerated their proliferation and resulted in them becoming more sensitive to AMPK pathway activators, such as biguanide (metformin) in both in vitro and in vivo studies [227, 228]. Metformin inhibits the over-activation of this pathway through upregulating lncRNA GAS5 expression, and finally induces apoptosis and inhibits the growth of BC cells [229]. As GAS5 is associated with invasion, growth, tumor promotion, proliferation, and apoptosis of BC (Table 1 and Additional file 1: Table S1), metformin can regulate these features by GAS5 antitumor activity. Metformin reverses EMT by inducing DNA methylation of the CpG-rich sequence at the gene downstream region in HOTAIR and downregulating the HOTAIR oncogenic expression in MDA-MB-231 BC cells [230]. The HOTAIR is associated with progression, metastasis, prognosis, cell growth, migration, invasion, apoptosis, and EMT in BC (Table 1 and Additional file 1: Table S1), so metformin can regulate these features by changing HOTAIR expression. Crosstalk between autophagy and Wnt/β-catenin signaling is frequent and is directly related to cell homeostasis [231]. There is also strong evidence from the current studies that inhibiting EMT results in the inactivation of Wnt/β-catenin, a primary mediator of EMT [232]. In breast cancer cells, metformin might also inhibit cell migration by inactivating Wnt/β-catenin [233]. Metformin treatment resulted in elevated endoplasmic reticulum stress, which was further intensified by knocking down MALAT1 expression [234]. MALAT1 is involved in metformin inhibitory role in the BC cells proliferation. Metformin increases the expression of HOTAIR, MALAT1, TUG1, LINC01121, and DICER1-AS1 in BC cells. MALAT1 knock-down in metformin-treated BC cells will increase the Bax/Bcl2 ratio and p21, Beclin1, VDAC1, LC3-II, CHOP and Bip expressions will decrease cyclin B1 [62]. As MALAT1 and TUG1 are associated with migration, invasion, metastasis, progression, EMT, relapse-free survival, proliferation, angiogenesis, motility, apoptosis, tumor growth, and tumor size of BC (Table 1 and Additional file 1: Table S1), metformin can regulate these features by MALAT1 and TUG1 activities. H19 is a transcription product of the H19 gene. H19 potentially influences gene expression in BC on several levels, including epigenetic regulation, transcriptional regulation, and posttranscriptional regulation. In BC tumorigenesis and progression, abnormal expression of H19 is governed by a number of molecular mechanisms, including microRNA-675 encoding, competing with endogenous RNA regulation, and interacting with Myc [235, 236]. Angiogenesis, cell death, inflammation, and apoptosis are some of the features that H19 stimulates [237]. There is growing evidence that ferroptosis, an emerging form of cell death that suppresses drug resistance and enhances the immune system’s ability to combat tumors, could be considered a new form of programmed cell death. In terms of cell necrosis, ferroptosis is defined as a process that depends on iron [238]. In case of ferroptosis, the membrane structure can be damaged owing to the formation of lipid peroxidation, which results in oxidative damage to the phospholipids, and in addition, there can be a high concentration of unsaturated fatty acids in the membrane [239]. One of the most common symptoms of ferroptosis is the excess of reactive oxygen species (ROS), which can be caused by metformin [240]. Metformin may induce ferroptosis by inhibiting autophagy via lncRNA H19 in BC subjects [238]. The effect of metformin on H19 lead to change in proliferation, invasion, migration, metastasis, cell cycle arrest, apoptosis, steroid receptor status, tumor size, disease-free survival, prognosis, stemness, EMT, and MET features of BC patients.
lncRNAs UCA1, H19, MALAT1, AFAP1-AS1, AC026904.1, and SNHG7 presented in Table 1 are associated with the effect of metformin on other types of cancer. Metformin promotes apoptosis and inhibits the proliferation of colon cancer cells by inhibiting UCA1 expression [241]. Metformin inhibits tumor cell invasion and migration partly by H19 downregulation [242] and decreases the expression of H19 in Endometrial Cancer [243]. Metformin decreases migration and invasion of cervical cancer cells by suppressing MALAT1 and disrupting of MALAT1/miR-142-3p sponge [244]. Metformin suppresses lung adenocarcinoma by downregulating AFAP1-AS1 and regulating the AFAP1-AS1/miR-3163/SPP1/PI3K/Akt/mTOR axis. [245]. Metformin suppresses hypopharyngeal cancer growth by decreasing SNHG7 expression through activating SAHH [246].
Other lncRNAs including AC006160.1, Loc100506691, lncRNA-AF085935, and HULC are also associated with cancers and metformin. Bladder cancer patients with high expression of AC006160.1 are sensitive to metformin [247]. Metformin anti-proliferative effects in gastric cancer may be associated with suppression of Loc100506691 (an oncogenic lncRNA) and Loc100506691-miR-26a-5p/miR-330-5p-CHAC1 axis [248]. In HepG2cells, metformin and EGCG combination shows anticarcinogenic effects by changes in proliferation, lncRNA-AF085935 expression, and apoptosis [249]. Metformin reduces HULC overexpression to inhibit HBV-induced hepatocellular carcinoma tumorigenesis [250].
Supplementary Information
Additional file 1: Table S1. lncRNAs associated with BC (continuation of Table 1).
Acknowledgements
Not applicable.
Abbreviations
- BC
Breast cancer
- lncRNA
Long non-coding RNAs
- T2D
Type 2 diabetes
- TNBC
Triple-negative breast cancer
- HER2
Human epidermal growth factor receptor 2
- MET
Mesenchymal-to-epithelial transition
- EMT
Epithelial-to-mesenchymal transition
- MMP-9
Matrix metalloproteinase 9
- P53
Tumor protein 53
- CTCs
Circulating tumor cells
- piRNAs
P-element-induced wimpy testis interacting RNAs
- ER
Estrogen receptor
- PR
Progesterone receptor
- mTOR
Mechanistic target of rapamycin
- EGFR
Epidermal growth factor receptor
- AMPK
AMP-activated protein kinase
- MAPK
Mitogen-activated protein kinases
- PKB
Phosphorylation of protein kinase B
- GRB2
Growth factor receptor-bound protein 2
- ERK
Signal-regulated kinase
- NFκB
Nuclear factor kappa
- STAT3
Signal transducer activator of transcription 3
- HIF1α
Hypoxia-inducible factor 1-alpha
- c-Myc
Cellular-Myc
- DICER
Double-stranded RNA specific endoribonuclease
- ACC
Acetyl-CoA carboxylase
- FASN
Fatty acid synthase
- IGFBP-3
Insulin-like growth factor binding protein 3
- NATs
Natural antisense RNAs
- ATG
Autophagy-related
- ER-positive
Estrogen receptor-positive
- ASOs
Antisense oligonucleotides
- LKB1
Liver kinase B1
- ROS
Reactive oxygen species
Author contributions
All authors contributed to the conception and design of the study. The literature review was performed by MG and ZNK. “BC” section was written by ASF, PE, and MG. “Metformin” section was written by AAM, MZ, and MA. “LncRNAs in BC” section was written by MG, AV, and ZNK. “Metformin action on lncRNAs in BC” section was written by MG, PE, and MMA. All authors commented on previous versions of the manuscript. MMA supervised the project and approved the final version. All authors read and approved the final manuscript.
Funding
This study was supported by Tehran University of Medical Sciences (1401-4-221-63487).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Basic Information About Breast Cancer | CDC. https://www.cdc.gov/cancer/breast/basic_info/index.htm. Accessed 10 July 2022.
- 2.Libby G, Donnelly LA, Donnan PT, Alessi DR, Morris AD, Evans JM. New users of metformin are at low risk of incident cancer: a cohort study among people with type 2 diabetes. Diabetes Care. 2009;32(9):1620–1625. doi: 10.2337/dc08-2175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Han B, Cui H, Kang L, Zhang X, Jin Z, Lu L, et al. Metformin inhibits thyroid cancer cell growth, migration, and EMT through the mTOR pathway. Tumor Biol. 2015;36(8):6295–6304. doi: 10.1007/s13277-015-3315-4. [DOI] [PubMed] [Google Scholar]
- 4.Qu C, Zhang W, Zheng G, Zhang Z, Yin J, He Z. Metformin reverses multidrug resistance and epithelial–mesenchymal transition (EMT) via activating AMP-activated protein kinase (AMPK) in human breast cancer cells. Mol Cell Biochem. 2014;386(1):63–71. doi: 10.1007/s11010-013-1845-x. [DOI] [PubMed] [Google Scholar]
- 5.Jalving M, Gietema JA, Lefrandt JD, de Jong S, Reyners AK, Gans RO, et al. Metformin: taking away the candy for cancer? Eur J Cancer. 2010;46(13):2369–2380. doi: 10.1016/j.ejca.2010.06.012. [DOI] [PubMed] [Google Scholar]
- 6.Liu B, Fan Z, Edgerton SM, Deng X-S, Alimova IN, Lind SE, et al. Metformin induces unique biological and molecular responses in triple negative breast cancer cells. Cell Cycle. 2009;8(13):2031–2040. doi: 10.4161/cc.8.13.8814. [DOI] [PubMed] [Google Scholar]
- 7.Chen TW, Liang YN, Feng D, Tao LY, Qi K, Zhang HY, et al. Metformin inhibits proliferation and promotes apoptosis of HER2 positive breast cancer cells by downregulating HSP90. J BUON. 2013;18(1):51–56. [PubMed] [Google Scholar]
- 8.Breast Cancer Statistics | How common is breast cancer? https://www.cancer.org/cancer/breast-cancer/about/how-common-is-breast-cancer.html. Accessed 10 July 2022.
- 9.Mondal P, Meeran SM. Long non-coding RNAs in breast cancer metastasis. Non-coding RNA Res. 2020;5(4):208–218. doi: 10.1016/j.ncrna.2020.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zeind CS, Carvalho MG. Koda-Kimble and Young’s applied therapeutics: the clinical use of drugs. 11. Philadelphia: LWW; 2018. [Google Scholar]
- 11.Feng Y, Spezia M, Huang S, Yuan C, Zeng Z, Zhang L, et al. Breast cancer development and progression: risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018;5(2):77–106. doi: 10.1016/j.gendis.2018.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Types of Breast Cancer | About breast cancer. https://www.cancer.org/cancer/breast-cancer/about/types-of-breast-cancer.html. Accessed 15 July 2022.
- 13.De Francesco EM, Cirillo F, Vella V, Belfiore A, Maggiolini M, Lappano R. Triple-negative breast cancer drug resistance, durable efficacy, and cure: how advanced biological insights and emerging drug modalities could transform progress. Expert Opin Ther Targets. 2022;26(6):513–535. doi: 10.1080/14728222.2022.2094762. [DOI] [PubMed] [Google Scholar]
- 14.Jaiyesimi IA, Buzdar AU, Hortobagyi G. Inflammatory breast cancer: a review. J Clin Oncol. 1992;10(6):1014–1024. doi: 10.1200/JCO.1992.10.6.1014. [DOI] [PubMed] [Google Scholar]
- 15.Parton M, Dowsett M, Smith I. Studies of apoptosis in breast cancer. BMJ. 2001;322(7301):1528–1532. doi: 10.1136/bmj.322.7301.1528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Huang Q-Y, Liu G-F, Qian X-L, Tang L-B, Huang Q-Y, Xiong L-X. Long non-coding RNA: dual effects on breast cancer metastasis and clinical applications. Cancers. 2019;11(11):1802. doi: 10.3390/cancers11111802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zibara K, Awada Z, Dib L, El-Saghir J, Al-Ghadban S, Ibrik A, et al. Anti-angiogenesis therapy and gap junction inhibition reduce MDA-MB-231 breast cancer cell invasion and metastasis in vitro and in vivo. Sci Rep. 2015;5(1):1–16. doi: 10.1038/srep12598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lappano R, Rigiracciolo DC, Belfiore A, Maggiolini M, De Francesco EM. Cancer associated fibroblasts: role in breast cancer and potential as therapeutic targets. Expert Opin Ther Targets. 2020;24(6):559–572. doi: 10.1080/14728222.2020.1751819. [DOI] [PubMed] [Google Scholar]
- 19.Giuli M, Giuliani E, Screpanti I, Bellavia D, Checquolo S. Notch signaling activation as a hallmark for triple-negative breast cancer subtype. J Oncol. 2019 doi: 10.1155/2019/8707053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Boudreau N, Myers C. Breast cancer-induced angiogenesis: multiple mechanisms and the role of the microenvironment. Breast Cancer Res. 2003;5(3):1–7. doi: 10.1186/bcr589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Santolla MF, Maggiolini M. The FGF/FGFR system in breast cancer: oncogenic features and therapeutic perspectives. Cancers. 2020;12(10):3029. doi: 10.3390/cancers12103029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li C-J, Chen H-M, Lai J-C. Diagnostic, prognostic, and predictive biomarkers in breast cancer. J Oncol. 2020 doi: 10.1155/2020/1835691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Afzal S, Hassan M, Ullah S, Abbas H, Tawakkal F, Khan MA. Breast cancer; discovery of novel diagnostic biomarkers, drug resistance, and therapeutic implications. Front Mol Biosci. 2022;9:783450. doi: 10.3389/fmolb.2022.783450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kopreski MS, Benko FA, Gocke CD. Circulating RNA as a tumor marker: detection of 5T4 mRNA in breast and lung cancer patient serum. Ann N Y Acad Sci. 2001;945(1):172–178. doi: 10.1111/j.1749-6632.2001.tb03882.x. [DOI] [PubMed] [Google Scholar]
- 25.Yin W-B, Yan M-G, Fang X, Guo J-J, Xiong W, Zhang R-P. Circulating circular RNA hsa_circ_0001785 acts as a diagnostic biomarker for breast cancer detection. Clin Chim Acta. 2018;487:363–368. doi: 10.1016/j.cca.2017.10.011. [DOI] [PubMed] [Google Scholar]
- 26.Souza KC, Evangelista AF, Leal LF, Souza CP, Vieira RA, Causin RL, et al. Identification of cell-free circulating microRNAs for the detection of early breast cancer and molecular subtyping. J Oncol. 2019 doi: 10.1155/2019/8393769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Luo D, Wilson JM, Harvel N, Liu J, Pei L, Huang S, et al. A systematic evaluation of miRNA: mRNA interactions involved in the migration and invasion of breast cancer cells. J Transl Med. 2013;11(1):1–14. doi: 10.1186/1479-5876-11-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Masuda M, Miki Y, Hata S, Takagi K, Sakurai M, Ono K, et al. An induction of microRNA, miR-7 through estrogen treatment in breast carcinoma. J Transl Med. 2012;10(1):1–9. doi: 10.1186/1479-5876-10-S1-S2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chang C-H, Fan T-C, Yu J-C, Liao G-S, Lin Y-C, Shih AC-C, et al. The prognostic significance of RUNX2 and miR-10a/10b and their inter-relationship in breast cancer. J Transl Med. 2014;12(1):1–12. doi: 10.1186/s12967-014-0257-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Vivacqua A, Muoio MG, Miglietta AM, Maggiolini M. Differential MicroRNA landscape triggered by estrogens in cancer associated fibroblasts (CAFs) of primary and metastatic breast tumors. Cancers. 2019;11(3):412. doi: 10.3390/cancers11030412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Qian L, Xie H, Zhang L, Zhao Q, Lü J, Yu Z. Piwi-interacting RNAs: a new class of regulator in human breast cancer. Front Oncol. 2021;11:695077. doi: 10.3389/fonc.2021.695077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fu A, Jacobs DI, Hoffman AE, Zheng T, Zhu Y. PIWI-interacting RNA 021285 is involved in breast tumorigenesis possibly by remodeling the cancer epigenome. Carcinogenesis. 2015;36(10):1094–1102. doi: 10.1093/carcin/bgv105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rojas LBA, Gomes MB. Metformin: an old but still the best treatment for type 2 diabetes. Diabetol Metab Syndr. 2013;5(1):1–15. doi: 10.1186/1758-5996-5-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Faria J, Negalha G, Azevedo A, Martel F. Metformin and breast cancer: molecular targets. J Mammary Gland Biol Neoplasia. 2019;24(2):111–123. doi: 10.1007/s10911-019-09429-z. [DOI] [PubMed] [Google Scholar]
- 35.Bailey CJ. Metformin: historical overview. Diabetologia. 2017;60(9):1566–1576. doi: 10.1007/s00125-017-4318-z. [DOI] [PubMed] [Google Scholar]
- 36.Dowling RJ, Goodwin PJ, Stambolic V. Understanding the benefit of metformin use in cancer treatment. BMC Med. 2011;9(1):1–6. doi: 10.1186/1741-7015-9-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Beysel S, Unsal IO, Kizilgul M, Caliskan M, Ucan B, Cakal E. The effects of metformin in type 1 diabetes mellitus. BMC Endocr Disord. 2018;18(1):1. doi: 10.1186/s12902-017-0228-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Drzewoski J, Hanefeld M. The current and potential therapeutic use of metformin—the good old drug. Pharmaceuticals (Basel) 2021;14(2):122. doi: 10.3390/ph14020122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.American Diabetes Association 8. Pharmacologic approaches to glycemic treatment: standards of medical care in diabetes—2018. Diabetes Care. 2018;41(Suppl 1):S73–S85. doi: 10.2337/dc18-S008. [DOI] [PubMed] [Google Scholar]
- 40.Leonaviciute D, Madsen B, Schmedes A, Buus NH, Rasmussen BS. Severe metformin poisoning successfully treated with simultaneous venovenous hemofiltration and prolonged intermittent hemodialysis. Case Rep Crit Care. 2018;2018:3868051. doi: 10.1155/2018/3868051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Cazzaniga M, Bonanni B, Guerrieri-Gonzaga A, Decensi A. Is it time to test metformin in breast cancer clinical trials? Cancer Epidemiol Biomark Prev. 2009;18(3):701–705. doi: 10.1158/1055-9965.EPI-08-0871. [DOI] [PubMed] [Google Scholar]
- 42.Xu H, Chen K, Jia X, Tian Y, Dai Y, Li D, et al. Metformin use is associated with better survival of breast cancer patients with diabetes: a meta-analysis. Oncologist. 2015;20(11):1236–1244. doi: 10.1634/theoncologist.2015-0096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Perl A. Activation of mTOR (mechanistic target of rapamycin) in rheumatic diseases. Nat Rev Rheumatol. 2016;12(3):169–182. doi: 10.1038/nrrheum.2015.172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Vazquez-Martin A, López-Bonet E, Oliveras-Ferraros C, Pérez-Martínez MC, Bernadó L, Menendez JA. Mitotic kinase dynamics of the active form of AMPK (phospho-AMPKαThr172) in human cancer cells. Cell Cycle. 2009;8(5):788–791. doi: 10.4161/cc.8.5.7787. [DOI] [PubMed] [Google Scholar]
- 45.Dowling RJ, Niraula S, Chang MC, Done SJ, Ennis M, McCready DR, et al. Changes in insulin receptor signaling underlie neoadjuvant metformin administration in breast cancer: a prospective window of opportunity neoadjuvant study. Breast Cancer Res. 2015;17(1):1–12. doi: 10.1186/s13058-015-0540-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Memmott RM, Mercado JR, Maier CR, Kawabata S, Fox SD, Dennis PA. Metformin prevents tobacco carcinogen-induced lung tumorigenesis. Cancer Prev Res (Phila) 2010;3(9):1066–1076. doi: 10.1158/1940-6207.CAPR-10-0055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Drzewoski J, Drozdowska A, Sliwińska A. Do we have enough data to confirm the link between antidiabetic drug use and cancer development? Pol Arch Med Wewn. 2011;121(3):81–87. [PubMed] [Google Scholar]
- 48.Kasznicki J, Sliwinska A, Drzewoski J. Metformin in cancer prevention and therapy. Ann Transl Med. 2014;2(6):57. doi: 10.3978/j.issn.2305-5839.2014.06.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.LeRoith D, Baserga R, Helman L, Roberts CT., Jr Insulin-like growth factors and cancer. Ann Intern Med. 1995;122(1):54–59. doi: 10.7326/0003-4819-122-1-199501010-00009. [DOI] [PubMed] [Google Scholar]
- 50.Kalender A, Selvaraj A, Kim SY, Gulati P, Brûlé S, Viollet B, et al. Metformin, independent of AMPK, inhibits mTORC1 in a rag GTPase-dependent manner. Cell Metab. 2010;11(5):390–401. doi: 10.1016/j.cmet.2010.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Vazquez-Martin A, Oliveras-Ferraros C, Menendez JA. The antidiabetic drug metformin suppresses HER2 (erbB-2) oncoprotein overexpression via inhibition of the mTOR effector p70S6K1 in human breast carcinoma cells. Cell Cycle. 2009;8(1):88–96. doi: 10.4161/cc.8.1.7499. [DOI] [PubMed] [Google Scholar]
- 52.Godsland IF. Insulin resistance and hyperinsulinaemia in the development and progression of cancer. Clin Sci. 2010;118(5):315–332. doi: 10.1042/CS20090399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wisastra R, Dekker FJ. Inflammation, cancer and oxidative lipoxygenase activity are intimately linked. Cancers. 2014;6(3):1500–1521. doi: 10.3390/cancers6031500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hirsch HA, Iliopoulos D, Struhl K. Metformin inhibits the inflammatory response associated with cellular transformation and cancer stem cell growth. Proc Natl Acad Sci. 2013;110(3):972–977. doi: 10.1073/pnas.1221055110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ma J, Guo Y, Chen S, Zhong C, Xue Y, Zhang Y, et al. Metformin enhances tamoxifen-mediated tumor growth inhibition in ER-positive breast carcinoma. BMC Cancer. 2014;14(1):1–11. doi: 10.1186/1471-2407-14-172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Arai M, Uchiba M, Komura H, Mizuochi Y, Harada N, Okajima K. Metformin, an antidiabetic agent, suppresses the production of tumor necrosis factor and tissue factor by inhibiting early growth response factor-1 expression in human monocytes in vitro. J Pharmacol Exp Ther. 2010;334(1):206–213. doi: 10.1124/jpet.109.164970. [DOI] [PubMed] [Google Scholar]
- 57.Blandino G, Valerio M, Cioce M, Mori F, Casadei L, Pulito C, et al. Metformin elicits anticancer effects through the sequential modulation of DICER and c-MYC. Nat Commun. 2012;3(1):1–11. doi: 10.1038/ncomms1859. [DOI] [PubMed] [Google Scholar]
- 58.Wahdan-Alaswad RS, Thor AD. Metformin activity against breast cancer: mechanistic differences by molecular subtype and metabolic conditions. Metformin. London: IntechOpen; 2020. [Google Scholar]
- 59.Cejuela M, Martin-Castillo B, Menendez JA, Pernas S. Metformin and breast cancer: where are we now? Int J Mol Sci. 2022;23(5):2705. doi: 10.3390/ijms23052705. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 60.Kaleağasıoğlu F, Ali DM, Berger MR. Multiple facets of autophagy and the emerging role of alkylphosphocholines as autophagy modulators. Front Pharmacol. 2020;11:547. doi: 10.3389/fphar.2020.00547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Lu G, Wu Z, Shang J, Xie Z, Chen C, Zhang C. The effects of metformin on autophagy. Biomed Pharmacother. 2021;137:111286. doi: 10.1016/j.biopha.2021.111286. [DOI] [PubMed] [Google Scholar]
- 62.Huang Y, Zhou Z, Zhang J, Hao Z, He Y, Wu Z, et al. lncRNA MALAT1 participates in metformin inhibiting the proliferation of breast cancer cell. J Cell Mol Med. 2021;25(15):7135–7145. doi: 10.1111/jcmm.16742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kang C, LeRoith D, Gallagher EJ. Diabetes, obesity, and breast cancer. Endocrinology. 2018;159(11):3801–3812. doi: 10.1210/en.2018-00574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Eliassen AH, Colditz GA, Rosner B, Willett WC, Hankinson SE. Adult weight change and risk of postmenopausal breast cancer. JAMA. 2006;296(2):193–201. doi: 10.1001/jama.296.2.193. [DOI] [PubMed] [Google Scholar]
- 65.Yu H, Rohan T. Role of the insulin-like growth factor family in cancer development and progression. J Natl Cancer Inst. 2000;92(18):1472–1489. doi: 10.1093/jnci/92.18.1472. [DOI] [PubMed] [Google Scholar]
- 66.Zhang AM, Wellberg EA, Kopp JL, Johnson JD. Hyperinsulinemia in obesity, inflammation, and cancer. Diabetes Metab J. 2021;45(3):285–311. doi: 10.4093/dmj.2020.0250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Collins KK. The diabetes-cancer link. Diabetes Spectrum. 2014;27(4):276–280. doi: 10.2337/diaspect.27.4.276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Roshan MH, Shing YK, Pace NP. Metformin as an adjuvant in breast cancer treatment. SAGE Open Med. 2019;7:2050312119865114. doi: 10.1177/2050312119865114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Scordamaglia D, Cirillo F, Talia M, Santolla MF, Rigiracciolo DC, Muglia L, et al. Metformin counteracts stimulatory effects induced by insulin in primary breast cancer cells. J Transl Med. 2022;20(1):1–19. doi: 10.1186/s12967-022-03463-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Evans JM, Donnelly LA, Emslie-Smith AM, Alessi DR, Morris AD. Metformin and reduced risk of cancer in diabetic patients. BMJ. 2005;330(7503):1304–1305. doi: 10.1136/bmj.38415.708634.F7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Col NF, Ochs L, Springmann V, Aragaki AK, Chlebowski RT. Metformin and breast cancer risk: a meta-analysis and critical literature review. Breast Cancer Res Treat. 2012;135(3):639–646. doi: 10.1007/s10549-012-2170-x. [DOI] [PubMed] [Google Scholar]
- 72.Franciosi M, Lucisano G, Lapice E, Strippoli GF, Pellegrini F, Nicolucci A. Metformin therapy and risk of cancer in patients with type 2 diabetes: systematic review. PLoS ONE. 2013;8(8):e71583. doi: 10.1371/journal.pone.0071583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Tang GH, Satkunam M, Pond GR, Steinberg GR, Blandino G, Schünemann HJ, et al. Association of metformin with breast cancer incidence and mortality in patients with type II diabetes: a GRADE-assessed systematic review and meta-analysismetformin therapy and breast cancer incidence and mortality. Cancer Epidemiol Biomark Prev. 2018;27(6):627–635. doi: 10.1158/1055-9965.EPI-17-0936. [DOI] [PubMed] [Google Scholar]
- 74.Zhang P, Li H, Tan X, Chen L, Wang S. Association of metformin use with cancer incidence and mortality: a meta-analysis. Cancer Epidemiol. 2013;37(3):207–218. doi: 10.1016/j.canep.2012.12.009. [DOI] [PubMed] [Google Scholar]
- 75.Yang T, Yang Y, Liu S. Association between metformin therapy and breast cancer incidence and mortality: evidence from a meta-analysis. J Breast Cancer. 2015;18(3):264–270. doi: 10.4048/jbc.2015.18.3.264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Zhang ZJ, Li S. The prognostic value of metformin for cancer patients with concurrent diabetes: a systematic review and meta-analysis. Diabetes Obes Metab. 2014;16(8):707–710. doi: 10.1111/dom.12267. [DOI] [PubMed] [Google Scholar]
- 77.Coyle C, Cafferty F, Vale C, Langley R. Metformin as an adjuvant treatment for cancer: a systematic review and meta-analysis. Ann Oncol. 2016;27(12):2184–2195. doi: 10.1093/annonc/mdw410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Cao X, Wu Y, Wang J, Liu K, Wang X. The effect of metformin on mortality among diabetic cancer patients: a systematic review and meta-analysis. JNCI Cancer Spectrum. 2017;1(1):pkx007. doi: 10.1093/jncics/pkx007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Wu Z, Qu B, Huang X, Song Y, Gao P, Shi J, et al. The potential adjunctive benefit of adding metformin to standard treatment in inoperable cancer patients: a meta-analysis of randomized controlled trials. Ann Transl Med. 2020;8(21):1404. doi: 10.21037/atm-20-4441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Morio K, Kurata Y, Kawaguchi-Sakita N, Shiroshita A, Kataoka Y. Efficacy of metformin in patients with breast cancer receiving chemotherapy or endocrine therapy: systematic review and meta-analysis. Ann Pharmacother. 2022;56(3):245–255. doi: 10.1177/10600280211025792. [DOI] [PubMed] [Google Scholar]
- 81.Wang Q, Ma X, Long J, Du X, Pan B, Mao H. Metformin and survival of women with breast cancer: a meta-analysis of randomized controlled trials. J Clin Pharm Ther. 2022;47(3):263–269. doi: 10.1111/jcpt.13500. [DOI] [PubMed] [Google Scholar]
- 82.Zhang Z-J, Yuan J, Bi Y, Wang C, Liu Y. The effect of metformin on biomarkers and survivals for breast cancer-a systematic review and meta-analysis of randomized clinical trials. Pharmacol Res. 2019;141:551–555. doi: 10.1016/j.phrs.2019.01.036. [DOI] [PubMed] [Google Scholar]
- 83.Farkhondeh T, Amirabadizadeh A, Aramjoo H, Llorens S, Roshanravan B, Saeedi F, et al. Impact of metformin on cancer biomarkers in non-diabetic cancer patients: a systematic review and meta-analysis of clinical trials. Curr Oncol. 2021;28(2):1412–1423. doi: 10.3390/curroncol28020134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Rahmani J, Manzari N, Thompson J, Gudi S, Chhabra M, Naik G, et al. The effect of metformin on biomarkers associated with breast cancer outcomes: a systematic review, meta-analysis, and dose–response of randomized clinical trials. Clin Transl Oncol. 2020;22(1):37–49. doi: 10.1007/s12094-019-02108-9. [DOI] [PubMed] [Google Scholar]
- 85.Barakat HE, Hussein RR, Elberry AA, Zaki MA, Elsherbiny RM. Factors influencing the anticancer effects of metformin on breast cancer outcomes: a systematic review and meta-analysis. Expert Rev Anticancer Ther. 2022;22(4):415–436. doi: 10.1080/14737140.2022.2051482. [DOI] [PubMed] [Google Scholar]
- 86.Park Y-M, Bookwalter D, O'Brien K, Jackson C, Weinberg C, Sandler D. A prospective study of type 2 diabetes, metformin use, and risk of breast cancer. Ann Oncol. 2021;32(3):351–359. doi: 10.1016/j.annonc.2020.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.García-Esquinas E, Guinó E, Castano-Vinyals G, Pérez-Gómez B, Llorca J, Altzibar JM, et al. Association of diabetes and diabetes treatment with incidence of breast cancer. Acta Diabetol. 2016;53(1):99–107. doi: 10.1007/s00592-015-0756-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Goodwin PJ, Chen BE, Gelmon KA, Whelan TJ, Ennis M, Lemieux J, et al. Effect of metformin vs placebo on invasive disease-free survival in patients with breast cancer: the MA. 32 randomized clinical trial. JAMA. 2022;327(20):1963–1973. doi: 10.1001/jama.2022.6147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Jarroux J, Morillon A, Pinskaya M. History, discovery, and classification of lncRNAs. Adv Exp Med Biol. 2017;1008:1–46. doi: 10.1007/978-981-10-5203-3_1. [DOI] [PubMed] [Google Scholar]
- 90.Marciniak M, iEwolucjonizmu ZG. Imprinting genomowy u ssaków: Najnowsze doniesienia. Post Biol Kom. 2008;35:243–252. [Google Scholar]
- 91.Johnsson P, Lipovich L, Grandér D, Morris KV. Evolutionary conservation of long non-coding RNAs; sequence, structure, function. Biochim Biophys Acta. 2014;1840(3):1063–1071. doi: 10.1016/j.bbagen.2013.10.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kumar V, Westra H-J, Karjalainen J, Zhernakova DV, Esko T, Hrdlickova B, et al. Human disease-associated genetic variation impacts large intergenic non-coding RNA expression. PLoS Genet. 2013;9(1):e1003201. doi: 10.1371/journal.pgen.1003201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Statello L, Guo C-J, Chen L-L, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021;22(2):96–118. doi: 10.1038/s41580-020-00315-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Fathi DB. Strategies to target long non-coding RNAs in cancer treatment: progress and challenges. Egypt J Med Hum Genet. 2020;21(1):41. doi: 10.1186/s43042-020-00074-4. [DOI] [Google Scholar]
- 95.Balas MM, Johnson AM. Exploring the mechanisms behind long noncoding RNAs and cancer. Non-coding RNA Res. 2018;3(3):108–117. doi: 10.1016/j.ncrna.2018.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Gao N, Li Y, Li J, Gao Z, Yang Z, Li Y, et al. Long non-coding RNAs: the regulatory mechanisms, research strategies, and future directions in cancers. Front Oncol. 2020;10:598817. doi: 10.3389/fonc.2020.598817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Sebastian-delaCruz M, Gonzalez-Moro I, Olazagoitia-Garmendia A, Castellanos-Rubio A, Santin I. The role of lncRNAs in gene expression regulation through mRNA stabilization. Noncoding RNA. 2021;7(1):3. doi: 10.3390/ncrna7010003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Giannakakis A, Zhang J, Jenjaroenpun P, Nama S, Zainolabidin N, Aau MY, et al. Contrasting expression patterns of coding and noncoding parts of the human genome upon oxidative stress. Sci Rep. 2015;5:9737. doi: 10.1038/srep09737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Begolli R, Sideris N, Giakountis A. LncRNAs as chromatin regulators in cancer: from molecular function to clinical potential. Cancers (Basel) 2019;11(10):1524. doi: 10.3390/cancers11101524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Wang W, Min L, Qiu X, Wu X, Liu C, Ma J, et al. Biological function of long non-coding RNA (LncRNA) Xist. Front Cell Dev Biol. 2021;9:645647. doi: 10.3389/fcell.2021.645647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Sang Y, Tang J, Li S, Li L, Tang X, Cheng C, et al. LncRNA PANDAR regulates the G1/S transition of breast cancer cells by suppressing p16(INK4A) expression. Sci Rep. 2016;6:22366. doi: 10.1038/srep22366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Cantile M, Di Bonito M, Cerrone M, Collina F, De Laurentiis M, Botti G. Long non-coding RNA HOTAIR in breast cancer therapy. Cancers (Basel) 2020;12(5):1197. doi: 10.3390/cancers12051197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Weidle UH, Birzele F, Kollmorgen G, Rueger R. Long non-coding RNAs and their role in metastasis. Cancer Genom Proteom. 2017;14(3):143–160. doi: 10.21873/cgp.20027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Dhamija S, Diederichs S. From junk to master regulators of invasion: lncRNA functions in migration, EMT and metastasis. Int J Cancer. 2016;139(2):269–280. doi: 10.1002/ijc.30039. [DOI] [PubMed] [Google Scholar]
- 105.Jiang M-C, Ni J-J, Cui W-Y, Wang B-Y, Zhuo W. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am J Cancer Res. 2019;9(7):1354. [PMC free article] [PubMed] [Google Scholar]
- 106.Jin H, Du W, Huang W, Yan J, Tang Q, Chen Y, et al. lncRNA and breast cancer: progress from identifying mechanisms to challenges and opportunities of clinical treatment. Molr Ther Nucleic Acids. 2021;25:613–637. doi: 10.1016/j.omtn.2021.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Sherif S, Mall R, Almeer H, Naik A, Al Homaid A, Thomas R, et al. Immune-related 3-lncRNA signature with prognostic connotation in a multi-cancer setting. J Transl Med. 2022;20(1):1–20. doi: 10.1186/s12967-022-03654-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Lai J, Chen B, Zhang G, Li X, Mok H, Liao N. Molecular characterization of breast cancer: a potential novel immune-related lncRNAs signature. J Transl Med. 2020;18(1):1–10. doi: 10.1186/s12967-020-02578-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Ma W, Zhao F, Yu X, Guan S, Suo H, Tao Z, et al. Immune-related lncRNAs as predictors of survival in breast cancer: a prognostic signature. J Transl Med. 2020;18(1):1–13. doi: 10.1186/s12967-020-02522-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Mehrpour Layeghi S, Arabpour M, Shakoori A, Naghizadeh MM, Mansoori Y, Tavakkoly Bazzaz J, et al. Expression profiles and functional prediction of long non-coding RNAs LINC01133, ZEB1-AS1 and ABHD11-AS1 in the luminal subtype of breast cancer. J Transl Med. 2021;19(1):1–17. doi: 10.1186/s12967-021-03026-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Li G-Y, Wang W, Sun J-Y, Xin B, Zhang X, Wang T, et al. Long non-coding RNAs AC026904. 1 and UCA1: a “one-two punch” for TGF-β-induced SNAI2 activation and epithelial–mesenchymal transition in breast cancer. Theranostics. 2018;8(10):2846. doi: 10.7150/thno.23463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Wu J, Xu W, Ma L, Sheng J, Ye M, Chen H, et al. Formononetin relieves the facilitating effect of lncRNA AFAP1-AS1-miR-195/miR-545 axis on progression and chemo-resistance of triple-negative breast cancer. Aging (Albany NY) 2021;13(14):18191. doi: 10.18632/aging.203156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Li Z, Hou P, Fan D, Dong M, Ma M, Li H, et al. The degradation of EZH2 mediated by lncRNA ANCR attenuated the invasion and metastasis of breast cancer. Cell Death Differ. 2017;24(1):59–71. doi: 10.1038/cdd.2016.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Tao W, Wang C, Zhu B, Zhang G, Pang D. 2019. LncRNA DANCR contributes to tumor progression via targetting miR-216a-5p in breast cancer: lncRNA DANCR contributes to tumor progression. Biosci Rep. [DOI] [PMC free article] [PubMed]
- 115.Yang F, Shen Y, Zhang W, Jin J, Huang D, Fang H, et al. An androgen receptor negatively induced long non-coding RNA ARNILA binding to miR-204 promotes the invasion and metastasis of triple-negative breast cancer. Cell Death Differ. 2018;25(12):2209–2220. doi: 10.1038/s41418-018-0123-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Li R-H, Chen M, Liu J, Shao C-C, Guo C-P, Wei X-L, et al. Long noncoding RNA ATB promotes the epithelial−mesenchymal transition by upregulating the miR-200c/Twist1 axe and predicts poor prognosis in breast cancer. Cell Death Dis. 2018;9(12):1–16. doi: 10.1038/s41419-018-1210-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Barton M, Santucci-Pereira J, Vaccaro OG, Nguyen T, Su Y, Russo J. BC200 overexpression contributes to luminal and triple negative breast cancer pathogenesis. BMC Cancer. 2019;19(1):1–17. doi: 10.1186/s12885-019-6179-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Xing Z, Lin A, Li C, Liang K, Wang S, Liu Y, et al. lncRNA directs cooperative epigenetic regulation downstream of chemokine signals. Cell. 2014;159(5):1110–1125. doi: 10.1016/j.cell.2014.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Lin X, Dinglin X, Cao S, Zheng S, Wu C, Chen W, et al. Enhancer-driven lncRNA BDNF-AS induces endocrine resistance and malignant progression of breast cancer through the RNH1/TRIM21/mTOR cascade. Cell Rep. 2020;31(10):107753. doi: 10.1016/j.celrep.2020.107753. [DOI] [PubMed] [Google Scholar]
- 120.Hu X, Liu Y, Du Y, Cheng T, Xia W. Long non-coding RNA BLACAT1 promotes breast cancer cell proliferation and metastasis by miR-150-5p/CCR2. Cell Biosci. 2019;9(1):1–9. doi: 10.1186/s13578-019-0274-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Gooding AJ, Zhang B, Jahanbani FK, Gilmore HL, Chang JC, Valadkhan S, et al. The lncRNA BORG drives breast cancer metastasis and disease recurrence. Sci Rep. 2017;7(1):1–18. doi: 10.1038/s41598-017-12716-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Zhang Y, Zhu M, Sun Y, Li W, Wang Y, Yu W. Upregulation of lncRNA CASC2 suppresses cell proliferation and metastasis of breast cancer via inactivation of the TGF-β signaling pathway. Oncol Res. 2019;27(3):379. doi: 10.3727/096504018X15199531937158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Han C, Li X, Fan Q, Liu G, Yin J. CCAT1 promotes triple-negative breast cancer progression by suppressing miR-218/ZFX signaling. Aging (Albany NY) 2019;11(14):4858. doi: 10.18632/aging.102080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Wu Z, Li Y, Wu Y, Wang Y, Nian W, Wang L, et al. Long non-coding RNA CCAT2 promotes the breast cancer growth and metastasis by regulating TGF-beta signaling pathway. Eur Rev Med Pharmacol Sci. 2017;21(4):706–714. [PubMed] [Google Scholar]
- 125.Wang Z, Yang B, Zhang M, Guo W, Wu Z, Wang Y, et al. lncRNA epigenetic landscape analysis identifies EPIC1 as an oncogenic lncRNA that interacts with MYC and promotes cell-cycle progression in cancer. Cancer Cell. 2018;33(4):706–720.e9. doi: 10.1016/j.ccell.2018.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Keshavarz M, Asadi MH. Long non-coding RNA ES 1 controls the proliferation of breast cancer cells by regulating the Oct4/Sox2/miR-302 axis. FEBS J. 2019;286(13):2611–2623. doi: 10.1111/febs.14825. [DOI] [PubMed] [Google Scholar]
- 127.Bai Y, Zhou X, Huang L, Wan Y, Li X, Wang Y. Long noncoding RNA EZR-AS1 promotes tumor growth and metastasis by modulating Wnt/β-catenin pathway in breast cancer. Exp Ther Med. 2018;16(3):2235–2242. doi: 10.3892/etm.2018.6461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Zhang Z, Sun L, Zhang Y, Lu G, Li Y, Wei Z. Long non-coding RNA FEZF1-AS1 promotes breast cancer stemness and tumorigenesis via targeting miR-30a/Nanog axis. J Cell Physiol. 2018;233(11):8630–8638. doi: 10.1002/jcp.26611. [DOI] [PubMed] [Google Scholar]
- 129.Ma F, Liu X, Zhou S, Li W, Liu C, Chadwick M, et al. Long non-coding RNA FGF13-AS1 inhibits glycolysis and stemness properties of breast cancer cells through FGF13-AS1/IGF2BPs/Myc feedback loop. Cancer Lett. 2019;450:63–75. doi: 10.1016/j.canlet.2019.02.008. [DOI] [PubMed] [Google Scholar]
- 130.Hu H, Wang Y, Zhang T, Zhang C, Liu Y, Li G, et al. Association of LncRNA-GACAT3 with MRI features of breast cancer and its molecular mechanism. J BUON. 2019;24:2377–2384. [PubMed] [Google Scholar]
- 131.Esmatabadi MJD, Motamedrad M, Sadeghizadeh M. Down-regulation of lncRNA, GAS5 decreases chemotherapeutic effect of dendrosomal curcumin (DNC) in breast cancer cells. Phytomedicine. 2018;42:56–65. doi: 10.1016/j.phymed.2018.03.022. [DOI] [PubMed] [Google Scholar]
- 132.Song R, Zhang J, Huang J, Hai T. Long non-coding RNA GHET1 promotes human breast cancer cell proliferation, invasion and migration via affecting epithelial mesenchymal transition. Cancer Biomark. 2018;22(3):565–573. doi: 10.3233/CBM-181250. [DOI] [PubMed] [Google Scholar]
- 133.Si H, Chen P, Li H, Wang X. Long non-coding RNA H19 regulates cell growth and metastasis via miR-138 in breast cancer. Am J Transl Res. 2019;11(5):3213. [PMC free article] [PubMed] [Google Scholar]
- 134.Wang Y, Zhang G, Han J. HIF1A-AS2 predicts poor prognosis and regulates cell migration and invasion in triple-negative breast cancer. J Cell Biochem. 2019;120(6):10513–10518. doi: 10.1002/jcb.28337. [DOI] [PubMed] [Google Scholar]
- 135.Lu PW, Li L, Wang F, Gu YT. Retracted: Effects of long non‐coding RNA HOST2 on cell migration and invasion by regulating MicroRNA let‐7b in breast cancer. Wiley Online Library; 2018. [DOI] [PubMed]
- 136.Mozdarani H, Ezzatizadeh V, Rahbar PR. The emerging role of the long non-coding RNA HOTAIR in breast cancer development and treatment. J Transl Med. 2020;18(1):1–15. doi: 10.1186/s12967-020-02320-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Liu M, Gou L, Xia J, Wan Q, Jiang Y, Sun S, et al. LncRNA ITGB2-AS1 could promote the migration and invasion of breast cancer cells through up-regulating ITGB2. Int J Mol Sci. 2018;19(7):1866. doi: 10.3390/ijms19071866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Zhou C, Wang X, Yang N, Xue S, Li W, Xie P. LncRNA LET function as a tumor suppressor in breast cancer development. Eur Rev Med Pharmacol Sci. 2018;22(18):6002–6007. doi: 10.26355/eurrev_201809_15935. [DOI] [PubMed] [Google Scholar]
- 139.Sas-Chen A, Aure MR, Leibovich L, Carvalho S, Enuka Y, Körner C, et al. LIMT is a novel metastasis inhibiting lnc RNA suppressed by EGF and downregulated in aggressive breast cancer. EMBO Mol Med. 2016;8(9):1052–1064. doi: 10.15252/emmm.201606198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Yuan C, Luo X, Duan S, Guo L. Long noncoding RNA LINC00115 promotes breast cancer metastasis by inhibiting miR-7. FEBS Open Bio. 2020;10(7):1230–1237. doi: 10.1002/2211-5463.12842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Wu J, Shuang Z, Zhao J, Tang H, Liu P, Zhang L, et al. Linc00152 promotes tumorigenesis by regulating DNMTs in triple-negative breast cancer. Biomed Pharmacother. 2018;97:1275–1281. doi: 10.1016/j.biopha.2017.11.055. [DOI] [PubMed] [Google Scholar]
- 142.Dong L, Qian J, Chen F, Fan Y, Long J. LINC00461 promotes cell migration and invasion in breast cancer through miR-30a-5p/integrin β3 axis. J Cell Biochem. 2019;120(4):4851–4862. doi: 10.1002/jcb.27435. [DOI] [PubMed] [Google Scholar]
- 143.Lu G, Li Y, Ma Y, Lu J, Chen Y, Jiang Q, et al. Long noncoding RNA LINC00511 contributes to breast cancer tumourigenesis and stemness by inducing the miR-185-3p/E2F1/Nanog axis. J Exp Clin Cancer Res. 2018;37(1):1–11. doi: 10.1186/s13046-018-0945-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Chen D, Zheng X, Cao Y, He X, Nian W, Zeng X, et al. Long non-coding RNA LINC00628 suppresses the growth and metastasis and promotes cell apoptosis in breast cancer. Eur Rev Med Pharmacol Sci. 2017;21(2):275–283. [PubMed] [Google Scholar]
- 145.Xia E, Shen Y, Bhandari A, Zhou X, Wang Y, Yang F, et al. Long non-coding RNA LINC00673 promotes breast cancer proliferation and metastasis through regulating B7–H6 and epithelial–mesenchymal transition. Am J Cancer Res. 2018;8(7):1273. [PMC free article] [PubMed] [Google Scholar]
- 146.Zhou W, Gong J, Chen Y, Chen J, Zhuang Q, Cao J, et al. Long noncoding RNA LINC00899 suppresses breast cancer progression by inhibiting miR-425. Aging (Albany NY) 2019;11(22):10144. doi: 10.18632/aging.102426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Song Z, Zhang X, Lin Y, Wei Y, Liang S, Dong C. LINC01133 inhibits breast cancer invasion and metastasis by negatively regulating SOX4 expression through EZH2. J Cell Mol Med. 2019;23(11):7554–7565. doi: 10.1111/jcmm.14625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Jiang M, Xiao Y, Liu D, Luo N, Gao Q, Guan Y. Overexpression of long noncoding RNA LINC01296 indicates an unfavorable prognosis and promotes tumorigenesis in breast cancer. Gene. 2018;675:217–224. doi: 10.1016/j.gene.2018.07.004. [DOI] [PubMed] [Google Scholar]
- 149.Li Y, Song Y, Wang Z, Zhang Z, Lu M, Wang Y. Long non-coding RNA LINC01787 drives breast cancer progression via disrupting miR-125b generation. Front Oncol. 2019;9:1140. doi: 10.3389/fonc.2019.01140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Xiong Y, Gu Y, Wang F, Li L, Zhu M, Wang N, et al. LINC01857 as an oncogene regulates CREB1 activation by interacting with CREBBP in breast cancer. J Cell Physiol. 2019;234(8):14031–14039. doi: 10.1002/jcp.28090. [DOI] [PubMed] [Google Scholar]
- 151.Jiang Z, Slater CM, Zhou Y, Devarajan K, Ruth KJ, Li Y, et al. LincIN, a novel NF90-binding long non-coding RNA, is overexpressed in advanced breast tumors and involved in metastasis. Breast Cancer Res. 2017;19(1):1–15. doi: 10.1186/s13058-017-0853-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Yan M, Zhang L, Li G, Xiao S, Dai J, Cen X. Long noncoding RNA linc-ITGB1 promotes cell migration and invasion in human breast cancer. Biotechnol Appl Biochem. 2017;64(1):5–13. doi: 10.1002/bab.1461. [DOI] [PubMed] [Google Scholar]
- 153.Hou P, Zhao Y, Li Z, Yao R, Ma M, Gao Y, et al. LincRNA-ROR induces epithelial-to-mesenchymal transition and contributes to breast cancer tumorigenesis and metastasis. Cell Death Dis. 2014;5(6):e1287. doi: 10.1038/cddis.2014.249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Ma T, Liang Y, Li Y, Song X, Zhang N, Li X, et al. LncRNA LINP1 confers tamoxifen resistance and negatively regulated by ER signaling in breast cancer. Cell Signal. 2020;68:109536. doi: 10.1016/j.cellsig.2020.109536. [DOI] [PubMed] [Google Scholar]
- 155.Huang X, Xie X, Liu P, Yang L, Chen B, Song C, et al. Adam12 and lnc015192 act as ceRNAs in breast cancer by regulating miR-34a. Oncogene. 2018;37(49):6316–6326. doi: 10.1038/s41388-018-0410-1. [DOI] [PubMed] [Google Scholar]
- 156.Luo L, Tang H, Ling L, Li N, Jia X, Zhang Z, et al. LINC01638 lncRNA activates MTDH-Twist1 signaling by preventing SPOP-mediated c-Myc degradation in triple-negative breast cancer. Oncogene. 2018;37(47):6166–6179. doi: 10.1038/s41388-018-0396-8. [DOI] [PubMed] [Google Scholar]
- 157.Wang S, Liang K, Hu Q, Li P, Song J, Yang Y, et al. JAK2-binding long noncoding RNA promotes breast cancer brain metastasis. J Clin Investig. 2017;127(12):4498–4515. doi: 10.1172/JCI91553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Beltrán-Anaya FO, Romero-Córdoba S, Rebollar-Vega R, Arrieta O, Bautista-Piña V, Dominguez-Reyes C, et al. Expression of long non-coding RNA ENSG 00000226738 (Lnc KLHDC 7B) is enriched in the immunomodulatory triple-negative breast cancer subtype and its alteration promotes cell migration, invasion, and resistance to cell death. Mol Oncol. 2019;13(4):909–927. doi: 10.1002/1878-0261.12446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Vennin C, Spruyt N, Robin Y-M, Chassat T, Le Bourhis X, Adriaenssens E. The long non-coding RNA 91H increases aggressive phenotype of breast cancer cells and up-regulates H19/IGF2 expression through epigenetic modifications. Cancer Lett. 2017;385:198–206. doi: 10.1016/j.canlet.2016.10.023. [DOI] [PubMed] [Google Scholar]
- 160.Richards EJ, Zhang G, Li Z-P, Permuth-Wey J, Challa S, Li Y, et al. Long non-coding RNAs (LncRNA) regulated by transforming growth factor (TGF) β: LncRNA-hit-mediated TGFβ-induced epithelial to mesenchymal transition in mammary epithelia. J Biol Chem. 2015;290(11):6857–6867. doi: 10.1074/jbc.M114.610915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Jia X, Shi L, Wang X, Luo L, Ling L, Yin J, et al. KLF5 regulated lncRNA RP1 promotes the growth and metastasis of breast cancer via repressing p27kip1 translation. Cell Death Dis. 2019;10(5):1–16. doi: 10.1038/s41419-019-1566-5. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 162.Shi S-J, Wang L-J, Yu B, Li Y-H, Jin Y, Bai X-Z. LncRNA-ATB promotes trastuzumab resistance and invasion-metastasis cascade in breast cancer. Oncotarget. 2015;6(13):11652. doi: 10.18632/oncotarget.3457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Zhou M, Hou Y, Yang G, Zhang H, Tu G, Du Y-E, et al. LncRNA-Hh strengthen cancer stem cells generation in twist-positive breast cancer via activation of hedgehog signaling pathway. Stem Cells. 2016;34(1):55–66. doi: 10.1002/stem.2219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Yi T, Zhou X, Sang K, Huang X, Zhou J, Ge L. Activation of lncRNA lnc-SLC4A1-1 induced by H3K27 acetylation promotes the development of breast cancer via activating CXCL8 and NF-kB pathway. Artif Cells Nanomed Biotechnol. 2019;47(1):3765–3773. doi: 10.1080/21691401.2019.1664559. [DOI] [PubMed] [Google Scholar]
- 165.Yang Y, Yang H, Xu M, Zhang H, Sun M, Mu P, et al. Long non-coding RNA (lncRNA) MAGI2-AS3 inhibits breast cancer cell growth by targeting the Fas/FasL signalling pathway. Hum Cell. 2018;31(3):232–241. doi: 10.1007/s13577-018-0206-1. [DOI] [PubMed] [Google Scholar]
- 166.Kwok ZH, Roche V, Chew XH, Fadieieva A, Tay Y. A non-canonical tumor suppressive role for the long non-coding RNA MALAT1 in colon and breast cancers. Int J Cancer. 2018;143(3):668–678. doi: 10.1002/ijc.31386. [DOI] [PubMed] [Google Scholar]
- 167.Arun G, Diermeier S, Akerman M, Chang K-C, Wilkinson JE, Hearn S, et al. Differentiation of mammary tumors and reduction in metastasis upon Malat1 lncRNA loss. Genes Dev. 2016;30(1):34–51. doi: 10.1101/gad.270959.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Cheng Y, Pan Y, Pan Y, Wang O. MNX1-AS1 is a functional oncogene that induces EMT and activates the AKT/mTOR pathway and MNX1 in breast cancer. Cancer Manag Res. 2019;11:803. doi: 10.2147/CMAR.S188007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Zhu M, Wang F, Mi H, Li L, Wang J, Han M, et al. Long noncoding RNA MEG3 suppresses cell proliferation, migration and invasion, induces apoptosis and paclitaxel-resistance via miR-4513/PBLD axis in breast cancer cells. Cell Cycle. 2020;19(23):3277–3288. doi: 10.1080/15384101.2020.1839700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Wang S, Ke H, Zhang H, Ma Y, Ao L, Zou L, et al. LncRNA MIR100HG promotes cell proliferation in triple-negative breast cancer through triplex formation with p27 loci. Cell Death Dis. 2018;9(8):1–11. doi: 10.1038/s41419-018-0869-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Zhu D, Zhang X, Lin Y, Liang S, Song Z, Dong C. MT1JP inhibits tumorigenesis and enhances cisplatin sensitivity of breast cancer cells through competitively binding to miR-24-3p. Am J Transl Res. 2019;11(1):245. [PMC free article] [PubMed] [Google Scholar]
- 172.Hu P, Chu J, Wu Y, Sun L, Lv X, Zhu Y, et al. NBAT1 suppresses breast cancer metastasis by regulating DKK1 via PRC2. Oncotarget. 2015;6(32):32410. doi: 10.18632/oncotarget.5609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Zhang M, Wu W, Wang Z, Wang X. lncRNA NEAT1 is closely related with progression of breast cancer via promoting proliferation and EMT. Eur Rev Med Pharmacol Sci. 2017;21(5):1020–1026. [PubMed] [Google Scholar]
- 174.Song X, Liu Z, Yu Z. LncRNA NEF is downregulated in triple negative breast cancer and correlated with poor prognosis. Acta Biochim Biophys Sin. 2019;51(4):386–392. doi: 10.1093/abbs/gmz021. [DOI] [PubMed] [Google Scholar]
- 175.Wu W, Chen F, Cui X, Yang L, Chen J, Zhao J, et al. LncRNA NKILA suppresses TGF-β-induced epithelial–mesenchymal transition by blocking NF-κB signaling in breast cancer. Int J Cancer. 2018;143(9):2213–2224. doi: 10.1002/ijc.31605. [DOI] [PubMed] [Google Scholar]
- 176.Jiang Y, Lin L, Zhong S, Cai Y, Zhang F, Wang X, et al. Overexpression of novel lncRNA NLIPMT inhibits metastasis by reducing phosphorylated glycogen synthase kinase 3β in breast cancer. J Cell Physiol. 2019;234(7):10698–10708. doi: 10.1002/jcp.27738. [DOI] [PubMed] [Google Scholar]
- 177.Li Y, Lv M, Song Z, Lou Z, Wang R, Zhuang M. Long non-coding RNA NNT-AS1 affects progression of breast cancer through miR-142-3p/ZEB1 axis. Biomed Pharmacother. 2018;103:939–946. doi: 10.1016/j.biopha.2018.04.087. [DOI] [PubMed] [Google Scholar]
- 178.Tan B-S, Yang M-C, Singh S, Chou Y-C, Chen H-Y, Wang M-Y, et al. LncRNA NORAD is repressed by the YAP pathway and suppresses lung and breast cancer metastasis by sequestering S100P. Oncogene. 2019;38(28):5612–5626. doi: 10.1038/s41388-019-0812-8. [DOI] [PubMed] [Google Scholar]
- 179.Yang Y-X, Wei L, Zhang Y-J, Hayano T, Pineiro Pereda MDP, Nakaoka H, et al. Long non-coding RNA p10247, high expressed in breast cancer (lncRNA-BCHE), is correlated with metastasis. Clin Exp Metastasis. 2018;35(3):109–121. doi: 10.1007/s10585-018-9901-2. [DOI] [PubMed] [Google Scholar]
- 180.Jadaliha M, Gholamalamdari O, Tang W, Zhang Y, Petracovici A, Hao Q, et al. A natural antisense lncRNA controls breast cancer progression by promoting tumor suppressor gene mRNA stability. PLoS Genet. 2018;14(11):e1007802. doi: 10.1371/journal.pgen.1007802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Liang Y, Song X, Li Y, Sang Y, Zhang N, Zhang H, et al. A novel long non-coding RNA-PRLB acts as a tumor promoter through regulating miR-4766-5p/SIRT1 axis in breast cancer. Cell Death Dis. 2018;9(5):1–16. doi: 10.1038/s41419-018-0582-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Gao X, Qin T, Mao J, Zhang J, Fan S, Lu Y, et al. PTENP1/miR-20a/PTEN axis contributes to breast cancer progression by regulating PTEN via PI3K/AKT pathway. J Exp Clin Cancer Res. 2019;38(1):1–14. doi: 10.1186/s13046-019-1260-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Tang J, Li Y, Sang Y, Yu B, Lv D, Zhang W, et al. LncRNA PVT1 regulates triple-negative breast cancer through KLF5/beta-catenin signaling. Oncogene. 2018;37(34):4723–4734. doi: 10.1038/s41388-018-0310-4. [DOI] [PubMed] [Google Scholar]
- 184.Liu B, Yao P, Xiao F, Guo J, Wu L, Yang Y. MYBL2-induced PITPNA-AS1 upregulates SIK2 to exert oncogenic function in triple-negative breast cancer through miR-520d-5p and DDX54. J Transl Med. 2021;19(1):1–17. doi: 10.1186/s12967-021-02956-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Wu H, Wang Y, Chen T, Li Y, Wang H, Zhang L, 2018. The N-terminal polypeptide derived from vMIP-II exerts its anti-tumor activity in human breast cancer by regulating lncRNA SPRY4-IT1. Biosci Rep. [DOI] [PMC free article] [PubMed]
- 186.Liu J, Song Z, Feng C, Lu Y, Zhou Y, Lin Y, et al. The long non-coding RNA SUMO1P3 facilitates breast cancer progression by negatively regulating miR-320a. Am J Transl Res. 2017;9(12):5594. [PMC free article] [PubMed] [Google Scholar]
- 187.Zhou B, Guo H, Tang J. Long non-coding RNA TFAP2A-AS1 inhibits cell proliferation and invasion in breast cancer via miR-933/SMAD2. Med Sci Monit Int Med J Exp Clin Res. 2019;25:1242. doi: 10.12659/MSM.912421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Liu Y, Du Y, Hu X, Zhao L, Xia W. Up-regulation of ceRNA TINCR by SP1 contributes to tumorigenesis in breast cancer. BMC Cancer. 2018;18(1):1–11. doi: 10.1186/s12885-018-4255-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Fan S, Yang Z, Ke Z, Huang K, Liu N, Fang X, et al. Downregulation of the long non-coding RNA TUG1 is associated with cell proliferation, migration, and invasion in breast cancer. Biomed Pharmacother. 2017;95:1636–1643. doi: 10.1016/j.biopha.2017.09.076. [DOI] [PubMed] [Google Scholar]
- 190.Li T, Liu Y, Xiao H, Xu G. Long non-coding RNA TUG1 promotes cell proliferation and metastasis in human breast cancer. Breast Cancer. 2017;24(4):535–543. doi: 10.1007/s12282-016-0736-x. [DOI] [PubMed] [Google Scholar]
- 191.Li H, Zhu L, Xu L, Qin K, Liu C, Yu Y, et al. Long noncoding RNA linc00617 exhibits oncogenic activity in breast cancer. Mol Carcinog. 2017;56(1):3–17. doi: 10.1002/mc.22338. [DOI] [PubMed] [Google Scholar]
- 192.Li X, Wu Y, Liu A, Tang X. Long non-coding RNA UCA1 enhances tamoxifen resistance in breast cancer cells through a miR-18a-HIF1α feedback regulatory loop. Tumor Biol. 2016;37(11):14733–14743. doi: 10.1007/s13277-016-5348-8. [DOI] [PubMed] [Google Scholar]
- 193.Zheng R, Lin S, Guan L, Yuan H, Liu K, Liu C, et al. Long non-coding RNA XIST inhibited breast cancer cell growth, migration, and invasion via miR-155/CDX1 axis. Biochem Biophys Res Commun. 2018;498(4):1002–1008. doi: 10.1016/j.bbrc.2018.03.104. [DOI] [PubMed] [Google Scholar]
- 194.Tariq A, Hao Q, Sun Q, Singh DK, Jadaliha M, Zhang Y, et al. LncRNA-mediated regulation of SOX9 expression in basal subtype breast cancer cells. RNA. 2020;26(2):175–185. doi: 10.1261/rna.073254.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Wang J, Xi C, Yang X, Lu X, Yu K, Zhang Y, et al. LncRNA WT1-AS inhibits triple-negative breast cancer cell migration and invasion by downregulating transforming growth factor β1. Cancer Biother Radiopharm. 2019;34(10):671–675. doi: 10.1089/cbr.2019.2925. [DOI] [PubMed] [Google Scholar]
- 196.Tian Y, Xia S, Ma M, Zuo Y. LINC00096 promotes the proliferation and invasion by sponging miR-383-5p and regulating RBM3 expression in triple-negative breast cancer. Onco Targets Ther. 2019;12:10569. doi: 10.2147/OTT.S229659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Li P, Zhou B, Lv Y, Qian Q. LncRNA HEIH regulates cell proliferation and apoptosis through miR-4458/SOCS1 axis in triple-negative breast cancer. Hum Cell. 2019;32(4):522–528. doi: 10.1007/s13577-019-00273-1. [DOI] [PubMed] [Google Scholar]
- 198.Mou E, Wang H. 2019. LncRNA LUCAT1 facilitates tumorigenesis and metastasis of triple-negative breast cancer through modulating miR-5702. Biosci Rep. [DOI] [PMC free article] [PubMed]
- 199.Wang Y, Wu S, Zhu X, Zhang L, Deng J, Li F, et al. LncRNA-encoded polypeptide ASRPS inhibits triple-negative breast cancer angiogenesis. J Exp Med. 2020 doi: 10.1084/jem.20190950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Wei M, Liu L, Wang Z. Long non-coding RNA heart and neural crest derivatives expressed 2-antisense RNA 1 overexpression inhibits the proliferation of cancer cells by reducing RUNX2 expression in triple-negative breast cancer. Oncol Lett. 2019;18(6):6775–6780. doi: 10.3892/ol.2019.11001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Wang G, Mou Z, Xu Y, Liu G, Wang D, Zhang H. LINC01096 knockdown inhibits progression of triple-negative breast cancer by increasing miR-3130-3p. Eur Rev Med Pharmacol Sci. 2019;23(17):7445–7456. doi: 10.26355/eurrev_201909_18854. [DOI] [PubMed] [Google Scholar]
- 202.Hung T, Wang Y, Lin MF, Koegel AK, Kotake Y, Grant GD, et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nat Genet. 2011;43(7):621–629. doi: 10.1038/ng.848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Zou Q, Zhou E, Xu F, Zhang D, Yi W, Yao J. A TP73-AS1/miR-200a/ZEB1 regulating loop promotes breast cancer cell invasion and migration. J Cell Biochem. 2018;119(2):2189–2199. doi: 10.1002/jcb.26380. [DOI] [PubMed] [Google Scholar]
- 204.Huan J, Xing L, Lin Q, Xui H, Qin X. Long noncoding RNA CRNDE activates Wnt/β-catenin signaling pathway through acting as a molecular sponge of microRNA-136 in human breast cancer. Am J Transl Res. 2017;9(4):1977. [PMC free article] [PubMed] [Google Scholar]
- 205.Wu J, Chen H, Ye M, Wang B, Zhang Y, Sheng J, et al. Downregulation of long noncoding RNA HCP5 contributes to cisplatin resistance in human triple-negative breast cancer via regulation of PTEN expression. Biomed Pharmacother. 2019;115:108869. doi: 10.1016/j.biopha.2019.108869. [DOI] [PubMed] [Google Scholar]
- 206.Shi Y, Lu H, Wang H. Downregulated lncRNA ADAMTS9-AS2 in breast cancer enhances tamoxifen resistance by activating microRNA-130a-5p. Eur Rev Med Pharmacol Sci. 2019;23(4):1563–1573. doi: 10.26355/eurrev_201902_17115. [DOI] [PubMed] [Google Scholar]
- 207.Mitobe Y, Ikeda K, Suzuki T, Takagi K, Kawabata H, Horie-Inoue K, et al. ESR1-stabilizing long noncoding RNA TMPO-AS1 promotes hormone-refractory breast cancer progression. Mol Cell Biol. 2019;39(23):e00261–e319. doi: 10.1128/MCB.00261-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Sun W, Li AQ, Zhou P, Jiang YZ, Jin X, Liu YR, et al. DSCAM-AS 1 regulates the G1/S cell cycle transition and is an independent prognostic factor of poor survival in luminal breast cancer patients treated with endocrine therapy. Cancer Med. 2018;7(12):6137–6146. doi: 10.1002/cam4.1603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Feng J, Wen T, Li Z, Feng L, Zhou L, Yang Z, et al. Cross-talk between the ER pathway and the lncRNA MAFG-AS1/miR-339-5p/CDK2 axis promotes progression of ER+ breast cancer and confers tamoxifen resistance. Aging (Albany NY) 2020;12(20):20658. doi: 10.18632/aging.103966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Shi Q, Li Y, Li S, Jin L, Lai H, Wu Y, et al. LncRNA DILA1 inhibits Cyclin D1 degradation and contributes to tamoxifen resistance in breast cancer. Nat Commun. 2020;11(1):1–15. doi: 10.1038/s41467-020-19349-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Du C, Wang Y, Zhang Y, Zhang J, Zhang L, Li J. LncRNA DLX6-AS1 contributes to epithelial–mesenchymal transition and cisplatin resistance in triple-negative breast cancer via modulating Mir-199b-5p/paxillin axis. Cell Transplant. 2020;29:0963689720929983. doi: 10.1177/0963689720929983. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 212.Zhang H, Zhang X-Y, Kang X-N, Jin L-J, Wang Z-Y. LncRNA-SNHG7 enhances chemotherapy resistance and cell viability of breast cancer cells by regulating miR-186. Cancer Manag Res. 2020;12:10163. doi: 10.2147/CMAR.S270328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Tang L, Chen Y, Chen H, Jiang P, Yan L, Mo D, et al. DCST1-AS1 promotes TGF-β-induced epithelial-mesenchymal transition and enhances chemoresistance in triple-negative breast cancer cells via ANXA1. Front Oncol. 2020;10:280. doi: 10.3389/fonc.2020.00280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Wang Z, Katsaros D, Biglia N, Shen Y, Loo L, Yu X, et al. ERα upregulates the expression of long non-coding RNA LINC00472 which suppresses the phosphorylation of NF-κB in breast cancer. Breast Cancer Res Treat. 2019;175(2):353–368. doi: 10.1007/s10549-018-05108-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Dong H, Wang W, Mo S, Chen R, Zou K, Han J, et al. SP1-induced lncRNA AGAP2-AS1 expression promotes chemoresistance of breast cancer by epigenetic regulation of MyD88. J Exp Clin Cancer Res. 2018;37(1):1–15. doi: 10.1186/s13046-018-0875-3. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 216.Dong H, Wang W, Mo S, Liu Q, Chen X, Chen R, et al. Long non-coding RNA SNHG14 induces trastuzumab resistance of breast cancer via regulating PABPC1 expression through H3K27 acetylation. J Cell Mol Med. 2018;22(10):4935–4947. doi: 10.1111/jcmm.13758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Pan Y, Pan Y, Cheng Y, Yang F, Yao Z, Wang O. Knockdown of LncRNA MAPT-AS1 inhibites proliferation and migration and sensitizes cancer cells to paclitaxel by regulating MAPT expression in ER-negative breast cancers. Cell Biosci. 2018;8(1):1–13. doi: 10.1186/s13578-018-0207-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Chang L, Hu Z, Zhou Z, Zhang H. Linc00518 contributes to multidrug resistance through regulating the MiR-199a/MRP1 axis in breast cancer. Cell Physiol Biochem. 2018;48(1):16–28. doi: 10.1159/000491659. [DOI] [PubMed] [Google Scholar]
- 219.Wang R, Zhang T, Yang Z, Jiang C, Seng J. Long non-coding RNA FTH 1P3 activates paclitaxel resistance in breast cancer through miR-206/ABCB 1. J Cell Mol Med. 2018;22(9):4068–4075. doi: 10.1111/jcmm.13679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Yang F, Liu Y-H, Dong S-Y, Ma R-M, Bhandari A, Zhang X-H, et al. A novel long non-coding RNA FGF14-AS2 is correlated with progression and prognosis in breast cancer. Biochem Biophys Res Commun. 2016;470(3):479–483. doi: 10.1016/j.bbrc.2016.01.147. [DOI] [PubMed] [Google Scholar]
- 221.Kong Y, Geng C, Dong Q. LncRNA PAPAS may promote triple-negative breast cancer by downregulating miR-34a. J Int Med Res. 2019;47(8):3709–3718. doi: 10.1177/0300060519850724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.García-Venzor A, Mandujano-Tinoco EA, Ruiz-Silvestre A, Sánchez JM, Lizarraga F, Zampedri C, et al. lncMat2B regulated by severe hypoxia induces cisplatin resistance by increasing DNA damage repair and tumor-initiating population in breast cancer cells. Carcinogenesis. 2020;41(11):1485–1497. doi: 10.1093/carcin/bgaa078. [DOI] [PubMed] [Google Scholar]
- 223.Sun W, Xu X, Jiang Y, Jin X, Zhou P, Liu Y, et al. Transcriptome analysis of luminal breast cancer reveals a role for LOL in tumor progression and tamoxifen resistance. Int J Cancer. 2019;145(3):842–856. doi: 10.1002/ijc.32185. [DOI] [PubMed] [Google Scholar]
- 224.Zeng Y, Wang G, Zhou C-F, Zhang H-B, Sun H, Zhang W, et al. LncRNA profile study reveals a three-LncRNA signature associated with the pathological complete response following neoadjuvant chemotherapy in breast cancer. Front Pharmacol. 2019;10:574. doi: 10.3389/fphar.2019.00574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Yao N, Fu Y, Chen L, Liu Z, He J, Zhu Y, et al. Long non-coding RNA NONHSAT101069 promotes epirubicin resistance, migration, and invasion of breast cancer cells through NONHSAT101069/miR-129-5p/Twist1 axis. Oncogene. 2019;38(47):7216–7233. doi: 10.1038/s41388-019-0904-5. [DOI] [PubMed] [Google Scholar]
- 226.Yu F, Wang L, Zhang B. Long non-coding RNA DRHC inhibits the proliferation of cancer cells in triple negative breast cancer by downregulating long non-coding RNA HOTAIR. Oncol Lett. 2019;18(4):3817–3822. doi: 10.3892/ol.2019.10683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Shaw RJ, Kosmatka M, Bardeesy N, Hurley RL, Witters LA, DePinho RA, et al. The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress. Proc Natl Acad Sci USA. 2004;101(10):3329–3335. doi: 10.1073/pnas.0308061100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Shackelford DB, Abt E, Gerken L, Vasquez DS, Seki A, Leblanc M, et al. LKB1 inactivation dictates therapeutic response of non-small cell lung cancer to the metabolism drug phenformin. Cancer Cell. 2013;23(2):143–158. doi: 10.1016/j.ccr.2012.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Jiang Y, Qian T, Li S, Xie Y, Tao M. Metformin reverses tamoxifen resistance through the lncRNA GAS5-medicated mTOR pathway in breast cancer. Ann Transl Med. 2022;10(6):366. doi: 10.21037/atm-22-795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Golshan M, Khaleghi S, Shafiee SM, Valaee S, Ghanei Z, Jamshidizad A, et al. Metformin modulates oncogenic expression of HOTAIR gene via promoter methylation and reverses epithelial–mesenchymal transition in MDA-MB-231 cells. J Cell Biochem. 2021;122(3–4):385–393. doi: 10.1002/jcb.29867. [DOI] [PubMed] [Google Scholar]
- 231.Pai SG, Carneiro BA, Mota JM, Costa R, Leite CA, Barroso-Sousa R, et al. Wnt/beta-catenin pathway: modulating anticancer immune response. J Hematol Oncol. 2017;10(1):1–12. doi: 10.1186/s13045-017-0471-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Lu W, Kang Y. Epithelial–mesenchymal plasticity in cancer progression and metastasis. Dev Cell. 2019;49(3):361–374. doi: 10.1016/j.devcel.2019.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Guo Z, Zhao M, Howard EW, Zhao Q, Parris AB, Ma Z, et al. Phenformin inhibits growth and epithelial–mesenchymal transition of ErbB2-overexpressing breast cancer cells through targeting the IGF1R pathway. Oncotarget. 2017;8(36):60342. doi: 10.18632/oncotarget.19466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Chen J, Zheng G, Guo H, Shi Z-N, Jiang J, Wang X-Y, et al. The effect of metformin treatment on endoplasmic reticulum (ER) stress induced by status epilepticus (SE) via the PERK-eIF2α-CHOP pathway. Bosn J Basic Med Sci. 2018;18(1):49. doi: 10.17305/bjbms.2017.2044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Wang J, Sun J, Yang F. The role of long non-coding RNA H19 in breast cancer. Oncol Lett. 2020;19(1):7–16. doi: 10.3892/ol.2019.11093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Dahariya S, Paddibhatla I, Kumar S, Raghuwanshi S, Pallepati A, Gutti RK. Long non-coding RNA: classification, biogenesis and functions in blood cells. Mol Immunol. 2019;112:82–92. doi: 10.1016/j.molimm.2019.04.011. [DOI] [PubMed] [Google Scholar]
- 237.Keniry A, Oxley D, Monnier P, Kyba M, Dandolo L, Smits G, et al. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat Cell Biol. 2012;14(7):659–665. doi: 10.1038/ncb2521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Chen J, Qin C, Zhou Y, Chen Y, Mao M, Yang J. Metformin may induce ferroptosis by inhibiting autophagy via lncRNA H19 in breast cancer. FEBS Open Bio. 2022;12(1):146–153. doi: 10.1002/2211-5463.13314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Lin H-Y, Ho H-W, Chang Y-H, Wei C-J, Chu P-Y. The evolving role of ferroptosis in breast cancer: translational implications present and future. Cancers. 2021;13(18):4576. doi: 10.3390/cancers13184576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Wang D, Tang L, Zhang Y, Ge G, Jiang X, Mo Y, et al. Regulatory pathways and drugs associated with ferroptosis in tumors. Cell Death Dis. 2022;13(6):1–15. doi: 10.1038/s41419-022-04927-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Guo J, Li Y, Duan H, Yuan L. Metformin suppresses the proliferation and promotes the apoptosis of colon cancer cells through inhibiting the expression of long noncoding RNA-UCA1. Onco Targets Ther. 2020;13:4169. doi: 10.2147/OTT.S245091. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 242.Yan L, Zhou J, Gao Y, Ghazal S, Lu L, Bellone S, et al. Regulation of tumor cell migration and invasion by the H19/let-7 axis is antagonized by metformin-induced DNA methylation. Oncogene. 2015;34(23):3076–3084. doi: 10.1038/onc.2014.236. [DOI] [PubMed] [Google Scholar]
- 243.Aminimoghaddam S, Fooladi B, Noori M, Klashami ZN, Hamidi AK, Amoli MM. The effect of metformin on expression of long non-coding RNA H19 in endometrial cancer. Med J Islam Repub Iran. 2021;35:155. doi: 10.47176/mjiri.35.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Xia C, Liang S, He Z, Zhu X, Chen R, Chen J. Metformin, a first-line drug for type 2 diabetes mellitus, disrupts the MALAT1/miR-142-3p sponge to decrease invasion and migration in cervical cancer cells. Eur J Pharmacol. 2018;830:59–67. doi: 10.1016/j.ejphar.2018.04.027. [DOI] [PubMed] [Google Scholar]
- 245.Qiu C, Li C, Zheng Q, Fang S, Xu J, Wang H, et al. Metformin suppresses lung adenocarcinoma by downregulating long non-coding RNA (lncRNA) AFAP1-AS1 and secreted phosphoprotein 1 (SPP1) while upregulating miR-3163. Bioengineered. 2022;13(5):11987–12002. doi: 10.1080/21655979.2021.2005981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Wu P, Tang Y, Fang X, Xie C, Zeng J, Wang W, et al. Metformin suppresses hypopharyngeal cancer growth by epigenetically silencing long non-coding RNA SNHG7 in FaDu cells. Front Pharmacol. 2019;10:143. doi: 10.3389/fphar.2019.00143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Liu J, Cui J, Zhao S, Wu M, Wang J, Zhang Y, et al. Ferroptosis-related long noncoding RNAs have excellent predictive ability for multiomic characteristics of bladder cancer. Oxid Med Cell Longev. 2022 doi: 10.1155/2022/9316847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Tseng H-H, Chen Y-Z, Chou N-H, Chen Y-C, Wu C-C, Liu L-F, et al. Metformin inhibits gastric cancer cell proliferation by regulation of a novel Loc100506691-CHAC1 axis. Mol Ther Oncolytics. 2021;22:180–194. doi: 10.1016/j.omto.2021.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Sabry D, Abdelaleem OO, El Amin Ali AM, Mohammed RA, Abdel-Hameed ND, Hassouna A, et al. Anti-proliferative and anti-apoptotic potential effects of epigallocatechin-3-gallate and/or metformin on hepatocellular carcinoma cells: in vitro study. Mol Biol Rep. 2019;46(2):2039–2047. doi: 10.1007/s11033-019-04653-6. [DOI] [PubMed] [Google Scholar]
- 250.Jiang Z, Liu H. Metformin inhibits tumorigenesis in HBV-induced hepatocellular carcinoma by suppressing HULC overexpression caused by HBX. J Cell Biochem. 2018;119(6):4482–4495. doi: 10.1002/jcb.26555. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- Tao W, Wang C, Zhu B, Zhang G, Pang D. 2019. LncRNA DANCR contributes to tumor progression via targetting miR-216a-5p in breast cancer: lncRNA DANCR contributes to tumor progression. Biosci Rep. [DOI] [PMC free article] [PubMed]
- Wu H, Wang Y, Chen T, Li Y, Wang H, Zhang L, 2018. The N-terminal polypeptide derived from vMIP-II exerts its anti-tumor activity in human breast cancer by regulating lncRNA SPRY4-IT1. Biosci Rep. [DOI] [PMC free article] [PubMed]
- Mou E, Wang H. 2019. LncRNA LUCAT1 facilitates tumorigenesis and metastasis of triple-negative breast cancer through modulating miR-5702. Biosci Rep. [DOI] [PMC free article] [PubMed]
Supplementary Materials
Additional file 1: Table S1. lncRNAs associated with BC (continuation of Table 1).
Data Availability Statement
Not applicable.