

Abstract
Inflammatory Bowel Diseases (IBD) are chronic, reoccurring, and debilitating conditions characterized by inflammation in the gastrointestinal tract, some of which can lead to more systemic complications and can include autoimmune dysfunction, a change in the taxonomic and functional structure of microbial communities in the gut, and complicated burdens in a person’s daily life. Like many diseases based in chronic inflammation, research on IBD has pointed towards a multifactorial origin involving factors of the person’s lifestyle, immune system, associated microbial communities, and environmental conditions. Treatment currently exists only as palliative care, and seeks to disrupt the feedback loop of symptoms by reducing inflammation and allowing as much of a return to homeostasis as possible. Various anti-inflammatory options have been explored, and this review focuses on the use of diet as an alternative means of improving gut health. Specifically, we highlight the connection between the role of sulforaphane from cruciferous vegetables in regulating inflammation and in modifying microbial communities, and to break down the role they play in IBD.
Keywords: inflammatory bowel disease, Crohn’s Disease, ulcerative colitis, broccoli, broccoli sprouts, sulforaphane, glucoraphanin, gut microbiota, dietary bioactives
1. Introduction
Chronic inflammatory diseases involve interactions between a macroorganism host and its immune system, the microorganisms which associate with or encounter that host, and the environmental conditions of the host which include the collective exposures to chemical, microbial, or other stressors. Understanding these complex diseases and ways to circumvent them requires a holistic research framework [1,2]. Not only do inflammatory diseases contribute to dramatic morbidity and mortality, but they can create negative knock-on effects such as social or lifestyle restrictions in service to disease management [3,4], and an increased risk for a variety of cancers [5]. Inflammatory Bowel Diseases (IBD) are notable examples of chronic diseases which involve host-microbe-environment interactions in the gastrointestinal (GI) tract of humans that can significantly disrupt daily life. Prolonged inflammation has also been linked to a higher risk of colorectal cancer (CRC) development [6–9].
There are two major presentations of IBD (Fig. 1). Ulcerative colitis (UC) is primarily limited to the colon, where inflammation occurs along the mucosal epithelial layer [10]. Crohn’s Disease (CD) is an autoimmune disorder where inflammation can occur along the entire length of the GI tract [7], can be transmural, and can be associated with additional complications such as fistulas and strictures. Some patients with IBD show overlapping pathologic features of UC and CD [11]. Both UC and CD patients experience fluctuations between active periods of inflammation and periods of reduced disease activity. Symptoms include abdominal pain, diarrhea, bloody stool, weight loss, and fatigue. In the gut, both presentations of IBD can involve compromised gut epithelial barrier function and alterations to the gut microbial communities, discussed in detail further in this review.
Figure 1.
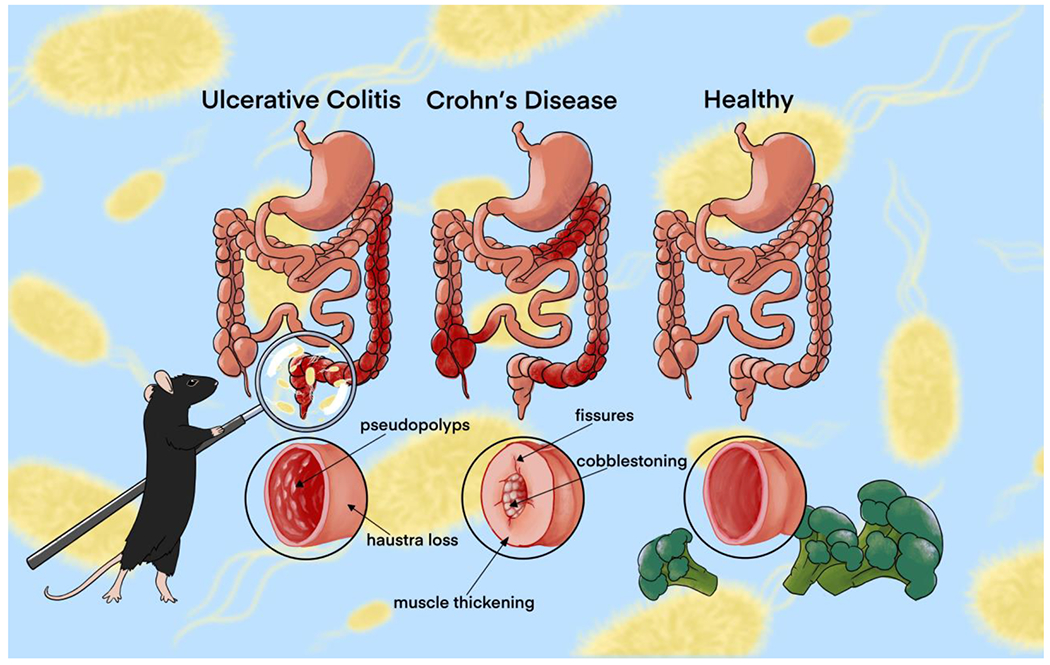
Primary Inflammatory Bowel Disease presentations, ulcerative colitis (UC) and Crohn’s disease (CD), compared to a healthy colon. UC is limited to the colon and is characterized by pseudopolyps and haustra loss (smoothing and shortening of the colon). CD can occur anywhere in the GI tract and is characterized by fissures, muscle thickening, and “cobblestoning”, which is the unique bubbling of the interior wall.
IBD has burgeoning global prevalence, with the greatest incidence rates in the United States, Canada, the United Kingdom, and western European countries and increasing incidence rates in nations which are assimilating those diets or industrialization of their food systems and sociopolitical or cultural structure [12,13]. The United States’ Centers for Disease Control and Prevention estimates 1.3% of U.S. adults have reported a diagnosis for IBD, and the risk for IBD development is highest in populations with lifestyle situations that are associated with increased stress and reduced access to resources, such as lower education rates, higher unemployment rates, or those living in poverty [14]. Diagnosis usually occurs between the ages of 20 and 40, and people diagnosed with IBD are more likely to have concurrent cardiovascular disease, cancer, arthritis, and kidney or liver disease, all of which typically have a chronic inflammation component. IBD poses a significant impact on the healthcare industry in the US. In 2015, health care expenditures for GI disorders totalled $135.9 billion, with IBD ranking in the top five most expensive, at $7.2 billion. From 2006 to 2014, the annual number of visits to the emergency department for IBD increased by 52%, and the number of hospitalizations from 2005 to 2014 increased by 13%. In addition, 13% of IBD cases are readmitted to the hospital within 30 days [15].
Palliative treatment for IBD reduces symptoms, and often targets inflammation as a way to recover host homeostasis and a functional microbial community in the gut, and achieve an overall reduction in side effects. However, current over-the-counter and prescription drugs for inflammation may cause ulcers or other symptoms [16]. The risk of developing colorectal cancer (CRC) increases with IBD duration [6–9]. Non-steroidal anti-inflammatory treatments (NSAIDs) and corticosteroids decrease risk of cancer incidence and mortality, and are widely available for the management of inflammatory diseases [17]. However, they have been associated with a variety of serious side effects, including stomach ulcers and a disordering of the microbial community structure in the gut (i.e. gut dysbiosis), and there is a growing need for non-steroidal treatment options that address chronic inflammation while preserving GI function and microbial communities.
Much is still unknown about IBD etiologies and development, however research has pointed towards a multifactorial origin. Environmental triggers for IBD include smoking, history of surgery, oral contraceptives, geography, socioeconomic status, infection, and diet [18,19], and host genetics and immune factors are also involved. Collectively, these factors are also known to alter host-associated microbial community structure and function, i.e. cause dysbiosis, which is a hallmark of IBD. Microbial-based therapeutics, such as probiotics or fecal microbial transplant (FMT) show promising results for resolving symptoms, but are often used as treatments rather than preventatives. However, diet is a crucial source of plant-derived bioactives which could provide non-steroidal anti-inflammatory compounds as a preventative strategy, and many of these metabolites have been demonstrated to drive host-microbial homeostasis and gut health [20]. This review explores a burgeoning research area: using dietary bioactives as an IBD intervention by directly providing anti-inflammatory compounds or indirectly providing the precursors which attract functional microorganisms to create anti-inflammatory compounds.
2. Current Status of Knowledge
2.1. Genetics and Immune Modulation of IBD
IBD has long been associated with genetic risk factors. There is increased prevalence in relatives of affected individuals as well as higher rates for disease among monozygotic versus dizygotic twins [21]. With the sequencing of the human genome, genome-wide association studies (GWAS) have elucidated over 200 gene loci associated with IBD [22,23]. Many of these genes play key roles in the proper functioning of the immune system, and their altered expression impacts important functions such as autophagy (ATG16L1, IRGM); mucosal barrier function and pathogen recognition and clearance (NOD2); and T cell response (IL10, IL21) [24]. Defects stemming from genetic alteration can result in impairment of proper immune cell function, leading to aberrant immune responses such as release of pro-inflammatory factors and dampened anti-inflammatory responses, further discussed below.
Dysregulated inflammation of the GI tract is a hallmark feature of IBD pathogenesis. Under normal conditions, inflammation is part of the innate immune response to harmful stimuli including microbial infiltration of tissues and tissue damage. Inflammation acts to provide a protective mucosal barrier between luminal microorganisms and the host epithelium [25,26], in an effort to neutralize the irritating stimulus and restore homeostasis. The dysregulated inflammatory and immune response to damage and/or pathogens leads to the chronicity of inflammation seen in IBD patients.
Acute inflammation can be initiated by the host upon recognition of pathogen-associated molecular patterns (PAMPs) on microbial cells, and damage-associated molecular patterns (DAMPs) released from damaged host cells [27], by pattern recognition receptors (PRRs) [28]. PRRs, which include the family of Toll-like receptors (TLRs), are upregulated in the intestinal mucosa of IBD patients, and activate signaling cascades that lead to the recruitment of immune cells in gut tissues [29]. Thus, IBD is marked by accumulations of CD4 and CD8 T lymphocytes at the disease site in attempts to ward off infections. Upon arrival, CD4 and CD8 may differentiate into T-helper cell types Th1 or Th2 [7]. Th1 and Th2 aggregation results in the increased production of proinflammatory cytokines, which coordinate the immune response. UC is reportedly driven by an aberrant Th2 response [30], while CD is linked to Th1 [31]. Acute inflammation becomes chronic when the target stimuli fail to be eliminated, and resolution pathways are dysregulated [5,25].
Pro-inflammatory interleukin (IL) cell signaling peptides IL-6 and IL-8 are upregulated and found in higher concentrations in the plasma and serum of patients with IBD and CRC, which has been implicated in the development and maintenance of these diseases [32–35]. Each cytokine is part of a complex signaling cascade, thus there are numerous targets that could produce the aforementioned effects. For example, IL-6 is part of the IL-6/JAK/STAT3 pathway, where STAT3 is a transcription factor for IL-6, and STAT3 inhibition results in IL-6 downregulation. Glycoprotein 130 (gp130) activates phosphorylation of STAT3 which results in dimerization, translocation, and IL-6 gene transcription [34–39]. IL-6 signaling and IL-6 trans signaling can result in TLR activation of pro-inflammatory signaling pathways, resulting in cancerous cell survival and evasion of detection by immune cells. Dietary bioactives such as sulforaphane, further discussed below, could act at multiple steps along the pathway to mitigate the inflammatory response.
IL-8 upregulation results in a pro-inflammatory response through increased neutrophil recruitment [25,32,40]. IL-8 transcription is dependent on the translocation of NF-κB p65 and p50 subunits across the nuclear membrane, and subsequent attachment at the IL-8 gene promoter [33]. Thus, inhibition of NF-κB results in downregulation of IL-8. The NF-κB subunits undergo nuclear translocation when TLR4 is activated at the cell surface and interacts with myeloid differentiation factor 88 (MyD88) [41,42], resulting in Bcl10 phosphorylation and combination with mucosa associated lymphoma translocation protein 1 (MALT1) [43]. The Bcl10/MALT1 complex ubiquitinates the IκB kinase-ɣ (IKKɣ), which phosphorylates IκB, causing dissociation from the p65 and p50 subunits and subsequent translocation. Due to the complexity of the signaling cascades involved in the production of ILs and furthered by their presence, there are many possible targets for intervention when contemplating reducing inflammation. Relevant targets for reduction include pro-inflammatory cytokines, such as IL-6, IL-12, IL-23 and IL-21, and targets for stimulation include anti-inflammatory cytokines, such as IL-10 and transforming growth factor-β (Fig. 2).
Figure 2.
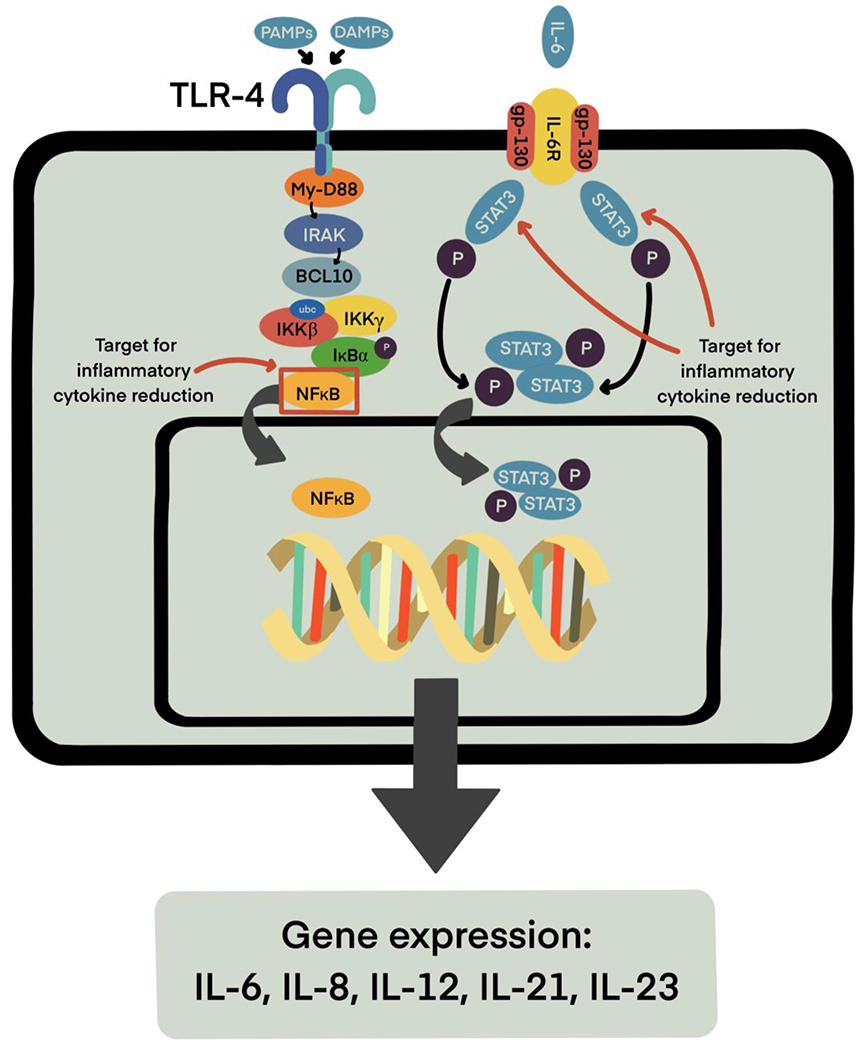
Production and activity of inflammatory cytokines IL-6, IL-8, IL-12, IL-21, and IL-23 rely on NF-κB and STAT3 signaling pathways. Targeting NF-κB and STAT3 may therefore result in reduction of these inflammatory molecules.
With over 200 identified risk loci [22,23], the genetics at play in IBD are complicated, to say the least. GWAS studies have indicated that UC and CD share 110/163 loci, reflecting the clinical similarity seen in these disorders, but these studies have also found loci distinct to each pathology as well as some detrimental in CD but protective in UC [22,44]. Moreover, more than 50% of the loci associated with IBD are also associated with other immune-mediated diseases, namely ankylosing spondylitis, psoriasis, and primary sclerosing cholangitis, indicating potential genetic links between inflammatory diseases previously considered distinct [44]. However, it is clear that a person’s genes are not the only factor in determining their risk for IBD, as the identified susceptibility genes account for less than 25% of predicted heritability [24]. Also of note, most GWAS studies have been conducted in broadly-defined Caucasian or Asian populations, and the same genes have not been implicated in GWAS studies in other populations [45]. Therefore, it is paramount to examine the other factors at play in the gut to expand our understanding of IBD pathogenesis.
2.2. Gut Microbiota, Biogeography, and IBD
In addition to inflammation and other innate immune responses (e.g. mucus production), invasion of pathogenic bacteria into body tissues is prevented by commensal bacteria [46]. The taxonomic and functional structure of microbial communities found along the GI tract are highly dependent on the diet, health status, age, and microbial encounters of the host [47–49]. Specific organs and sites within organs foster different environmental conditions that can create spatial niches for microbial taxa, an ecological occurrence known as biogeography [50–52]. For example, pH changes dramatically along the GI tract and is linked to microbial diversity and density. The upper intestine is more acidic and has a higher oxygen concentration than the colon, resulting in a microbial population of primarily fast growing, facultative anaerobes [53,54]. Here, the microbial community is curated by bile salts from the stomach which are bactericidal and stimulate the growth of Firmicutes phyla over Bacteroidetes [55]. Primary families of bacteria found in the upper intestine include Lactobacillaceae and Enterobacteriaceae. In contrast to the upper intestine, the colon has slower transit times [56] and lower concentrations of bile salts [55], and therefore has greater microbial abundance and diversity [53]. Primary families found in the colon include Bacteroidaceae, Prevotellaceae, Rikenellaceae, Lachnospiraceae, and Ruminococcaceae [57].
Initial colonization of the human GI tract occurs at birth via exposures to environmental sources of microorganisms. However, the most functional exposures have been linked to vaginal birth, breast feeding, and other sources of human-associated microbiota [58–60]. This first population of the gut is crucial in developing and maintaining epithelial barrier function [61], for example by stimulating epithelial cell division and growth, mucus production, angiogenesis (which supports epithelial cells), and neurogenesis (which supports gut motility). Early-life GI infections and disease have not been associated with IBD risk later in life [62], however, disruptions in the process of host-association and microbially-induced immune tolerance have been associated with IBD risk [63].
Importantly, microbial community diversity in the GI is much reduced in animal and human models of IBD, suggesting that more understanding may be gained by assessing microbial makeup at the species or strain levels, as key ecological roles could be left open as certain taxa are absent from the gut [64].
IBD patients often have a less acidic GI tract [65] due to altered diet or treatments, and this may result in alterations to gut biogeography such as small intestinal bacterial overgrowth [66]. Additionally, patients with UC have reduced bicarbonate secretion [67,68] and inflammation was found to alter bicarbonate transport in the ileum potentially affecting nutrient availability to resident microbes [69]. Bacterial lactate production is increased during periods of severe colitis, resulting in reduced production of short chain fatty acids (SCFAs) [68,70]. Interestingly, increases in lactate production during periods of active colitis is accompanied by increases in fecal counts of Lactobacilli and Enterococcus, but reduced Lactobacilli in biopsy samples, potentially due to the alteration of the mucous layer [71]. The mucosal-associated bacterial fraction are altered in IBD patients as compared to healthy participants [72,73]. Reactive oxygen species (ROS) and reactive nitrogen species (RNS) are important to cell processes and as anti-pathogen molecules. However, increased numbers of ROS and RNS are reported in IBD patients, and pathogenesis of UC has been linked to ROS [74].
Patients with GI inflammation were found to have altered richness of microbial taxonomic profiles, however, literature on this phenomenon has inconsistent findings, likely due to a combination of small sample sizes, nuances in methods, relying on fecal microbiota as a proxy for other GI populations, and the situation-specific trends of the complex human gut microbiome. It is not always clear which taxa may be contributing to inflammation and which are altered simply through changing environmental conditions in an inflamed gut. Symptoms may also be related to the loss of species associated with health benefits, such as UC patients with decreased abundance of Roseburia hominis and Faecalibacterium prausnitzii [75]. Conversely, CD patients have increased abundance in Faecalibacterium prausnitzii [76], as well as in Ruminococcus gnavus, and decreased abundance in Dialister invisus and Bifidobacterium adolescentis [77].
In addition to these nuances, species-level competition in the gut can have life-stage-specific benefits or detriments. For example, exposure to microbial diversity [78] and even non-pathogenic strains of Escherichia coli early in life can prime the immune system [79], yet UC and CD patients have been reported to have an increased abundance of pathogenic strains of E. coli during disease flare-ups [75,80]. This shift may be triggered by low-fiber/high-fat diets which are known to shift host metabolites, as well as entice E. coli and other pathobionts to the gut, which further shifts host-microbial interactions, increases inflammation, and results in a slightly high oxygen content along the epithelial wall which could result in the creation of cell-damaging reactive oxygen species [81]. Similarly, infection with certain strains of Helicobacter pylori, best known for causing ulcerative infections in the stomach and intestines, is associated with a large reduction in IBD risk as its presence triggers more immune monitoring [82]. However, H. pylori is also found in patients with IBD, and is commonly used to trigger CD-like symptoms in immune-deficient mouse models.
Complicated species-level antagonism can remodel gut communities [83]. Bacteroides fragilis has been identified as a gut commensal and early-life colonizer [84], and exhibits competition against other Bacteroides species in the gut. However, if unseated as the dominant species, B. vulgatus will increase in abundance and become dominant, and its proteases and other activities can induce colitis in mouse models [85]. Bacteroides have a complicated relationship with human health, and differences in genome and phenome are associated with disease [86] and colorectal cancer risk [87]. FMT has been used to “reset” the gut microbiota in a number of GI conditions, by providing a diverse and functional community from a healthy donor to a patient. FMT can impact intestinal permeability, the production of short-chain fatty acids (especially butyrate) and secondary bile acids, compete against pathogenic strains, and more, and has been used successfully to treat UC and CD. A systematic review of controlled trials found that UC patients had only a ~20% FMT success rate, while CD patients experienced a ~60% FMT success rate [88]. This is not surprising, considering that UC often results from chemical or physical damage (e.g. high salt diets) and the change in the microbiota is a side effect, whereas microbiota are actively involved in CD.
2.3. Dietary Factors and IBD Risk
It is now evident that diet is playing a major role in the increasing prevalence of IBD. The highest incidence of IBD occurs in the United States and Europe, where the diet has increasingly shifted toward high quantities of animal protein and fat in combination with sugar, processed foods, and low quantities of plant-based foods - referred to as a Westernized diet. In addition, as countries have Westernized their diets, they have also seen an increase in the rates of IBD [24,46]. The link between diet and IBD risk is well established in mouse models, as well as in humans where it is also recognized that diet is often accompanied by other social/lifestyle or environment co-factors [46,48,89].
Diet is a crucial source of plant-derived bioactives which could provide a non-steroidal effect, and many of these play an important role in driving microbial homeostasis and gut health [20]. For example, dietary fibers are crucial to short-chain fatty acid production (e.g. butyrate) which can support intestinal cell health and function and increase gut motility, and high fiber diets have been found to improve fecal markers of dysbiosis in patients with UC [90]. Another commonly researched plant metabolite group is polyphenols, which have been found to decrease inflammation, improve microbial abundance and diversity, and reduce oxidative stress [91–93]. There is a growing need for treatment options that address chronic inflammation while preserving GI function and promoting commensal microbial communities. Studies have shown that Westernized diets are detrimental to the gut microbiota diversity, and can lead to a pre-diabetic state, and changes in mucosal integrity, similar to that seen in IBD [24,94]. Therefore, positive changes in diet, achieved by incorporating more plant-based foods, may be a promising route in the prevention and treatment of IBD.
Epidemiological studies have investigated the impact of high fat/high protein and plant-based diets on microbial abundance and diversity [95]. High fat diets such as the Westernized diets result in increases in Firmicutes and Proteobacteria in both humans and mice [96–99]. Predictably, high fat diets that are low in plant derived dietary fibers and carbohydrates result in less abundant SCFA producing bacteria such as Roseburia spp, Eubacterium rectale, and Ruminococcus bromii, and increases in bile tolerant bacteria such as Alistepes spp, and Bilophila spp [97,98]. Bacterial makeup changes rapidly in response to antibiotic treatment, smoking, lack of exercise, dietary changes, and dietary additives and contaminants such as pesticides and heavy metals [100]. Sudden and abnormal changes in the abundance and diversity of gut microbiota have been associated with symptoms such as bloating and stomach pain, and can result in an over active immune response to commensal bacteria [101].
2.4. Targeting IBD with Broccoli Sprout Bioactives
Cruciferous vegetables are known for being rich in bioactives including fibers, flavonoids, and other antioxidants [102,103]. Epidemiological studies have shown that diets containing cruciferous vegetables, such as broccoli and broccoli sprouts, have inflammation-reducing benefits, are associated with lower cancer risk, and have specifically been shown to be an effective treatment in IBS, UC, and CD [104,105]. High consumption of cruciferous vegetables has been associated with decreased serum levels of pro-inflammatory cytokines, including IL-1β, TNFα, and IL-6 [106,107], as well as the inhibition of NFκB [107,108]. More specifically, the glucosinolates (GSLs) that are uniquely abundant in cruciferous vegetables can be metabolized into a variety of compounds with dramatically different actions and activity levels. GSLs are nitrogen- and sulfur-containing compounds which give cruciferous vegetables their distinct odor, which acts as a detractant against generalist herbivorous insects but an attractant for specialist herbivorous insects [109]. GSLs and many of their byproducts have well-demonstrated anti-insect activities [109], yet are cited to have numerous health benefits in humans.
GSLs can be metabolized into isothiocyanates (ITCs), a group of dietary bioactive metabolites with anti-inflammatory and anti-cancer activities [106,107,110–113]. When in the presence of plant-derived myrosinase enzymes, GSLs are hydrolyzed into glucose and an unstable molecule, thiohydroxamate-O-sulphanate, which is immediately broken down into ITCs (Fig. 3A). Which ITC product is formed depends on the GSL side chain, the pH of the environment, and the specifier proteins present [114]. Myrosinase and GSLs are housed in separate compartments in the plant, and come into contact with each other when the plant is damaged by herbivory/mastication, crushing, or heating. Epithiospecifier protein (ESP) is also present in the plant cell cytoplasm along with myrosinase, and converts GSL to ITC-nitrile, an alternative conversion product which is biologically inactive in humans [115]. ESP conversion of GSLs is more efficient, and therefore rates of conversion to ITC-nitrile are higher in raw broccoli plants (Fig. 2A). ESP is inactivated by low heat, which allows primary myrosinase conversion in mildly heated broccoli [116]. Myrosinase is denatured in the presence of high temperatures, and cooking vegetables results in significantly lower concentrations of ITC in the vegetables as GSLs are left intact [117,118].
Figure 3.
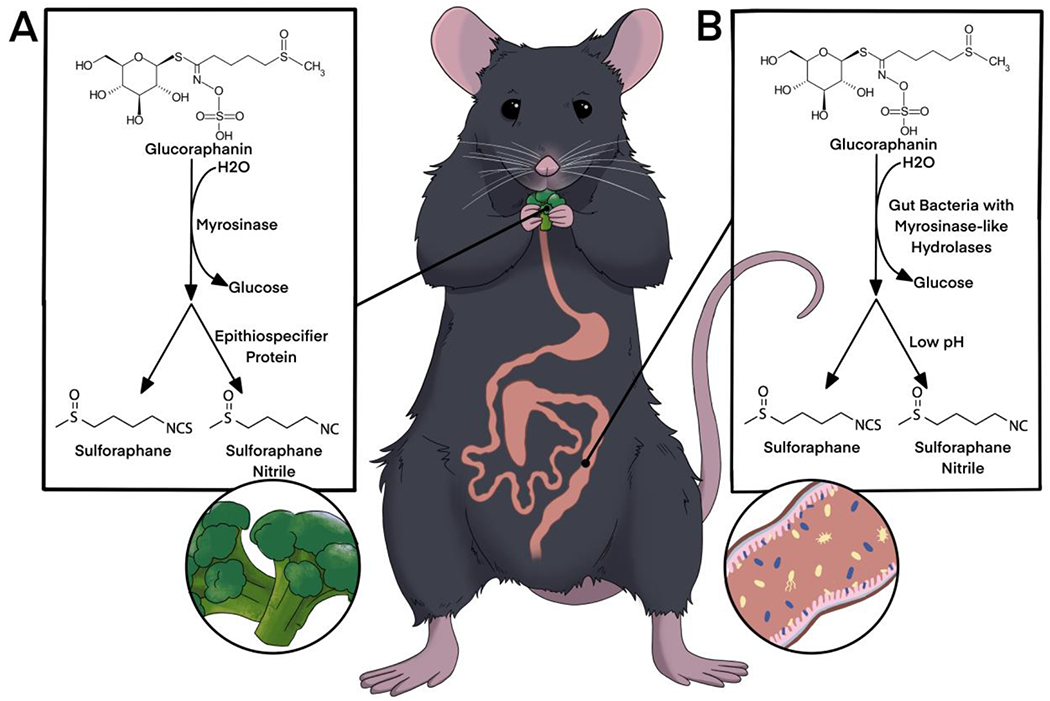
(A) Glucoraphanin hydrolysis in the presence of plant myrosinase upon damage to the broccoli plant. Epithiospecifier protein preferentially converts glucoraphanin to sulforaphane-nitrile. (B) Glucoraphanin hydrolysis has been demonstrated by gut bacteria in the ceceum and colon of mammals. Low pH environments favor conversion to sulforaphane-nitrile.
Of recent special interest, and one of the most well-studied ITCs, is sulforaphane (SFN) [119]. SFN is the primary hydrolysis product of glucoraphanin (GLR, also known as 4-methylsulfinylbutyl glucosinolate), which is the primary GSL found in mature broccoli, and at 15 times greater quantities in broccoli sprouts [109,120,121]. SFN has been shown to operate through inflammation pathways by inhibiting NF-κB, thereby downregulating pro-inflammatory cytokines important to IBD development and maintenance [122–125]. SFN inhibition of NF-κB translocation has been established, however the underlying mechanisms are yet to be elucidated. It has been theorized that SFN inhibits NF-κB by interacting with Keap1, which inhibits polyubiquitination and binding of Nrf2 and thereby allows Nrf2 nuclear accumulation [122]. Nrf2 activity is inversely linked to NF-κB activity, perhaps due to competition for CREB binding protein (CBP) [123,124]. Additionally, it is possible that SFN plays a direct role in inactivation of NF-κB by interacting with cysteine residues crucial to NF-κB activation [125]. Securing of these thiol groups leads to impairment of redox-sensitive DNA binding, ultimately preventing NF-κB from gene attachment. Mechanisms for STAT3 phosphorylation inhibition by SFN are also poorly understood. However, proposed mechanisms include inhibition of STAT3-dependent luciferase reporter activity, and modification of miR-124-3p expression [89,112].
Recent studies suggest the interaction between cruciferous vegetables, primarily broccoli and broccoli sprouts, and gut microbiota could contribute to stronger protective effects against GI inflammatory diseases and/or inflammation-associated tumorigenesis. Some studies have proposed that SFN is metabolized by GI bacteria [126,127], while SFN has also been shown to affect the composition of the gut microbiota, provide protective effects against dysbiosis, improve barrier function, and reduce the severity of IBD-like colitis in mouse models as discussed in detail further along in this review. As the gut microbiota contribute to the generation of broccoli sprout-derived bioactives [126,128–131], alterations in the gut microbiota may have significant impact on both the bioavailability and efficacy of broccoli sprout-derived bioactives (Fig. 3B).
2.5. Gut Microbiota-Mediated Metabolism of Glucosinolates
Mammals, including rats, mice and humans, do not produce digestive enzymes analogous to myrosinase for converting GSLs to ITCs [126,128,132,133]. Animal studies have been used to further investigate this conversion. In rats, SFN metabolites appeared in the urine after oral administration of purified GLR [134]. Consistently, one of our previous studies showed that mice fed a diet of steamed broccoli sprouts, which contained the highest level of GLR and minimal amount of SFN as compared to other preparations (raw or mild-heat-treated), have plasma and tissue SFN levels similar to mice fed steamed broccoli sprouts with active myrosinase added back in prior to consumption [131]. Germ-free rats failed to hydrolyze GSLs from Brussels sprouts diet [132]. Collectively, these data indicate a second conversion method occurring in the GI tract, where cecal and colonic microbiota have been found to have myrosinase-like hydrolases, and which have been demonstrated to convert GLR to SFN both in vivo and in vitro [121,126].
Animal studies also suggest that intact GSLs can reach the colon [129] and that the cecum/colon is the site of GSL conversion to the bioactive ITCs [126,129]. There is direct evidence of GLR hydrolysis in the rat cecal lumen in situ and absorption of the resulting SFN across the cecal enterocytes [126]. ITCs were detected in the mesenteric plasma 2 hours after the injection of GLR to the rat cecum, as compared to 15 minutes after the direct injection of SFN, indicating a conversion process of GLR to SFN in cecum [126]. The ex vivo incubation in the study supported the role of cecal microbiota in GLR hydrolysis as well, though ITC nitriles were the main hydrolytic products at the low pH environment [126].
There is a lot of indirect evidence to support the important role of human gut microbiota in metabolizing GSLs, however, not much clarity has been achieved for specific microorganisms responsible for converting GSLs to ITCs. In humans, urinary ITC excretion after consumption of cooked broccoli decreased dramatically in participants who were pretreated with oral antibiotics and bowel cleansing [128], supporting an important role for the gut microbiota in the metabolism of GSLs to its bioactive forms when plant-derived myrosinase is inactivated by cooking. Despite the inactivation of myrosinase in cooked broccoli, studies have found low amounts of SFN and other GLR hydrolysis products in the plasma and urine of people who have consumed a cooked broccoli meal [135,136]. One crossover study found a delayed appearance of ITCs in the plasma and urine from people consuming the dietary supplement of broccoli sprouts that lack myrosinase [137]. In addition, population studies continue to show an ITC-related reduction in cancer risk in the US, where the majority of consumers prefer cooked broccoli [138], supporting the idea that this might be a physiologically-relevant process.
GSLs are preferentially converted to ITC-nitrile under acidic conditions [139]. Luang-In et al. found that B. adolescentis, B. pseudocatenulatum, and B. longum primarily produced ITC-nitriles [140–143] perhaps due to dominant activity of a sulfatase over a myrosinase-like enzyme [141,144]. Luang-In further investigated the presence of putative myrosinases in L. agilis R16 and E. coli VL8 (bacteria known to convert GSLs to ITCs) and were unable to identify sequences similar to plant derived myrosinase enzymes. Wu et al. assessed GSL hydrolysis in mice fed broccoli and sinigrin, and found that bacteria in the families Clostridiaceae, Lachnospiraceae, and Porphyromonadaceae were positively associated with myrosinase activity [145]. Beta-glucosidase activity has been correlated with the abundance of members of the Lachnospiraceae and Ruminococcaceae families that potentially have glycoside hydrolases [146,147]. Several microorganisms isolated from the mammalian gut [148] appear to have myrosinase-like glycoside hydrolases that can cleave the glycoside moiety from GSLs, and there is evidence for GLR hydrolysis to SFN by cecal microbiota both ex vivo and in vivo to SFN [149–151]. Some bacterial species isolated from the human gut appear to have myrosinase-like enzymes that cleave the glycoside moiety from GSLs when incubated with GSLs in vitro, such as Escherichia coli, Bacteroides thetaiotaomicron, Enterococcus faecalis, Enterococcus faecium, Lactobacillus spp., Peptostreptococcus spp., and Bifidobacterium spp. [129,141,149,150,152]. Lactobacillus plantarum and Lactococcus lactis were capable of metabolizing GLR and glucoerucin into SFN nitrile and erucin nitrile [153]. Lactobacillus agilis was shown to convert sinigrin to allyl ITC [149]. BT2159-BT2156 was recently identified as an important gene cluster in GSL conversion to ITCs by B. thetaiotaomicron [154]. More studies are needed to elucidate the specific bacteria and/or bacterial consortia that can metabolize dietary glucosinolates to generate health-beneficial bioactives in the context of human microbial community.
2.6. Modulation of Gut Microbiota by Sulforaphane/Broccoli/Broccoli Sprouts
Broccoli is a significant source of dietary fibers, which are known to modify the gut microbiome [155–157]. The specific roles fiber and ITCs play in the modification of gut microbiota is unclear, however, studies focusing specifically on purified SFN and modification of the gut microbiome have revealed some of these nuances. Additionally, improved lipid metabolism markers and reduced triacylglycerols were observed in mice fed a broccoli stem diet [158] and human cells treated with purified SFN [159]. Broccoli diets result in increases in the Bacteroides fraction of human gut microbiota, and interestingly BMI seems to have an impact on the modification of microbiota by broccoli diets [160].
Many studies have suggested the critical role of plant-based myrosinase in ensuring maximal bioavailability of ITCs, and the insufficiency of gut microbiota-based myrosinase as the primary method of ITC production [137]. A study by Clark et al. found that subjects taking a broccoli sprout supplement with no active myrosinase, thus conversion of GSLs to ITCs was primarily dependent on the subject’s gut microbiota, had SFN metabolite levels 5 times lower than those subjects consuming fresh broccoli sprouts [137]. Feeding rats with 10% freeze-dried broccoli for 1 or 2 weeks resulted in improved ability of cecal microbiota to hydrolyze GLR ex vivo [120]. Liou et al. explored growth inhibition by ITCs on specific bacterial strains and found that some species are inhibited by the presence of ITCs in growth media while others are not, suggesting that ITCs can have an impact on the abundance of specific bacteria [154]. These studies suggest changes in gut microbiota with exposure to GLR or broccoli feeding that promote SFN formation, and the seemingly conflicting results may reflect the different behavior of isolated or mixed cultures in a laboratory setting and the complex environments of the gut.
Specifically, a number of studies have assessed the impacts of SFN and SFN-enriched broccoli diets on the abundance and diversity of the gut microbiota [160]. Both broccoli and SFN diets have been associated with protection against symptoms of colitis, yet result in only minor changes to gut microbiota in humans [160], although this may reflect the use of 3% genetic distance cutoffs to assign operational taxonomic units to represent a bacterial species rather than using phylotypes or sequence variants which are more precise. It is also highly likely that the protective effects from an SFN-enriched diet affect microbial activity more than community structure. Mice given SFN have increased Bacteroidetes to Firmicutes ratios [145,161,162], with reported increases in abundance and richness of Proteobacteria, Bacteroides, Clostridium spp. and butyrate-producing bacteria [163]. Furthermore, mice fed raw broccoli experienced increases in Bacteroidetes and decreases in Proteobacteria and Desulfovibrionaceae [145], and many pathogens identified in IBD patients do belong to this phylum or family, respectively [164,165]. SFN and SFN-enriched broccoli have also been associated with decreases in Desulfovibrionaceae [93] and Mucispirillum schaedleri [163], which are associated with pro-inflammatory activity in the GI tract.
Removing potentially pathogenic species through dietary components, or increasing overall bacterial diversity and promoting competitive exclusion of pathogens, would both result in a change to the host-microbial dynamics which would improve homeostasis in the gut. Recently, studies have investigated SFN as an antibacterial agent. SFN has a dose-dependent effect against both gram-negative and gram-positive bacteria. However, SFN is more effective against gram-negative bacteria such as E. coli and Salmonella enterica, than it is against gram-positive bacteria [166–168]. Additionally, SFN has been found to inactivate urease produced by Helicobacter pylori [169], and modify production of nitric oxide in human patients with H. pylori infection [170], potentially leading to an inability to colonize the GI tract by this bacteria.
Jun et al. assessed SFN-mediated microbial changes in an aging mouse model, and found that old mice fed an SFN diet experienced increases in microbial diversity, shifting the microbiota of old mice, and ultimately increasing the similarities between old and young mice [161]. They additionally noted an increase in Allobaculum, a bacteria associated with increased epithelial function. Zhang et al. assessed SFN modification of the gut microbiome in a DSS mouse model of UC. SFN significantly protected against community modification by DSS, and compared to DSS groups without SFN, SFN groups experienced decreases in the abundance of bacterial families Erysipelototrichaeae and Campylobacteraceae, both families associated with elevated intestinal inflammation or raised risk of IBD, as well as increases in Bacteroidales_S24-7 candidate family and Rikenellaceae, which have been associated with protection of intestinal health or against intestinal inflammation [162].
3. Conclusions
Dietary intervention is an equitable and in-demand strategy, as it creates a palliative care prevention and treatment strategy which is more affordable and accessible than a purified-supplement or medical treatment. While purified compounds represent important research and treatment options, a whole-food solution would ensure a wider adoption of this strategy and greater public health impact in the general community. Moreover, dietary interventions which are paired with other healthy-lifestyle improvements are shown to have a greater impact on reducing chronic diseases [171], and built environment infrastructure (e.g. grocery stores, public transportation) which reduces the burden of accessing healthy resources and thus improves fiber intake during dietary interventions [172]. At present, conflicting study results and the complicated nature of IBD, diet, and lifestyle have precluded the ability to make clear diet recommendations for all IBD patients. However, we can make recommendations based on harm-reduction, and diet needs to be part of a holistic treatment plan, to encourage patients to make dietary choices that promote commensal bacteria colonization and avoid choices which inadvertently exacerbate gut microbiome shifts.
It is becoming clear from the immense collective body of research on IBD that dietary bioactives and modification of the gut microbiota have an important and promising role in disease modification for individuals with IBD going forward. The data discussed in this review support the important role of microbial conversion of GSLs to ITCs. However, the structural and functional complexity of the gut microbiota make mechanistic studies challenging. Additionally, research comparing the differences between fiber and GSL modification of the gut microbiome is lacking, and therefore discussions regarding GSLs are missing critical aspects of a whole food approach to IBD intervention. Studies on identifying specific bacterial populations that are capable of metabolizing GSL precursors are inconclusive, partly due to the limitation of the assay methods, including the lack of appropriate in vitro and in vivo models to recapitulate the complex context of human microbial community. As gut microbiota is an extremely complicated ecosystem, better experimental methods need to be developed to accurately and efficiently study gut microbiota-mediated metabolism of GSLs. It is possible that the conversion of GSLs to ITCs is not achieved by one bacteria species, instead multiple players are involved in the process in a certain way. For example, an in vitro model has been used to mimic the human gut environment, in which SFN and SFN nitrile were detected in the culture medium that contained human gut microflora and purified GLR [173]. The use of organoid or microfluidics-based organ-on-a-chip combined with specific bacterial inoculation may provide real-time, high-throughput capacity, and studies using human microbiota-associated or humanized animal models may have the opportunity to evaluate spatial resolution of microbial populations in a more relevant context.
Another important consideration for human studies is that each individual’s gut microbiota is different, influenced by genetic and environmental factors such as diet and physical activity levels, as well as body weight and health conditions. The inter-individual variation will likely add extra complexity to the studies. Many human studies reported inter-individual variation in GSL metabolism. Notably, while there was less inter-individual variation in urinary ITC levels if ITCs were ingested, substantially different amounts of urinary ITCs were found if GSLs or cruciferous vegetables were given [128,136,150,174–177]. In a clinical trial, all subjects consumed 200 g steamed broccoli in one meal, resulting in highly varied levels of urinary ITC excretion, which was associated with varied levels of ex vivo GLR degradation by fecal bacterial culture from these subjects [150].
Biogeography has long been an ecological concept used to study the distribution of animals and plants with respect to geography, as it influences environmental conditions, food supply, dispersal, and more. Microbial ecology began using the term in the early 2000s to describe environmentally-located microbial communities [178], and as our research technology has improved, we have been able to identify microbial biogeography - or spatial and temporal distribution - in host-associated communities. Microbial biogeography is exquisitely clear in the gut as an ecological principle [50,51,179], and is critical to understanding how microbially-mediated dietary bioactives can be used to reduce inflammation and other complex diseases in the gut. Future research must take microbial biogeography into account when assessing the action of diet in this and other host-microbial-diet interactions. Combination and sequential therapy will be in part informed by what is learned from the microbial biogeography of the GI tract.
Highlights.
Modulation of gut microbial communities by broccoli/broccoli sprout bioactives.
Gut microbiota-mediated metabolism of broccoli/broccoli sprout-derived bioactives.
Importance of the interplay in reducing inflammation in inflammatory bowel diseases.
Statement of Significance:
This review highlights the importance of the interplay between broccoli sprout bioactives and gut microbiota and its important role in reducing inflammation in inflammatory bowel disease.
Acknowledgements
The authors would like to thank Brigitte Lavoie (University of Vermont) and Grace Chen (University of Michigan), as well as undergraduate researchers Dorien Baudewyns (Husson University), Louisa Colucci (Husson University), and Joe Balkan (Tufts University), for their work on the laboratory research which forms the broader collaboration that led to the generation of this review. All authors have read and approved the final manuscript. This project was supported by the USDA National Institute of Food and Agriculture, Hatch Project Numbers ME0-22102 (Ishaq) and ME0-22303 (Li) through the Maine Agricultural & Forest Experiment Station, and USDA-NIFA-AFRI Foundational Program [Grant No. 2018-67017-27520]. It was also supported by NIH grant NOA R21AT011203 (Mawe).
Maine Agricultural and Forest Experiment Station Publication Number [XXXX - will add upon acceptance]
Financial Support
This project was supported by the USDA-NIFA Hatch Project Numbers ME0-22102 (Ishaq) and ME0-22303 (Li) through the Maine Agricultural & Forest Experiment Station, and USDA-NIFA-AFRI Foundational Program [Grant No. 2018-67017-27520]. It was also supported by NIH grant NOA R21AT011203 (Mawe).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflict of Interest
The authors declare no conflicts of interest.
References
- [1].Wang J, Chen W-D, Wang Y-D. The Relationship Between Gut Microbiota and Inflammatory Diseases: The Role of Macrophages. Front Microbiol 2020;11:1065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Ahn J, Hayes RB. Environmental Influences on the Human Microbiome and Implications for Noncommunicable Disease. Annu Rev Public Health 2021;42:277–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Kim YS, Jung S-A, Lee K-M, Park SJ, Kim TO, Choi CH, et al. Impact of inflammatory bowel disease on daily life: an online survey by the Korean Association for the Study of Intestinal Diseases. Intest Res 2017;15:338–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Robertson N, Gunn S, Piper R. Psychological and Social Factors Associated with Pain in Inflammatory Bowel Disease: A Systematic Literature Review of the Evidence in Adult and Pediatric Studies. Crohn’s Colitis 360 2019;1:otz003. [Google Scholar]
- [5].Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature 2008;454:436–44. [DOI] [PubMed] [Google Scholar]
- [6].Klampfer L. Cytokines, inflammation and colon cancer. Current Cancer Drug Targets 2013;11:451–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Zhang Y-Z. Inflammatory bowel disease: Pathogenesis. World J Gastroenterol 2014;20:91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Matricon J, Barnich N, Ardid D. Immunopathogenesis of inflammatory bowel disease. Self Nonself 2010;1:299–309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Rubin DC, Shaker A, Levin MS. Chronic intestinal inflammation: Inflammatory bowel disease and colitis-associated colon cancer. Frontiers in Immunology 2012;3:00107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Qin X. Why is damage limited to the mucosa in ulcerative colitis but transmural in Crohn’s disease? World J Gastrointest Pathophysiol 2013;4:63–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Odze RD. A contemporary and critical appraisal of “indeterminate colitis.” Mod Pathol 2015;28:S30–46. [DOI] [PubMed] [Google Scholar]
- [12].Molodecky NA, Soon IS, Rabi DM, Ghali WA, Ferris M, Chernoff G, et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012;142:46–54.e42; quiz e30. [DOI] [PubMed] [Google Scholar]
- [13].Ng SC, Shi HY, Hamidi N, Underwood FE, Tang W, Benchimol EI, et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies. Lancet 2017;390:2769–78. [DOI] [PubMed] [Google Scholar]
- [14].CDC. Data and Statistics. US Centers for Disease Control and Prevention; 2021. https://www.cdc.gov/ibd/data-statistics.htm (accessed February 15, 2022). [Google Scholar]
- [15].Peery AF, Crockett SD, Murphy CC, Lund JL, Dellon ES, Williams JL, et al. Burden and Cost of Gastrointestinal, Liver, and Pancreatic Diseases in the United States: Update 2018. Gastroenterology 2019;156:254–72.e11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Jahnavi K, Reddy PP, Vasudha B, Narender B. Non-steroidal anti-inflammatory drugs: an overview. Journal of Drug Delivery and Therapeutics 2019;9:442–8. [Google Scholar]
- [17].Vonkeman HE, van de Laar MAFJ. Nonsteroidal anti-inflammatory drugs: adverse effects and their prevention. Semin Arthritis Rheum 2010;39:294–312. [DOI] [PubMed] [Google Scholar]
- [18].Loftus EV Jr. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 2004;126:1504–17. [DOI] [PubMed] [Google Scholar]
- [19].Vedamurthy A, Ananthakrishnan AN. Influence of Environmental Factors in the Development and Outcomes of Inflammatory Bowel Disease. Gastroenterol Hepatol 2019;2:72–82. [PMC free article] [PubMed] [Google Scholar]
- [20].Viennois E, Gewirtz AT, Chassaing B. Chronic Inflammatory Diseases: Are We Ready for Microbiota-based Dietary Intervention? Cell Mol Gastroenterol Hepatol 2019;8:61–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Halme L, Paavola-Sakki P, Turunen U, Lappalainen M, Farkkila M, Kontula K. Family and twin studies in inflammatory bowel disease. World J Gastroenterol 2006;12:3668–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Jostins L, Ripke S, Weersma RK, Duerr RH, McGovern DP, Hui KY, et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012;491:119–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Liu JZ, van Sommeren S, Huang H, Ng SC, Alberts R, Takahashi A, et al. Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nat Genet 2015;47:979–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Lee M, Chang EB. Inflammatory Bowel Diseases (IBD) and the Microbiome-Searching the Crime Scene for Clues. Gastroenterology 2021;160:524–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Chen L, Deng H, Cui H, Fang J, Zuo Z, Deng J, et al. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018;9:7204–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Ferrero-Miliani L, Nielsen OH, Andersen PS, Girardin SE. Chronic inflammation: importance of NOD2 and NALP3 in interleukin-1beta generation. Clin Exp Immunol 2007;147:227–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Roh JS, Sohn DH. Damage-Associated Molecular Patterns in Inflammatory Diseases. Immune Netw 2018;18:e27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Eom T, Kim YS, Choi CH, Sadowsky MJ, Unno T. Current understanding of microbiota- and dietary-therapies for treating inflammatory bowel disease. J Microbiol 2018;56:189–98. [DOI] [PubMed] [Google Scholar]
- [29].Tanaka K. Expression of Toll-like receptors in the intestinal mucosa of patients with inflammatory bowel disease. Expert Rev Gastroenterol Hepatol 2008;2:193–6. [DOI] [PubMed] [Google Scholar]
- [30].Targan SR, Karp LC. Defects in mucosal immunity leading to ulcerative colitis. Immunol Rev 2005;206:296–305. [DOI] [PubMed] [Google Scholar]
- [31].Cobrin GM, Abreu MT. Defects in mucosal immunity leading to Crohn’s disease. Immunol Rev 2005;206:277–95. [DOI] [PubMed] [Google Scholar]
- [32].Mazzucchelli L, Hauser C, Zgraggen K, Wagner H, Hess M, Laissue JA, et al. Expression of Interleukin-8 Gene in Inflammatory Bowel Disease is Related to the Histological Grade of Active Inflammation. American Journal of Pathology 1994;144:997–1007. [PMC free article] [PubMed] [Google Scholar]
- [33].Hoffmann E, Dittrich-Breiholz O, Holtmann H, Kracht M. Multiple control of interleukin-8 gene expression. Journal of Leukocyte Biology 2002;72:847–55. [PubMed] [Google Scholar]
- [34].Lin W-W, Karin M. A cytokine-mediated link between innate immunity, inflammation, and cancer. J Clin Invest 2007;117:1175–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Szkaradkiewicz A, Marciniak R, Chudzicka-Strugała I, Wasilewska A, Drews M, Majewski P, et al. Proinflammatory cytokines and IL-10 in inflammatory bowel disease and colorectal cancer patients. Arch Immunol Ther Exp 2009;57:291–4. [DOI] [PubMed] [Google Scholar]
- [36].Gabay C Interleukin-6 and chronic inflammation. Arthritis Res Ther 2006;8:S3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Neurath MF, Finotto S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine Growth Factor Rev 2011;22:83–9. [DOI] [PubMed] [Google Scholar]
- [38].Turner MD, Nedjai B, Hurst T, Pennington DJ. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochimica et Biophysica Acta - Molecular Cell Research 2014;1843:2563–82. [DOI] [PubMed] [Google Scholar]
- [39].Murakami M, Hirano T. A four-step model for the IL-6 amplifier, a regulator of chronic inflammations in tissue-specific MHC class ll-associated autoimmune diseases. Front Immunol 2011;2:00022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Jung HC, Eckmann L, Yang S-K, Panja A, Fierer J, Morzycka-Wroblewska E, et al. A Distinct Array of Proinflammatory Cytokines Is Expressed in Human Colon Epithelial Cells in Response to Bacterial Invasion. Journal of Clinical Investigations 1995;95:55–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Chilton PM, Embry CA, Mitchell TC. Effects of differences in lipid A structure on TLR4 pro-inflammatory signaling and inflammasome activation. Frontiers in Immunology 2012;3:00154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Lawrence T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harbor Perspectives in Biology 2009;1. 10.1101/cshperspect.a001651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Bhattacharyya S, Dudeja PK, Tobacman JK. ROS, Hsp27, and IKKbeta mediate dextran sodium sulfate (DSS) activation of IkappaBa, NFkappaB, and IL-8. Inflamm Bowel Dis 2009;15:673–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Ellinghaus D, Bethune J, Petersen B-S, Franke A. The genetics of Crohn’s disease and ulcerative colitis--status quo and beyond. Scand J Gastroenterol 2015;50:13–23. [DOI] [PubMed] [Google Scholar]
- [45].Alharbi RS, Shaik NA, Almahdi H, ElSokary HA, Jamalalail BA, Mosli MH, et al. Genetic association study of NOD2 and IL23R amino acid substitution polymorphisms in Saudi Inflammatory Bowel Disease patients. Journal of King Saud University - Science 2022;34:101726. [Google Scholar]
- [46].Levine A, Sigall Boneh R, Wine E. Evolving role of diet in the pathogenesis and treatment of inflammatory bowel diseases. Gut 2018;67:1726–38. [DOI] [PubMed] [Google Scholar]
- [47].Singh RK, Chang H-W, Yan D, Lee KM, Ucmak D, Wong K, et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med 2017;15:73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Johnson AJ, Vangay P, Al-Ghalith GA, Hillmann BM, Ward TL, Shields-Cutler RR, et al. Daily Sampling Reveals Personalized Diet-Microbiome Associations in Humans. Cell Host Microbe 2019;25:789–802.e5. [DOI] [PubMed] [Google Scholar]
- [49].Amato KR, Arrieta M-C, Azad MB, Bailey MT, Broussard JL, Bruggeling CE, et al. The human gut microbiome and health inequities. Proc Natl Acad Sci U S A 2021;118:e2017947118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Tropini C, Earle KA, Huang KC, Sonnenburg JL. The Gut Microbiome: Connecting Spatial Organization to Function. Cell Host Microbe 2017;21:433–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Hillman ET, Lu H, Yao T, Nakatsu CH. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ 2017;32:300–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Lynch SV, Pedersen O. The Human Intestinal Microbiome in Health and Disease. N Engl J Med 2016;375:2369–79. [DOI] [PubMed] [Google Scholar]
- [53].Donaldson GP, Lee SM, Mazmanian SK. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol 2016;14:20–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Nugent SG, Kumar D, Rampton DS, Evans DF. Intestinal luminal pH in inflammatory bowel disease: possible determinants and implications for therapy with aminosalicylates and other drugs. Gut 2001;48:571–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Ridlon JM, Kang DJ, Hylemon PB, Bajaj JS. Bile acids and the gut microbiome. Curr Opin Gastroenterol 2014;30:332–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Maurer AH. Gastrointestinal Motility, Part 2: Small-Bowel and Colon Transit. J Nucl Med Technol 2016;44:12–8. [DOI] [PubMed] [Google Scholar]
- [57].Gu S, Chen D, Zhang J-N, Lv X, Wang K, Duan L-P, et al. Bacterial community mapping of the mouse gastrointestinal tract. PLoS One 2013;8:e74957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Huurre A, Kalliomäki M, Rautava S, Rinne M, Salminen S, Isolauri E. Mode of delivery - effects on gut microbiota and humoral immunity. Neonatology 2008;93:236–40. [DOI] [PubMed] [Google Scholar]
- [59].Kelly D, King T, Aminov R. Importance of microbial colonization of the gut in early life to the development of immunity. Mutat Res 2007;622:58–69. [DOI] [PubMed] [Google Scholar]
- [60].Harmsen HJ, Wildeboer-Veloo AC, Raangs GC, Wagendorp AA, Klijn N, Bindels JG, et al. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J Pediatr Gastroenterol Nutr 2000;30:61–7. [DOI] [PubMed] [Google Scholar]
- [61].Guo F, Cai D, Li Y, Gu H, Qu H, Zong Q, et al. How Early-Life Gut Microbiota Alteration Sets Trajectories for Health and Inflammatory Bowel Disease? Frontiers in Nutrition 2021;8:690073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Bernstein CN, Burchill C, Targownik LE, Singh H, Roos LL. Events Within the First Year of Life, but Not the Neonatal Period, Affect Risk for Later Development of Inflammatory Bowel Diseases. Gastroenterology 2019;156:2190–7.e10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Wang M, Monaco MH, Donovan SM. Impact of early gut microbiota on immune and metabolic development and function. Semin Fetal Neonatal Med 2016;21:380–7. [DOI] [PubMed] [Google Scholar]
- [64].Gibbons SM. Keystone taxa indispensable for microbiome recovery. Nature Microbiology 2020;5:1067–8. [DOI] [PubMed] [Google Scholar]
- [65].Press AG, Hauptmann IA, Hauptmann L, Fuchs B, Fuchs M, Ewe K, et al. Gastrointestinal pH profiles in patients with inflammatory bowel disease. Aliment Pharmacol Ther 1998;12:673–8. [DOI] [PubMed] [Google Scholar]
- [66].Compare D, Pica L, Rocco A, De Giorgi F, Cuomo R, Sarnelli G, et al. Effects of long-term PPI treatment on producing bowel symptoms and SIBO. European Journal of Clinical Investigation 2011;41:380–6. 10.1111/j.1365-2362.2010.02419.x. [DOI] [PubMed] [Google Scholar]
- [67].Caprilli R, Frieri G, Latella G, Vernia P, Santoro ML. Faecal excretion of bicarbonate in ulcerative colitis. Digestion 1986;35:136–42. [DOI] [PubMed] [Google Scholar]
- [68].Vernia P, Caprilli R, Latella G, Barbetti F, Magliocca FM, Cittadini M. Fecal lactate and ulcerative colitis. Gastroenterology 1988;95:1564–8. [DOI] [PubMed] [Google Scholar]
- [69].Zhang H, Ameen N, Melvin JE, Vidyasagar S. Acute inflammation alters bicarbonate transport in mouse ileum. J Physiol 2007;581:1221–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Kaczmarczyk O, Dąbek-Drobny A, Woźniakiewicz M, Paśko P, Dobrowolska-Iwanek J, Woźniakiewicz A, et al. Fecal Levels of Lactic, Succinic and Short-Chain Fatty Acids in Patients with Ulcerative Colitis and Crohn Disease: A Pilot Study. J Clin Med Res 2021;10. 10.3390/jcm10204701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Premkumar K, Ramadass B, Ramakrishna BS. Fecal microbiota in inflammatory bowel disease: Studies of lactobacillus and bifidobacteria diversity. Gastroenterology, Hepatology and Endoscopy Practice 2022;2:91. [Google Scholar]
- [72].Swidsinski A, Ladhoff A, Pernthaler A, Swidsinski S, Loening-Baucke V, Ortner M, et al. Mucosal flora in inflammatory bowel disease. Gastroenterology 2002;122:44–54. [DOI] [PubMed] [Google Scholar]
- [73].Gevers D, Kugathasan S, Denson LA, Vázquez-Baeza Y, Van Treuren W, Ren B, et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host Microbe 2014;15:382–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Guan G, Lan S. Implications of Antioxidant Systems in Inflammatory Bowel Disease. Biomed Res Int 2018;2018:1290179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Machiels K, Joossens M, Sabino J, De Preter V, Arijs I, Eeckhaut V, et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014;63:1275–83. [DOI] [PubMed] [Google Scholar]
- [76].Hansen R, Russell RK, Reiff C, Louis P, McIntosh F, Berry SH, et al. Microbiota of De-Novo Pediatric IBD: Increased Faecalibacterium prausnitzii and Reduced Bacterial Diversity in Crohn’s But Not in Ulcerative Colitis. Official Journal of the American College of Gastroenterology ∣ ACG 2012;107:1913. [DOI] [PubMed] [Google Scholar]
- [77].Joossens M, Huys G, Cnockaert M, De Preter V, Verbeke K, Rutgeerts P, et al. Dysbiosis of the faecal microbiota in patients with Crohn’s disease and their unaffected relatives. Gut 2011;60:631–7. [DOI] [PubMed] [Google Scholar]
- [78].Mulder IE, Schmidt B, Lewis M, Delday M, Stokes CR, Bailey M, et al. Restricting microbial exposure in early life negates the immune benefits associated with gut colonization in environments of high microbial diversity. PLoS One 2011;6:e28279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Shirkey TW, Siggers RH, Goldade BG, Marshall JK, Drew MD, Laarveld B, et al. Effects of commensal bacteria on intestinal morphology and expression of proinflammatory cytokines in the gnotobiotic pig. Exp Biol Med 2006;231:1333–45. [DOI] [PubMed] [Google Scholar]
- [80].Mirsepasi-Lauridsen HC, Vallance BA, Krogfelt KA, Petersen AM. Escherichia coli Pathobionts Associated with Inflammatory Bowel Disease. Clin Microbiol Rev 2019;32:e00060–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Lee J-Y, Cevallos SA, Byndloss MX, Tiffany CR, Olsan EE, Butler BP, et al. High-Fat Diet and Antibiotics Cooperatively Impair Mitochondrial Bioenergetics to Trigger Dysbiosis that Exacerbates Pre-inflammatory Bowel Disease. Cell Host Microbe 2020;28:273–84.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Axelrad JE, Cadwell KH, Colombel J-F, Shah SC. The role of gastrointestinal pathogens in inflammatory bowel disease: a systematic review. Therap Adv Gastroenterol 2021;14:17562848211004493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Ross BD, Verster AJ, Radey MC, Schmidtke DT, Pope CE, Hoffman LR, et al. Human gut bacteria contain acquired interbacterial defence systems. Nature 2019;575:224–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Verster AJ, Ross BD, Radey MC, Bao Y, Goodman AL, Mougous JD, et al. The Landscape of Type VI Secretion across Human Gut Microbiomes Reveals Its Role in Community Composition. Cell Host Microbe 2017;22:411–9.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Mills RH, Dulai PS, Vázquez-Baeza Y, Sauceda C, Daniel N, Gerner RR, et al. Multi-omics analyses of the ulcerative colitis gut microbiome link Bacteroides vulgatus proteases with disease severity. Nat Microbiol 2022;7:262–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Zafar H, Saier MH Jr. Gut Bacteroides species in health and disease. Gut Microbes 2021;13:1–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Kordahi MC, Stanaway IB, Avril M, Chac D, Blanc M-P, Ross B, et al. Genomic and functional characterization of a mucosal symbiont involved in early-stage colorectal cancer. Cell Host Microbe 2021;29:1589–98.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [88].Lopez J, Grinspan A. Fecal Microbiota Transplantation for Inflammatory Bowel Disease. Gastroenterol Hepatol 2016;12:374–9. [PMC free article] [PubMed] [Google Scholar]
- [89].Hahm E-R, Singh SV. Sulforaphane inhibits constitutive and interleukin-6-induced activation of signal transducer and activator of transcription 3 in prostate cancer cells. Cancer Prev Res (Phila) 2010;3:484–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Fritsch J, Garces L, Quintero MA, Pignac-Kobinger J, Santander AM, Fernández I, et al. Low-Fat, High-Fiber Diet Reduces Markers of Inflammation and Dysbiosis and Improves Quality of Life in Patients With Ulcerative Colitis. Clin Gastroenterol Hepatol 2021;19:1189–99.e30. [DOI] [PubMed] [Google Scholar]
- [91].Li H, Christman LM, Li R, Gu L. Synergic interactions between polyphenols and gut microbiota in mitigating inflammatory bowel diseases. Food Funct 2020;11:4878–91. [DOI] [PubMed] [Google Scholar]
- [92].Pap N, Fidelis M, Azevedo L, do Carmo MAV, Wang D, Mocan A, et al. Berry polyphenols and human health: evidence of antioxidant, anti-inflammatory, microbiota modulation, and cell-protecting effects. Current Opinion in Food Science 2021;42:167–86. [Google Scholar]
- [93].Wu M, Luo Q, Nie R, Yang X, Tang Z, Chen H. Potential implications of polyphenols on aging considering oxidative stress, inflammation, autophagy, and gut microbiota. Crit Rev Food Sci Nutr 2021;61:2175–93. [DOI] [PubMed] [Google Scholar]
- [94].Schroeder BO, Birchenough GMH, Ståhlman M, Arike L, Johansson MEV, Hansson GC, et al. Bifidobacteria or Fiber Protects against Diet-Induced Microbiota-Mediated Colonic Mucus Deterioration. Cell Host Microbe 2018;23:27–40.e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Conlon MA, Bird AR. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2014;7:17–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [96].Turnbaugh PJ, Ridaura VK, Faith JJ, Rey FE, Knight R, Gordon JI. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Science Translational Medicine 2009;1:6ra14–6ra14. 10.1126/scitranslmed.3000322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [97].David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, Wolfe BE, et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014;505:559–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Filippo CD, Cavalieri D, Paola MD, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proceedings of the National Academy of Sciences 2010;107:14691–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Hildebrandt MA, Hoffmann C, Sherrill-Mix SA, Keilbaugh SA, Hamady M, Chen Y-Y, et al. High-fat diet determines the composition of the murine gut microbiome independently of obesity. Gastroenterology 2009;137:1716–24.e1–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Tu P, Chi L, Bodnar W, Zhang Z, Gao B, Bian X, et al. Gut Microbiome Toxicity: Connecting the Environment and Gut Microbiome-Associated Diseases. Toxics 2020;8:19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Zhang Y-J, Li S, Gan R-Y, Zhou T, Xu D-P, Li H-B. Impacts of gut bacteria on human health and diseases. Int J Mol Sci 2015;16:7493–519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Plumb GW, Price KR, Rhodes MJ, Williamson G. Antioxidant properties of the major polyphenolic compounds in broccoli. Free Radio Res 1997;27:429–35. [DOI] [PubMed] [Google Scholar]
- [103].Kurilich AC, Tsau GJ, Brown A, Howard L, Klein BP, Jeffery EH, et al. Carotene, tocopherol, and ascorbate contents in subspecies of Brassica oleracea. J Agric Food Chem 1999;47:1576–81. [DOI] [PubMed] [Google Scholar]
- [104].Tse G, Eslick GD. Cruciferous vegetables and risk of colorectal neoplasms: a systematic review and meta-analysis. Nutr Cancer 2014;66:128–39. [DOI] [PubMed] [Google Scholar]
- [105].Huang L, Li B-L, He C-X, Zhao Y-J, Yang X-L, Pang B, et al. Sulforaphane inhibits human bladder cancer cell invasion by reversing epithelial-to-mesenchymal transition via directly targeting microRNA-200c/ZEB1 axis. J Funct Foods 2018;41:118–26. [Google Scholar]
- [106].Burnett JP, Lim G, Li Y, Shah RB, Lim R, Paholak HJ, et al. Sulforaphane enhances the anticancer activity of taxanes against triple negative breast cancer by killing cancer stem cells. Cancer Lett 2017;394:52–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [107].Liu X-L, Zhang X-T, Meng J, Zhang H-F, Zhao Y, Li C, et al. ING5 knockdown enhances migration and invasion of lung cancer cells by inducing EMT via EGFR/PI3K/Akt and IL-6/STAT3 signaling pathways. Oncotarget 2017;8:54265–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [108].Mazarakis N, Snibson K, Licciardi PV, Karagiannis TC. The potential use of l-sulforaphane for the treatment of chronic inflammatory diseases: A review of the clinical evidence. Clin Nutr 2020;39:664–75. [DOI] [PubMed] [Google Scholar]
- [109].Bartnik M, Facey PC. Chapter 8 - Glycosides. In: Badal S, Delgoda R, editors. Pharmacognosy, Boston: Academic Press; 2017, p. 101–61. [Google Scholar]
- [110].Burnett JP, Korkaya H, Ouzounova MD, Jiang H, Conley SJ, Newman BW, et al. Trastuzumab resistance induces EMT to transform HER2+ PTEN− to a triple negative breast cancer that requires unique treatment options. Sci Rep 2015;5:1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [111].Sun L, Burnett J, Zou H, Sun D. Overexpression of ALDH1A1 is associated with poor prognosis and therapeutic effects of sulforaphane in esophageal cancer. Int J Clin Exp Pathol 2016;9:11670–5. [Google Scholar]
- [112].Li X, Zhao Z, Li M, Liu M, Bahena A, Zhang Y, et al. Sulforaphane promotes apoptosis, and inhibits proliferation and self-renewal of nasopharyngeal cancer cells by targeting STAT signal through miRNA-124-3p. Biomed Pharmacother 2018;103:473–81. [DOI] [PubMed] [Google Scholar]
- [113].Li Y, Zhang T, Korkaya H, Liu S, Lee H-F, Newman B, et al. Sulforaphane, a dietary component of broccoli/broccoli sprouts, inhibits breast cancer stem cells. Clin Cancer Res 2010;16:2580–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [114].Hanschen FS, Klopsch R, Oliviero T, Schreiner M, Verkerk R, Dekker M. Optimizing isothiocyanate formation during enzymatic glucosinolate breakdown by adjusting pH value, temperature and dilution in Brassica vegetables and Arabidopsis thaliana. Sci Rep 2017;7:40807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Matusheski NV, Swarup R, Juvik JA, Mithen R, Bennett M, Jeffery EH. Epithiospecifier protein from broccoli (Brassica oleracea L. ssp. italica) inhibits formation of the anticancer agent sulforaphane. J Agric Food Chem 2006;54:2069–76. [DOI] [PubMed] [Google Scholar]
- [116].Westphal A, Riedl KM, Cooperstone JL, Kamat S, Balasubramaniam VM, Schwartz SJ, et al. High-Pressure Processing of Broccoli Sprouts: Influence on Bioactivation of Glucosinolates to Isothiocyanates. J Agric Food Chem 2017;65:8578–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [117].Sangkret S, Pongmalai P, Devahastin S, Chiewchan N. Enhanced production of sulforaphane by exogenous glucoraphanin hydrolysis catalyzed by myrosinase extracted from Chinese flowering cabbage (Brassica rapa var. parachinensis). Sci Rep 2019;9:9882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [118].Howard LA, Jeffery EH, Wallig MA, Klein BP. Retention of Phytochemicals in Fresh and Processed Broccoli. Journal of Food Science 1997;62:1098–104. [Google Scholar]
- [119].Bones AM, Rossiter JT. The enzymic and chemically induced decomposition of glucosinolates. Phytochemistry 2006;67:1053–67. [DOI] [PubMed] [Google Scholar]
- [120].Angelino D, Dosz EB, Sun J, Hoeflinger JL, Van Tassell ML, Chen P, et al. Myrosinase-dependent and -independent formation and control of isothiocyanate products of glucosinolate hydrolysis. Front Plant Sci 2015;6:831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [121].Angelino D, Jeffery E. Glucosinolate hydrolysis and bioavailability of resulting isothiocyanates: Focus on glucoraphanin. J Funct Foods 2014;7:67–76. [Google Scholar]
- [122].Petkovic M, Leal EC, Alves I, Bose C, Palade PT, Singh P, et al. Dietary supplementation with sulforaphane ameliorates skin aging through activation of the Keap1-Nrf2 pathway. J Nutr Biochem 2021;98:108817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [123].Ziady AG, Sokolow A, Shank S, Corey D, Myers R, Plafker S, et al. Interaction with CREB binding protein modulates the activities of Nrf2 and NF-κB in cystic fibrosis airway epithelial cells. Am J Physiol Lung Cell Mol Physiol 2012;302:L1221–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [124].Sun C-C, Li S-J, Yang C-L, Xue R-L, Xi Y-Y, Wang L, et al. Sulforaphane attenuates muscle inflammation in dystrophin-deficient mdx mice via NF-E2-related factor 2 (Nrf2)-mediated inhibition of NF-κB signaling pathway. J Biol Chem 2015;290:17784–95. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- [125].Heiss E, Herhaus C, Klimo K, Bartsch H, Gerhäuser C. Nuclear Factor κB Is a Molecular Target for Sulforaphane-mediated Anti-inflammatory Mechanisms. J Biol Chem 2001;276:32008–15. [DOI] [PubMed] [Google Scholar]
- [126].Lai R-H, Miller MJ, Jeffery E. Glucoraphanin hydrolysis by microbiota in the rat cecum results in sulforaphane absorption. Food Funct 2010;1:161–6. [DOI] [PubMed] [Google Scholar]
- [127].Xu X, Dai M, Lao F, Chen F, Hu X, Liu Y, et al. Effect of glucoraphanin from broccoli seeds on lipid levels and gut microbiota in high-fat diet-fed mice. J Funct Foods 2020;68:103858. [Google Scholar]
- [128].Shapiro TA, Fahey JW, Wade KL, Stephenson KK, Talalay P. Human metabolism and excretion of cancer chemoprotective glucosinolates and isothiocyanates of cruciferous vegetables. Cancer Epidemiol Biomarkers Prev 1998;7:1091–100. [PubMed] [Google Scholar]
- [129].Elfoul L, Rabot S, Khelifa N, Quinsac A, Duguay A, Rimbault A. Formation of allyl isothiocyanate from sinigrin in the digestive tract of rats monoassociated with a human colonic strain of Bacteroides thetaiotaomicron. FEMS Microbiol Lett 2001;197:99–103. [DOI] [PubMed] [Google Scholar]
- [130].Vermeulen M, Klöpping-Ketelaars IWAA, van den Berg R, Vaes WHJ. Bioavailability and kinetics of sulforaphane in humans after consumption of cooked versus raw broccoli. J Agric Food Chem 2008;56:10505–9. [DOI] [PubMed] [Google Scholar]
- [131].Li Y, Zhang T, Li X, Zou P, Schwartz SJ, Sun D. Kinetics of sulforaphane in mice after consumption of sulforaphane-enriched broccoli sprout preparation. Mol Nutr Food Res 2013;57:2128–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [132].Rouzaud G, Rabot S, Ratcliffe B, Duncan AJ. Influence of plant and bacterial myrosinase activity on the metabolic fate of glucosinolates in gnotobiotic rats. Br J Nutr 2003;90:395–404. [DOI] [PubMed] [Google Scholar]
- [133].Bheemreddy RM, Jeffery EH. The metabolic fate of purified glucoraphanin in F344 rats. J Agric Food Chem 2007;55:2861–6. [DOI] [PubMed] [Google Scholar]
- [134].Lai R-H, Keck A-S, Wallig MA, West LG, Jeffery EH. Evaluation of the safety and bioactivity of purified and semi-purified glucoraphanin. Food Chem Toxicol 2008;46:195–202. [DOI] [PubMed] [Google Scholar]
- [135].Fahey JW, Wehage SL, Holtzclaw WD, Kensler TW, Egner PA, Shapiro TA, et al. Protection of humans by plant glucosinolates: efficiency of conversion of glucosinolates to isothiocyanates by the gastrointestinal microflora. Cancer Prev Res (Phila) 2012;5:603–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [136].Conaway CC, Getahun SM, Liebes LL, Pusateri DJ, Topham DK, Botero-Omary M, et al. Disposition of glucosinolates and sulforaphane in humans after ingestion of steamed and fresh broccoli. Nutr Cancer 2000;38:168–78. [DOI] [PubMed] [Google Scholar]
- [137].Clarke JD, Hsu A, Riedl K, Bella D, Schwartz SJ, Stevens JF, et al. Bioavailability and inter-conversion of sulforaphane and erucin in human subjects consuming broccoli sprouts or broccoli supplement in a cross-over study design. Pharmacol Res 2011;64:456–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [138].Jeffery EH, Araya M. Physiological effects of broccoli consumption. Phytochem Rev 2009;8:283–98. [Google Scholar]
- [139].Gil V, MacLeod AJ. The effects of pH on glucosinolate degradation by a thioglucoside glucohydrolase preparation. Phytochemistry 1980;19:2547–51. [Google Scholar]
- [140].Luang-In V, Albaser AA, Nueno-Palop C, Bennett MH, Narbad A, Rossiter JT. Glucosinolate and Desulfo-glucosinolate Metabolism by a Selection of Human Gut Bacteria. Curr Microbiol 2016;73:442–51. [DOI] [PubMed] [Google Scholar]
- [141].Luang-In V, Narbad A, Cebeci F, Bennett M, Rossiter JT. Identification of Proteins Possibly Involved in Glucosinolate Metabolism in L. agilis R16 and E. coli VL8. Protein J 2015;34:135–46. [DOI] [PubMed] [Google Scholar]
- [142].Luang-In V, Narbad A, Nueno-Palop C, Mithen R, Bennett M, Rossiter JT. The metabolism of methylsulfinylalkyl- and methylthioalkyl-glucosinolates by a selection of human gut bacteria. Mol Nutr Food Res 2014;58:875–83. [DOI] [PubMed] [Google Scholar]
- [143].Luang-In V, Deeseenthum S, Udomwong P, Saengha W, Gregori M. Formation of sulforaphane and iberin products from Thai cabbage fermented by myrosinase-positive bacteria. Molecules 2018;23:955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [144].Wathelet J-P, Iori R, Leoni O, Rollin P, Mabon N, Marlier M, et al. A recombinant β-O-glucosidase from Caldocellum saccharolyticum to hydrolyse desulfoglucosinolates. Biotechnol Lett 2001;23:443–6. [Google Scholar]
- [145].Wu Y, Shen Y, Zhu Y, Mupunga J, Zou L, Liu C, et al. Broccoli ingestion increases the glucosinolate hydrolysis activity of microbiota in the mouse gut. Int J Food Sci Nutr 2019;70:585–94. [DOI] [PubMed] [Google Scholar]
- [146].Flores R, Shi J, Gail MH, Gajer P, Ravel J, Goedert JJ. Association of fecal microbial diversity and taxonomy with selected enzymatic functions. PLoS One 2012;7:e39745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [147].Blanco G, Sánchez B, Fdez-Riverola F, Margolles A, Lourenço A. In silico Approach for Unveiling the Glycoside Hydrolase Activities in Faecalibacterium prausnitzii Through a Systematic and Integrative Large-Scale Analysis. Front Microbiol 2019;10:517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [148].Narbad A, Rossiter JT. Gut Glucosinolate Metabolism and Isothiocyanate Production. Mol Nutr Food Res 2018;62:e1700991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [149].Palop ML, LLanos Palop M, Smiths JP, ten Brink B. Degradation of sinigrin by Lactobacillus agilis strain R16. International Journal of Food Microbiology 1995;26:219–29. [DOI] [PubMed] [Google Scholar]
- [150].Li F, Hullar MAJ, Beresford SAA, Lampe JW. Variation of glucoraphanin metabolism in vivo and ex vivo by human gut bacteria. Br J Nutr 2011;106:408–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [151].Liu X, Wang Y, Hoeflinger JL, Neme BP, Jeffery EH, Miller MJ. Dietary Broccoli Alters Rat Cecal Microbiota to Improve Glucoraphanin Hydrolysis to Bioactive Isothiocyanates. Nutrients 2017/3;9:262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [152].Rabot S, Nugon-Baudon L, Raibaud P, Szylit O. Rape-seed meal toxicity in gnotobiotic rats: influence of a whole human faecal flora or single human strains of Escherichia coli and Bacteroides vulgatus. Br J Nutr 1993;70:323–31. [DOI] [PubMed] [Google Scholar]
- [153].Mullaney JA, Kelly WJ, McGhie TK, Ansell J, Heyes JA. Lactic acid bacteria convert glucosinolates to nitriles efficiently yet differently from enterobacteriaceae. J Agric Food Chem 2013;61:3039–46. [DOI] [PubMed] [Google Scholar]
- [154].Liou CS, Sirk SJ, Diaz CAC, Klein AP, Fischer CR, Higginbottom SK, et al. A Metabolic Pathway for Activation of Dietary Glucosinolates by a Human Gut Symbiont. Cell 2020;180:717–28.e19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [155].Tanes C, Bittinger K, Gao Y, Friedman ES, Nessel L, Paladhi UR, et al. Role of dietary fiber in the recovery of the human gut microbiome and its metabolome. Cell Host Microbe 2021;29:394–407.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [156].Liu M, Li X, Zhou S, Wang TTY, Zhou S, Yang K, et al. Dietary fiber isolated from sweet potato residues promotes a healthy gut microbiome profile. Food Funct 2020;11:689–99. [DOI] [PubMed] [Google Scholar]
- [157].Cui J, Lian Y, Zhao C, Du H, Han Y, Gao W, et al. Dietary Fibers from Fruits and Vegetables and Their Health Benefits via Modulation of Gut Microbiota. Compr Rev Food Sci Food Saf 2019;18:1514–32. [DOI] [PubMed] [Google Scholar]
- [158].Zandani G, Kaftori-Sandler N, Sela N, Nyska A, Madar Z. Dietary broccoli improves markers associated with glucose and lipid metabolism through modulation of gut microbiota in mice. Nutrition 2021:111240. [DOI] [PubMed] [Google Scholar]
- [159].Choi K-M, Lee Y-S, Sin D-M, Lee S, Lee MK, Lee Y-M, et al. Sulforaphane inhibits mitotic clonal expansion during adipogenesis through cell cycle arrest. Obesity 2012;20:1365–71. [DOI] [PubMed] [Google Scholar]
- [160].Kaczmarek JL, Liu X, Charron CS, Novotny JA, Jeffery EH, Seifried HE, et al. Broccoli consumption affects the human gastrointestinal microbiota. J Nutr Biochem 01/2019;63:27–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [161].Jun S-R, Cheema A, Bose C, Boerma M, Palade PT, Carvalho E, et al. Multi-Omic Analysis Reveals Different Effects of Sulforaphane on the Microbiome and Metabolome in Old Compared to Young Mice. Microorganisms 2020;8:1500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [162].Zhang Y, Tan L, Li C, Wu H, Ran D, Zhang Z. Sulforaphane alter the microbiota and mitigate colitis severity on mice ulcerative colitis induced by DSS. AMB Express 2020;10:119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [163].Zandani G, Anavi-Cohen S, Tsybina-Shimshilashvili N, Sela N, Nyska A, Madar Z. Broccoli Florets Supplementation Improves Insulin Sensitivity and Alters Gut Microbiome Population-A Steatosis Mice Model Induced by High-Fat Diet. Front Nutr 2021;8:680241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [164].Paramsothy S, Nielsen S, Kamm MA, Deshpande NP, Faith JJ, Clemente JC, et al. Specific Bacteria and Metabolites Associated With Response to Fecal Microbiota Transplantation in Patients With Ulcerative Colitis. Gastroenterology 2019;156:1440–54.e2. [DOI] [PubMed] [Google Scholar]
- [165].Zhang L, Liu F, Xue J, Lee SA, Liu L, Riordan SM. Bacterial Species Associated With Human Inflammatory Bowel Disease and Their Pathogenic Mechanisms. Front Microbiol 2022;13:801892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [166].Abukhabta S, Khalil Ghawi S, Karatzas KA, Charalampopoulos D, McDougall G, Allwood JW, et al. Sulforaphane-enriched extracts from glucoraphanin-rich broccoli exert antimicrobial activity against gut pathogens in vitro and innovative cooking methods increase in vivo intestinal delivery of sulforaphane. Eur J Nutr 2021;60:1263–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [167].Aires A, Mota VR, Saavedra MJ, Rosa EAS, Bennett RN. The antimicrobial effects of glucosinolates and their respective enzymatic hydrolysis products on bacteria isolated from the human intestinal tract. J Appl Microbiol 2009;106:2086–95. [DOI] [PubMed] [Google Scholar]
- [168].Rosendale D, Butts C, de Guzman E, Maddox I, Martell S, McIntyre L, et al. The contribution of dietary broccoli sprouts towards the microbial metabolite profile in the hind gut of mice. Int J Food Sci Technol 2012;47:1328–32. [Google Scholar]
- [169].Fahey JW, Stephenson KK, Wade KL, Talalay P. Urease from Helicobacter pylori is inactivated by sulforaphane and other isothiocyanates. Biochem Biophys Res Commun 2013;435:1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [170].Mirmiran P, Bahadoran Z, Ghasemi A, Jeddi S, Azizi F. Short communications. J Funct Foods 2017;C:356–8. [Google Scholar]
- [171].Lamers CR, de Roos NM, Heerink HH, van de Worp-Kalter LA, Witteman BJM. P550 Short term effects of a combined lifestyle intervention in patients with Inflammatory Bowel Disease. J Crohns Colitis 2021;15:S518. [Google Scholar]
- [172].Wedick NM, Ma Y, Olendzki BC, Procter-Gray E, Cheng J, Kane KJ, et al. Access to healthy food stores modifies effect of a dietary intervention. Am J Prev Med 2015;48:309–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [173].Kellingray L, Saha S, Doleman J, Narbad A, Mithen R. Investigating the metabolism of glucoraphanin by the human gut microbiota using ex vivo culturing methods. Proceedings of the Nutrition Society, vol. 73(OCE1), Cambridge University Press; 2014, p. E34. [Google Scholar]
- [174].Getahun SM, Chung FL. Conversion of glucosinolates to isothiocyanates in humans after ingestion of cooked watercress. Cancer Epidemiol Biomarkers Prev 1999;8:447–51. [PubMed] [Google Scholar]
- [175].Shapiro TA, Fahey JW, Wade KL, Stephenson KK, Talalay P. Chemoprotective glucosinolates and isothiocyanates of broccoli sprouts: metabolism and excretion in humans. Cancer Epidemiol Biomarkers Prev 2001;10:501–8. [PubMed] [Google Scholar]
- [176].Rouzaud G, Young SA, Duncan AJ. Hydrolysis of glucosinolates to isothiocyanates after ingestion of raw or microwaved cabbage by human volunteers. Cancer Epidemiol Biomarkers Prev 2004;13:125–31. [DOI] [PubMed] [Google Scholar]
- [177].Shapiro TA, Fahey JW, Dinkova-Kostova AT, Holtzclaw WD, Stephenson KK, Wade KL, et al. Safety, tolerance, and metabolism of broccoli sprout glucosinolates and isothiocyanates: a clinical phase I study. Nutr Cancer 2006;55:53–62. [DOI] [PubMed] [Google Scholar]
- [178].Martiny JBH, Bohannan BJM, Brown JH, Colwell RK, Fuhrman JA, Green JL, et al. Microbial biogeography: putting microorganisms on the map. Nat Rev Microbiol 2006;4:102–12. [DOI] [PubMed] [Google Scholar]
- [179].Gu M, Samuelson DR, de la Rua NM, Charles TP, Taylor CM, Luo M, et al. Host innate and adaptive immunity shapes the gut microbiota biogeography. Microbiol Immunol 2022;In press. 10.1111/1348-0421.12963. [DOI] [PMC free article] [PubMed] [Google Scholar]