Tang et al. 10.1073/pnas.0607374103. |
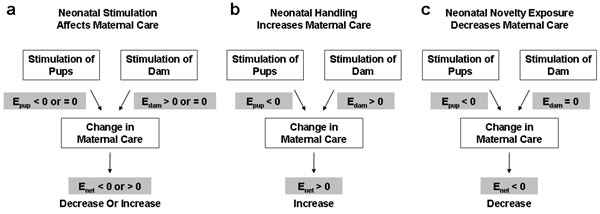
Fig. 4. Parallel-process model of early stimulation effect on maternal care. (a) The combined effects (Enet) of pup and dam stimulation on maternal care can be either an increase or decrease in maternal care. Shaded areas indicate isolated effects of pup (Epup) or dam (Edam) stimulation on maternal care. (b) Case of neonatal handling. A positive Enet can be achieved by the positive Edam overriding the negative Epup. (c) Case of neonatal novelty exposure. A negative Enet can be explained by the negative Epup alone because neonatal novelty exposure did not involve differential maternal stimulation (Edam = 0).
Supporting Methods
Study 1
Animals.
All experimental procedures were in accordance with the Institutional Animal Care and Use Committee at the University of New Mexico. Pregnant dams arrived at the vivarium 16-17 days before giving birth. Litters were culled within 24 h of birth. Because of the scope of the current study, we maximized the number of male pups and included female pups only to maintain constant litter size. On postnatal day (P) 1, pups were assigned to Novel and Home groups in a pseudorandom fashion, with equal numbers of males and female of roughly equivalent body sizes assigned to each group. Pups were housed with their dams in translucent plastic cages (51 ´25 ´ 22 cm) with food and water ad libitum. Pups were weaned on P22, and thereafter they were housed individually.Morris water task
. Rats were tested in a moving-platform version of the Morris water task, in which a different platform location was used each day (1). The hidden-platform location was selected pseudorandomly from one of four quadrants and two distances from the center of the pool without repetition. The pool, 120 cm in diameter and 52 cm in depth, was filled with tap water (~21°C) mixed with ~1 cup of powdered milk. Several conspicuous distal cues were available. On each of the 8 days of testing, each rat entered the pool from four different locations (N, S, E, W) in a pseudorandom sequence in a total of four trials. The order of water maze testing was counterbalanced between Novel and Home groups, and experimenters were blind to the animals' group identity.Corticosterone (CORT) response to expected and unexpected stressors
. The CORT response to an expected versus an unexpected stressor was assessed. The expected stressor was exposure to four trials of the water task as was experienced on the previous 8 days of water task testing. The unexpected stressor was exposure to a novel open-field during the 1-min interval between T1 and T2 of the water task. Half of the rats were exposed to the open-field on testing day 9 and no open-field on testing day 10, with the other half tested in reverse order. For each rat, four blood samples at different time points were obtained by tail nick. Before water maze training, two baseline blood samples were collected on consecutive days and averaged to obtain a single baseline CORT measure (CB). Two additional samples were collected on testing days: one after the expected water maze stressor (CWM) and the other after superimposing an unexpected open-field stressor on the expected stressor (COF+WM). The order of water maze testing and blood collection was counterbalanced between Novel and Home groups, and experimenters were blind to the animals' group identity. Blood samples were collected in EDTA-coated tubes and centrifuged. Serum was removed and stored at -20°C until an RIA was performed. Serum CORT concentrations were measured in a single assay using a Coat-A-Count corticosterone kit (Diagnostic Products, Los Angeles, CA). The lower limit of detectability was 20.45 ng/ml, and the intraassay coefficient of variation was 10.9%.Social competition
. Rats were tested for their ability to obtain chocolate rewards in the presence of a competitor. First, rats were familiarized with melted chocolate drops (Hershey's Milk Chocolate Chips), which were applied to the interior wall of their housing cages for 3-4 days before training. For 6 days of individual training carried out in a testing room, a testing apparatus containing a narrow runway was inserted into the home cage. During the first 2 training days, chocolate drops were applied at the front, along the side walls, and at the end of the runway to shape the behavior of runway entering. For the remaining 4 days of training, one chocolate drop was made available only at the end of the runway. On each training trial, after a brief tone (3 s), the opaque divider in front of the runway was removed, allowing the rat access to the chocolate for the next 60 s. Between trials, the apparatus was wiped clean, and a new chocolate drop was applied for the next trial. During the first 5 training days, rats were given a minimum of 8 and a maximum of 14 trials to reach a criterion of consuming the chocolate during 3 consecutive trials. On the last training day, rats were given 8 trials regardless of their performance, and total number of rewards obtained and latency to obtain rewards from the time of the divider removal were assessed for each rat. Food restriction was applied throughout training to maintain body weights at ~90% of baseline weight.Competition between a pair of nonsibling Novel and Home rats of similar weights was observed on 2 consecutive days in the same testing room in clean non-home cages lined with fresh bedding. Pairs of rats were placed simultaneously into the testing cage and given 5 min to habituate to the novel cage and the presence of a conspecific. At the beginning of each of the 8 trials per day, the pair was first confined to half of the testing cage by the divider, and the testing apparatus was placed into the cage on the opposite side of the divider. After the 3-s tone, the divider was removed. When one of the pair entered the runway, narrow enough to allow only one rat at a time to enter, the rat was given 3 s to consume the reward at the end of the runway before the trial was terminated and the apparatus removed. If neither rat obtained the reward within 60 s, the trial was terminated. Between trials, the end of the runway was wiped clean, and a new reward was applied. A trial was considered a competition trial if both rats attempted to enter the runway during the 60-s interval (~45% of trials). The winner of a competition trial was defined as the rat that obtained the chocolate first. Latency of the winner to obtain the reward was measured for each trial.
Statistical analysis
. Because multiple rats from the same litters were used, we first tested for a litter effect on all dependent measures. Significant litter effects were not found; therefore, all statistical tests were performed using individual rat as the unit of analysis. The effect size is reported as Cohen's d (large, >0.8; medium, >0.5; small, <0.2) (2); all effect sizes within the present study were medium to large.Study 2
Pup retrieval latency.
Within the same groups of rats described in Study 1, we examined maternal retrieval of pups after the neonatal novelty exposure procedure on P1-8. To habituate dams to the presence of the container, a clean plastic open-top container (depth = 3.2 cm) was placed in the dam's cage for 10 min on at least 2 consecutive days »5 days before the expected birth date. On novelty exposure days, the location of the container was placed at the opposite end of the cage from the original nest location. Each day, a clean container was used for each litter. Dams typically completed retrieval of their entire litters within 10 min. The observation ended at 10 min if the dam was no longer engaged in retrieval or had not retrieved at all; otherwise, the dam was allowed to complete the retrieval. Within this small time window, frequencies of other maternal-care behaviors were very low, and therefore we report only retrieval-related measures here. Retrieval behavior was observed for the first 8 postnatal days because after this day pups began to crawl out of the containers, hence circumventing maternal retrieval. If a pup was picked up more than once by the dam, only the latency of the first pickup was recorded; if a pup was not retrieved, which happened rarely, no latency was recorded. An average latency for each dam was computed across days.Relation of pup retrieval to active nursing
. A second experiment was performed on a separate group of 15 pregnant Long-Evans hooded rats that arrived at our laboratory 12 days before giving birth (Harlan, Indianapolis, IN). Litters were culled and housed as described previously except the light cycle began at 0800. Pup retrieval was observed after the dam and pups were removed from the home cage for varying intervals of 0, 3, and 15 min delays (n = 5 per group). The variations in intervals were used to create variations in maternal-care behaviors and to determine potential influences of separation duration on maternal-care behavior. Because no effect of separation duration was detected, this variable received no further analysis. Three litters were excluded from analysis because two did not retrieve across all days within the time period allotted, and one was an outlier. Active nursing was observed during five around-the-clock sessions: at 0900, 1300, and 1600 during the light period and at 0600 and 2100 during the dark period. Observations during the dark cycle were made under red light illumination. During each session, 25 cycles of 3-min observations were made. During a cycle, each of the 15 litters was observed sequentially for 10s. For each dam, a total of 125 observations were made daily (25 cycles per period ´ 5 periods per day = 125 observations per day). The following behaviors were scored: (i) licking and grooming, (ii) arched-back nursing, (iii) blanket nursing, (iv) passive nursing, and (v) no contact with pups (for detailed description, see ref. 3). Interrater reliability was assessed by using 75 observations from P1, and it was computed for each of the maternal measures among three raters; all reliability scores were ³0.90.Discussion: Variables Influencing Maternal Care
One of the conclusions that can be drawn from the present data is that neonatal handling and neonatal novelty exposure procedures produced opposite effects on maternal care, with the former increasing and the latter decreasing maternal-care behaviors. We suggest that both findings can be explained within a common framework, in which the final maternal-care outcome is jointly determined by at least two inputs: an effect by the stimulation of pups (Epup) and an effect by the stimulation of the dam (Edam; Fig. 4a). The first effect provides a negative input to the maternal-care outcome (indicated by the shaded area on the left) through differential signaling of the pups, with stimulated pups receiving lower priority in maternal care, as supported by Young's (4) and our current findings. The second effect provides a positive input through differential maternal stimulation (indicated by the shaded area on the right), with dams that experienced brief maternal separation providing more maternal care (5-8).
In the context of neonatal handling, the positive effect of differential stimulation between dams of handled and nonhandled pups must override the negative effect of pup stimulation on maternal-care behavior, giving rise to a net increase in maternal care toward handled pups (Fig. 4b). In the context of neonatal novelty exposure, because the same dams cared for Novel and Home pups in our within-litter design, there is no differential dam stimulation. Differential pup stimulation is the only pathway that contributes to differential maternal care, giving rise to a net decrease in maternal care (Fig. 4c).
1. Whishaw IQ (1985) Physiol Behav 35:139-143.
2. Rosenthal R, Rosnow RL (1984) Essentials of Behavioral Research: Methods and Data Analysis (McGraw-Hill Co., New York).
3. Myers MM, Brunelli SA, Squire JM, Shindeldecker RD, Hofer MA (1989) Dev Psychobiol 22:29-53.
4. Young RD (1965) Psychonom Sci 3:295-296.
5. Thoman EB, Levine S (1970) Physiol Behav 5:1417-1421.
6. Liu D, Diorio J, Tannenbaum B, Caldji C, Francis D, Freedman A, Sharma S, Pearson D, Plotsky PM, Meaney MJ (1997) Science 277:1659-1662.
7. Francis D, Diorio J, Liu D, Meaney MJ (1999) Science 286:1155-1158.
8. Macri S, Mason GJ, Wurbel H (2004) Eur J Neurosci 20:1017-1024.