Files in this Data Supplement:
Fig. S1. Alignment of ANX1, ANX2 and FER amino acid sequences and ANX-YFP fusions in tobacco epidermal cells. (A) Alignment of ANX1, ANX2 and FER amino acid sequences. The alignment was performed using Jellyfish (LabVelocity). Residues conserved across the three sequences are shaded black, whereas residues conserved across two sequences are shaded gray. FER shares 51.1% identity with both ANX1 and ANX2, whereas ANX1 and ANX2 are 85.6% identical. The dashed line underlines the kinase domains, whereas the triple line indicates the transmembrane domains. ANX1 (NP_187120); ANX2 (NP_198220); FER (NP_190723). (B) Plasmids encoding YFP, FLS2-YFP, ANX1-YFP and ANX2-YFP under the control of the constitutive 35S promoter were transiently expressed in tobacco epidermal tissue. Filters are indicated at the top. Similar to the membrane-localized FLS2-YFP fusion protein, ANX1-YFP and ANX2-YFP fusion proteins localize uniformly at the cell periphery.
Fig. S2. Localization of ANX-YFP fusion proteins in plasmolysed or dead pollen tubes and in germinating pollen grains. (A) Plasmolysis of ANX-YFP-expressing pollen tubes confirms the plasma membrane localization of ANX1-YFP and ANX2-YFP fusion proteins. Note also that unlike ACA9-YFP-expressing pollen tubes, a weak residual fluorescence signal is consistently found close to the cell wall of ANX1-YFP and ANX2-YFP expressing pollen tubes after protoplast retraction (white arrows). (B) Pollen grains from pACA9-ANX1-YFP-expressing transgenic T1 Arabidopsis line. Asterisk indicates a non-fluorescent pollen grain (left). On the right, the strongest fluorescent ANX1-YFP signal (arrows) is located polarly towards the emerging tip of a germinating fluorescent pollen grain. (C) A dead pollen tube from a pACA9-ANX2-YFP-expressing transgenic T1 Arabidopsis line displays a completely different fluorescent pattern to actively growing pollen tubes (see Fig. 2). Upper panel, bright-field micrograph; bottom panel, YFP filter. Scale bars: 5 m in A; 7.5 m in B; 15 m in C.
Fig. S3. Pollination of the male-sterile dde2-2 mutant pistils by pollen from anx1-1/anx1-1 anx2-1/ANX2, anx1-2/anx1-2 anx2-2/ANX2 and anx1-2/anx1-2 anx2-2/anx2-2 reveals the male sterility of the anx1-2/anx1-2 anx2-2/anx2-2 mutant. Twelve dde2-2 pistils for each pollen donor genotype (indicated on the left) were manually pollinated resulting in normal developing siliques with pollen from anx1-1/anx1-1anx2-1/ANX2 (second panel) and anx1-2/anx1-2anx2-2/ANX2 (third panel) plants. In clear contrast, none of the twelve pistils pollinated by anx1-2/anx1-2 anx2-2/anx2-2 pollen (bottom panel) could produce a seed-containing silique. The green arrows indicate the position of the pistils manually pollinated.
Fig. S4. Time-lapse imaging of anx1-2 anx2-2 mutant pollen growth in vitro shows that mutant pollen can suffer discharge several times. (A) Imaging of anx1-2 anx2-2 mutant pollen from 1 to 6 hours and 30 minutes of incubation in vitro on pollen germination medium. White arrows indicate the direction of the first pollen discharge, and white arcs show the emerging bulges or tips that precede a discharge. Yellow arrows show the direction of the second discharge, and the green arrow indicates the direction of the third discharge. Time is in hours, minutes and seconds after the start of the incubation. (B) Rare example of a anx1-2 anx2-2 mutant pollen that has discharged a first time (white arrow) but was able to recover to produce a long pollen tube which ultimately discharged (yellow arrow). Scale bars: 25 m in A; 10 m in B.
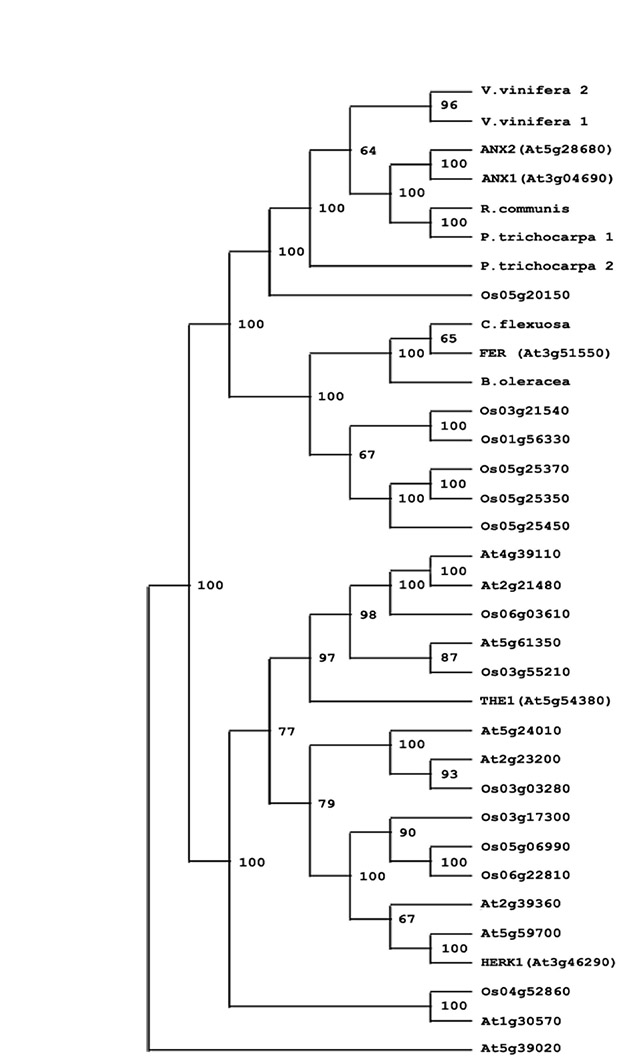
Fig. S5. Phylogenetic tree showing ANX1 and ANX2 homologs in Arabidopsis thaliana (At, Brassicaceae), Oryza sativa (Os, Poaceae), Cardamine flexuosa, Brassica oleracea (Brassicaceae), Populus trichocarpa (Salicaceae), Vitis vinifera (Vitaceae) and Ricinus communis (Euphorbiaceae). Bootstrap numbers at the forks indicate how many times the sequences to the right of the fork occurred in the same group of 100 trees. A subset of 34 sequences was used. At5g39020 protein sequence was used as an out-group. Multiple alignments were performed with CLUSTALW (Thompson et al., 1994). Phylogenetic analysis was performed with the PHYLIP package (http://evolution.genetics.washington.edu/phylip/) using the protein sequence parsimony method (PROTPARS) on 100 bootstrap replicates with jumbling of the order of sequences three times for each replicate. GenBank submission numbers for ANX1 and ANX2 homologs: Cardamine flexuosa (ABT18099), Brassica oleracea (ABT18094), Ricinus communis (EEF31088), Populus trichocarpa 1 (XP_002319081), P. trichocarpa 2 (XP_002316044), Vitis vinifera 1 (CAO40132), V. vinifera 2 (CAO44912).
Reference
Thompson, J. D., Higgins, D. G. and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673-4680.
Movie 1. Time-lapse imaging of actively growing Arabidopsis pollen tubes expressing pACA9-ANX1-YFP shows recycling of the membrane-bound ANX1-RLK at the pollen tube tip. The clear inverse cone pool of vesicles moving from the apex to the basal pole of the pollen tube is consistent with endocytosis of membrane materials at the apex of the tube. Although harder to observe, small vesicles moving from the basal pole to the subapical zone of the apex were also present and are consistent with exocytosis not at the apex itself, but at the subapical zone in agreement with the putative ACE-H model (Zonia and Meunik, 2009). Note that the drop in fluorescence intensity over time is due to photobleaching. Images were recorded every 3 seconds.
Movie 2. Time-lapse imaging of actively growing Arabidopsis pollen tubes expressing pACA9-ANX2-YFP shows recycling of the membrane-bound ANX2-RLK at the pollen tube tip. The clear inverse cone pool of vesicles moving from the apex to the basal pole of the pollen tube is consistent with endocytosis of membrane materials at the apex of the tube. Although harder to observe, small vesicles moving from the basal pole to the subapical zone of the apex were also present and are consistent with exocytosis not at the apex itself, but at the subapical zone in agreement with the putative ACE-H model (Zonia and Meunik, 2009). Note that the drop in fluorescence intensity over time is due to photobleaching. Images were recorded every 20 seconds.
Movie 3. Explosive discharge of an anx1 anx2 mutant pollen emerging tip at the subapical zone of the tip. Note that the tip had stopped growing before bursting at the subapical zone of the apex. Images were recorded every second for 30 seconds. See corresponding Fig. 4B.
Movie 4. Montage of time-lapse imaging over several hours of anx1 anx2 mutant pollen showing that mutant pollen can discharge several times. See corresponding Fig. S4.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}