 1 controls basement membrane assembly in the myotome
1 controls basement membrane assembly in the myotomeFiles in this Data Supplement:
Fig. S1. Formation of a laminin-containing myotomal basement membrane is impaired in Gli compound mutant embryos. (A-L) Myf5, laminin and myogenin protein expression was examined by in situ hybridization and immunohistochemistry with confocal imaging in forelimb somites of E9.5 wild-type (A,E,I), Gli2+/−;Gli3−/− (B,F,J), Gli2−/−;Gli3+/− (C,G,K), and Gli2−/−;Gli3−/− (D,H,L) mouse embryos using a DIG-labelled Myf5 probe, anti-Myf5 (red), anti pan-laminin (red) and anti-myogenin (green) antibodies. (A,E) Myf5-expressing cells populate the myotome as thin layer underneath the dermomyotome in wild-type embryos. (B,F) Gli2+/−;Gli3−/− embryos display little defect in myotome formation. By contrast, Gli2−/−;Gli3+/− (C,G) and Gli2−/−;Gli3−/− (D,H) embryos have abnormally patterned myotomal cells (white arrows). (I) A near-continuous basement membrane forms underneath the myotome (white arrows) and on the basal side of the dermomyotome (yellow arrowhead) in wild-type embryos. (J) In Gli2+/−;Gli3−/− somites, no myotomal basement membrane defect is observed, although gaps are visible (white arrows). By contrast, Gli2−/−;Gli3+/− (K) and Gli2−/−;Gli3−/− (L) display a severe phenotype that resembles Shh−/− somites, with no myotomal basement membrane forming, although short fragments of assembled Laminin are associated with myogenin-positive cells (white arrows). Note the presence of myogenin-positive cells in the ventral somite. Magnification: ×400 (left panels) and ×630 (right panels).
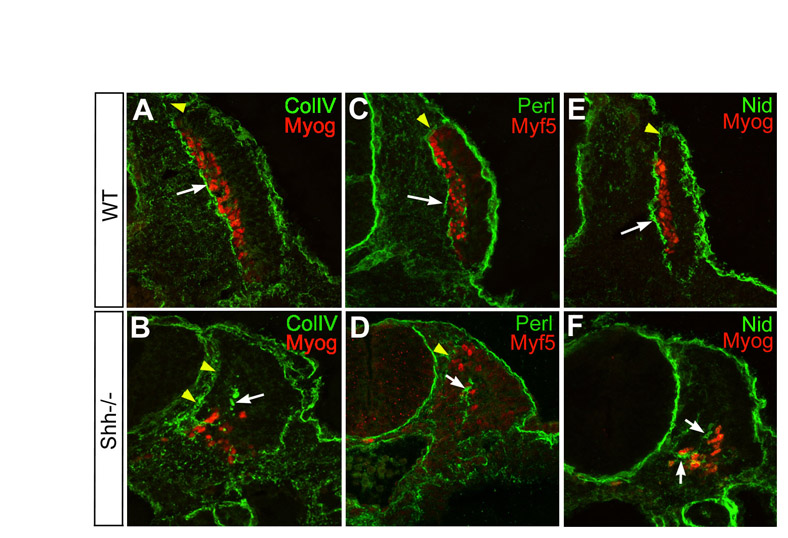
Fig. S2. Extracellular matrix components are synthesised and secreted in Shh−/−embryos. (A-F) Collagen IV (A,B), perlecan (C,D) and nidogen (E,F) protein expression (in green) in relation to Myf5 (C,D) or myogenin (A,B,E,F) distribution (in red) was examined by immunohistochemistry and confocal imaging in forelimb somites of E9.5 wild-type (A,C,E), Shh−/− (B,D,F) embryos. (A,C,E) In wild-type somites, collagen IV, perlecan and nidogen have incorporated into the assembled myotomal basement membrane (white arrows) that underlies the myotome containing Myf5 and myogenin-positive cells. (B,D,F) By contrast, collagen IV, perlecan and nidogen fail to assemble into a continuous sheet around myogenin- and Myf5-positive myotomal cells, although the presence of clumps indicates that they are efficiently synthesised and secreted (white arrows). Note also that the dermomyotomal basement membrane forms normally (yellow arrowheads). Magnification: ×400.
Fig. S3. Expression of the laminin receptors α6β1 integrin and dystroglycan in Gli compound mutant embryos. α6β1 integrin (green), Pax3 (red), β-dystroglycan (red) and Myf5 (green) protein distribution was examined by immunohistochemistry and confocal imaging in forelimb somites of E9.5 Gli2+/−;Gli3−/− and Gli2−/−;Gli3+/− embryos. α6β1 Integrin and β-dystroglycan are apparently expressed normally in Gli2+/−;Gli3−/−, although the presence of Pax3-positive cells (red) in the myotomal compartment indicates some level of disruption in the myotomal basement membrane function. In Gli2−/−;Gli3+/− embryos, α6β1 integrin and β-dystroglycan remain expressed at the surface of myotomal cells. However, myotomal cells begin to display aberrant migratory behaviour and there is an increased number of Pax3-positive cells in the myotome compartment. Magnification: ×400.
Fig. S4. The laminin receptors α6β1 integrin and dystroglycan are active in Shh−/−embryos. (A-D) α-dystroglycan (A,B) and active β1-integrin(CD29) (C,D) protein distribution in relation to Myf5 expression was examined by immunohistochemistry in E9.5 wild-type (A,C) and Shh−/− (B,D) embryos. (A) α-dystroglycan (red) is expressed at the cell surface of wild-type Myf5-positive myotomal cells (red arrow). (B) Myf5-positive cells (green) maintain α-dystroglycan (red) expression in Shh−/− somites. (C) Myf5-positive myotomal cells (green) are CD29 positive (red) indicating activity of integrins. Note that CD29 is detected at higher levels in cells located deeper in the myotome (red arrow). (D) Despite their mispatterning, Myf5-positive cells are labelled with the CD29 antibody (red arrow). Magnification: ×400.
Fig. S5. Partial recovery of basement membrane assembly in Shh−/−;Gli3−/−embryos. (A-H) β1 laminin subunit (green) and Myf5 (red) protein distribution was examined by immunohistochemistry and confocal imaging in interlimb and forelimb somites of E9.5 Shh−/− (A,B), Shh−/−;Gli3+/− (C,D), Shh−/−;Gli3−/− (E,F) and wild-type (G,H) embryos. (A,B) Although secreted, laminin fails to assemble into a continuous basement membrane, causing a ventral migration of Myf5-expressing MPCs (red arrow) at all axial levels of Shh−/− embryos. (CD) Despite, the partial recovery of Myf5 expression, no basement membrane assembly is observed at interlimb or forelimb levels in Shh−/−;Gli3+/− embryos. (E,F) Likewise, no basement membrane is observed at interlimb levels in Shh−/−;Gli3−/− embryos. However, a discontinuous basement membrane is present at forelimb level (F) in Shh−/−;Gli3−/− somites (white arrows), indicating that a partial recovery is occurring. Aberrant ventral migration of MPCs remains visible in interlimb and forelimb somites (E,F, red arrows). Note the concomitant recovery of epaxial Myf5 activation at interlimb and forelimb levels (white asterisk). (G,H) E9.5 wild-type embryos display a fully assembled myotomal basement membrane at forelimb, but not interlimb level. Yellow arrowheads indicate the dermomyotomal basement membrane. (I,J) Lama1 expression remains defective in Shh−/−;Gli3+/− embryos, although reduced expression can be observed in forelimb somites (black arrows). Compare neural tube and pro-mesonephros staining in J with Fig. 5A′ and 5E′. Magnification: ×400.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}