Files in this Data Supplement:
Fig. S1. Expression and purification of recombinant Sec61βOPG. (A) Sequences of tail-anchored protein derivatives used in this study. Predicated transmembrane domains are highlighted in yellow and the opsin N-glycosylation tag derived from bovine opsin (OPG) is in green. For Sec61βOPG, an endogenous cysteine at residue 39 was mutated to a serine and an endogenous serine at residue 77 was replaced with a cysteine whilst for cytochrome b5 (Cytb5) Ser119 was altered to a cysteine (all changes shown in red font). (B) Sec61βOPG in pHisTrx vector was expressed in E. coli and the resulting fusion protein purified on NiNTA agarose before removal of the HisTrx tag using thrombin. The purified protein was resolved on SDS-PAGE and stained with Coomassie Brilliant Blue, full-length and N-terminally truncated forms of Sec61βOPG are indicated. A minor contaminant corresponding to E. coli DNAK is also identified (•), the removal of this component by an additional step of ion exchange chromatography did not alter the membrane integration properties of the resulting recombinant protein preparation (our unpublished data). (C) The ability of recombinant Sec61βOPG to integrate into sheep pancreatic microsomes in the presence of rabbit reticulocyte lysate was determined using N-glycosylation of the C-terminal OPG tag. N-glycosylation was confirmed by EndoH digestion, and the N-glycosylated (+gly), non-glycosylated (-gly) and truncated forms (trunc.) of Sec61βOPG are shown.
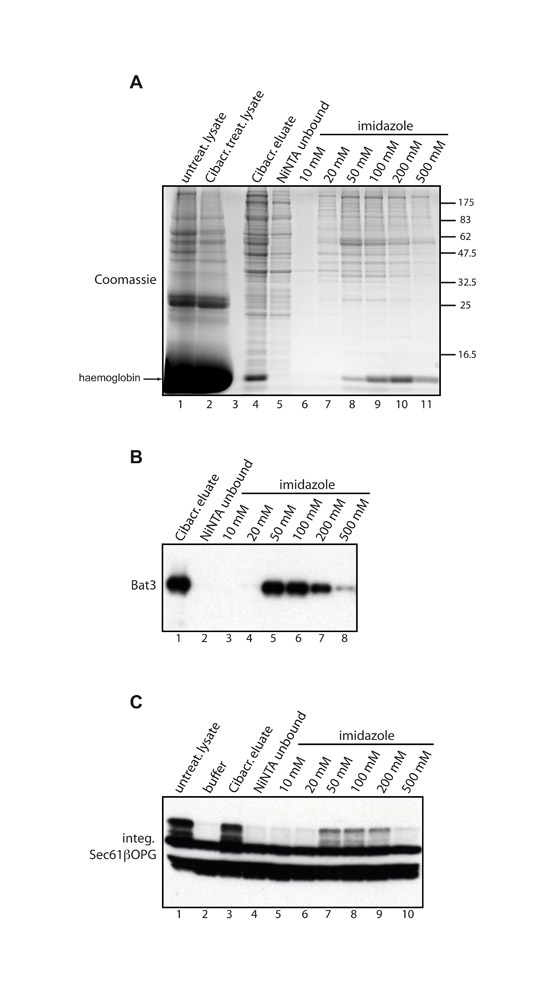
Fig. S2. NiNTA agarose binding profile of the Cibacron-eluted material. Cibacron Blue agarose (600 l) was incubated with 5 ml rabbit reticulocyte lysate for 2 hours at 4°C, the beads were then extensively washed and the bound proteins eluted with 1.5 M NaCl (480 l). The eluate was desalted, concentration of imidazole adjusted to 5 mM and the resulting protein solution incubated with NiNTA agarose (100 l bead volume). The unbound fraction was collected, beads washed and the bound proteins eluted using a step-wise gradient of increasing imidazole concentrations from 10 mM to 500 mM. After desalting, 5 l lysates and 10 l of each fraction were run on an SDS-PAGE gel that was later stained with Coomassie Brilliant Blue (A). In parallel, 2 l from each fraction were used for immunoblotting with anti-Bat3 antibody (B). The capacity of each fraction to restore the integration competence of the Cibacron-treated lysate was tested (C) by adding 5 l buffer R as a control (lane 2), or 5 l of each of the fractions indicated (lanes 3-10), to 20 l of the Cibacron-treated lysate and then carrying out a Sec61βOPG integration reaction and visualising the results by immunoblotting with anti-opsin antibody (cf. Figs 3 and 4).
Fig. S3. Immunofluorescence analysis of Bat3 and GFP-Sed5 in S. cerevisiae. The subcellular localisation of full-length Bat3 (A) and an N-terminal fragment of Bat3 (B) (cf. Fig. 6A) was determined by immunofluorescence microscopy in Δmdy2 (Δget5) strain and compared with DAPI staining of the nucleus. (C) The subcellular localisations of full-length Bat3 and GFP-Sed5 were determined by immunofluorescence microscopy in wild-type and Δmdy2 (Δget5) cells that were also stained with DAPI to reveal the nucleus. (D) The subcellular localisation of a Bat3 ΔNLS mutant was determined as described for A and B. (E) Immunoblot confirming efficient expression of the Bat3 ΔNLS mutant in Δmdy2 (Δget5) S. cerevisiae cells.
{kind=link}
{kind=link}
{kind=link}