Files in this Data Supplement:
Table S1. Strains used and attachment at cytokinesis.
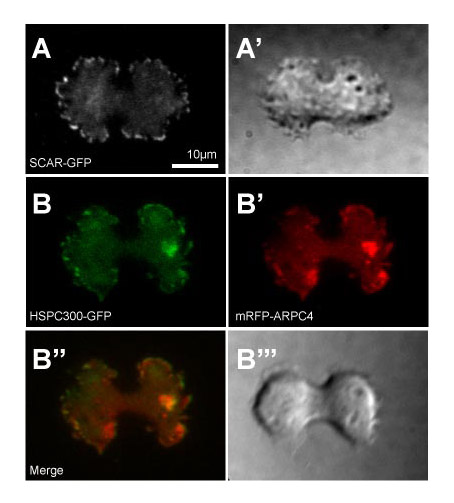
Fig. S1. Localisation of different SCAR complex GFP probes at cytokinesis. (A) SCAR-GFP single expression and (B) HSPC300-GFP, (B′) mRFP-ARPC4 coexpression in Ax3 cells. Images were taken by TIRF microscopy. A′ and B′′′ are the DIC images of the respective cells taken at the same time. Scale bar: 10 m.
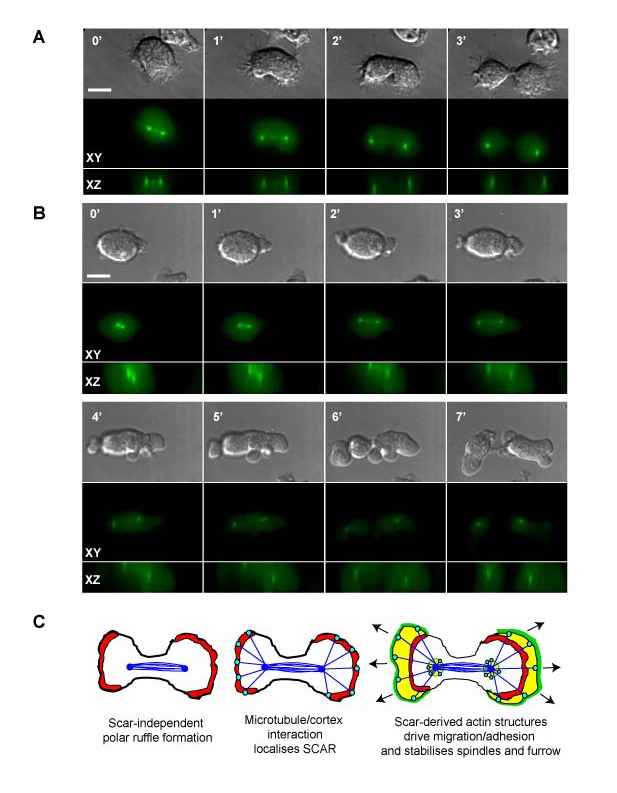
Fig. S2. 3D analysis of spindle orientation in (A) Ax3 and (B) IR46 cells expressing GFP-tubulin. A Z-series of images was captured at each time point. The images were then reconstructed to give both an extended focus (XY) image (looking down onto each cell) as well as a vertical (XZ) section through the cell along the plane of the spindle. Scale bar: 10 m. (C) Model of the processes involved in Dictyostelium cytokinesis and how they might affect the spindle orientation. Blue lines depict microtubules; red indicates SCAR-independent actin polymerisation; green indicates SCAR complex localisation/activity and yellow the resultant actin.
Fig. S3. Blebbistatin treatment of SCAR complex mutants. Cells were grown in either 40 M blebbistatin or DMSO control for 3 days and then fixed, stained with DAPI, and the number of nuclei per cell counted. At least 100 cells were counted for each sample, and the values plotted are the means ± standard deviation of three independent experiments. Hspc300, napA, pir121 and abi mutant cells were all generated in an Ax3 background and the results for scar-null cells in an Ax2 background are shown here. *P<0.005 (Student’s t-test)
Fig. S4. Generation of an inducible scar/mhcA double mutant. Western blot of control (Ax3), scar-null (IR46) and IR46 cells expressing SCAR under the control of the Tet-On promoter. IR46::pJSK321 cells were grown overnight in the concentrations of doxycycline indicated. Equal quantities of total cell lysates were probed with a rabbit anti-SCAR antibody.
Fig. S5. SCAR is essential for traction-mediated cytofission. Multinucleate control (Ax3) and scar-null (IR46) cells were generated by growth in shaking culture for 48 hour in the presence of 40 M blebbistatin. Cells were then seeded onto glass coverslips, allowed to adhere and blebbistatin washed out by three successive changes of medium. The division of these multinucleate cells was then recorded by timelapse microscopy (A and B) Scale bar: 50 m. This was quantified by counting the number of individual cells within the field of view and normalising to the initial number at the time points shown. Values plotted in B are the means ± standard deviations of three independent experiments.
Movie 1. Localisation of HSPC300-GFP and mRFP-ARPC4 in dividing Ax3 (wild-type) Dictyostelium cells. Images taken by TIRF.
Movie 2. Localisation of HSPC300-GFP and mRFP-actin in dividing Ax3 (wild-type) Dictyostelium cells. Images taken by TIRF.
Movie 3. Colocalisation of HSPC300-GFP to MiDASes. Ax3 cells coexpressing HSPC300-GFP and mRFP-actin were imaged by TIRF microscopy. Note the transient actin-rich MiDASes in the centre of each daughter cell.
Movie 4. Morphology and movement of control (Ax3) Dictyostelium cells undergoing cytokinesis
Movie 5. Morphology and movement of scar-null (IR46) Dictyostelium cells undergoing cytokinesis.
Movie 6. Morphology of control Drosophila cells (labelled with GFP-Moesin) dividing within a wild-type tissue. Movie shows a repeated series of Z-sections through the tissue.
Movie 7. Morphology of scar-null Drosophila cells (labelled with GFP-Moesin) dividing within a wild-type tissue. Movie shows a repeated series of Z-sections through the tissue.
Movie 8. Localisation of HSPC300-GFP and mRFP-actin in dividing JK107 (mhcA-null) cells expressing SCAR.
Movie 9. Localisation of HSPC300-GFP and mRFP-actin in dividing JK107 (mhcA-null) cells with SCAR expression repressed.
Movie 10. Localisation of HSPC300-GFP and mRFP-actin in dividing eb1-null cells.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}