Files in this Data Supplement:
Fig. S1. Heightened levels of fn1 expression in han mutants. (A,B) In situ hybridization for myl7 illustrates the han mutant phenotype at the stage when embryos were collected for RNA extraction and microarray analysis. Dorsal views, anterior up. Scale bar: 50 µm. (A) Wild-type cardiomyocytes have initiated cardiac fusion with contacts between posterior subsets of contralateral cells and are proceeding to complete formation of the cardiac cone with the merger of anterior subsets of contralateral cells. (B) By contrast, han mutant embryos exhibit two separated populations of cardiomyocytes. (C,D) Bar charts indicating levels of gene expression, relative gapdh, as detected by qRT-PCR in wild-type and han mutant embryos. Values represent the mean (±s.e.m.) from triplicate samples. (C) Microarray analysis revealed a 2.0-fold upregulation of fn1 in han mutants (see Table S1), and validation of this result by qRT-PCR indicated that fn1 expression is increased 1.6-fold in han mutants. (D) Expression of hand2 was undetectable by qRT-PCR in han mutants, whereas microarray analysis indicated a 5.3-fold downregulation of hand2 in han mutants (see Table S1). PCR primers used were: fn1, 5′-GAATCCCCGAGCCAATCAA-3′ and 5′-TTTTATGTTTCCCCGTTCCTT-3′; hand2, 5′-TGCGCTCGAATCAACAAATG-3′ and 5′-TATACCCTACGGCGAACCACA-3′; gapdh, 5′-CCCCACCCCCAATGTCTCT-3′ and 5′-AACCTGGTGCTCCGTGTATCC-3′.
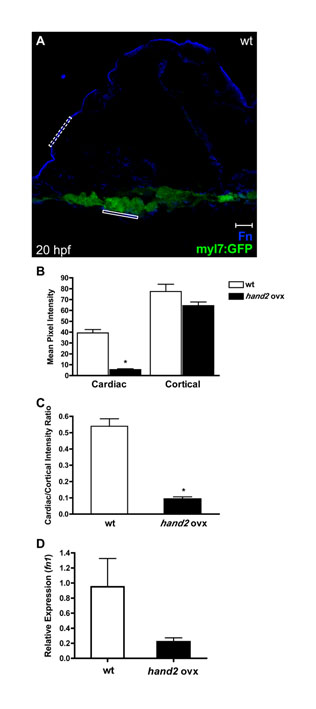
Fig. S2. Quantification confirms reduced Fn deposition basal to the myocardium in embryos overexpressing hand2. (A) Representative transverse confocal section in an embryo expressing Tg(myl7:egfp) (green) exemplifies the method for quantification of Fn immunofluorescence (blue). Scale bar: 10 µm. For each image, we used ImageJ software to select a cardiac region of interest (r.o.i.) (solid rectangle) basal to a group of at least three cardiomyocytes and then measured the mean pixel intensity of the r.o.i. in the blue channel. Similarly, we measured the mean pixel intensity in a comparably sized r.o.i. (dashed rectangle) along the exterior cortex of the embryo. Measurements were taken from three consecutive confocal images per cryosection in three different cryosections per embryo, and at least three embryos were sampled for each condition. (B) Bar chart comparing the levels of Fn deposition in terms of mean pixel intensity (±s.e.m.) for each r.o.i. measured. Asterisk indicates statistically significant difference from wild type (P<0.0001, two-tailed unpaired Student�s t-test). In embryos overexpressing hand2, deposition of Fn adjacent to the cardiomyocytes is reduced ∼85%, whereas cortical deposition of Fn is relatively unaffected. (C) Bar chart comparing levels of Fn deposition adjacent to the cardiomyocytes, normalized relative to cortical Fn levels. Values represent the mean ratio (±s.e.m.) for each condition; asterisk indicates statistically significant difference from wild type (P<0.0001, two-tailed unpaired Student�s t-test). (D) Bar chart indicating levels of fn1 expression, relative to hprt expression, as detected by qRT-PCR in wild-type embryos and in embryos overexpressing hand2. Values represent the mean (±s.e.m.) from triplicate samples. fn1 expression is decreased 4-fold in embryos overexpressing hand2. PCR primers used were: fn1, 5′-GAATCCCCGAGCCAATCAA-3′ and 5′-TTTTATGTTTCCCCGTTCCTT-3′; hprt, 5′-ATCCGCCTCAAGAGTTACCAA-3′ and 5′-TCTAGCAGCGTTTTCATCGTT-3′.
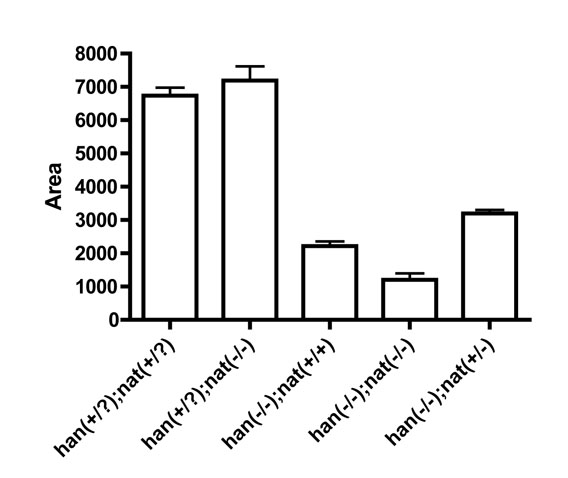
Fig. S3. Reduction of fn1 function slightly improves cardiomyocyte production in hand2 mutants. Bar chart comparing the production of cardiomyocytes in embryos of the indicated genotypes, as represented by the area of myl7 expression at 20 hpf. Values represent the mean area (±s.e.m.) of five han−/−;nat−/− embryos and at least ten embryos from each other category, collected from at least three independent clutches. Areas were measured using ImageJ as previously described (Marques et al., 2008). According to Tukey�s multiple comparison test following a significant ANOVA (P<0.0001), all categories exhibit significant differences from wild type (P<0.001), except for han+/?;nat−/− (P>0.05). Relative to han−/−;nat+/+, all categories exhibit significant differences (P<0.001), including han−/−;nat+/− (P<0.05), but excepting han−/−;nat−/− (P>0.05). Although statistically significant, the improvement of cardiomyocyte production in han−/−;nat+/− embryos is relatively modest and does not approach restoration of wild-type cardiomyocyte production. It is important to note that the assessment of cardiomyocyte production via measurement of the area of myl7 expression does not take into account the three-dimensional stacking of cardiomyocytes. However, we observe similar stacking in wild-type and han−/−;nat+/− embryos: examination of transverse sections similar to those shown in Figs 2 and 4 reveals that cardiomyocytes are stacked 1 to 2 tiers deep in both cases (n=5 of each genotype). Therefore, it is reasonable to compare the areas of myl7 expression in these two contexts to reach the conclusion that cardiomyocyte production is not rescued in han−/−;nat+/− embryos. By contrast, examination of transverse sections indicates that cardiomyocytes are stacked 1 to 3 tiers deep in han mutant embryos and 2 or more tiers deep in nat mutant and han−/−;nat−/− embryos (n≥4 of each genotype), leaving open the possibility that we are underestimating cardiomyocyte production when measuring areas of myl7 expression in these cases.
Marques, S. R., Lee, Y., Poss, K. D. and Yelon, D. (2008). Reiterative roles for FGF signaling in the establishment of size and proportion of the zebrafish heart. Dev. Biol.321, 397-406.
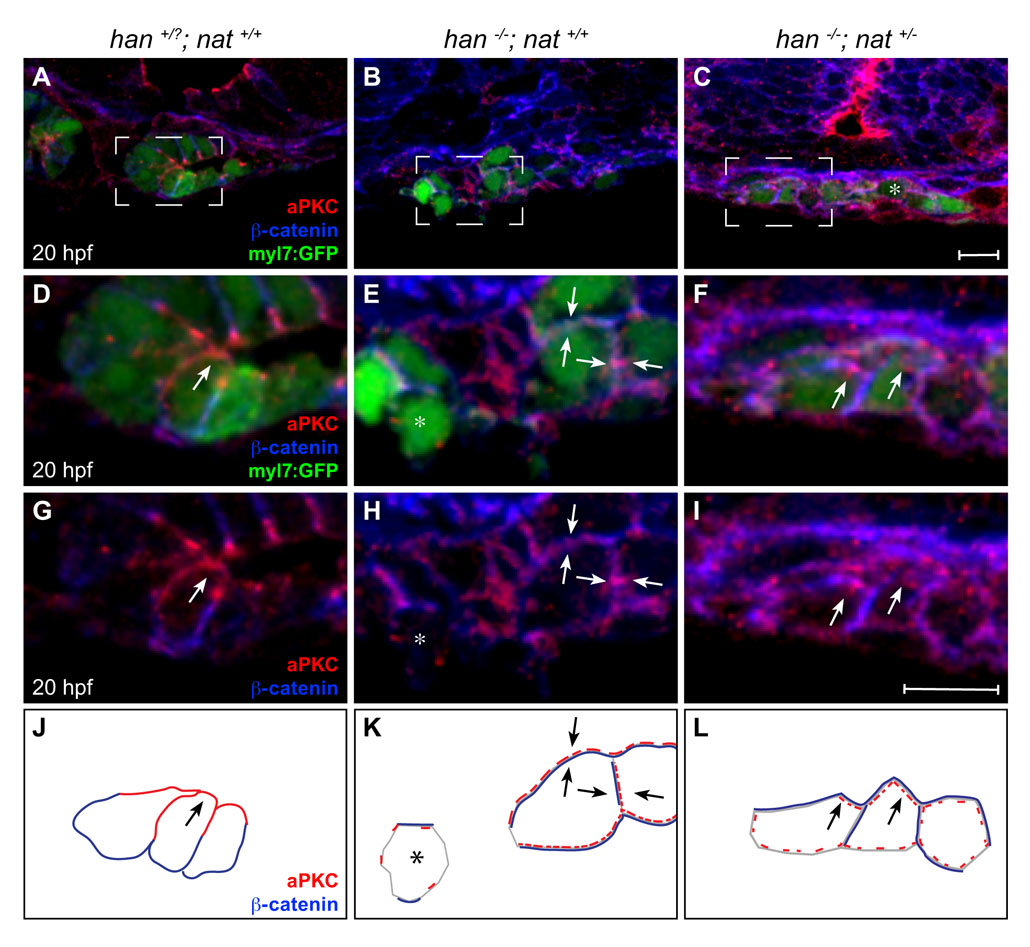
Fig. S4. Poor rescue of myocardial apicobasal polarity in han−/−;nat+/− embryos. (A-I) Transverse confocal sections of the right lateral mesoderm in embryos expressing Tg(myl7:egfp) (green). Dorsal is up. Immunofluorescence detects aPKC (red) and β-catenin (blue). Boxed regions in A-C are shown at higher magnification in D-I. At this stage, the wild-type myocardium has begun the asymmetric involution that displaces a medial portion of the myocardial epithelium dorsally (Rohr et al., 2008). Scale bars: 10 µm. (J-L) Cartoons schematize cardiomyocyte-associated localization of aPKC (red) and β-catenin (blue) shown in D-I. Wild-type cardiomyocytes are organized in a cohesive layer (A) and exhibit apical enrichment of aPKC (D,G,J, arrow) and basolateral enrichment of β-catenin (D,G,J). In han embryos, the small clusters of cardiomyocytes (B) exhibit irregular localization of aPKC and β-catenin, either diffusely around the cell (E,H,K, arrows) or nearly absent (E,H,K, asterisk). Only slight improvements are seen in han−/−;nat+/− embryos: aPKC is either distributed diffusely (C, asterisk) or partially enriched apically (F,I,L, arrows), and β-catenin is either distributed diffusely or partially enriched laterally (F,I,L). We note that previous work indicated an absence of aPKC localization in han mutant cardiomyocytes (Trinh et al., 2005). We suspect that the diffuse and irregular aPKC localization observed here (E,H,K) reflects increased sensitivity of our current protocol, which uses a different fixation method and thinner sections.
Rohr, S., Otten, C. and Abdelilah-Seyfried, S. (2008). Asymmetric involution of the myocardial field drives heart tube formation in zebrafish. Circ. Res.102, e12-e19.
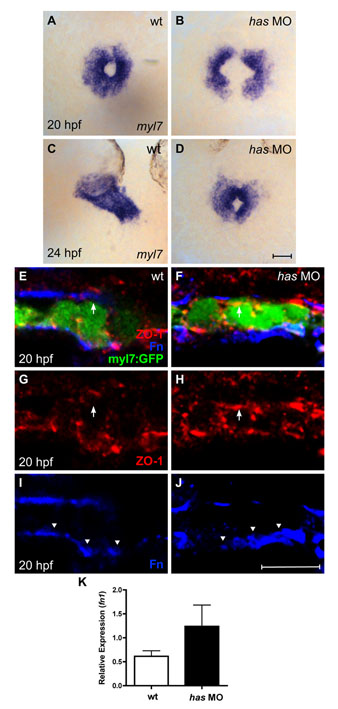
Fig. S5. Loss of apkci function does not disrupt basal deposition of Fn. (A-D) In situ hybridization for myl7 comparing embryos injected with an anti-apkci morpholino (has MO) with their wild-type siblings. Dorsal views, anterior up. Scale bar: 50 µm. Although cardiac fusion proceeds in embryos lacking apkci function, heart tube extension is arrested (Horne-Badovinac et al., 2001; Peterson et al., 2001; Rohr et al., 2006). (E-J) Transverse confocal sections of the left lateral mesoderm in embryos expressing Tg(myl7:egfp) (green). Dorsal is up. Immunofluorescence detects ZO-1 (red) and Fn (blue). Scale bar: 10 µm. Wild-type cardiomyocytes exhibit lateral localization of ZO-1 (E,G) and basal deposition of Fn (E,I, arrowheads). By contrast, has morphant cardiomyocytes exhibit defective apicobasal polarity (Rohr et al., 2006): for example, ZO-1 is diffusely localized (F,H), including localization in the apical domain, where it is not found in wild-type cardiomyocytes (E-H, arrows). Despite the disruption of some aspects of apicobasal polarity, Fn deposition remains restricted to the basal domain of has morphant cardiomyocytes (F,J, arrowheads). Altogether, the has morphant phenotype shares many similarities with the han−/−;nat+/− phenotype. In each case, cardiac fusion proceeds, but heart tube extension arrests; additionally, the myocardium is a relatively aligned layer that exhibits basal Fn deposition despite its other apicobasal polarity defects. (K) Bar chart indicates levels of fn1 expression, relative to hprt expression, as detected by qRT-PCR in wild-type and has morphant embryos. Values represent the mean (±s.e.m.) from triplicate samples. fn1 expression is increased 2-fold in has morphants, comparable to the increase observed in han mutants (see Fig. S1C). However, our qRT-PCR data monitor fn1 expression in whole embryos, rather than in the specific locations relevant to cardiac fusion. In this regard, it is notable that the global increase in fn1 expression in has morphants does not block cardiac fusion (D) and that these embryos also exhibit normal basal deposition of Fn (J). PCR primers used were: fn1, 5′-GAATCCCCGAGCCAATCAA-3′ and 5′-TTTTATGTTTCCCCGTTCCTT-3′; hprt, 5′-ATCCGCCTCAAGAGTTACCAA-3′ and 5′-TCTAGCAGCGTTTTCATCGTT-3′.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}