

Abstract
Human embryonic stem cells (hESCs) offer a platform to bridge what we have learned from animal studies to human biology. Using oligodendrocyte differentiation as a model system, we show that sonic hedgehog (SHH)-dependent sequential activation of the transcription factors OLIG2, NKX2.2 and SOX10 is required for sequential specification of ventral spinal OLIG2-expressing progenitors, pre-oligodendrocyte precursor cells (pre-OPCs) and OPCs from hESC-derived neuroepithelia, indicating that a conserved transcriptional network underlies OPC specification in human as in other vertebrates. However, the transition from pre-OPCs to OPCs is protracted. FGF2, which promotes mouse OPC generation, inhibits the transition of pre-OPCs to OPCs by repressing SHH-dependent co-expression of OLIG2 and NKX2.2. Thus, despite the conservation of a similar transcriptional network across vertebrates, human stem/progenitor cells may respond differently to those of other vertebrates to certain extrinsic factors.
Keywords: Glia, Myelination, Neuron-glial switch, Purmorphamine, Transplantation
Introduction
Oligodendrocytes, the myelinating glia in the central nervous system (CNS), are differentiated chiefly from neuroepithelial (NE) cells in the ventral neural tube. These NE cells express the helix-loop-helix transcription factor OLIG2 in response to ventrally derived sonic hedgehog (SHH) (Lu et al., 2000; Zhou et al., 2000). In the spinal cord, the OLIG2-expressing progenitors give rise to motoneurons during the neurogenic phase. Thereafter, OLIG2-expressing progenitors downregulate neurogenic transcription factors such as neurogenin 2 (NGN2) and PAX6, begin to express the oligodendroglial transcription factors NKX2.2 and SOX10 (Lee et al., 2005; Marquardt and Pfaff, 2001; Qi et al., 2001; Stolt et al., 2004; Sugimori et al., 2007; Zhou et al., 2001), and become migratory oligodendrocyte precursor cells (OPCs) that also express the surface markers platelet-derived growth factor receptor alpha (PDGFRα) and membrane proteoglycan NG2 [also known as chrondroitin sulfate proteoglycan 4 (CSPG4)]. In addition to the ventral source, a smaller population of oligodendrocytes is generated independently of SHH from the dorsal neural tube (Cai et al., 2005; Fogarty et al., 2005; Vallstedt et al., 2005). Nevertheless, expression of OLIG2 is a prerequisite step for the dorsal precursor cells to become OPCs (Cai et al., 2005). OLIG2 is expressed in human OPCs, as shown in limited studies on human fetal brain tissues (Jakovcevski and Zecevic, 2005). Whether OLIG2 expression is necessary for human OPC specification and how neural progenitors differentiate into OPCs in humans remain elusive.
Embryonic stem cells (ESCs), isolated from a blastocyst embryo, can differentiate into all cell lineages of the organism (Evans and Kaufman, 1981; Thomson et al., 1998). They thus offer an in vitro model system for studying early mammalian development, including oligodendrocyte specification. As in the developing mouse neural tube, SHH induces OLIG2 expression and promotes the generation of OPCs from mouse ESC (mESC)-derived neural progenitors (Billon et al., 2002). The SHH-induced OPC differentiation from mESCs in vitro retains the correct timing seen in embryonic development (Billon et al., 2002; Du et al., 2006; Samanta and Kessler, 2004), which may be accounted for by activation of the intrinsic transcriptional networks. Hence, the nature of in vitro OPC differentiation from mESCs is consistent with what has been learned from in vivo development. Nevertheless, insight into the molecular mechanism underlying OPC specification in vertebrates and the ease of OPC generation from mESCs have not yet been translated to human primates. Several groups have reported the production of oligodendrocytes from hESCs (Izrael et al., 2007; Kang et al., 2007; Nistor et al., 2005); however, in none of the reports was SHH used for differentiation nor SHH signaling analyzed. It is not known whether the SHH-dependent transcriptional network is required for human OPC specification, as in other vertebrates.
Besides SHH signaling, fibroblast growth factor 2 (FGF2) promotes OPC generation from rodent neural stem/progenitor cells (Avellana-Adalid et al., 1996; Zhang et al., 1998). This effect may be SHH-dependent (Gabay et al., 2003) or -independent (Chandran et al., 2003; Kessaris et al., 2004). Similar to OPC differentiation from rodent neural progenitors, OPCs can be efficiently differentiated from mESCs in response to FGF2 followed by PDGF (Brustle et al., 1999). However, attempts from several independent laboratories to generate OPCs from FGF2-expanded human neural stem/progenitor cells using similar approaches as for their mouse counterparts have been consistently unsuccessful (Chandran et al., 2004; Roy et al., 1999; Zhang et al., 2000). Similarly, hESC-derived neural progenitors, following expansion in the presence of FGF2 or EGF, appear to give rise to OPCs at low efficiency (Izrael et al., 2007; Kang et al., 2007). It seems that stem/progenitor cells from human and rodents respond to FGF2 differently in oligodendrocyte differentiation.
Using a chemically defined oligodendrocyte differentiation system, we present evidence that SHH-dependent induction of OLIG2, co-expression of OLIG2 and NKX2.2, and activation of SOX10 and PDGFRα, are essential for OPC specification from hESCs, indicating that a conserved transcriptional network underlies OPC specification in human as in other vertebrates. The transition from the OLIG2 and NKX2.2-expressing pre-OPCs to OPCs in human is a uniquely protracted process. FGF2 blocks the transition of pre-OPCs to OPCs by disrupting SHH-dependent co-expression of OLIG2 and NKX2.2.
Materials and Methods
Maintenance and oligodendrocyte differentiation of hESCs
hESCs (line H9, passage 21 to 40; H1, passage 25 to 35) were maintained as described (Thomson et al., 1998). Differentiated colonies were marked and removed physically with a pipette tip and the stem cell state was monitored by expression of OCT4 (POU5F1) and SSEA4 (Pankratz et al., 2007). hESCs were differentiated to primitive NE in an adherent colony culture for 10 days in a neural induction medium consisting of DMEM/F12, N2 supplement and non-essential amino acids (Invitrogen, Carlsbad, CA, USA), as detailed elsewhere (Pankratz et al., 2007; Zhang et al., 2001). To pattern the primitive NE to ventral spinal progenitors, differentiation cultures at day 10 were treated with retinoic acid (RA, 100 nM; Sigma, St Louis, MO, USA) followed by RA and SHH (100 ng/ml; R&D Systems, Minneapolis, MN, USA) (Li et al., 2005) or purmorphamine (1 μM; Calbiochem, San Diego, CA, USA) (Li et al., 2008; Sinha and Chen, 2006) at day 14 for 2 weeks. NE cells grew rapidly and formed neural tube-like rosettes at day 14-17 of differentiation. The NE colonies were gently blown off the surface using a pipette and grown as floating clusters in suspension in the same medium. OLIG2-expressing neural progenitors usually appear at ∼day 24 and differentiate to motoneurons at the fifth week (day 28-35) (Li et al., 2005). At day 35, the culture was switched to a glial differentiation medium consisting of DMEM, N1 supplement, T3 (60 ng/ml), biotin (100 ng/ml) and cAMP (1 μM) (all from Sigma). A cocktail of cytokines consisting of the PDGF-AA isoform, insulin-like growth factor 1 (IGF1) and neurotrophin 3 (NT3; NTF3) (all at 10 ng/ml from R&D Systems) were added to promote the expansion of the progenitor population. The cultures were fed every other day and the progenitor clusters disaggregated into smaller cell aggregates with a polished glass Pasteur pipette every week. FGF2 (10 ng/ml), EGF (10 ng/ml), noggin (100 ng/ml) (all from R&D Systems), SHH or purmorphamine was added to examine their effect on motoneuron differentiation, progenitor proliferation and OPC specification. To differentiate OPCs to oligodendrocytes, progenitor clusters were incubated in ACCUTASE (Innovative Cell Technologies, San Diego, CA, USA) at 37°C for 5 minutes. After removal of ACCUTASE, the loosened clusters were triturated and plated onto glass coverslips coated with polyornithine and laminin in the glial differentiation medium without SHH/purmorphamine. The concentration of PDGF-AA, IGF1 and NT3 was 5 ng/ml each.
Lentivirus-mediated RNAi transfection
OLIG2 and non-silencing control RNAi constructs were made using the pGIPZ vector (Open Biosystems, Huntsville, AL, USA). Lentivirus was produced by co-transfecting the pGIPZ RNAi plasmid and packaging plasmid into 293T cells by the calcium-phosphate method, as described (Xia et al., 2008). Neural progenitors at day 35 were transfected by directly adding the concentrated lentivirus to the cell cultures at a multiplicity of infection of 10.
RNA extraction and RT-PCR
Total RNA was extracted from cells using Trizol reagent (Invitrogen) and reverse-transcribed using the SuperScript III First-Strand Synthesis System (Invitrogen). RT-PCR was performed as described (Pankratz et al., 2007). Primers are listed in Table 1.
Table 1. Primers (5′ to 3′) for real-time PCR.
| Human gene | Forward | Reverse |
|---|---|---|
| GAPDH | TCGACAGTCAGCCGCATCTTCTTT | ACCAAATCCGTTGACTCCGACCTT |
| GLI1 | GGCTGCAGTAAAGCCTTCAGCAAT | TGCAGCCAGGGAGCTTACATACAT |
| GLI2 | AGGACAAGGCCAACATCGTCATCT | TCCCTGGCTTTCATTTGCAGTTGG |
| GLI3 | TTGCACAAAGGCCTACTCGAGACT | CTTGTTGCAACCTTCGTGCTCACA |
| NKX2.2 | GACAACTGGTGGCAGATTTCGCTT | AGCCACAAAGAAAGGAGTTGGACC |
| OLIG2 | GGTAAGTGCGCAATGCTAAGCTGT | TACAAAGCCCAGTTTGCAACGCAG |
| SHH | AAGATCTCCAGAAACTCCGAGCGA | CGAGATGGCCAAAGCGTTCAACTT |
Immunocytochemistry and microscopy
Immunofluorescence on coverslip cultures and free-floating brain sections was described previously (Pankratz et al., 2007; Zhang et al., 2001). The primary antibodies used are listed in Table 2. The immunostained samples were visualized with a Nikon TE600 fluorescence microscope (Nikon Instruments, Melville, NY, USA) fitted with a SPOT camera (Diagnostic Instruments, Sterling Heights, MI, USA) or with a Nikon C1 laser-scanning confocal microscope. For counting immunostained cells, fields of fixed area were randomly selected from biological replicates using ImageJ (W. Rasband, NIMH, Bethesda, MD, USA). Statistical analyses were performed using one-way ANOVA (Tukey or Dunnett for multiple comparisons) in the R environment (R Development Core Team, Vienna, Austria).
Table 2. Antibodies.
| Antibody | Isotype | Dilution | Source | Cat. # |
|---|---|---|---|---|
| OLIG2 | Goat IgG | 1:500 | Santa Cruz | sc-19969 |
| NKX2.2 | Mouse IgG | 1:50 | DSHB, Iowa City, IA | 74.5A5 |
| βIII-tubulin | Mouse IgG | 1:5000 | Sigma | T 8660 |
| GFAP | Rabbit IgG | 1:5000 | DAKO | Z0344 |
| NG2 | Mouse IgG | 1:400 | BD Pharmingen | 554275 |
| HB9 | Mouse IgG | 1:50 | DSHB, Iowa City, IA | 81.5C10 |
| PDGFRα | Rabbit IgG | 1:1000 | Gift from Dr William Stallcup | |
| O4 | Mouse IgM | 1:50 | Chemicon | MAB345 |
| MBP | Rabbit IgG | 1:500 | Chemicon | AB5622 |
| Human nuclei | Mouse IgG | 1:200 | Chemicon | MAB1281 |
Fluorescence-activated cell sorting (FACS)
Single-cell suspensions were fixed and stained as described (Pankratz et al., 2007). FACS was performed with a FACSCalibur flow cytometer and analyzed using CellQuest Pro (BD Biosciences, San Diego, CA, USA).
Transplantation of OPCs into the shiverer mouse brain and tissue preparation
Animal experiments were performed following protocols approved by the Animal Care and Use Committees at the University of Wisconsin-Madison. Progenitors enriched for OPCs after 10-12 weeks of hESC differentiation were dissociated with ACCUTASE and prepared in artificial cerebral spinal fluid at 50,000 cells/μl. About 2 μl of cell suspension (100,000 cells in total) was transplanted into the lateral ventricle of a newborn shiverer mouse with a glass pipette (Zhang et al., 2001). The transplantation site was on the right-hand side, 1 mm from the midline between the Bregman and Lambda. Cells were injected 1 mm deep so as to target the ventricle and future corpus callosum. Animals were anesthetized and perfused with 4% paraformaldehyde 3 months after transplantation. Brain tissues were processed for immunohistochemistry and electron microscopy as described (Zhang et al., 1998; Zhang et al., 2001).
Results
Expression of OLIG2 in hESC-derived progenitors is essential for OPC specification
hESCs differentiate to primitive NE at day 8-10, and this can be efficiently patterned to region-specific progenitors in response to particular morphogens (Li et al., 2005; Pankratz et al., 2007). To examine the role of OLIG2 in human oligodendrocyte differentiation, we treated the primitive NE with retinoic acid (RA; 0.1 μM) from day 10 to promote the specification of spinal progenitors. By day 14, multiple neural tube-like rosettes formed in each colony, which were detached and grown in suspension in the same neural medium with or without SHH (100 ng/ml) (Fig. 1A). At day 24 (or 10 days following SHH treatment), a subset of NE cells began to express OLIG2, which peaked at week 4 to 5 (Fig. 1B,C). In the absence of SHH, very few OLIG2-expressing cells (6%) were observed (Fig. 1B). To determine whether the generation of these few OLIG2+ cells depends on endogenous SHH, cyclopamine (5 μM), an inhibitor of hedgehog signaling, was added at day 10 together with RA. The presence of cyclopamine nearly eliminated the presence of OLIG2+ cells (Fig. 1B). Together, these results indicate that SHH, especially exogenous SHH, is required for OLIG2 expression in the hESC-derived NE.
Fig. 1. SHH-dependent OLIG2 expression for hESC differentiation to OPCs.
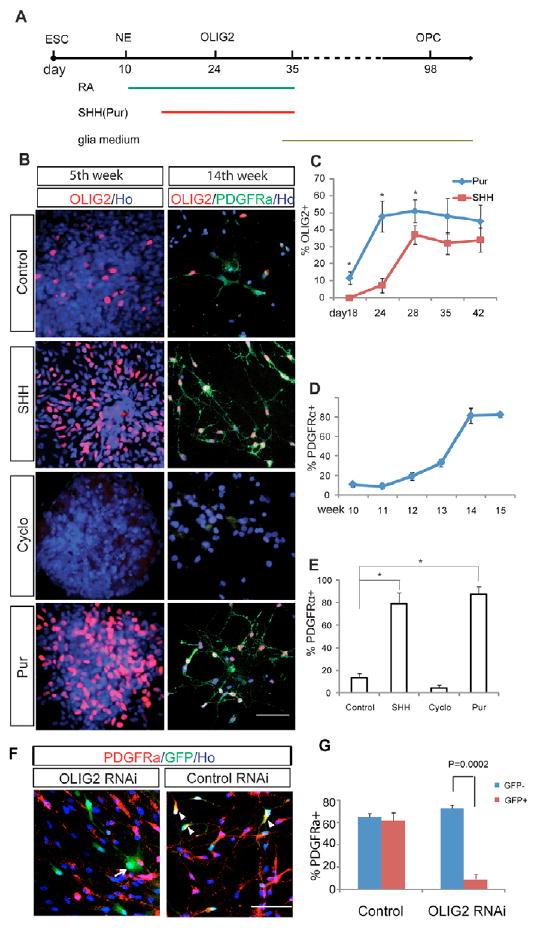
(A) Experimental paradigm showing differentiation of OLIG2 progenitors and OPCs. (B) OLIG2-expressing progenitors and OLIG2+ PDGFRα+ OPCs are present in the control, SHH-treated and purmorphamine (Pur)-treated cultures, but rarely in the cyclopamine (Cyclo)-treated cultures at day 35 (5th week) and day 98 (14th week). (C) Time course of OLIG2 expression in response to Pur (1 μM) and SHH (100 ng/ml). *P=0.011, 0.008 and 0.014 at day 18, 23 and 28, respectively. (D) PDGFRα+ OPCs first appear after 10 weeks of differentiation and reach a plateau at the fifteenth week. (E) Comparative effect of SHH, purmorphamine and cyclopamine on the efficiency of OPC generation at the fifteenth week. *P<0.0001. The percentage PDGFRα in D and E represents the proportion of PDGFRα+ cells among total cells. (F) OLIG2 RNAi cells do not express PDGFRα (arrows), whereas control RNAi cells are positive for PDGFRα (arrowheads). (G) Quantification of the effect of OLIG2 RNAi on the percentage of PDGFRα+ cells. Cells infected with the control RNAi virus (GFP+) generated a similar proportion of PDGFRα-expressing cells to those without viral infection (GFP−), whereas the cells infected with OLIG2 RNAi virus (GFP+) generated fewer PDGFRα+ OPCs than the non-infected population (GFP−). Ho, Hoechst 33258-stained nuclei. Scale bars: 50 μm.
To determine whether OLIG2 expression is essential for human OPC specification, at day 35, after the peak of OLIG2 progenitor generation, the cultures were switched to glial differentiation medium (Fig. 1A). OPCs co-expressing PDGFRα and OLIG2 began to appear at the tenth week of differentiation, reaching a peak in the fourteenth week (Fig. 1D). In the cultures with few OLIG2 progenitors (the control and cyclopamine groups), there were few PDGFRα+ OPCs. (Fig. 1B,E). Therefore, there was a clear correlation between the OLIG2 expression at week 5 and the presence of OPCs at week 14.
The requirement of OLIG2 expression for human OPC generation was verified by knocking down OLIG2 expression from day 35 with lentivirus-delivered RNAi. The expression of OLIG2 RNAi, indicated by the co-expression of GFP, inhibited the generation of PDGFRα+ cells at week 14, whereas the expression of OLIG2 and PDGFRα was not altered in the control RNAi-transfected cultures (compare the GFP+ and GFP− cells in Fig. 1F,G; see Fig. S1A,B in the supplementary material). Thus, OLIG2 expression is required for human OPC specification.
To further confirm the necessity of SHH signaling in OLIG2 expression and OPC specification, we replaced SHH with purmorphamine, a small molecule that activates the hedgehog signaling pathway (Sinha and Chen, 2006), in the above differentiation cultures. Treatment with 1 μM purmorphamine (Li et al., 2008) resulted in earlier (by 4-5 days) and more potent induction of OLIG2 expression (Fig. 1B,C). At week 14, a similar population of PDGFRα+ OPCs was generated as in the SHH-treated cultures (Fig. 1E). These results support the proposal that OLIG2 expression induced by SHH signaling is essential for human OPC specification. Since purmorphamine replicates the effect of SHH in OPC generation, yet is chemically stable, we have presented the data that were generated using purmorphamine.
SHH signaling is required for inducing and maintaining OLIG2 and NKX2.2-expressing pre-OPCs
At the time of OLIG2 progenitor generation (day 24), examination of OPC markers including SOX10, NKX2.2 and PDGFRα indicated that only NKX2.2 was present, but it did not co-express with OLIG2 (Fig. 2A,B). Following neuronal differentiation at the fifth week, OLIG2 progenitors from hESCs no longer produced motoneurons (Li et al., 2005). We asked whether these OLIG2 progenitors were now becoming OPCs. At day 35, ∼40% of the OLIG2+ cells co-expressed NKX2.2 (Fig. 2B). In the chick and mouse brainstem it is thought that OLIG2 and NKX2.2 co-expression defines the region for OPC generation, and OLIG2+ NKX2.2+ progenitors quickly acquire the expression of OPC markers including PDGFRα, NG2 and SOX10 (Finzsch et al., 2008; Liu et al., 2007; Richardson et al., 2006). However, the human OLIG2+ NKX2.2+ cells were negative for PDGFRα, NG2 and SOX10 (see below). We refer to these cells as pre-OPCs for convenience of description.
Fig. 2. SHH-dependent induction of pre-OPCs.
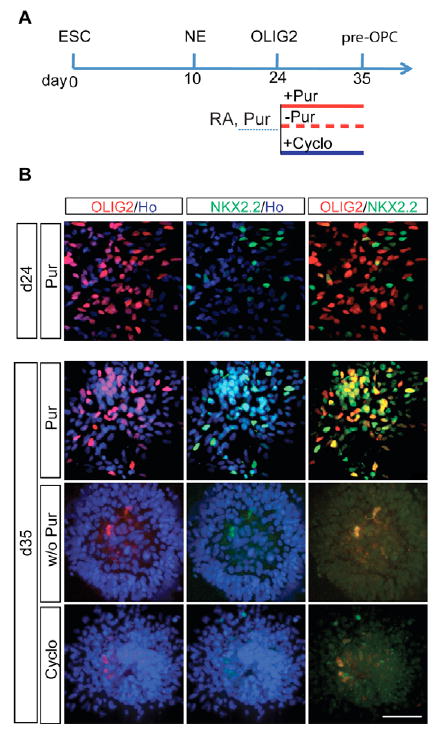
(A) Experimental design showing three groups treated with or without purmorphamine (Pur) or cyclopamine (Cyclo) from day 24 to 35. (B) OLIG2 and NKX2.2 are expressed at day 24, but not in the same cells. At day 35, OLIG2 and NKX2.2 co-express only in cultures with SHH or purmorphamine. Scale bar: 50 μm.
We then asked whether the induction of pre-OPCs is dependent upon SHH signaling. OLIG2 progenitor cultures at day 24 (Fig. 1A) were continued either with or without purmorphamine through day 35, or cyclopamine was added to block endogenous SHH (Fig. 2A). In the presence of purmorphamine, a population of OLIG2+ NKX2.2+ pre-OPCs appeared (Fig. 2B). By contrast, removal of purmorphamine or addition of cyclopamine resulted in a lack of cells expressing OLIG2 and/or NKX2.2 (Fig. 2B). These results indicate that not only the maintenance of OLIG2 expression, but also the induction of pre-OPCs is dependent on SHH signaling, especially exogenous SHH.
To determine whether the maintenance of pre-OPCs is dependent on SHH, the pre-OPC cultures (day 35) were grown in the absence or presence of purmorphamine (1 μM) or cyclopamine (5 μM) (Fig. 3A). At day 50, cells expressing OLIG2 and NKX2.2 were present in the cultures with or without continued purmorphamine, although ∼40% more OLIG2+ NKX2.2+ cells were seen in the purmorphamine-treated cultures as compared with cells without purmorphamine (Fig. 3B,C). However, the cultures treated with cyclopamine had very few cells expressing OLIG2 and/or NKX2.2, even though the cultures grew similarly to those of the other groups (Fig. 3B,C). At week 14, PDGFRα+ OPCs were present in the cultures with or without purmorphamine, but not in the cyclopamine-treated cultures (Fig. 3B,D). Thus, SHH signaling is necessary to maintain the co-expression of OLIG2 and NKX2.2 in pre-OPCs, and it subsequently promotes OPC generation even though exogenous SHH is no longer needed.
Fig. 3. Requirement of SHH signaling for maintenance of OLIG2 expression.
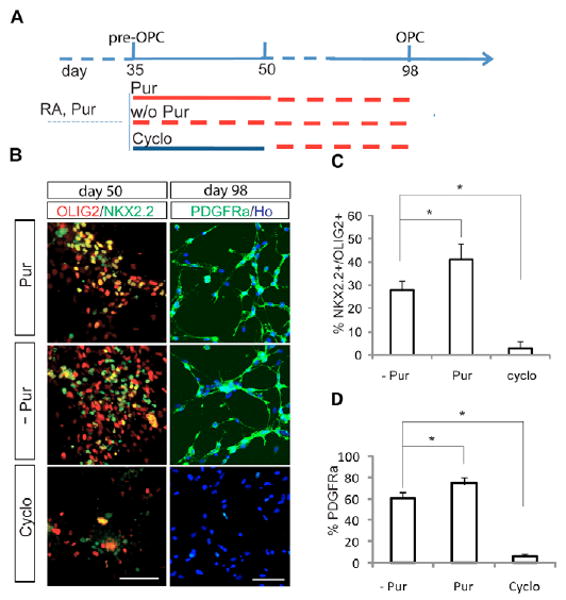
(A) Experimental design showing OLIG2-enriched cells treated with purmorphamine or cyclopamine until day 50. (B) At day 50, an OLIG2+ NKX2.2+ population is present in cultures with or without purmorphamine, but not in cultures with cyclopamine. PDGFRα+ OPCs appear in cultures with or without purmorphamine, but not in cultures with cyclopamine, at day 100. (C) Quantitative analysis showing the proportion of OLIG2+ NKX2.2+ cells (*P=0.023 between groups with and without purmorphamine; *P=0.0015 between groups without purmorphamine and with cyclopamine) at day 50. (D) Proportion of PDGFRα+ OPCs at the fourteenth week. *P=0.035 between groups with and without purmorphamine; *P<0.0001 between groups without purmorphamine and with cyclopamine. Scale bar: 50 μm.
FGF2 increases pre-OPCs by inhibiting motoneuron differentiation
In rodents, OLIG2-expressing spinal progenitors are a source of both motoneurons and OPCs (Lu et al., 2002; Zhou and Anderson, 2002). We hypothesized that repressing neurogenic potential might result in more OLIG2 progenitors for OPC generation during the gliogenic phase. Two mitogens, FGF2 (10 ng/ml) and EGF (20 ng/ml), were applied to the cultures along with purmorphamine from day 24 (before motoneuron differentiation) (Fig. 4A). FGF2 completely blocked HB9 (MNX1) expression at day 35, unlike the cultures without FGF2, which contained 38% (38.4±5.3%) HB9-expressing motoneurons. Whereas cultures with FGF2 alone had very few OLIG2+ or HB9+ cells, those with both purmorphamine and FGF2 had a much larger population (55%) of OLIG2+ cells (Fig. 4B,C; see Fig. S2 in the supplementary material). These OLIG2+ cells, after removal of FGF2 and addition of RA and SHH, a condition for efficient motoneuron differentiation (Li et al., 2005), no longer generated HB9+ cells (see Fig. S3 in the supplementary material). EGF had a moderate effect on motoneuron differentiation, but did not significantly alter the OLIG2+ population (Fig. 4B,C).
Fig. 4. FGF2 inhibits motoneuron differentiation and increases pre-OPCs.
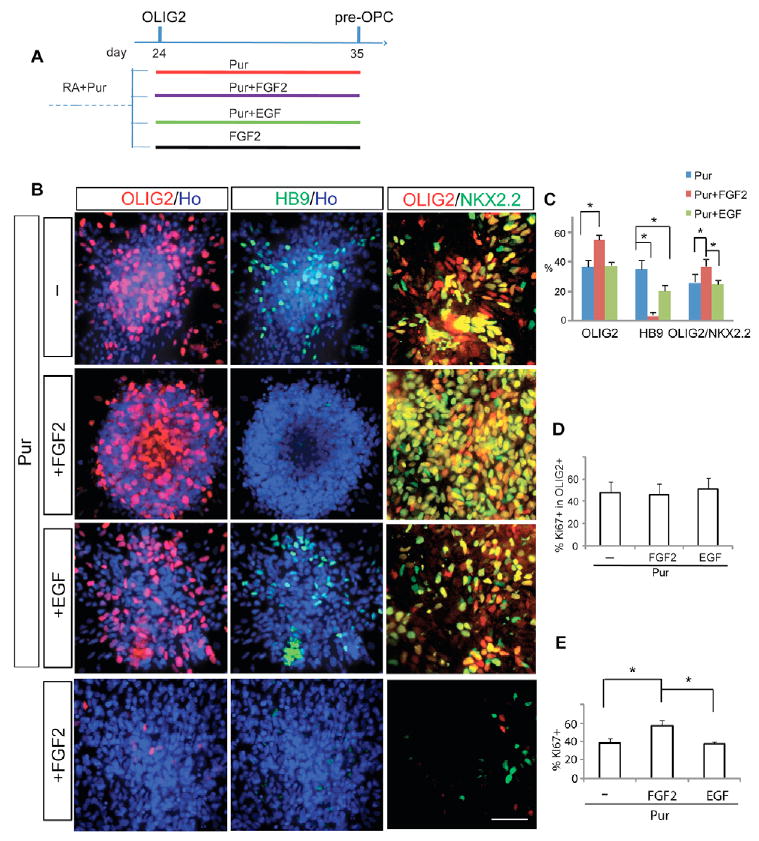
(A) Experimental design showing that the OLIG2+ cells at day 24 are further cultured for 12 days in the presence of purmorphamine or FGF2 alone, or in a combination of purmorphamine and FGF2 or EGF. (B) FGF2, but not EGF, blocks HB9 expression (column 2). (C) In the presence of purmorphamine, FGF2 but not EGF increases the proportion of OLIG2+ cells by ∼50% and decreases the HB9+ population to less than 5%, whereas EGF moderately decreases HB9+ cells. (D) Neither FGF2 nor EGF selectively alters the Ki67+ proportion in OLIG2+ cells. (E) There is a moderate increase in Ki67+ cells in FGF2-treated (for 10 days) cultures. *P<0.05 between the paired groups. Scale bar: 50 μm.
To assess whether the increased OLIG2+ population is contributed by the mitogenic effect of FGF2, we immunostained OLIG2+ cells with an antibody against Ki67, a protein expressed in dividing cells. The proportion of Ki67+ cells among the OLIG2+ population was not altered by FGF2 (Fig. 4D), even though we observed an overall 50% increase in Ki67+ cells in the cultures with FGF2 as compared with those without FGF2 (Fig. 4E). Since the OLIG2 progenitors and the HB9+ motoneurons accounted for ∼70% of the total population and FGF2 caused a drastic reduction of post-mitotic motoneurons from 38% to less than 5% (Fig. 4C), the increased Ki67+ population following FGF2 treatment can be largely attributed to the shrinking population of non-dividing motoneurons. Indeed, EGF had less effect in inhibiting motoneuron differentiation and did not alter the proportion of OLIG2+ or Ki67+ cells significantly (Fig. 4C,D). TUNEL labeling showed no obvious differences in cell death among the three groups (not shown). The increased OLIG2+ cell population is likely to be contributed by the accumulation of OLIG2 progenitors that failed to differentiate to motoneurons in the presence of FGF2, rather than simply by FGF2-induced proliferation of OLIG2 progenitors. Thus, FGF2 might increase the pre-OPCs by preventing OLIG2 progenitors from differentiating to motoneurons.
FGF2 inhibits the transition of pre-OPCs to OPCs by disrupting the SHH-dependent co-expression of OLIG2 and NKX2.2
FGF2 promotes OPC generation from mESCs and rodent neural stem/progenitor cells (Brustle et al., 1999; Chandran et al., 2003; Gabay et al., 2003; Kessaris et al., 2004). We asked whether FGF2 could similarly enhance the generation of OPCs from human pre-OPCs. hESCs were differentiated for 35 days in the presence of purmorphamine and FGF2 to enrich for pre-OPCs (Fig. 4A). The enriched pre-OPCs from day 35 were continued with purmorphamine until day 50. From day 35, the cells were cultured in the presence or absence of FGF2 (Fig. 5A). Weekly examination of the differentiation cultures revealed that by the fourteenth week, the majority of the cells (84%) in the group without FGF2 exhibited a typical bipolar or tripolar OPC morphology. These cells expressed multiple OPC markers including PDGFRα, SOX10 and NG2 (Fig. 5B,C). Very few cells were positive for the neuronal protein βIII-tubulin (5.1±4.4%) or the astrocytic antigen GFAP (4.7±2.7%) (Fig. 5B,D). By contrast, the FGF2-treated cultures possessed few PDGFRα+ OPCs (4.3±3.1%), whereas the majority of the cell populations were βIII-tubulin+ neurons (53±6.8%) and GFAP+ progenitors/astrocytes (34.7±5.2%) (Fig. 5B-D). Thus, FGF2 inhibits OPC generation and instead favors that of neurons or astrocytes.
Fig. 5. FGF2 inhibits the transition of pre-OPCs to OPCs by blocking co-expression of OLIG2 and NKX2.2.
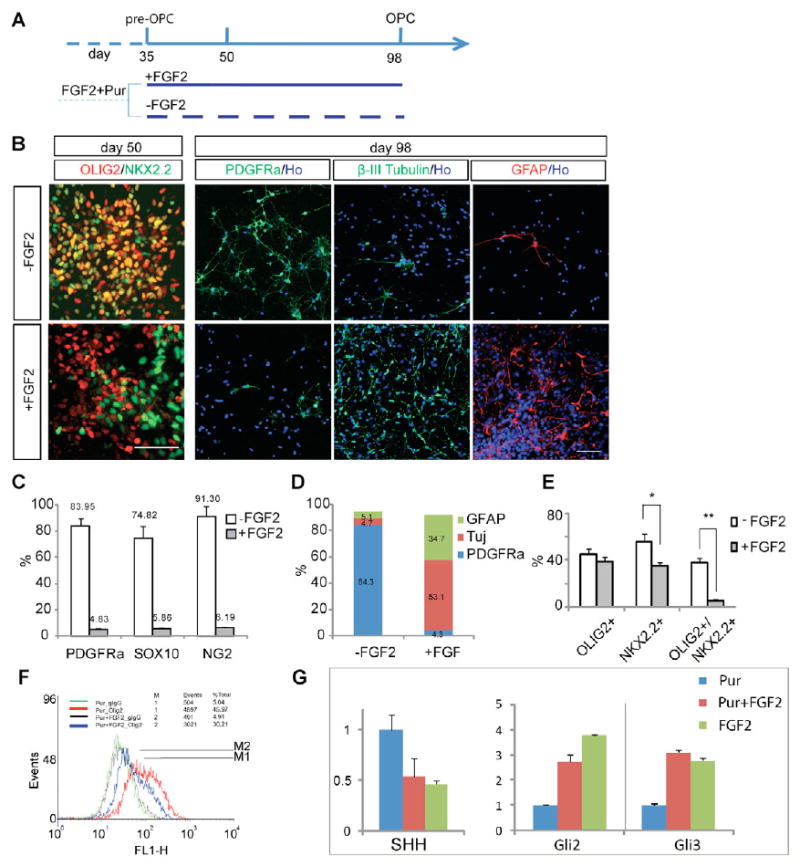
(A) Experimental design showing that the gliogenic OLIG2 progenitors at day 35 are treated with or without FGF2 and then examined at day 50 or day 98. (B) At day 50 (left column), the majority of the cells co-express OLIG2 and NKX2.2, whereas few double-positive cells are present in cultures with FGF2. At day 98, cultures without FGF2 (upper row) contain mostly PDGFRα+ cells with few cells positive for βIII-tubulin or GFAP. In the presence of FGF2, the majority of cells are positive for βIII-tubulin or GFAP (lower row). Scale bar: 100 μm. (C) A higher proportion of cells expressing OPC markers (percentage PDGFRα+, SOX10+ or NG2+ cells among total cells) is generated in the absence of FGF2 at day 98. (D) The population of PDGFRα+, βIII-tubulin+ and GFAP+ cells in the presence or absence of FGF2 at day 98. (E) FGF2 treatment results in a lower proportion of OLIG2+ NKX2.2+ cells at day 50. *P=0.048, **P=0.0018; n=3. (F) FACS analysis at day 50 shows a slight decrease (10%) in the OLIG2+ population and a leftward shift (indicating decreased expression level in individual cells) for the group with FGF2 (blue) compared with that without FGF2 (red). (G) Quantitative RT-PCR analysis at day 50 showing that FGF2 decreases SHH expression by half and increases the transcription of GLI2 and GLI3.
To investigate how FGF2 inhibits the transition of pre-OPCs to OPCs, the pre-OPC cultures were treated with or without FGF2 at day 35 and analyzed at day 50 (Fig. 5A). FGF2 treatment resulted in a lack of OLIG2+ NKX2.2+ cells, as compared with 40% double-positive cells in the cultures without FGF2 (Fig. 5B, left panel). The OLIG2+ cell population appeared similar, but the immunofluorescence intensity was lower in the FGF2-treated cells (Fig. 5B,E), which was confirmed by the leftward shift of the curve in FACS analysis (Fig. 5F). The NKX2.2+ cell population decreased significantly upon FGF2 treatment, while the fluorescence intensity of individual cells remained unchanged (Fig. 5B,E). Since our data suggested that the induction and maintenance of OLIG2 and NKX2.2 expression is crucial for human OPC specification (Fig. 3), we hypothesized that FGF2 blocks co-expression of OLIG2 and NKX2.2 by disrupting SHH signaling. Quantitative RT-PCR analyses indicated that FGF2 reduces endogenous SHH expression by half (Fig. 5G). The intracellular repressive mediators of SHH signaling, GLI2 and GLI3 (Ruiz i Altaba et al., 2007), were significantly upregulated by FGF2 alone or FGF2 plus purmorphamine (Fig. 5G). This might account for the loss of OLIG2 and NKX2.2 co-expression in pre-OPCs, hence inhibiting subsequent transition to OPCs.
hESC-derived OPCs differentiate to oligodendrocytes in vitro and produce myelin sheaths in vivo
Analyses of the signaling requirements during multiple stages of OPC specification have resulted in the present 3-month-long protocol for OPC generation from hESCs (Fig. 6A). After 14 weeks of hESC differentiation, the plated OPCs exhibited a bipolar morphology (Fig. 6B,C), with the vast majority co-expressing OLIG2 and NKX2.2 (Fig. 6D). These cells also expressed SOX10 (Fig. 6E). In addition to the oligodendroglial transcription factors, these cells co-expressed PDGFRα or NG2 on the membrane and processes (Fig. 6E-G). Over the course of the subsequent 2-4 weeks, these bipolar cells became multipolar and were positive for the immature oligodendrocyte marker O4 (Fig. 6H,I). The O4+ cells were no longer positive for PDGFRα (see Fig. S4 in the supplementary material), similar to our previous finding using fetal brain tissue-derived progenitor cultures (Zhang et al., 2000). The expression of another commonly used OPC marker, NG2, often overlapped with that of O4 (see Fig. S4 in the supplementary material). With further culturing for 6-8 weeks, most cells died in our oligodendroglia-enriched cultures under the serum-free conditions. Nevertheless, surviving cells exhibited more-elaborated processes, occasionally with membrane sheaths. These cells were positively labeled for myelin basic protein (MBP) (Fig. 6J), which is expressed by mature oligodendrocytes.
Fig. 6. Maturation and myelination of hESC-derived OPCs.
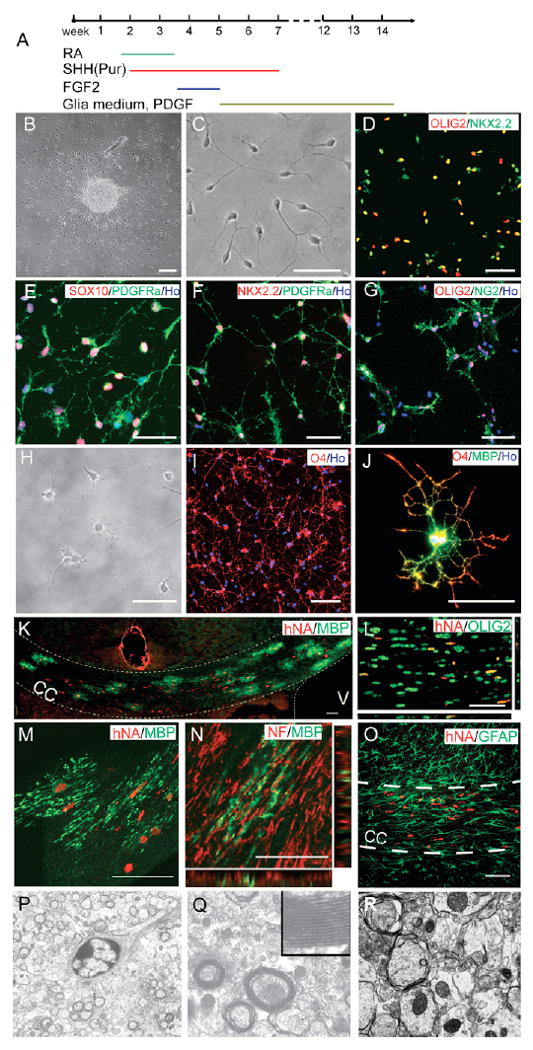
(A) Summary of procedures for directed differentiation of OPCs from hESCs. (B) Bipolar cells are present surrounding the progenitor cluster 3 days after plating. (C) Dissociated OPCs exhibit a bipolar morphology. (D-G) The OPCs co-express OLIG2+ NKX2.2+ (D), SOX10+ PDGFRα+ (E), NKX2.2+ PDGFRα+ (F) and OLIG2+ NG2+ (G). (H, I) Six weeks following OPC differentiation, cells exhibit multipolar and web-like processes and express O4. (J) After an additional month, some cells express MBP. (K) Three months after transplantation, the grafted cells, as revealed by hNA and MBP, preferentially localize in the corpus callosum (CC, outlined). (L) All of the human cells are positive for OLIG2. (M) The human cells exhibit multiple MBP+ processes. (N) Confocal analysis shows adjacent expression of MBP+ human cell fibers with NF+ axons in the corpus callosum. (O) The hNA+ cells are negative for GFAP. (P) Electron micrograph showing a typical oligogodendrocyte surrounded by many myelin profiles in the grafted brain. (Q) Higher magnification from P showing compact myelin sheaths in the grafted brain with identifiable major dense lines (inset). (R) Electron micrograph showing thin non-compact sheaths wrapping around axons in the corpus callusom of a shiverer mouse without transplant. Magnification: P, 6000×; Q,R, 20,000×; Q inset, 60,000×. Scale bars: 50 μm in B-I,K-O; 10 μm in J.
The myelination potential of the in vitro-produced human OPCs was investigated by transplanting the OPCs (differentiated from hESCs for 12 weeks) into the ventricle of neonatal shiverer mice. Shiverer mice do not produce endogenous compact myelin sheaths owing to a complete lack of MBP. At 3 months following transplantation, grafted human cells, identified by specific human nuclear antigen (hNA) expression, were present preferentially in the corpus callosum (Fig. 6K). Virtually every hNA-expressing cell was also positive for OLIG2 (Fig. 6L). Most of the human cells displayed numerous processes and immunostained positively for MBP (Fig. 6M). These MBP-expressing processes connected to multiple neurofilament-positive axons in the corpus callosum (Fig. 6N). No human cells co-labeled for GFAP (Fig. 6O). Toluidine Blue staining on the semi-thin sections showed numerous myelin sheaths in grafted animals, but not in non-transplanted shiverer mice (not shown). Electron microscopy analyses confirmed the presence of compact myelin sheaths in the corpus callosum of the transplanted mice (Fig. 6P,Q), but not in the non-transplanted shiverer mice (Fig. 6R).
Discussion
Inaccessibility to human embryo experimentation calls for an alternative, in vitro model to study human cells or tissues directly. In recent years, directed neural differentiation from hESCs has allowed a re-examination of the fundamental principles of early neural development learned from vertebrate studies (Elkabetz et al., 2008; Li et al., 2005; Pankratz et al., 2007; Roy et al., 2006; Watanabe et al., 2007; Yan et al., 2005). In the present study, we have revealed that human oligodendrocyte development involves a transcriptional network of nearly identical sequence to that observed in vertebrate models. Following expression of OLIG2 and genesis of motoneurons, the human OLIG2 progenitors become pre-OPCs by co-expressing NKX2.2, and finally differentiate into OPCs by activation of SOX10 and PDGFRα. Blocking OLIG2 expression inhibits OPC production, confirming the requirement of OLIG2 for human OPC specification. The vast majority of human OLIG2 progenitors are generated in a SHH-dependent manner, because cultures without exogenous SHH, or those in which endogenous SHH signaling has been blocked with cyclopamine, have few OLIG2 progenitors and OPCs. We also found that SHH is not only crucial for efficiently inducing pre-OPCs, but it is also required for the transition from pre-OPCs to OPCs, for which endogenous SHH signaling is sufficient (Fig. 7). Thus, the SHH-dependent signaling network underlying vertebrate OPC development is conserved in humans (Fig. 7).
Fig. 7. Regulation of OPC specification from hESCs by SHH and FGF2.
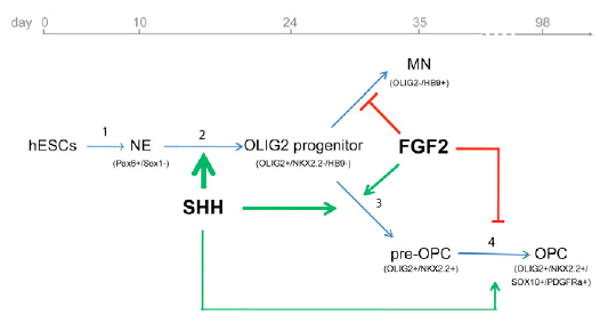
Human ESCs (hESCs) are differentiated to neuroepithelial (NE) cells without exogenous growth factors (1), then to OLIG2-expressing ventral spinal progenitors (2) and OLIG2+ NKX2.2+ pre-OPCs (3) before becoming SOX10+ PDGFRα+ OPCs (4). The induction of OLIG2 progenitors (2) and pre-OPCs (3) is largely dependent upon exogenous SHH (thick arrows). SHH produced in the culture (thin arrow) is sufficient for the transition of pre-OPCs to OPCs (4). FGF2 increases the pre-OPCs during the neurogenic phase through inhibition of motoneuron differentiation (3), but blocks the transition of pre-OPCs to OPCs by inhibiting the SHH-dependent co-expression of OLIG2 and NKX2.2 in pre-OPCs (4).
Our study also reveals unique aspects of human OPC generation. In our human OPC differentiation cultures, NKX2.2 is the first OPC-related transcription factor that is co-expressed with OLIG2 at the fifth week, preceding the expression of PDGFRα. This expression pattern resembles that in the chick and in the mouse hindbrain (Vallstedt et al., 2005; Zhou et al., 2001), but differs from that in the mouse spinal cord, where NKX2.2 expression is induced after PDGFRα+ OPCs are formed (Fu et al., 2002; Qi et al., 2001). This might be partly related to the hindbrain/cervical spinal identity of our human progenitors patterned by RA (Li et al., 2005), although species differences cannot be excluded. We have also found a protracted transition period from pre-OPCs at the fifth week to human OPCs at the fourteenth week. In chick and mouse brainstem, the OLIG2+ NKX2.2+ progenitors quickly express PDGFRα and become migrating OPCs (Vallstedt et al., 2005; Zhou et al., 2001). OLIG2 progenitors differentiated from mESCs also co-express PDGFRα and NG2 shortly following motoneuron differentiation (Billon et al., 2002; Du et al., 2006). It is possible that our serum-free culture conditions are not optimal for acquisition of the OPC identity at an earlier time. The addition of FGF2, EGF, SHH or noggin, however, did not accelerate the transition of pre-OPCs to OPCs (data not shown). Similarly, removal of PDGF, NT3 and/or IGF1 did not alter the time course of OPC generation either (data not shown). Alternatively, this protracted transition might be intrinsically controlled. The long OPC-specification process (3 months) appears to match the earliest appearance of OPCs in human embryos at the end of the first trimester (Grever et al., 1997; Jakovcevski and Zecevic, 2005; Weidenheim et al., 1996). One potential explanation for the late appearance of OPCs is a need for expanding neurogenic progenitors, as large numbers of neurons are needed for the evolutionarily enlarged human brain and spinal cord.
FGF2 is a mitogen for rodent and human neural stem/progenitor cells and enhances the generation of OPCs in rodents (Avellana-Adalid et al., 1996; Brustle et al., 1999; Gabay et al., 2003; Zhang et al., 1998). In the present study, it is interesting that following the induction of OLIG2, FGF2 nearly completely blocked motoneuron differentiation and preferentially increased the proportion of OLIG2 progenitors. The inhibition of motoneuron differentiation is not attributable to the mitogenic effect of FGF2 preventing the OLIG2 progenitors from exiting the cell cycle. Nor does FGF2 preferentially promote the survival of the OLIG2 progenitors. It is likely that FGF2 enhances the transition from the neurogenic OLIG2 progenitors to pre-OPCs (Fig. 7). Although fine dissection of the mechanism is not possible with the available system, this finding provides, as it stands, a simple way of switching neurogenesis to gliogenesis from a pool of progenitors.
Continued use of FGF2 inhibited the generation of OPCs from hESCs. This is reminiscent of observations in the past decade that human neural stem/progenitor cells, following expansion with FGF2, or FGF2 and EGF, rarely produce oligodendrocytes in vitro (Chandran et al., 2004; Roy et al., 1999; Zhang et al., 2000). Even when the human neural progenitor cultures are enriched for OPCs, the OPCs quickly disappear after culturing in the presence of FGF2 (Grever et al., 1999). Our present finding that co-expression of OLIG2 and NKX2.2 is suppressed by FGF2 in the pre-OPCs explains the phenomenon. The inhibitory effect appears specific to FGF2, as EGF did not affect the co-expression of OLIG2 and NKX2.2 and subsequent generation of OPCs (Fig. 4). FGF2 induces OPC generation from mouse neural stem/progenitor cells by activating endogenous SHH signaling (Gabay et al., 2003) or by as yet unknown SHH-independent pathways (Chandran et al., 2003; Kessaris et al., 2004). In our human cell differentiation system, FGF2 inhibits endogenous SHH expression and significantly increases the level of GLI2 and GLI3, which are downstream repressors of SHH signaling (Ruiz i Altaba et al., 2007), thus disrupting co-expression of OLIG2 and NKX2.2 in pre-OPCs. This results in the loss of pre-OPCs and the conversion to progenitors expressing OLIG2 (at a low level) or NKX2.2, which generates astrocytes or neurons (Fig. 5B,D). Nevertheless, we do not exclude the possibility that FGF does this through other oligodendroglial transcription factors, such as SOX10 and ASCL1 (Liu et al., 2007; Sugimori et al., 2008), or by selectively promoting the survival/expansion of neuronal progenitors in our long-term cultures.
In the present study, we developed an effective strategy for reproducibly directing hESCs to an enriched population of OPCs with an unequivocal oligodendrocyte identity and myelination potential. In the previous reports of OPC differentiation from hESCs, SHH was not applied and the expression of OLIG2 in neural progenitors was not examined by single-cell-resolution immunocytochemistry, but instead by PCR on bulk cultures (Izrael et al., 2007; Kang et al., 2007). Based on our findings, we believe that the OPCs described in those reports are differentiated from OLIG2 progenitors spontaneously induced by endogenous or alternatively activated SHH signaling. The near pure population of ‘OPCs’ generated from hESCs without application of SHH (Nistor et al., 2005) cannot be explained by our model, nor has the result been replicated by us or any other reports. The identity of the OPCs in that report has not been unequivocally verified either.
The SHH-dependent transcriptional network underlying human OPC specification and the time course of OPC generation, consistent with that in human embryo development, revealed in the present study suggest that the hESC differentiation system is a useful tool for understanding the biology of human cells. The divergent responses to common growth factors such as FGF2 between human and other vertebrate cells might be related to the in vitro system, but may also reflect the nature of normal human development. Similar to our present finding, the maintenance of human and mouse ESCs depends on the same transcriptional network (Boyer et al., 2005). However, the common extrinsic factor, BMP, maintains the self-renewal of mESCs but induces cell differentiation of hESCs (Xu et al., 2002; Ying et al., 2003). This seemingly ‘trivial’ deviation has slowed down the translation of findings from mESCs to the establishment of hESCs (Evans and Kaufman, 1981; Thomson et al., 1998). Similarly, over the past decade, many laboratories have stumbled in trying to replicate the finding in rodents, so as to differentiate human neural stem/progenitors to OPCs. Thus, the confirmation of conserved principles and the revelation of ‘nuances’ using the hESC differentiation system might bear significant consequences.
Supplementary Material
Acknowledgments
This study was supported by the National Institute of Neurological Disorders and Stroke (R01 NS045926), the National Multiple Sclerosis Society, a gift from the Busta family and the Bleser family, and partly by a core grant to the Waisman Center from the National Institute of Child Health and Human Development (P30 HD03352). Deposited in PMC for release after 12 months.
Footnotes
Supplementary material for this article is available at http://dev.biologists.org/cgi/content/full/dev.029447/DC1
References
- Avellana-Adalid V, Nait-Oumesmar B, Lachapelle F, Baron-Van Evercooren A. Expansion of rat oligodendrocyte progenitors into proliferative ‘oligospheres’ that retain differentiation potential. J Neurosci Res. 1996;45:558–570. doi: 10.1002/(SICI)1097-4547(19960901)45:5<558::AID-JNR6>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- Billon N, Jolicoeur C, Ying QL, Smith A, Raff M. Normal timing of oligodendrocyte development from genetically engineered, lineage-selectable mouse ES cells. J Cell Sci. 2002;115:3657–3665. doi: 10.1242/jcs.00049. [DOI] [PubMed] [Google Scholar]
- Boyer LA, Lee TI, Cole MF, Johnstone SE, Levine SS, Zucker JP, Guenther MG, Kumar RM, Murray HL, Jenner RG, et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell. 2005;122:947–956. doi: 10.1016/j.cell.2005.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brustle O, Jones KN, Learish RD, Karram K, Choudhary K, Wiestler OD, Duncan ID, McKay RD. Embryonic stem cell-derived glial precursors: a source of myelinating transplants. Science. 1999;285:754–756. doi: 10.1126/science.285.5428.754. [DOI] [PubMed] [Google Scholar]
- Cai J, Qi Y, Hu X, Tan M, Liu Z, Zhang J, Li Q, Sander M, Qiu M. Generation of oligodendrocyte precursor cells from mouse dorsal spinal cord independent of Nkx6 regulation and Shh signaling. Neuron. 2005;45:41–53. doi: 10.1016/j.neuron.2004.12.028. [DOI] [PubMed] [Google Scholar]
- Chandran S, Kato H, Gerreli D, Compston A, Svendsen CN, Allen ND. FGF-dependent generation of oligodendrocytes by a hedgehog-independent pathway. Development. 2003;130:6599–6609. doi: 10.1242/dev.00871. [DOI] [PubMed] [Google Scholar]
- Chandran S, Compston A, Jauniaux E, Gilson J, Blakemore W, Svendsen C. Differential generation of oligodendrocytes from human and rodent embryonic spinal cord neural precursors. Glia. 2004;47:314–324. doi: 10.1002/glia.20011. [DOI] [PubMed] [Google Scholar]
- Du ZW, Li XJ, Nguyen GD, Zhang SC. Induced expression of Olig2 is sufficient for oligodendrocyte specification but not for motoneuron specification and astrocyte repression. Mol Cell Neurosci. 2006;33:371–380. doi: 10.1016/j.mcn.2006.08.007. [DOI] [PubMed] [Google Scholar]
- Elkabetz Y, Panagiotakos G, Al Shamy G, Socci ND, Tabar V, Studer L. Human ES cell-derived neural rosettes reveal a functionally distinct early neural stem cell stage. Genes Dev. 2008;22:152–165. doi: 10.1101/gad.1616208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292:154–156. doi: 10.1038/292154a0. [DOI] [PubMed] [Google Scholar]
- Finzsch M, Stolt CC, Lommes P, Wegner M. Sox9 and Sox10 influence survival and migration of oligodendrocyte precursors in the spinal cord by regulating PDGF receptor alpha expression. Development. 2008;135:637–646. doi: 10.1242/dev.010454. [DOI] [PubMed] [Google Scholar]
- Fogarty M, Richardson WD, Kessaris N. A subset of oligodendrocytes generated from radial glia in the dorsal spinal cord. Development. 2005;132:1951–1959. doi: 10.1242/dev.01777. [DOI] [PubMed] [Google Scholar]
- Fu H, Qi Y, Tan M, Cai J, Takebayashi H, Nakafuku M, Richardson W, Qiu M. Dual origin of spinal oligodendrocyte progenitors and evidence for the cooperative role of Olig2 and Nkx2.2 in the control of oligodendrocyte differentiation. Development. 2002;129:681–693. doi: 10.1242/dev.129.3.681. [DOI] [PubMed] [Google Scholar]
- Gabay L, Lowell S, Rubin LL, Anderson DJ. Deregulation of dorsoventral patterning by FGF confers trilineage differentiation capacity on CNS stem cells in vitro. Neuron. 2003;40:485–499. doi: 10.1016/s0896-6273(03)00637-8. [DOI] [PubMed] [Google Scholar]
- Grever WE, Weidenheim KM, Tricoche M, Rashbaum WK, Lyman WD. Oligodendrocyte gene expression in the human fetal spinal cord during the second trimester of gestation. J Neurosci Res. 1997;47:332–340. doi: 10.1002/(sici)1097-4547(19970201)47:3<332::aid-jnr11>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- Grever WE, Zhang S, Ge B, Duncan ID. Fractionation and enrichment of oligodendrocytes from developing human brain. J Neurosci Res. 1999;57:304–314. doi: 10.1002/(SICI)1097-4547(19990801)57:3<304::AID-JNR2>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- Izrael M, Zhang P, Kaufman R, Shinder V, Ella R, Amit M, Itskovitz-Eldor J, Chebath J, Revel M. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Mol Cell Neurosci. 2007;34:310–323. doi: 10.1016/j.mcn.2006.11.008. [DOI] [PubMed] [Google Scholar]
- Jakovcevski I, Zecevic N. Olig transcription factors are expressed in oligodendrocyte and neuronal cells in human fetal CNS. J Neurosci. 2005;25:10064–10073. doi: 10.1523/JNEUROSCI.2324-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang SM, Cho MS, Seo H, Yoon CJ, Oh SK, Choi YM, Kim DW. Efficient induction of oligodendrocytes from human embryonic stem cells. Stem Cells. 2007;25:419–424. doi: 10.1634/stemcells.2005-0482. [DOI] [PubMed] [Google Scholar]
- Kessaris N, Jamen F, Rubin LL, Richardson WD. Cooperation between sonic hedgehog and fibroblast growth factor/MAPK signalling pathways in neocortical precursors. Development. 2004;131:1289–1298. doi: 10.1242/dev.01027. [DOI] [PubMed] [Google Scholar]
- Lee SK, Lee B, Ruiz EC, Pfaff SL. Olig2 and Ngn2 function in opposition to modulate gene expression in motor neuron progenitor cells. Genes Dev. 2005;19:282–294. doi: 10.1101/gad.1257105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li XJ, Du ZW, Zarnowska ED, Pankratz M, Hansen LO, Pearce RA, Zhang SC. Specification of motoneurons from human embryonic stem cells. Nat Biotechnol. 2005;23:215–221. doi: 10.1038/nbt1063. [DOI] [PubMed] [Google Scholar]
- Li XJ, Hu BY, Jones SA, Zhang YS, Lavaute T, Du ZW, Zhang SC. Directed differentiation of ventral spinal progenitors and motor neurons from human embryonic stem cells by small molecules. Stem Cells. 2008;26:886–893. doi: 10.1634/stemcells.2007-0620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Z, Hu X, Cai J, Liu B, Peng X, Wegner M, Qiu M. Induction of oligodendrocyte differentiation by Olig2 and Sox10: evidence for reciprocal interactions and dosage-dependent mechanisms. Dev Biol. 2007;302:683–693. doi: 10.1016/j.ydbio.2006.10.007. [DOI] [PubMed] [Google Scholar]
- Lu QR, Yuk D, Alberta JA, Zhu Z, Pawlitzky I, Chan J, McMahon AP, Stiles CD, Rowitch DH. Sonic hedgehog-regulated oligodendrocyte lineage genes encoding bHLH proteins in the mammalian central nervous system. Neuron. 2000;25:317–329. doi: 10.1016/s0896-6273(00)80897-1. [DOI] [PubMed] [Google Scholar]
- Lu QR, Sun T, Zhu Z, Ma N, Garcia M, Stiles CD, Rowitch DH. Common developmental requirement for Olig function indicates a motor neuron/oligodendrocyte connection. Cell. 2002;109:75–86. doi: 10.1016/s0092-8674(02)00678-5. [DOI] [PubMed] [Google Scholar]
- Marquardt T, Pfaff SL. Cracking the transcriptional code for cell specification in the neural tube. Cell. 2001;106:651–654. doi: 10.1016/s0092-8674(01)00499-8. [DOI] [PubMed] [Google Scholar]
- Nistor GI, Totoiu MO, Haque N, Carpenter MK, Keirstead HS. Human embryonic stem cells differentiate into oligodendrocytes in high purity and myelinate after spinal cord transplantation. Glia. 2005;49:385–396. doi: 10.1002/glia.20127. [DOI] [PubMed] [Google Scholar]
- Pankratz MT, Li XJ, Lavaute TM, Lyons EA, Chen X, Zhang SC. Directed neural differentiation of human embryonic stem cells via an obligated primitive anterior stage. Stem Cells. 2007;25:1511–1520. doi: 10.1634/stemcells.2006-0707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi Y, Cai J, Wu Y, Wu R, Lee J, Fu H, Rao M, Sussel L, Rubenstein J, Qiu M. Control of oligodendrocyte differentiation by the Nkx2.2 homeodomain transcription factor. Development. 2001;128:2723–2733. doi: 10.1242/dev.128.14.2723. [DOI] [PubMed] [Google Scholar]
- Richardson WD, Kessaris N, Pringle N. Oligodendrocyte wars. Nat Rev Neurosci. 2006;7:11–18. doi: 10.1038/nrn1826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy NS, Wang S, Harrison-Restelli C, Benraiss A, Fraser RA, Gravel M, Braun PE, Goldman SA. Identification, isolation, and promoter-defined separation of mitotic oligodendrocyte progenitor cells from the adult human subcortical white matter. J Neurosci. 1999;19:9986–9995. doi: 10.1523/JNEUROSCI.19-22-09986.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy NS, Cleren C, Singh SK, Yang L, Beal MF, Goldman SA. Functional engraftment of human ES cell-derived dopaminergic neurons enriched by coculture with telomerase-immortalized midbrain astrocytes. Nat Med. 2006;12:1259–1268. doi: 10.1038/nm1495. [DOI] [PubMed] [Google Scholar]
- Ruiz i Altaba A, Mas C, Stecca B. The Gli code: an information nexus regulating cell fate, stemness and cancer. Trends Cell Biol. 2007;17:438–447. doi: 10.1016/j.tcb.2007.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samanta J, Kessler JA. Interactions between ID and OLIG proteins mediate the inhibitory effects of BMP4 on oligodendroglial differentiation. Development. 2004;131:4131–4142. doi: 10.1242/dev.01273. [DOI] [PubMed] [Google Scholar]
- Sinha S, Chen JK. Purmorphamine activates the Hedgehog pathway by targeting Smoothened. Nat Chem Biol. 2006;2:29–30. doi: 10.1038/nchembio753. [DOI] [PubMed] [Google Scholar]
- Stolt CC, Lommes P, Friedrich RP, Wegner M. Transcription factors Sox8 and Sox10 perform non-equivalent roles during oligodendrocyte development despite functional redundancy. Development. 2004;131:2349–2358. doi: 10.1242/dev.01114. [DOI] [PubMed] [Google Scholar]
- Sugimori M, Nagao M, Bertrand N, Parras CM, Guillemot F, Nakafuku M. Combinatorial actions of patterning and HLH transcription factors in the spatiotemporal control of neurogenesis and gliogenesis in the developing spinal cord. Development. 2007;134:1617–1629. doi: 10.1242/dev.001255. [DOI] [PubMed] [Google Scholar]
- Sugimori M, Nagao M, Parras CM, Nakatani H, Lebel M, Guillemot F, Nakafuku M. Ascl1 is required for oligodendrocyte development in the spinal cord. Development. 2008;135:1271–1281. doi: 10.1242/dev.015370. [DOI] [PubMed] [Google Scholar]
- Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–1147. doi: 10.1126/science.282.5391.1145. [DOI] [PubMed] [Google Scholar]
- Vallstedt A, Klos JM, Ericson J. Multiple dorsoventral origins of oligodendrocyte generation in the spinal cord and hindbrain. Neuron. 2005;45:55–67. doi: 10.1016/j.neuron.2004.12.026. [DOI] [PubMed] [Google Scholar]
- Watanabe K, Ueno M, Kamiya D, Nishiyama A, Matsumura M, Wataya T, Takahashi JB, Nishikawa S, Muguruma K, Sasai Y. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nat Biotechnol. 2007;25:681–686. doi: 10.1038/nbt1310. [DOI] [PubMed] [Google Scholar]
- Weidenheim KM, Bodhireddy SR, Rashbaum WK, Lyman WD. Temporal and spatial expression of major myelin proteins in the human fetal spinal cord during the second trimester. J Neuropathol Exp Neurol. 1996;55:734–745. doi: 10.1097/00005072-199606000-00008. [DOI] [PubMed] [Google Scholar]
- Xia X, Ayala M, Thiede BR, Zhang SC. In vitro- and in vivo-induced transgene expression in human embryonic stem cells and derivatives. Stem Cells. 2008;26:525–533. doi: 10.1634/stemcells.2007-0710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu RH, Chen X, Li DS, Li R, Addicks GC, Glennon C, Zwaka TP, Thomson JA. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat Biotechnol. 2002;20:1261–1264. doi: 10.1038/nbt761. [DOI] [PubMed] [Google Scholar]
- Yan Y, Yang D, Zarnowska ED, Du Z, Werbel B, Valliere C, Pearce RA, Thomson JA, Zhang SC. Directed differentiation of dopaminergic neuronal subtypes from human embryonic stem cells. Stem Cells. 2005;23:781–790. doi: 10.1634/stemcells.2004-0365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ying QL, Nichols J, Chambers I, Smith A. BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell. 2003;115:281–292. doi: 10.1016/s0092-8674(03)00847-x. [DOI] [PubMed] [Google Scholar]
- Zhang SC, Lundberg C, Lipsitz D, O'Connor LT, Duncan ID. Generation of oligodendroglial progenitors from neural stem cells. J Neurocytol. 1998;27:475–489. doi: 10.1023/a:1006953023845. [DOI] [PubMed] [Google Scholar]
- Zhang SC, Ge B, Duncan ID. Tracing human oligodendroglial development in vitro. J Neurosci Res. 2000;59:421–429. doi: 10.1002/(SICI)1097-4547(20000201)59:3<421::AID-JNR17>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- Zhang SC, Wernig M, Duncan ID, Brustle O, Thomson JA. In vitro differentiation of transplantable neural precursors from human embryonic stem cells. Nat Biotechnol. 2001;19:1129–1133. doi: 10.1038/nbt1201-1129. [DOI] [PubMed] [Google Scholar]
- Zhou Q, Anderson DJ. The bHLH transcription factors OLIG2 and OLIG1 couple neuronal and glial subtype specification. Cell. 2002;109:61–73. doi: 10.1016/s0092-8674(02)00677-3. [DOI] [PubMed] [Google Scholar]
- Zhou Q, Wang S, Anderson DJ. Identification of a novel family of oligodendrocyte lineage-specific basic helix-loop-helix transcription factors. Neuron. 2000;25:331–343. doi: 10.1016/s0896-6273(00)80898-3. [DOI] [PubMed] [Google Scholar]
- Zhou Q, Choi G, Anderson DJ. The bHLH transcription factor Olig2 promotes oligodendrocyte differentiation in collaboration with Nkx2.2. Neuron. 2001;31:791–807. doi: 10.1016/s0896-6273(01)00414-7. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.