

Abstract
Epidemiological studies demonstrate that a high dietary intake of carotenoids may offer protection against age-related macular degeneration, cancer and cardiovascular and neurodegenerative diseases. Humans cannot synthesize carotenoids and depend on their dietary intake. Major carotenoids that have been found in human plasma can be divided into two groups, carotenes (nonpolar molecules, such as β-carotene, α-carotene or lycopene) and xanthophylls (polar carotenoids that include an oxygen atom in their structure, such as lutein, zeaxanthin and β-cryptoxanthin). Only two dietary carotenoids, namely lutein and zeaxanthin (macular xanthophylls), are selectively accumulated in the human retina. A third carotenoid, meso-zeaxanthin, is formed directly in the human retina from lutein. Additionally, xanthophylls account for about 70% of total carotenoids in all brain regions. Some specific properties of these polar carotenoids must explain why they, among other available carotenoids, were selected during evolution to protect the retina and brain. It is also likely that the selective uptake and deposition of macular xanthophylls in the retina and brain are enhanced by specific xanthophyll-binding proteins. We hypothesize that the high membrane solubility and preferential transmembrane orientation of macular xanthophylls distinguish them from other dietary carotenoids, enhance their chemical and physical stability in retina and brain membranes and maximize their protective action in these organs. Most importantly, xanthophylls are selectively concentrated in the most vulnerable regions of lipid bilayer membranes enriched in polyunsaturated lipids. This localization is ideal if macular xanthophylls are to act as lipid-soluble antioxidants, which is the most accepted mechanism through which lutein and zeaxanthin protect neural tissue against degenerative diseases.
Keywords: carotenoids, macular xanthophylls, zeaxanthin, lutein, neural tissue, lipid antioxidants, age-related macular degeneration (AMD), age-related neurodegenerative diseases
1. Introduction
Major carotenoids that are present in the human diet can be divided into two groups, carotenes (nonpolar carotenoids, such as β-carotene, α-carotene and lycopene) and xanthophylls (polar carotenoids, such as lutein, zeaxanthin and β-cryptoxanthin) (see Figure 1 for their structures). Xanthophylls form less than 20% of the total carotenoids in the human diet. Already in the blood plasma, the amount of xanthophylls increases to about 40% (Figure 2A) [1]. It should be noted that the serum xanthophyll-to-carotenoid ratio varies among the population and depends on diet. This preferential intake of xanthophylls is further enhanced at the level of neural tissues. Xanthophylls are preferentially accumulated from circulation into the brain [1,2] and retina [3,4]. Recently, several carotenoids (including lutein, zeaxanthin, β-cryptoxanthin, α-cryptoxanthin, β-carotene, α-carotene and lycopene) have been identified in the human brain [2]. Surprisingly, xanthophylls are the most abundant carotenoids in this tissue; the evaluation by Craft et al. [2] indicates that xanthophylls account for about 65% of total carotenoids in all brain regions. Similar preferential accumulation of up to 72% of xanthophylls from circulation into the brain was reported by Johnson et al. [1]. Thus, brain tissue, similar to the retina, preferentially accumulates macular xanthophylls (lutein and zeaxanthin); additionally, it accumulates cryptoxanthin, which is not present in the retina. This preferential accumulation is maximal in the case of the retina, where only xanthophylls are present [3,4]. Figure 2A illustrates the enhanced tissue preferences and selectivity for the accumulation of xanthophylls.
Figure 1.

Chemical structures of carotenoids (xanthophylls and carotenes) abundant in food, blood plasma and neural tissue.
Figure 2.

(A) Preferential accumulation of xanthophylls in the brain and retina tissues expressed as a percent of xanthophylls in the total carotenoid pool; (B) preferential accumulation of zeaxanthin over lutein in the brain and retina tissues expressed as the zeaxanthin-to-lutein ratio. Data adapted from [1,2,4,9–12].
Only two dietary carotenoids, namely lutein and zeaxanthin (macular xanthophylls), are selectively accumulated in the human retina. The highest concentration of macular xanthophylls is found in the outer plexiform layer, which is a layer of neuronal synapses between photoreceptor cells and secondary neurons [3,5]. Macular xanthophylls are also present in the layer of rod outer segments [6,7] and in retinal pigment epithelium cells [8]. In addition to the preferential accumulation of xanthophylls from food into the neural tissue, there is a significant increase in the zeaxanthin-to-lutein ratio in neural tissue, as compared to that in the dietary intake of these xanthophylls, and in blood plasma. In human serum, the zeaxanthin-to-lutein ratio ranges from 1:7 to 1:4 [1,9–12], which is consistent with the relatively high lutein content in fruits and vegetables as compared to the content of zeaxanthin.
Zeaxanthin is the dominant xanthophyll in only a few food products, such as the goji berry and orange pepper [13,14]. Thus, the dietary intake of lutein is much higher than that of zeaxanthin, with the evaluated dietary zeaxanthin-to-lutein ratio of 1:12 to 1:5 [11,15,16]. As indicated in Figure 2B, this ratio is increased first on the level of the serum and next when macular xanthophylls are selectively accumulated in the neural tissue. The zeaxanthin-to-lutein ratio in the retina increases to the value of 1:2 in the retina periphery and 2:1 in the central macula [17]. The significant part of the total retina zeaxanthin is represented by its stereoisomer meso-zeaxanthin (see Figure 1 for its structure), which is formed directly in the retina from lutein [17]. The reported zeaxanthin-to-lutein ratio in the human brain ranges from 1:3 to 1:1.4 [1,2]. All of these data suggest that xanthophylls are selectively accumulated and stored in neural tissue and indicate that zeaxanthin (and meso-zeaxanthin) has a particular role in the neural tissue different from that of lutein. It is commonly accepted that both lutein and zeaxanthin are potent lipid-soluble antioxidants. However, the data presented in Figure 2B demonstrate that the retina and brain, which are tissues susceptible to lipid peroxidation, preferentially accumulate zeaxanthin from a human diet poor in zeaxanthin. Furthermore, the synthesis of meso-zeaxanthin in the retina directly from lutein invites the intriguing question: Is the appearance of meso-zeaxanthin in the retina a tissue response to a zeaxanthin-poor diet?
In plants, xanthophylls are present in the pigment-protein complexes of the photosynthetic apparatus [18]. As lipid-soluble molecules, they are also associated with the lipid-bilayer portion of the photosynthetic plant apparatus [18–20], especially under light stress conditions [21]. Their presence in the lipid-bilayer portion of membranes in prokaryotes is well established [22–24]. In these organisms, xanthophylls are associated with membranes subjected to light and oxygen stress. These membranes are particularly rich in polyunsaturated lipids and, under oxidative stress, produce high levels of reactive oxygen species (ROS). These two features of plant thylakoid and prokaryote membranes are shared with the retina photoreceptor segment and neuron membranes. The light harvesting system of plants contains about 75% polyunsaturated lipids [25]. Similarly, photoreceptors of the retina and the neuronal endings and synaptosomes are very rich in long-chain polyunsaturated fatty acids (PUFAs), especially docosahexaenoic acid (DHA) [26–28]. In all of these tissues, only polar carotenoids, namely xanthophylls, are present as lipid-soluble antioxidants. Functional association of xanthophylls as a lipid-soluble antioxidant in membranes rich in polyunsaturated phospholipids was shown by Jagannadham et al. [24] for Antarctic bacteria. To maintain membrane fluidity when grown at a low temperature, these bacteria synthesize a greater proportion of unsaturated fatty acids, which correlates with the synthesis of zeaxanthin.
All of the above indicates that xanthophylls (polar carotenoids) in contrast to carotenes (nonpolar carotenoids) are effectively involved in the protection of lipids in biological membranes rich in PUFAs, which are especially susceptible to oxidative destruction. Epidemiological studies indicate that a xanthophyll-rich diet and xanthophyll supplementation can impede the onset of age-related neurodegenerative diseases, such as age-related macular degeneration (AMD), Alzheimer’s disease and dementia [2,29–33]. The most accepted mechanism through which xanthophylls protect the neural tissue against degenerative diseases is their action as lipid-soluble antioxidants [34–37]. The direct and indirect antioxidant actions of xanthophylls involve blue light filtration [38,39], quenching of singlet oxygen [40] and scavenging of free radicals [41–43]. In vitro, these abilities of xanthophylls are not significantly better than those of other carotenoids. Therefore, it must be some specific property of these xanthophylls that could help explain their preferential (brain) or selective (retina) presence in the human neural tissue and why nature has chosen them from more than 20 other carotenoids present in blood plasma. Norman Krinsky in his reviews [44,45] indicated that one such property is the disposition of xanthophylls in biological membranes. This review is responsive to those articles with the central hypothesis that states: the high membrane solubility, preferential transmembrane orientation and selective concentration of xanthophylls in the most vulnerable regions of lipid bilayer membranes enriched in polyunsaturated lipids distinguish them from other dietary carotenoids, enhance their chemical and physical stability in retina and brain membrane and maximizes their protective action in these organs. We cannot rule out the possible function of specific xanthophyll-binding proteins that can enhance the selective uptake and deposition of macular xanthophylls in the retina and brain. Both interactions of macular xanthophylls with membranes and specific proteins are significant. However, in this review, we will discuss mainly xanthophyll-membrane interactions.
2. High Membrane Solubility and Transmembrane Location
Carotenoids are transported in human blood plasma exclusively by lipoproteins. The segregation of polar (xanthophylls) and nonpolar (carotenes) carotenoid molecules already occurs on the level of carotenoid transport, where carotene molecules are associated primarily with the low-density lipoproteins (LDLs) and xanthophylls are carried by the high-density lipoproteins (HDLs) [46,47]. The reported ratio of lutein in HDLs to lutein in LDLs is approximately 3:1, whereas the ratio of lycopene in HDLs to lycopene in LDLs is about 1:2 [48]. Probably, the HDL transport is critical for the delivery of xanthophylls to the neural tissue. This statement is supported by results obtained by Connor et al. [49], which showed the correlation between HDL deficiency and macular xanthophylls’ deposition in the chicken retina. HDL-deficient mutant chickens fed a high-lutein diet accumulated less lutein and zeaxanthin in the retinal tissue than control chickens fed the same diet. The proportion of the surface phospholipids to the core lipids (cholesteryl esters and triglycerides) in HDLs and LDLs is different. In HDLs, the ratio of phospholipids to core lipids is 1.4:1, whereas in LDLs, this ratio is 0.3:1 [48]. Furthermore, Borel et al. [48] have suggested that this initial segregation of carotenoids is a consequence of the preferential solubility of xanthophylls in phospholipids. HDLs are also enriched in phospholipids containing PUFAs [50]. We think that the association of xanthophylls with HDLs is a result of their high solubility in a PUFA environment. This statement is supported by the findings of Delyfer et al. [51], which showed that a high plasma level of total PUFAs correlates with the high concentration of xanthophylls in the retina. Additional evidence for the correlation between retinal carotenoids and lipids in HDLs has been provided by Renzi et al. [52].
Macular xanthophylls are very soluble in lipid bilayer membranes. The reported solubility thresholds in fluid-phase phospholipid model membranes are in the area of 10 mol% for zeaxanthin and 15 mol% for lutein [53]. A value of 5 mol% also was reported for zeaxanthin incorporated into unilamellar vesicles formed with dipalmitoylphosphatidylcholine [54]. An even lower solubility threshold in lipid bilayer membranes (about 1 mol%) was reported for canthaxanthin, a polar carotenoid with keto groups [55]. Nonpolar β-carotene starts to aggregate at a concentration as low as 0.5 mol% [56]. Mono-polar β-cryptoxanthin is less soluble in the phospholipid bilayer than dipolar xanthophylls [57]. This tendency was confirmed by Socaciu et al. [58], who measured the incorporation ratio of different carotenoids in different phospholipid model membranes. They confirmed high incorporation for xanthophylls and low incorporation for β-carotene. All of these indicate that the high membrane solubility of macular xanthophylls is one of the major characteristics that distinguishes them from other dietary carotenoids.
At a high concentration, xanthophylls can significantly affect membrane properties. They significantly shift the main phase transition of phospholipid bilayers to a lower temperature and decrease the cooperativity of the main phase transition [57,59]. The effect of β-carotene on the membrane phase transition is negligible. Xanthophylls also increase the order of phospholipid membranes [57] and decrease alkyl-chain motion in the fluid phase [60]. At a concentration of 10 mol%, they significantly increase the hydrophobicity of the membrane interior [61]. These effects are the strongest for dipolar xanthophylls, significantly weaker for monopolar xanthophylls (β-cryptoxanthin) and negligible for nonpolar carotenoids (β-carotene).
The high incorporation rate of xanthophylls into lipid membranes is reduced when cholesterol is present in the phospholipid bilayer [58,62]. The decline in incorporation was the strongest for polar carotenoid zeaxanthin. We observed a similar tendency when spin-labeled lutein was incorporated into saturated phosphatidylcholine membranes [63]. Spin-labeled lutein was completely insoluble when cholesterol (30 mol%) was present in these membranes. We think that in the phospholipid bilayer, the xanthophyll-cholesterol interaction is weaker than the xanthophyll-phospholipid interaction. Two polar groups of xanthophyll molecules interact with opposite surfaces of the membrane, and its rigid bar-like portion crosses the entire membrane. The cholesterol molecule is located in one leaflet of the bilayer, and its rigid plate-like portion extends to the depth of the seventh to ninth carbon in the lipid bilayer. When these molecules are located next to each other in the phospholipid bilayer, a free space is created in the membrane center. Cholesterol molecules are forced to sink deeper into the bilayer, which is energetically unfavorable, because it allows water to access the hydrophobic surface of alkyl chains (indicated schematically in Figure 3). These unfavorable interactions caused by xanthophylls and cholesterol are avoided by their separation in the phospholipid membranes (see Section 3).
Figure 3.

Schematic drawing showing the location of the xanthophyll molecule in the cholesterol-rich and cholesterol-poor membrane domains. An unfavorable interaction with cholesterol in the cholesterol-rich domain is indicated.
The transmembrane orientation of xanthophylls in phospholipid bilayers is well documented [64–70]. The presence of polar hydroxyl groups at the ends of macular xanthophyll molecules (see Figure 1) ensures their perpendicular or nearly perpendicular orientation in the bilayer (as shown in Figure 3). The transmembrane localization of macular xanthophylls in retinal membranes can also explain their very slow removal from the retina (about 50 days), observed after discontinuation of xanthophyll supplements given to healthy volunteers [71]. Similar effects were also observed by Hammond et al. [72]. For xanthophylls to be stable in membranes, they must anchor at opposite membrane surfaces. Cholesterol molecules, which also are anchored by the −OH group, but at only one membrane surface (located in one membrane leaflet), can be completely replaced in the retina every six to seven days, and in humans, this replacement may be even more rapid [73]. The incorporation yield of macular xanthophylls into liposomes and cells of retinal pigmented epithelium is five- to 10-times greater than the incorporation yield of canthaxanthin and 20- to 40-times greater than the incorporation yield of β-carotene [74]. These observations suggest that anchoring xanthophyll molecules at opposite membrane surfaces is significant not only in enhancing their effects on membrane properties [57,66,69], but also in stabilizing these molecules in membranes of the human retina (shown schematically in Figure 4). It should be noted that Sujak et al. [75] reported results indicating the existence of two orthogonally-oriented pools of lutein in model phosphatidylcholine membranes, one following the orientation of zeaxanthin (perpendicular to the membrane surface) and the second parallel to the membrane surface. Furthermore, Monte Carlo [76] and molecular dynamics [77] simulations of the behavior of lutein in phosphatidylcholine bilayers have shown that these two orientations are very probable. The existence of two orthogonally-oriented pools of lutein in the membrane may enhance its physiological role as a blue light filter. All of the above allowed us to conclude that solubility, orientation and organization of macular xanthophylls in lipid bilayer membranes depend on their structure, as well as on the lipid environment.
Figure 4.

Schematic drawing explaining the physical stability of dipolar xanthophylls in the lipid-bilayer membranes. A hydrophobic barrier across the lipid bilayer is indicated. To remove the dipolar xanthophyll molecule from the bilayer, one of its polar −OH groups has to cross the hydrophobic (energy) barrier. ΔEH is the energy needed to pull the polar −OH group of the dipolar xanthophyll across this barrier.
Interesting conclusions can be made by comparing the antioxidant properties of xanthophylls to the antioxidant properties of other dietary carotenoids investigated in organic solvents and in lipid bilayer membranes. Zeaxanthin and β-carotene show similar antioxidant properties in organic solutions. However, their antioxidant properties differ when incorporated into membranes [78]. Zeaxanthin was shown to react with free radicals slightly more effectively than β-cryptoxanthin and much more effectively than β-carotene [79,80]. β-carotene and lycopene are able to react efficiently only with radicals generated inside the membrane. Xanthophylls, with their hydroxyl groups exposed to an aqueous environment, can also scavenge free radicals generated in the aqueous phase [81]. The above examples allow us to conclude that the presence of polar hydroxyl groups at the ends of xanthophyll molecules and their transmembrane orientation enhance their stability in membranes and their antioxidant properties and, thus, maximize their protective action in biological membranes. Those properties also distinguish xanthophylls from other dietary carotenoids.
3. Location in the Most Vulnerable Regions of Lipid Membranes
In the human retina, the concentration of xanthophylls reaches a level between 0.1 and 1 mM in the central fovea [5,71]. Xanthophylls are accumulated mainly in the region of photoreceptor axons [5], where they act as a blue light filter, and within photoreceptor outer segments (POS) [6,7], where their antioxidant functions are assumed. Although xanthophylls in POS constitute about 10% to 25% of the amount in the entire retina [6,7], the local concentration of xanthophylls in membranes of the rod outer segment is ~70% higher than in residual retina membranes [7]. The concentration of xanthophylls in the human retina is, however, high enough for effective blue light filtration and antioxidant action. To understand the basic mechanisms of these actions, it is necessary to better understand the carotenoid-membrane interaction. For systems with a high carotenoid concentration (in bacteria and plants where the local carotenoid concentration in the lipid bilayer can reach a value of a few mol%), it is most significant to understand how carotenoids affect membrane physical properties, structure and dynamics, as well as the lateral organization of the lipid bilayer (its domain structure). For systems with a low carotenoid concentration, it is especially important to understand how the membrane itself—membrane composition, structure and lateral organization—affects the organization of carotenoids in the lipid bilayer, including their solubility (monomeric versus aggregated state), orientation (transmembrane versus parallel) and localization (distribution between membrane domains). Furthermore, knowledge of the bulk-membrane physical properties, which are not uniform across the lipid bilayer and can differ by membrane domain, is significant to better understand chemical reactions and physical processes that take place in the lipid bilayer membrane and involve carotenoids [82].
Retinal pigment epithelium membranes and disk membranes of rod outer segments are laterally heterogeneous and contain raft domains [83–86]. Raft domains, enriched in cholesterol and sphingolipids, were isolated as detergent-resistant membrane (DRM) fractions. Remaining detergent-soluble membrane (DSM) fractions, formed by bulk lipids surrounding raft domains, were rich in long-chain PUFAs. Additionally, rhodopsin, the main protein of POS membranes that is responsible for the first stages of visual signal transduction and is located in the bulk domain of the POS membrane [87], was isolated mostly with the DSM fraction [83,84,86]. Rafts in photoreceptor cell membranes are involved in the regulation of the G-protein-mediated pathway of phototransduction [85]. Aggregation of small, unstable rafts in bigger platforms (observed, for example, in retinal pigment epithelium cells) is supposed to enhance signal transduction to the cell interior and cause a specific reaction in the cell, such as apoptosis [88].
Rhodopsin requires the presence of polyunsaturated lipids (DHA) for its activity [89–91], and thus, their colocalization is functionally justified. Furthermore, phospholipids containing very long-chain PUFAs (VLC-PUFAs, >C24, with 3 to 9 double bonds) likely play a unique and important role in the retina because they are necessary for cell survival, and their loss leads to cell death [92,93]. It has been suggested that they are tightly bound to rhodopsin and that their unusually long chains may partially surround the α-helical segments of rhodopsin [94]. Epidemiological studies of long-chain polyunsaturated phospholipid intake suggest a protective role against the incidence of advanced AMD [95,96]. It was shown that lipids containing VLC-PUFAs are reduced in the retina of aged eyes and severely reduced in the retina of AMD eyes [97]. As indicated by the authors, their results support the potential value of interventions to increase retinal VLC-PUFAs in the prevention and treatment of AMD. All of these indicate that the colocalization of rhodopsin with polyunsaturated lipids is functionally justified. However, this colocalization creates a dangerous situation for both rhodopsin and polyunsaturated lipids, especially during illumination, when reactive oxygen species can be produced by photosensitizers [98,99]. It should be noted that phospholipids containing long-chain and very long-chain PUFAs, which are involved in preventing AMD, are also very prone to oxidative damage. Fortunately, to protect the retina against oxidative damage, nature has used xanthophylls as an effective protector that can absorb damaging blue light, neutralize photosensitizers and reactive oxygen species and scavenge free radicals.
Our investigations, made on a model of POS membranes, indicate that xanthophylls were about 14-times more concentrated in the unsaturated bulk domain (enriched in polyunsaturated lipids and isolated as DSM) and excluded from the raft domain (enriched in saturated lipids and cholesterol and isolated as DRM) [100]. A similar distribution also was found in membranes made of a raft-forming mixture where macular xanthophylls lutein and zeaxanthin were about eight-times more concentrated in the bulk, unsaturated domain than in the raft domain [101]. A similar distribution has been observed for monopolar β-cryptoxanthin, but not for nonpolar β-carotene, which was more uniformly distributed between the DRM and DSM domains. Results indicating the distribution of carotenoids between raft and bulk domains are summarized in Figure 5. Colocalization of xanthophyll molecules with polyunsaturated lipids is ideal if xanthophylls are to act as a lipid antioxidant, which is the most accepted mechanism through which lutein and zeaxanthin protect the retina from AMD [44,45,102]. It should significantly enhance the effectiveness of xanthophylls, especially when the local concentration of xanthophylls in the membrane is not very high. Studies by Wisniewska-Becker et al. [103] show that preferential colocalization of xanthophylls with polyunsaturated lipids really enhances their antioxidant activity. The observed inhibition of lipid peroxidation by lutein was significantly greater in membranes containing raft domains than in homogenous membranes.
Figure 5.
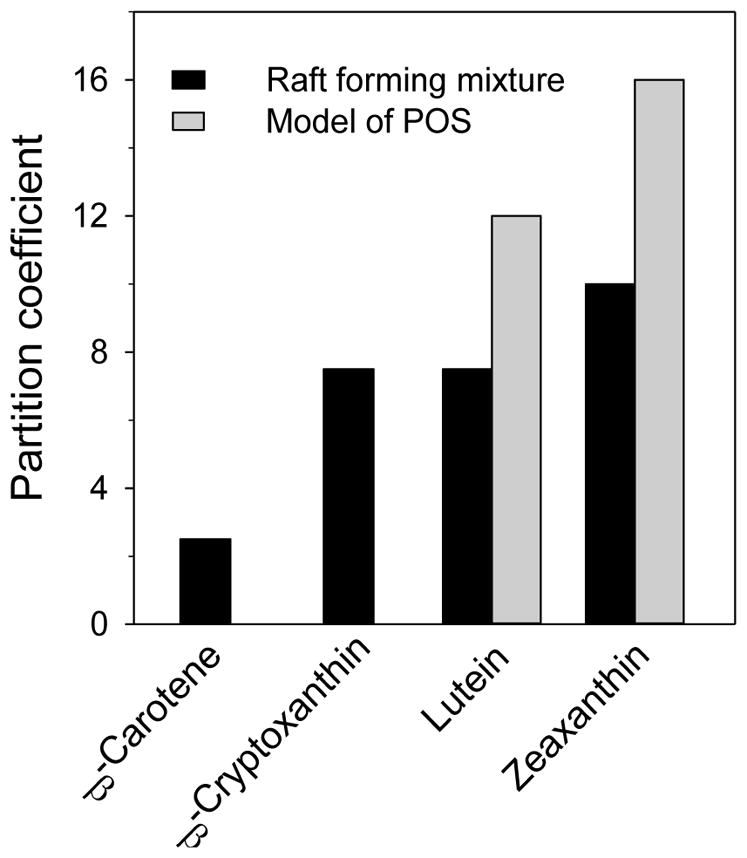
Partition coefficient of dipolar xanthophylls (lutein and zeaxanthin), monopolar xanthophyll (β-cryptoxanthin) and nonpolar carotenoid (β-carotene) between the bulk (unsaturated) domain and the raft (saturated) domain in the membrane made of the raft-forming mixture and in the model of photoreceptor outer segment (POS) membranes. The unsaturated domain in the model of POS membranes is abundant in DHA, with six double bonds. For more details, see [100,101].
4. Photo-Related and Dark Antioxidant Action
It is commonly believed that xanthophylls protect eye retinas mainly through photo-related antioxidant action, while in the brain, the dark antioxidant action of xanthophylls is expected. In this section, we will describe the mechanisms of both actions, which are summarized in the diagram in Figure 6.
Figure 6.

Diagram illustrating processes through which xanthophylls are protecting membranes against oxidative damage. Photo-related and dark processes are actively involved in protecting the retina, while only dark processes are assumed to be active in the protection of brain tissue. The broken rectangles indicate processes that are not yet fully confirmed as involved in protecting the eye retina and are included here as purported processes.
In the retina, macular xanthophylls are localized mostly in Henle’s layer [64] and form a filter for blue light. Although most ultraviolet is absorbed by the cornea [104] and lens [105], some fraction of blue radiation reaches the retina, may activate potent retinal photosensitizers (such as all-trans retinal, cytochrome c oxidase, porphyrins and the major chromophore of lipofuscin, bisretinoid A2E [99,106–109]) and, consequently, generates reactive oxygen species. Thus, blue light absorption can be considered an indirect antioxidant action, because it prevents blue light from generating reactive oxygen species that can damage photoreceptor cells. Blue light absorption by macular xanthophylls is extremely important for young eyes, for which lens transparency is almost 95%. During aging, the lens gradually loses its transparency, becomes yellowish [105] and better filters UV and blue light. Thus, in older age, the blue light filtration performed by macular xanthophylls becomes relatively less important.
The blue light filtration antioxidant mechanism is based on the high extinction coefficient of macular xanthophylls for the absorption spectrum band of 390 to 540 nm with maximum absorption at around 450 nm. The most damaging impact on the retina has been shown for exposure to high-energy light with maximum absorption at around 440 nm [99,110], which is also maximum absorption for xanthophyll molecules. Xanthophylls dissolved in lipid bilayers may exist not only in the form of monomers, but also in aggregate arrangements. Two formed aggregates are well known: the “card-pack” arrangement (H-aggregate), where the shift to a shorter wavelength is observed (blue shift), and the “head-to-tail” organization (J-aggregate), with the shift to a longer wavelength (red shift). Typically, in phospholipid membranes, macular xanthophylls are present as monomers or can form H-aggregates with a blue-shifted absorption spectrum and maximum absorption at 380 nm. Sujak et al. [54] observed that, in lipid bilayers, lutein has a higher degree of aggregation than zeaxanthin and, because of that, possesses a wider bandwidth in UV and blue light absorption. Interestingly, infant retinas, which have more exposure to blue and ultraviolet light, have more lutein and less zeaxanthin relative to adult retinas [17]. Better filtration of blue light by lutein may justify its unique distribution in the adult retina and its high concentration in the infant retina. Junghans et al. [111] have investigated the blue light filter efficiency of four plasma carotenoids (lutein, zeaxanthin, β-carotene and lycopene) incorporated into membranes of liposomes loaded with a hydrophilic fluorescent dye, Lucifer yellow, excitable by blue light. Fluorescent emission of the dye was lower in liposomes with carotenoids as compared to the control, indicating a filter effect. Macular xanthophylls lutein and zeaxanthin exhibited the highest blue light absorption activity as compared to liposomes containing nonpolar carotenoids. Blue-light filter efficacy is ordered as lutein > zeaxanthin > β-carotene > lycopene. These results indicate that xanthophylls (especially lutein) are the best blue light filters among all carotenoids available in blood plasma.
In the retina, macular xanthophylls may not only act as blue light filters, but may also optimize visual performance. The layer of macular xanthophylls is believed to reduce chromatic aberrations, glare disability and light scattering, which enhance vision contrast [112,113]. This way, xanthophylls can positively affect the cognitive function of the brain, especially in the aged population [114–116].
Xanthophylls are capable of quenching excited triplet states of potent singlet oxygen photosensitizers. These quenching processes belong to the direct photo-related antioxidant actions of xanthophylls. By this mechanism, the largest part of excess energy is transferred from potentially harmful triplets of photosensitizers to xanthophylls and dissipated as heat. This mechanism is well established for porphyrins, but has not yet been clearly confirmed to work for photosensitizers in the retina. In the retina, photo activation of rhodopsin leads to isomerization of its chromophore, 11-cis-retinal to all-trans-retinal, which, under certain conditions, can act as a photosensitizer. Free all-trans-retinal may absorb light and transfer energy from its excited triplet state to molecular oxygen, generating singlet oxygen [98]. It is postulated that the close proximity of xanthophylls, which are also located in the bulk domain (see Section 3), allows effective energy transfer from excited all-trans retinal to xanthophyll and prevents singlet oxygen generation by this photosensitizer [117].
Carotenoids have been known to be the most effective singlet oxygen quenchers, and their activities are much higher than that of other retinal antioxidants, tocopherols and thiols [118,119]. They are able to quench singlet oxygen by two different mechanisms. The first mechanism, which involves energy transfer, is termed physical quenching and is considered the major pathway of singlet oxygen deactivation. According to this mechanism, carotenoid molecules deactivate singlet oxygen to the nonreactive triplet state. During that process, carotenoid molecules become excited to the triplet state and can return to the ground state, dissipating the excess energy as heat. The profit of the physical quenching is that carotenoids may act without alternation of their own chemical structure. The second mechanism is called chemical quenching. It involves a chemical reaction between carotenoid and singlet oxygen, which results in pigment autoxidation. The capacity of major plasma carotenoids to quench singlet oxygen in an organic solvent mainly depends on the number of conjugated double bonds in the chromophore, but also varies with functional groups [120]. Thus, zeaxanthin (with its 11 conjugated double bonds) has a higher ability to quench singlet oxygen than lutein (with its 10 conjugated double bonds) (see Figure 1). The inactivation of singlet oxygen may also occur through chemical quenching involving autoxidation of the carotenoids, a process that consumes the carotenoids themselves. Chemical quenching is reported to contribute less than 0.05% to the overall singlet oxygen quenching by carotenoids [121]. Nevertheless, it has been reported that the retina contains macular xanthophyll metabolites, indicating that chemical quenching can take place in biological tissues [122,123]. The degradation of four major plasma carotenoids, induced by UV light in the presence of rose bengal, has been studied by van Kuijk and his co-workers [124,125]. Higher degradation rates were found for nonpolar carotenoids as compared to macular xanthophylls. Furthermore, studies of the autoxidation of carotenoids incorporated in pig liver microsomes [126] give similar results: nonpolar carotenoids, such as β-carotene and lycopene, had degraded totally, whereas polar carotenoids had degraded much slower, and zeaxanthin was shown to be the most stable carotenoid. We can conclude that the high chemical stability of macular xanthophylls distinguishes them from other dietary carotenoids.
The conjugated double bond system is primarily responsible for the high chemical reactivity of carotenoids, not only with singlet oxygen [118,120], but also with free radicals [127]. Carotenoids are capable of intercepting peroxyl radical and inhibiting phospholipid peroxidation. Selective localization of xanthophylls in domains rich in polyunsaturated phospholipids (see Section 3) and, therefore, susceptible to free-radical-induced damage is ideal for their chemical dark antioxidant action. Carotenoids scavenge lipid peroxyl radicals by forming radical adducts [127] that are less reactive than lipid alkyl peroxyl radicals. Thus, carotenoids are effective chain-breaking antioxidants, which delay the oxidation of biological membranes by trapping the chain-initiating or chain-propagating peroxyl radicals. This dark antioxidant action should form the major mechanism through which xanthophylls protect brain tissue against neurodegenerative diseases. However, direct mechanisms of xanthophyll actions in the brain are not yet investigated in detail and not yet defined.
5. Concluding Remarks
Carotenoids are important dietary prophylactic agents in numerous degenerative diseases. Among them, macular xanthophylls play a unique position in their influence on the health and functions of the retina and brain. Their selective presence throughout the central nervous system tissue has led to the questions of not only how these dietary carotenoids influence the retina and brain, but also why nature, during the evolutionary process, chose macular xanthophylls from other carotenoids available in the human diet to perform those specific functions. In this review, we informed researchers about the importance of the xanthophyll-membrane interactions that enhance their chemical and physical stability in the retina and brain membranes and maximize their protective action in these organs.
In this section, we make additional comments about the selective accumulation of macular xanthophylls, lutein and zeaxanthin, in the outer plexiform layer (which comprises a dense network of synapses in the retina) [64]. It is commonly accepted that the primary function for these xanthophylls is to provide a filter for blue light [111]. We think that equally significant is the direct antioxidant action of xanthophylls to protect membranes of neuronal endings and synaptosomes, which are rich in polyunsaturated phospholipids. In a similar manner, xanthophylls can act in the phospholipid environment of the grey matter of the brain, which is rich in PUFAs [128,129]. Interestingly, in these two neural tissues, only polar carotenoids (namely xanthophylls) are present as lipid-soluble antioxidants. Embryologically, the retina is part of the brain. Additionally, polyunsaturated phospholipids, especially DHA, are present in high amounts in the retina (rod outer segment and synaptic area) and in the brain, primarily in synaptosomes [93,130–132]. Retina and brain tissues are also very well oxygenated, which makes them particularly accessible to oxidative damage. Thus, they need relatively stable lipid-soluble antioxidants for their protection. Data indicate that xanthophylls serve this function well.
Throughout this review, we indicated that there are several physical and chemical properties that distinguish xanthophylls, especially macular xanthophylls, from other carotenoids available in the human diet as lipid-soluble antioxidants of neural tissues. They include high membrane solubility, preferential transmembrane orientation and selective concentration in the polyunsaturated lipid environment. Xanthophylls also have greater resistance to autoxidation than carotenes, which is an important feature, especially for neural tissue, where turnover of the antioxidant pole is slow. All of these unique properties enhance xanthophylls’ chemical and physical stability in biological membranes and maximize their protective actions. Less chemically-stable carotenes can be more easily oxidized in lipid bilayer membranes [126] and can even function as prooxidant compounds [133,134]. These unique properties were “recognized” during evolution when mechanisms for the selective accumulation of xanthophylls in the retina and brain were developed. These mechanisms are not yet understood in detail. Some suggestions about selective xanthophyll transport are included in Section 2. Selective accumulation suggests involvement of specific xanthophyll-binding proteins. Some of these membrane-associated, xanthophyll-binding proteins already have been identified and characterized, including zeaxanthin-binding protein [135] and lutein-binding protein [136]. The unanswered question is whether these proteins are only selective transporters of macular xanthophylls or whether they can store xanthophylls.
It is clear from Figure 2A that evolution selected xanthophylls, including lutein and zeaxanthin, to play their protective role in neural tissue. However, we found very intriguing the fact that the human diet is poor in zeaxanthin and relatively rich in lutein, while comparatively, the amount of zeaxanthin in the retina and brain is significantly enhanced (see Figure 2). Zeaxanthin, with its 11 double conjugated bonds, has a higher ability to quench singlet oxygen than lutein, with its 10 conjugated double bonds [120] (see Figure 1). This property of zeaxanthin as the best lipid-soluble antioxidant among xanthophylls was recognized during evolution. We also think that the evolutionary selection of zeaxanthin as the best lipid-soluble antioxidant in different organisms (bacteria, plants, humans) and tissues (human retina, human brain) is associated with the requirement to protect the highly unsaturated regions of the lipid membranes. To compensate for the poor presence of zeaxanthin in the human diet, nature developed a mechanism to transform lutein, which is the most abundant xanthophyll in the diet, to an isomer of zeaxanthin, namely meso-zeaxanthin, directly in the retina (see Figure 1 for the chemical structures). The detailed mechanism of this isomerization process is unknown; however, we think that the nature of the overall strategy (appearance of zeaxanthin molecules in phospholipid environment susceptible to oxidation) used here is similar to that in plants during the violaxanthin cycle [137]. During this cycle, violaxanthin is partially transformed to zeaxanthin through chemical de-epoxidation, whereas during isomerization of lutein in the retina, rearrangement of bonds within the functional groups occurs [17,138]. We can summarize that the distribution of xanthophylls in human tissues and organs is biologically controlled by their solubility, specific binding proteins (this control is structure dependent, distinguishing zeaxanthin and lutein) and the site of formation (as in the case of meso-zeaxanthin).
The neuronal conduction and synaptic neurotransmission is affected by the membrane state. Neuronal lipid bilayer membranes in the brain and retina are especially vulnerable to oxidative damage because of their enrichment in PUFAs and exposure to high oxygen concentration. Only dipolar xanthophylls, with their high solubility in polyunsaturated phospholipid membrane domains and high resistance to autoxidation, serve the role of protector in these neural membranes. Lutein and zeaxanthin are present at a high concentration in the synaptosomal membranes of the outer plexiform layer, which links the brain and retina tissues. Although the most accepted function of these xanthophylls is blue light filtration, we postulate that equally significant is the protection of these highly unsaturated membranes through the trapping of chain-initiating and/or chain-propagating peroxyl radicals. Through this function, xanthophylls can effectively protect the neuronal synapses region in both the retina and brain tissues. This dark antioxidant action is indicated in Figure 6.
Acknowledgments
This work was supported by Grants EY015526, EB002052, EB001980 and EY001931 from the National Institutes of Health.
Footnotes
Author Contributions: J.W., M.Z. and W.K.S. all contributed to the preparation of the manuscript. All authors reviewed the manuscript.
Conflicts of Interest: The authors declare no conflict of interest.
Contributor Information
Mariusz Zareba, Email: mariusz@mcw.edu.
Witold Karol Subczynski, Email: subczyn@mcw.edu.
References
- 1.Johnson EJ, Vishwanathan R, Johnson MA, Hausman DB, Davey A, Scott TM, Green RC, Miller LS, Gearing M, Woodard J, et al. Relationship between Serum and Brain Carotenoids, alpha-Tocopherol, and Retinol Concentrations and Cognitive Performance in the Oldest Old from the Georgia Centenarian Study. J Aging Res. 2013;2013:951786. doi: 10.1155/2013/951786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Craft NE, Haitema TB, Garnett KM, Fitch KA, Dorey CK. Carotenoid, tocopherol, and retinol concentrations in elderly human brain. J Nutr Health Aging. 2004;8:156–162. [PubMed] [Google Scholar]
- 3.Bone RA, Landrum JT, Tarsis SL. Preliminary identification of the human macular pigment. Vis Res. 1985;25:1531–1535. doi: 10.1016/0042-6989(85)90123-3. [DOI] [PubMed] [Google Scholar]
- 4.Bone RA, Landrum JT, Fernandez L, Tarsis SL. Analysis of the macular pigment by HPLC: Retinal distribution and age study. Investig Ophthalmol Vis Sci. 1988;29:843–849. [PubMed] [Google Scholar]
- 5.Snodderly DM, Auran JD, Delori FC. The macular pigment. II. Spatial distribution in primate retinas. Investig Ophthalmol Vis Sci. 1984;25:674–685. [PubMed] [Google Scholar]
- 6.Rapp LM, Maple SS, Choi JH. Lutein and zeaxanthin concentrations in rod outer segment membranes from perifoveal and peripheral human retina. Investig Ophthalmol Vis Sci. 2000;41:1200–1209. [PubMed] [Google Scholar]
- 7.Sommerburg OG, Siems WG, Hurst JS, Lewis JW, Kliger DS, van Kuijk FJ. Lutein and zeaxanthin are associated with photoreceptors in the human retina. Curr Eye Res. 1999;19:491–495. doi: 10.1076/ceyr.19.6.491.5276. [DOI] [PubMed] [Google Scholar]
- 8.Bernstein PS, Khachik F, Carvalho LS, Muir GJ, Zhao DY, Katz NB. Identification and quantitation of carotenoids and their metabolites in the tissues of the human eye. Exp Eye Res. 2001;72:215–223. doi: 10.1006/exer.2000.0954. [DOI] [PubMed] [Google Scholar]
- 9.Bone RA, Landrum JT, Dixon Z, Chen Y, Llerena CM. Lutein and zeaxanthin in the eyes, serum and diet of human subjects. Exp Eye Res. 2000;71:239–245. doi: 10.1006/exer.2000.0870. [DOI] [PubMed] [Google Scholar]
- 10.Curran-Celentano J, Hammond BR, Jr, Ciulla TA, Cooper DA, Pratt LM, Danis RB. Relation between dietary intake serum concentrations and retinal concentrations of lutein zeaxanthin in adults in a Midwest population. Am J Clin Nutr. 2001;74:796–802. doi: 10.1093/ajcn/74.6.796. [DOI] [PubMed] [Google Scholar]
- 11.Olmedilla-Alonso B, Beltran-de-Miguel B, Estevez-Santiago R, Cuadrado-Vives C. Markers of lutein and zeaxanthin status in two age groups of men and women: Dietary intake, serum concentrations, lipid profile and macular pigment optical density. Nutr J. 2014;13:52. doi: 10.1186/1475-2891-13-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wang W, Connor SL, Johnson EJ, Klein ML, Hughes S, Connor WE. Effect of dietary lutein and zeaxanthin on plasma carotenoids and their transport in lipoproteins in age-related macular degeneration. Am J Clin Nutr. 2007;85:762–769. doi: 10.1093/ajcn/85.3.762. [DOI] [PubMed] [Google Scholar]
- 13.Peng Y, Ma C, Li Y, Leung KS, Jiang ZH, Zhao Z. Quantification of zeaxanthin dipalmitate and total carotenoids in Lycium fruits (Fructus Lycii) Plant Foods Hum Nutr. 2005;60:161–164. doi: 10.1007/s11130-005-9550-5. [DOI] [PubMed] [Google Scholar]
- 14.Sommerburg O, Keunen JE, Bird AC, van Kuijk FJ. Fruits and vegetables that are sources for lutein and zeaxanthin: The macular pigment in human eyes. Br J Ophthalmol. 1998;82:907–910. doi: 10.1136/bjo.82.8.907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Beatty S, Nolan J, Kavanagh H, O’Donovan O. Macular pigment optical density and its relationship with serum and dietary levels of lutein and zeaxanthin. Arch Biochem Biophys. 2004;430:70–76. doi: 10.1016/j.abb.2004.03.015. [DOI] [PubMed] [Google Scholar]
- 16.Nebeling LC, Forman MR, Graubard BI, Snyder RA. Changes in carotenoid intake in the United States: The 1987 and 1992 National Health Interview Surveys. J Am Diet Assoc. 1997;97:991–996. doi: 10.1016/S0002-8223(97)00239-3. [DOI] [PubMed] [Google Scholar]
- 17.Bone RA, Landrum JT, Friedes LM, Gomez CM, Kilburn MD, Menendez E, Vidal I, Wang W. Distribution of lutein and zeaxanthin stereoisomers in the human retina. Exp Eye Res. 1997;64:211–218. doi: 10.1006/exer.1996.0210. [DOI] [PubMed] [Google Scholar]
- 18.Demmig-Adams B, Gilmore AM, Adams WW., III Carotenoids 3: In vivo function of carotenoids in higher plants. FASEB J. 1996;10:403–412. doi: 10.1096/fasebj.10.4.8647339. [DOI] [PubMed] [Google Scholar]
- 19.Niyogi KK, Bjorkman O, Grossman AR. The roles of specific xanthophylls in photoprotection. Proc Natl Acad Sci USA. 1997;94:14162–14167. doi: 10.1073/pnas.94.25.14162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Havaux M. Carotenoids as membrane stabilizers in chloroplasts. Trends Plant Sci. 1998;3:147–151. [Google Scholar]
- 21.Sarry JE, Montillet JL, Sauvaire Y, Havaux M. The protective function of the xanthophyll cycle in photosynthesis. FEBS Lett. 1994;353:147–150. doi: 10.1016/0014-5793(94)01028-5. [DOI] [PubMed] [Google Scholar]
- 22.Chamberlain NR, Mehrtens BG, Xiong Z, Kapral FA, Boardman JL, Rearick JI. Correlation of carotenoid production, decreased membrane fluidity, and resistance to oleic acid killing in Staphylococcus aureus 18Z. Infect Immun. 1991;59:4332–4337. doi: 10.1128/iai.59.12.4332-4337.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chattopadhyay MK, Jagannadham MV, Vairamani M, Shivaji S. Carotenoid pigments of an antarctic psychrotrophic bacterium Micrococcus roseus: Temperature dependent biosynthesis, structure, and interaction with synthetic membranes. Biochem Biophys Res Commun. 1997;239:85–90. doi: 10.1006/bbrc.1997.7433. [DOI] [PubMed] [Google Scholar]
- 24.Jagannadham MV, Chattopadhyay MK, Subbalakshmi C, Vairamani M, Narayanan K, Rao CM, Shivaji S. Carotenoids of an Antarctic psychrotolerant bacterium, Sphingobacterium antarcticus, and a mesophilic bacterium, Sphingobacterium multivorum. Arch Microbiol. 2000;173:418–424. doi: 10.1007/s002030000163. [DOI] [PubMed] [Google Scholar]
- 25.McConn M, Browse J. Polyunsaturated membranes are required for photosynthetic competence in a mutant of Arabidopsis. Plant J. 1998;15:521–530. doi: 10.1046/j.1365-313x.1998.00229.x. [DOI] [PubMed] [Google Scholar]
- 26.Acar N, Berdeaux O, Gregoire S, Cabaret S, Martine L, Gain P, Thuret G, Creuzot-Garcher CP, Bron AM, Bretillon L. Lipid composition of the human eye: Are red blood cells a good mirror of retinal and optic nerve fatty acids? PLoS ONE. 2012;7:e35102. doi: 10.1371/journal.pone.0035102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Salem N, Jr, Kim H, Yergey JA. Docosahexoenoic acid: Membrane function and metabolism. In: Simopolous AP, Kifer RR, Martin RE, editors. Health Effects of Polyunsaturated Fatty Acids in Seafoods. Academic Press; New York, NY, USA: 1986. pp. 263–317. [Google Scholar]
- 28.Ikemoto A, Kobayashi T, Watanabe S, Okuyama H. Membrane fatty acid modifications of PC12 cells by arachidonate or docosahexaenoate affect neurite outgrowth but not norepinephrine release. Neurochem Res. 1997;22:671–678. doi: 10.1023/a:1027393724676. [DOI] [PubMed] [Google Scholar]
- 29.The Age-Related Eye Disease Study 2 (AREDS2) Research Group. Lutein + zeaxanthin and omega-3 fatty acids for age-related macular degeneration: The Age-Related Eye Disease Study 2 (AREDS2) randomized clinical trial. JAMA. 2013;309:2005–2015. doi: 10.1001/jama.2013.4997. [DOI] [PubMed] [Google Scholar]
- 30.Johnson EJ. Role of lutein and zeaxanthin in visual and cognitive function throughout the lifespan. Nutr Rev. 2014;72:605–612. doi: 10.1111/nure.12133. [DOI] [PubMed] [Google Scholar]
- 31.Mares-Perlman JA, Millen AE, Ficek TL, Hankinson SE. The body of evidence to support a protective role for lutein and zeaxanthin in delaying chronic disease. Overview. J Nutr. 2002;132:518S–524S. doi: 10.1093/jn/132.3.518S. [DOI] [PubMed] [Google Scholar]
- 32.Ribaya-Mercado JD, Blumberg JB. Lutein and zeaxanthin and their potential roles in disease prevention. J Am Coll Nutr. 2004;23:567S–587S. doi: 10.1080/07315724.2004.10719427. [DOI] [PubMed] [Google Scholar]
- 33.Berr C, Balansard B, Arnaud J, Roussel AM, Alperovitch A. Cognitive decline is associated with systemic oxidative stress: The EVA study. Etude du Vieillissement Arteriel. J Am Geriatr Soc. 2000;48:1285–1291. doi: 10.1111/j.1532-5415.2000.tb02603.x. [DOI] [PubMed] [Google Scholar]
- 34.Martin HD, Ruck C, Schmidt M, Sell S, Beutner S, Mayer B, Walsh R. Chemistry of carotenoid oxidation and free radical reactions. Pure Appl Chem. 1999;71:2253–2262. [Google Scholar]
- 35.Bohm F, Edge R, Truscott G. Interactions of dietary carotenoids with activated (singlet) oxygen and free radicals: Potential effects for human health. Mol Nutr Food Res. 2012;56:205–216. doi: 10.1002/mnfr.201100222. [DOI] [PubMed] [Google Scholar]
- 36.Chucair AJ, Rotstein NP, Sangiovanni JP, During A, Chew EY, Politi LE. Lutein and zeaxanthin protect photoreceptors from apoptosis induced by oxidative stress: Relation with docosahexaenoic acid. Investig Ophthalmol Vis Sci. 2007;48:5168–5177. doi: 10.1167/iovs.07-0037. [DOI] [PubMed] [Google Scholar]
- 37.Moeller SM, Jacques PF, Blumberg JB. The potential role of dietary xanthophylls in cataract and age-related macular degeneration. J Am Coll Nutr. 2000;19:522S–527S. doi: 10.1080/07315724.2000.10718975. [DOI] [PubMed] [Google Scholar]
- 38.Jaffe GJ, Wood IS. Retinal phototoxicity from the operating microscope: A protective effect by the fovea. Arch Ophthalmol. 1988;106:445–446. doi: 10.1001/archopht.1988.01060130487004. [DOI] [PubMed] [Google Scholar]
- 39.Kirschfeld K. Carotenoid pigments: Their possible role in protecting against photooxidation in eyes and photoreceptor cells. Proc R Soc B Biol Sci. 1982;216:71–85. doi: 10.1098/rspb.1982.0061. [DOI] [PubMed] [Google Scholar]
- 40.Cantrell A, McGarvey DJ, Truscott TG, Rancan F, Bohm F. Singlet oxygen quenching by dietary carotenoids in a model membrane environment. Arch Biochem Biophys. 2003;412:47–54. doi: 10.1016/s0003-9861(03)00014-6. [DOI] [PubMed] [Google Scholar]
- 41.Stahl W, Junghans A, de Boer B, Driomina ES, Briviba K, Sies H. Carotenoid mixtures protect multilamellar liposomes against oxidative damage: Synergistic effects of lycopene and lutein. FEBS Lett. 1998;427:305–308. doi: 10.1016/s0014-5793(98)00434-7. [DOI] [PubMed] [Google Scholar]
- 42.Trevithick-Sutton CC, Foote CS, Collins M, Trevithick JR. The retinal carotenoids zeaxanthin and lutein scavenge superoxide and hydroxyl radicals: A chemiluminescence and ESR study. Mol Vis. 2006;12:1127–1135. [PubMed] [Google Scholar]
- 43.Lim BP, Nagao A, Terao J, Tanaka K, Suzuki T, Takama K. Antioxidant activity of xanthophylls on peroxyl radical-mediated phospholipid peroxidation. Biochim Biophys Acta. 1992;1126:178–184. doi: 10.1016/0005-2760(92)90288-7. [DOI] [PubMed] [Google Scholar]
- 44.Krinsky NI. Possible biologic mechanisms for a protective role of xanthophylls. J Nutr. 2002;132:540S–542S. doi: 10.1093/jn/132.3.540S. [DOI] [PubMed] [Google Scholar]
- 45.Krinsky NI, Landrum JT, Bone RA. Biologic mechanisms of the protective role of lutein and zeaxanthin in the eye. Annu Rev Nutr. 2003;23:171–201. doi: 10.1146/annurev.nutr.23.011702.073307. [DOI] [PubMed] [Google Scholar]
- 46.Loane E, Nolan JM, O’Donovan O, Bhosale P, Bernstein PS, Beatty S. Transport and retinal capture of lutein and zeaxanthin with reference to age-related macular degeneration. Surv Ophthalmol. 2008;53:68–81. doi: 10.1016/j.survophthal.2007.10.008. [DOI] [PubMed] [Google Scholar]
- 47.Parker RS. Absorption, metabolism, and transport of carotenoids. FASEB J. 1996;10:542–551. [PubMed] [Google Scholar]
- 48.Borel P, Grolier P, Armand M, Partier A, Lafont H, Lairon D, Zais-Braesco V. Carotenoids in biological emulsions: Solubility, surface-to-core distribution, and release from lipid droplets. J Lipid Res. 1996;37:250–261. [PubMed] [Google Scholar]
- 49.Connor WE, Duell PB, Kean R, Wang Y. The prime role of HDL to transport lutein into the retina: Evidence from HDL-deficient WHAM chicks having a mutant ABCA1 transporter. Investig Ophthalmol Vis Sci. 2007;48:4226–4231. doi: 10.1167/iovs.06-1275. [DOI] [PubMed] [Google Scholar]
- 50.Kontush A, Lhomme M, Chapman MJ. Unraveling the complexities of the HDL lipidome. J Lipid Res. 2013;54:2950–2963. doi: 10.1194/jlr.R036095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Delyfer MN, Buaud B, Korobelnik JF, Rougier MB, Schalch W, Etheve S, Vaysse C, Combe N, Goff ML, Wolf-Schnurrbusch UE, et al. Association of macular pigment density with plasma omega-3 fatty acids: The PIMAVOSA study. Investig Ophthalmol Vis Sci. 2012;53:1204–1210. doi: 10.1167/iovs.11-8721. [DOI] [PubMed] [Google Scholar]
- 52.Renzi LM, Hammond BR, Jr, Dengler M, Roberts R. The relation between serum lipids and lutein and zeaxanthin in the serum and retina: Results from cross-sectional case-control and case study designs. Lipids Health Dis. 2012;11:33. doi: 10.1186/1476-511X-11-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sujak A, Gruszecki WI. Organization of mixed monomolecular layers formed with the xanthophyll pigments lutein or zeaxanthin and dipalmitoylphosphatidylcholine at the argon-water interface. J Photochem Photobiol B. 2000;59:42–47. doi: 10.1016/s1011-1344(00)00133-0. [DOI] [PubMed] [Google Scholar]
- 54.Sujak A, Okulski W, Gruszecki WI. Organisation of xanthophyll pigments lutein and zeaxanthin in lipid membranes formed with dipalmitoylphosphatidylcholine. Biochim Biophys Acta. 2000;1509:255–263. doi: 10.1016/s0005-2736(00)00299-6. [DOI] [PubMed] [Google Scholar]
- 55.Sujak A, Gabrielska J, Milanowska J, Mazurek P, Strzalka K, Gruszecki WI. Studies on canthaxanthin in lipid membranes. Biochim Biophys Acta. 2005;1712:17–28. doi: 10.1016/j.bbamem.2005.03.010. [DOI] [PubMed] [Google Scholar]
- 56.Kennedy TA, Liebler DC. Peroxyl radical scavenging by beta-carotene in lipid bilayers. Effect of oxygen partial pressure. J Biol Chem. 1992;267:4658–4663. [PubMed] [Google Scholar]
- 57.Wisniewska A, Widomska J, Subczynski WK. Carotenoid-membrane interactions in liposomes: Effect of dipolar, monopolar, and nonpolar carotenoids. Acta Biochim Pol. 2006;53:475–484. [PubMed] [Google Scholar]
- 58.Socaciu C, Jessel R, Diehl HA. Competitive carotenoid and cholesterol incorporation into liposomes: Effects on membrane phase transition, fluidity, polarity and anisotropy. Chem Phys Lipids. 2000;106:79–88. doi: 10.1016/s0009-3084(00)00135-3. [DOI] [PubMed] [Google Scholar]
- 59.Widomska J, Kostecka-Gugala A, Latowski D, Gruszecki WI, Strzalka K. Calorimetric studies of the effect of cis-carotenoids on the thermotropic phase behavior of phosphatidylcholine bilayers. Biophys Chem. 2009;140:108–114. doi: 10.1016/j.bpc.2008.12.002. [DOI] [PubMed] [Google Scholar]
- 60.Yin JJ, Subczynski WK. Effects of lutein and cholesterol on alkyl chain bending in lipid bilayers: A pulse electron spin resonance spin labeling study. Biophys J. 1996;71:832–839. doi: 10.1016/S0006-3495(96)79284-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wisniewska A, Subczynski WK. Effects of polar carotenoids on the shape of the hydrophobic barrier of phospholipid bilayers. Biochim Biophys Acta. 1998;1368:235–246. doi: 10.1016/s0005-2736(97)00182-x. [DOI] [PubMed] [Google Scholar]
- 62.Grolier P, Zais-Braesco V, Zelmire L, Fessi H. Incorporation of carotenoids in aqueous systems: Uptake by cultured rat hepatocytes. Biochim Biophys Acta. 1992;1111:135–138. doi: 10.1016/0005-2736(92)90283-r. [DOI] [PubMed] [Google Scholar]
- 63.Wisniewska A, Draus J, Subczynski WK. Is a fluid-mosaic model of biological membranes fully relevant? Studies on lipid organization in model and biological membranes. Cell Mol Biol Lett. 2003;8:147–159. [PubMed] [Google Scholar]
- 64.Bone RA, Landrum JT. Macular pigment in Henle fiber membranes: A model for Haidinger’s brushes. Vis Res. 1984;24:103–108. doi: 10.1016/0042-6989(84)90094-4. [DOI] [PubMed] [Google Scholar]
- 65.Gruszecki WI, Sielewiesiuk J. Orientation of xanthophylls in phosphatidylcholine multibilayers. Biochim Biophys Acta. 1990;1023:405–412. doi: 10.1016/0005-2736(90)90133-9. [DOI] [PubMed] [Google Scholar]
- 66.Gruszecki WI, Strzalka K. Carotenoids as modulators of lipid membrane physical properties. Biochim Biophys Acta. 2005;1740:108–115. doi: 10.1016/j.bbadis.2004.11.015. [DOI] [PubMed] [Google Scholar]
- 67.Subczynski WK, Markowska E, Sielewiesiuk J. Effect of polar carotenoids on the oxygen diffusion-concentration product in lipid bilayers. An EPR spin label study. Biochim Biophys Acta. 1991;1068:68–72. doi: 10.1016/0005-2736(91)90061-c. [DOI] [PubMed] [Google Scholar]
- 68.Subczynski WK, Markowska E, Sielewiesiuk J. Spin-label studies on phosphatidylcholine-polar carotenoid membranes: Effects of alkyl-chain length and unsaturation. Biochim Biophys Acta. 1993;1150:173–181. doi: 10.1016/0005-2736(93)90087-g. [DOI] [PubMed] [Google Scholar]
- 69.Gruszecki WI. Carotenoids in membranes. In: Frank HA, Young A, Britton G, Cogdell RJ, editors. The Photochemistry of Carotenoids. Kluwer Academic Publishers; Berlin, Gremany: 1999. pp. 363–379. [Google Scholar]
- 70.Gruszecki WI, Sujak A, Strzalka K, Radunz A, Schmid GH. Organisation of xanthophyll-lipid membranes studied by means of specific pigment antisera, spectrophotometry and monomolecular layer technique lutein versus zeaxanthin. Z Naturforsch C. 1999;54:517–525. doi: 10.1515/znc-1999-7-810. [DOI] [PubMed] [Google Scholar]
- 71.Landrum JT, Bone RA, Joa H, Kilburn MD, Moore LL, Sprague KE. A one year study of the macular pigment: The effect of 140 days of a lutein supplement. Exp Eye Res. 1997;65:57–62. doi: 10.1006/exer.1997.0309. [DOI] [PubMed] [Google Scholar]
- 72.Hammond BR, Jr, Johnson EJ, Russell RM, Krinsky NI, Yeum KJ, Edwards RB, Snodderly DM. Dietary modification of human macular pigment density. Investig Ophthalmol Vis Sci. 1997;38:1795–1801. [PubMed] [Google Scholar]
- 73.Tserentsoodol N, Sztein J, Campos M, Gordiyenko NV, Fariss RN, Lee JW, Fliesler SJ, Rodriguez IR. Uptake of cholesterol by the retina occurs primarily via a low density lipoprotein receptor-mediated process. Mol Vis. 2006;12:1306–1318. [PubMed] [Google Scholar]
- 74.Shafaa MW, Diehl HA, Socaciu C. The solubilisation pattern of lutein, zeaxanthin, canthaxanthin and beta-carotene differ characteristically in liposomes, liver microsomes and retinal epithelial cells. Biophys Chem. 2007;129:111–119. doi: 10.1016/j.bpc.2007.05.007. [DOI] [PubMed] [Google Scholar]
- 75.Sujak A, Gabrielska J, Grudzinski W, Borc R, Mazurek P, Gruszecki WI. Lutein and zeaxanthin as protectors of lipid membranes against oxidative damage: The structural aspects. Arch Biochem Biophys. 1999;371:301–307. doi: 10.1006/abbi.1999.1437. [DOI] [PubMed] [Google Scholar]
- 76.Okulski W, Sujak A, Gruszecki WI. Dipalmitoylphosphatidylcholine membranes modified with carotenoid pigment lutein: Experiment versus Monte Carlo simulation study of the membrane organization. Biochim Biophys Acta. 2008;1778:2105–2118. doi: 10.1016/j.bbamem.2008.04.002. [DOI] [PubMed] [Google Scholar]
- 77.Pasenkiewicz-Gierula M, Baczynski K, Murzyn K, Markiewicz M. Orientation of lutein in a lipid bilayer -revisited. Acta Biochim Pol. 2012;59:115–118. [PubMed] [Google Scholar]
- 78.Woodall AA, Britton G, Jackson MJ. Carotenoids and protection of phospholipids in solution or in liposomes against oxidation by peroxyl radicals: Relationship between carotenoid structure and protective ability. Biochim Biophys Acta. 1997;1336:575–586. doi: 10.1016/s0304-4165(97)00007-x. [DOI] [PubMed] [Google Scholar]
- 79.Woodall AA, Britton G, Jackson MJ. Antioxidant activity of carotenoids in phosphatidylcholine vesicles: Chemical and structural considerations. Biochem Soc Trans. 1995;23:133S. doi: 10.1042/bst023133s. [DOI] [PubMed] [Google Scholar]
- 80.Woodall AA, Lee SW, Weesie RJ, Jackson MJ, Britton G. Oxidation of carotenoids by free radicals: Relationship between structure and reactivity. Biochim Biophys Acta. 1997;1336:33–42. doi: 10.1016/s0304-4165(97)00006-8. [DOI] [PubMed] [Google Scholar]
- 81.Britton G. Structure and properties of carotenoids in relation to function. FASEB J. 1995;9:1551–1558. [PubMed] [Google Scholar]
- 82.Subczynski WK, Widomska J, Feix JB. Physical properties of lipid bilayers from EPR spin labeling and their influence on chemical reactions in a membrane environment. Free Radic Biol Med. 2009;46:707–718. doi: 10.1016/j.freeradbiomed.2008.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Boesze-Battaglia K, Dispoto J, Kahoe MA. Association of a photoreceptor-specific tetraspanin protein, ROM-1, with triton X-100-resistant membrane rafts from rod outer segment disk membranes. J Biol Chem. 2002;277:41843–41849. doi: 10.1074/jbc.M207111200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Martin RE, Elliott MH, Brush RS, Anderson RE. Detailed characterization of the lipid composition of detergent-resistant membranes from photoreceptor rod outer segment membranes. Investig Ophthalmol Vis Sci. 2005;46:1147–1154. doi: 10.1167/iovs.04-1207. [DOI] [PubMed] [Google Scholar]
- 85.Nair KS, Balasubramanian N, Slepak VZ. Signal-dependent translocation of transducin, RGS9–1-Gbeta5L complex, and arrestin to detergent-resistant membrane rafts in photoreceptors. Curr Biol. 2002;12:421–425. doi: 10.1016/s0960-9822(02)00691-7. [DOI] [PubMed] [Google Scholar]
- 86.Seno K, Kishimoto M, Abe M, Higuchi Y, Mieda M, Owada Y, Yoshiyama W, Liu H, Hayashi F. Light- and guanosine 5′-3-O-(thio)triphosphate-sensitive localization of a G protein and its effector on detergent-resistant membrane rafts in rod photoreceptor outer segments. J Biol Chem. 2001;276:20813–20816. doi: 10.1074/jbc.C100032200. [DOI] [PubMed] [Google Scholar]
- 87.Polozova A, Litman BJ. Cholesterol dependent recruitment of di22:6-PC by a G protein-coupled receptor into lateral domains. Biophys J. 2000;79:2632–2643. doi: 10.1016/S0006-3495(00)76502-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Lincoln JE, Boling M, Parikh AN, Yeh Y, Gilchrist DG, Morse LS. Fas signaling induces raft coalescence that is blocked by cholesterol depletion in human RPE cells undergoing apoptosis. Investig Ophthalmol Vis Sci. 2006;47:2172–2178. doi: 10.1167/iovs.05-1167. [DOI] [PubMed] [Google Scholar]
- 89.Anderson RE, Benolken RM, Dudley PA, Landis DJ, Wheeler TG. Proceedings: Polyunsaturated fatty acids of photoreceptor membranes. Exp Eye Res. 1974;18:205–213. doi: 10.1016/0014-4835(74)90149-3. [DOI] [PubMed] [Google Scholar]
- 90.Litman BJ, Mitchell DC. A role for phospholipid polyunsaturation in modulating membrane protein function. Lipids. 1996;31:S193–S197. doi: 10.1007/BF02637075. [DOI] [PubMed] [Google Scholar]
- 91.Mitchell DC, Straume M, Litman BJ. Role of sn-1-saturated,sn-2-polyunsaturated phospholipids in control of membrane receptor conformational equilibrium: Effects of cholesterol and acyl chain unsaturation on the metarhodopsin I in equilibrium with metarhodopsin II equilibrium. Biochemistry. 1992;31:662–670. doi: 10.1021/bi00118a005. [DOI] [PubMed] [Google Scholar]
- 92.Agbaga MP, Brush RS, Mandal MN, Henry K, Elliott MH, Anderson RE. Role of Stargardt-3 macular dystrophy protein (ELOVL4) in the biosynthesis of very long chain fatty acids. Proc Natl Acad Sci USA. 2008;105:12843–12848. doi: 10.1073/pnas.0802607105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Agbaga MP, Mandal MN, Anderson RE. Retinal very long-chain PUFAs: New insights from studies on ELOVL4 protein. J Lipid Res. 2010;51:1624–1642. doi: 10.1194/jlr.R005025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Aveldano MI. Phospholipid species containing long and very long polyenoic fatty acids remain with rhodopsin after hexane extraction of photoreceptor membranes. Biochemistry. 1988;27:1229–1239. doi: 10.1021/bi00404a024. [DOI] [PubMed] [Google Scholar]
- 95.Parekh N, Voland RP, Moeller SM, Blodi BA, Ritenbaugh C, Chappell RJ, Wallace RB, Mares JA. Association between dietary fat intake and age-related macular degeneration in the Carotenoids in Age-Related Eye Disease Study (CAREDS): An ancillary study of the Women’s Health Initiative. Arch Ophthalmol. 2009;127:1483–1493. doi: 10.1001/archophthalmol.2009.130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Sangiovanni JP, Chew EY. The role of omega-3 long-chain polyunsaturated fatty acids in health and disease of the retina. Prog Retin Eye Res. 2005;24:87–138. doi: 10.1016/j.preteyeres.2004.06.002. [DOI] [PubMed] [Google Scholar]
- 97.Liu A, Chang J, Lin Y, Shen Z, Bernstein PS. Long-chain and very long-chain polyunsaturated fatty acids in ocular aging and age-related macular degeneration. J Lipid Res. 2010;51:3217–3229. doi: 10.1194/jlr.M007518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Delmelle M. An investigation of retinal as a source of singlet oxygen. Photochem Photobiol. 1978;27:731–734. [Google Scholar]
- 99.Rozanowska M, Sarna T. Light-induced damage to the retina: Role of rhodopsin chromophore revisited. Photochem Photobiol. 2005;81:1305–1330. doi: 10.1562/2004-11-13-IR-371. [DOI] [PubMed] [Google Scholar]
- 100.Wisniewska A, Subczynski WK. Distribution of macular xanthophylls between domains in a model of photoreceptor outer segment membranes. Free Radic Biol Med. 2006;41:1257–1265. doi: 10.1016/j.freeradbiomed.2006.07.003. [DOI] [PubMed] [Google Scholar]
- 101.Wisniewska A, Subczynski WK. Accumulation of macular xanthophylls in unsaturated membrane domains. Free Radic Biol Med. 2006;40:1820–1826. doi: 10.1016/j.freeradbiomed.2006.01.016. [DOI] [PubMed] [Google Scholar]
- 102.Landrum JT, Bone RA. Mechanistic evidence for eye diseases and carotenoids. In: Krinsky NI, Mayne ST, Sies H, editors. Carotenoids in Health and Disease. Marcel Dekker; New York, NY, USA: 2004. pp. 445–472. [Google Scholar]
- 103.Wisniewska-Becker A, Nawrocki G, Duda M, Subczynski WK. Structural aspects of the antioxidant activity of lutein in a model of photoreceptor membranes. Acta Biochim Pol. 2012;59:119–124. [PMC free article] [PubMed] [Google Scholar]
- 104.Cullen AP. Photokeratitis and other phototoxic effects on the cornea and conjunctiva. Int J Toxicol. 2002;21:455–464. doi: 10.1080/10915810290169882. [DOI] [PubMed] [Google Scholar]
- 105.Dillon J, Atherton SJ. Time resolved spectroscopic studies on the intact human lens. Photochem Photobiol. 1990;51:465–468. doi: 10.1111/j.1751-1097.1990.tb01738.x. [DOI] [PubMed] [Google Scholar]
- 106.Boulton M, Rozanowska M, Rozanowski B. Retinal photodamage. J Photochem Photobiol B. 2001;64:144–161. doi: 10.1016/s1011-1344(01)00227-5. [DOI] [PubMed] [Google Scholar]
- 107.Gorgels TG, van ND. Ultraviolet and green light cause different types of damage in rat retina. Investig Ophthalmol Vis Sci. 1995;36:851–863. [PubMed] [Google Scholar]
- 108.Grimm C, Wenzel A, Williams T, Rol P, Hafezi F, Reme C. Rhodopsin-mediated blue-light damage to the rat retina: Effect of photoreversal of bleaching. Investig Ophthalmol Vis Sci. 2001;42:497–505. [PubMed] [Google Scholar]
- 109.Pautler EL, Morita M, Beezley D. Hemoprotein(s) mediate blue light damage in the retinal pigment epithelium. Photochem Photobiol. 1990;51:599–605. doi: 10.1111/j.1751-1097.1990.tb01972.x. [DOI] [PubMed] [Google Scholar]
- 110.Arnault E, Barrau C, Nanteau C, Gondouin P, Bigot K, Vienot F, Gutman E, Fontaine V, Villette T, Cohen-Tannoudji D, et al. Phototoxic action spectrum on a retinal pigment epithelium model of age-related macular degeneration exposed to sunlight normalized conditions. PLoS ONE. 2013;8:e71398. doi: 10.1371/journal.pone.0071398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Junghans A, Sies H, Stahl W. Macular pigments lutein and zeaxanthin as blue light filters studied in liposomes. Arch Biochem Biophys. 2001;391:160–164. doi: 10.1006/abbi.2001.2411. [DOI] [PubMed] [Google Scholar]
- 112.Hammond BR, Jr, Elliot JG. Multiple influences of hantophylls within the visual system. In: Landrum JT, Nolan J, editors. Carotenoids and Retinal Disease. CRC Press, Taylor and Francis Group; Boca Raton, FL, USA: 2013. pp. 147–170. [Google Scholar]
- 113.Wooten BR, Hammond BR. Macular pigment: Influences on visual acuity and visibility. Prog Retin Eye Res. 2002;21:225–240. doi: 10.1016/s1350-9462(02)00003-4. [DOI] [PubMed] [Google Scholar]
- 114.Feeney J, Finucane C, Savva GM, Cronin H, Beatty S, Nolan JM, Kenny RA. Low macular pigment optical density is associated with lower cognitive performance in a large, population-based sample of older adults. Neurobiol Aging. 2013;34:2449–2456. doi: 10.1016/j.neurobiolaging.2013.05.007. [DOI] [PubMed] [Google Scholar]
- 115.Johnson EJ, McDonald K, Caldarella SM, Chung HY, Troen AM, Snodderly DM. Cognitive findings of an exploratory trial of docosahexaenoic acid and lutein supplementation in older women. Nutr Neurosci. 2008;11:75–83. doi: 10.1179/147683008X301450. [DOI] [PubMed] [Google Scholar]
- 116.Vishwanathan R, Iannaccone A, Scott TM, Kritchevsky SB, Jennings BJ, Carboni G, Forma G, Satterfield S, Harris T, Johnson KC, et al. Macular pigment optical density is related to cognitive function in older people. Age Ageing. 2014;43:271–275. doi: 10.1093/ageing/aft210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kim SR, Nakanishi K, Itagaki Y, Sparrow JR. Photooxidation of A2-PE, a photoreceptor outer segment fluorophore, and protection by lutein and zeaxanthin. Exp Eye Res. 2006;82:828–839. doi: 10.1016/j.exer.2005.10.004. [DOI] [PubMed] [Google Scholar]
- 118.DiMascio P, Kaiser S, Sies H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch Biochem Biophys. 1989;274:532–538. doi: 10.1016/0003-9861(89)90467-0. [DOI] [PubMed] [Google Scholar]
- 119.DiMascio P, Murphy ME, Sies H. Antioxidant defense systems: The role of carotenoids tocopherols and thiols. Am J Clin Nutr. 1991;53:194S–200S. [PubMed] [Google Scholar]
- 120.Conn PF, Schalch W, Truscott TG. The singlet oxygen and carotenoid interaction. J Photochem Photobiol B. 1991;11:41–47. doi: 10.1016/1011-1344(91)80266-k. [DOI] [PubMed] [Google Scholar]
- 121.Stahl W, Sies H. Antioxidant effects of carotenoids: Implication in photoprotection in humans. In: Packer L, editor. Implication in Photoprotection in Humans: Handbook of Antioxidants. Marcel Dekker; Basel, Switzerland: 2002. pp. 223–234. [Google Scholar]
- 122.Bhosale P, Bernstein PS. Quantitative measurement of 3′-oxolutein from human retina by normal-phase high-performance liquid chromatography coupled to atmospheric pressure chemical ionization mass spectrometry. Anal Biochem. 2005;345:296–301. doi: 10.1016/j.ab.2005.07.006. [DOI] [PubMed] [Google Scholar]
- 123.Khachik F, Bernstein PS, Garland DL. Identification of lutein and zeaxanthin oxidation products in human and monkey retinas. Investig Ophthalmol Vis Sci. 1997;38:1802–1811. [PubMed] [Google Scholar]
- 124.Hurst JS, Contreras JE, Siems WG, van Kuijk FJ. Oxidation of carotenoids by heat and tobacco smoke. BioFactors. 2004;20:23–35. doi: 10.1002/biof.5520200103. [DOI] [PubMed] [Google Scholar]
- 125.Siems WG, Sommerburg O, van Kuijk FJ. Lycopene and beta-carotene decompose more rapidly than lutein and zeaxanthin upon exposure to various pro-oxidants in vitro. BioFactors. 1999;10:105–113. doi: 10.1002/biof.5520100204. [DOI] [PubMed] [Google Scholar]
- 126.Socaciu C, Jessel R, Diehl HA. Carotenoid incorporation into microsomes: Yields, stability and membrane dynamics. Spectrochim Acta A Mol Biomol Spectrosc. 2000;56:2799–2809. doi: 10.1016/s1386-1425(00)00375-9. [DOI] [PubMed] [Google Scholar]
- 127.Burton GW, Ingold KU. beta-Carotene: An unusual type of lipid antioxidant. Science. 1984;224:569–573. doi: 10.1126/science.6710156. [DOI] [PubMed] [Google Scholar]
- 128.Bradbury J. Docosahexaenoic acid (DHA): An ancient nutrient for the modern human brain. Nutrients. 2011;3:529–554. doi: 10.3390/nu3050529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Sastry PS. Lipids of nervous tissue: Composition and metabolism. Prog Lipid Res. 1985;24:69–176. doi: 10.1016/0163-7827(85)90011-6. [DOI] [PubMed] [Google Scholar]
- 130.Breckenridge WC, Gombos G, Morgan IG. The lipid composition of adult rat brain synaptosomal plasma membranes. Biochim Biophys Acta. 1972;266:695–707. doi: 10.1016/0006-3002(72)90012-1. [DOI] [PubMed] [Google Scholar]
- 131.Fliesler SJ, Anderson RE. Chemistry and metabolism of lipids in the vertebrate retina. Prog Lipid Res. 1983;22:79–131. doi: 10.1016/0163-7827(83)90004-8. [DOI] [PubMed] [Google Scholar]
- 132.Wiegand RD, Anderson RE. Phospholipid molecular species of frog rod outer segment membranes. Exp Eye Res. 1983;37:159–173. doi: 10.1016/0014-4835(83)90075-1. [DOI] [PubMed] [Google Scholar]
- 133.Zhang P, Omaye ST. Antioxidant and prooxidant roles for beta-carotene, alpha-tocopherol and ascorbic acid in human lung cells. Toxicol In Vitro. 2001;15:13–24. doi: 10.1016/s0887-2333(00)00054-0. [DOI] [PubMed] [Google Scholar]
- 134.Palozza P. Evidence for pro-oxidant effects of carotenoids in vitro and in vivo: Implications in health and disease. In: Krinsky NI, Mayne ST, Sies H, editors. Carotenoids in Health and Disease. Marcel Dekker; New York, NY, USA: 2004. pp. 127–149. [Google Scholar]
- 135.Bhosale P, Bernstein PS. Vertebrate and invertebrate carotenoid-binding proteins. Arch Biochem Biophys. 2007;458:121–127. doi: 10.1016/j.abb.2006.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Bhosale P, Li B, Sharifzadeh M, Gellermann W, Frederick JM, Tsuchida K, Bernstein PS. Purification and partial characterization of a lutein-binding protein from human retina. Biochemistry. 2009;48:4798–4807. doi: 10.1021/bi9004478. [DOI] [PubMed] [Google Scholar]
- 137.Havaux M, Niyogi KK. The violaxanthin cycle protects plants from photooxidative damage by more than one mechanism. Proc Natl Acad Sci USA. 1999;96:8762–8767. doi: 10.1073/pnas.96.15.8762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Khachik F. Partial synthesis of serum carotenoids and their metabolites. Acta Biochim Pol. 2012;59:75–78. [PubMed] [Google Scholar]