

Summary
Microtubules are non-covalent mesoscale polymers central to the eukaryotic cytoskeleton. Microtubule structure, dynamics and mechanics are modulated by a cell’s choice of tubulin isoforms and posttranslational modifications, a “tubulin code” which is thought to support the diverse morphology and dynamics of microtubule arrays across various cell types, cell cycle and developmental stages. We give a brief historical overview of research into tubulin diversity and highlight recent progress towards uncovering the mechanistic underpinnings of the tubulin code. As a large number of essential pathways converge upon the microtubule cytoskeleton, understanding how cells utilize tubulin diversity is crucial to understanding cellular physiology and disease.
Introduction
Microtubules are one of the four non-covalent polymers that constitute the cytoskeleton, an integrated filamentous protein network that gives cells their shape, facilitates their movement, organizes their cytoplasm and builds the spindle needed to segregate genetic material. Microtubules are highly dynamic and oscillate stochastically between periods of growth and depolymerization, a property known as “dynamic instability” (Figure 1; (Horio, 1986; Mitchison, 1984; Walker, 1988)). The microtubule building block is the αβ-tubulin heterodimer. It assembles head-to-tail to form a polar protofilament with α-tubulin exposed on one end and β-tubulin at the other (reviewed in (Nogales, 2001)). Protofilaments assemble laterally to form a hollow cylinder (Figure 1). Most eukaryotic cells make microtubules with 13 protofilaments (Tilney et al., 1973). At the resolution of the optical microscope, all microtubules look the same, but this is deceptive. Microtubules in cells are highly complex, composed of multiple tubulin isoforms and functionalized with diverse evolutionarily conserved posttranslational modifications. This tubulin diversity alters intrinsic microtubule properties such as their dynamics and mechanics, as well as the recruitment and activity of motors and microtubule-associated proteins (MAPs). This variation has been interpreted as a “tubulin code” by which cells regulate microtubule diversity to achieve specific, physiological goals (Janke and Bulinski, 2011; Verhey and Gaertig, 2007; Yu et al., 2015). How cells write and read the tubulin code is largely unknown, but the advent of new tools for in vitro and in vivo tubulin manipulation, and the identification of an extensive list of tubulin-modifying enzymes is now bringing this fundamental problem into focus.
Figure 1. Microtubule structure and dynamics.

Microtubules are non-covalent cylindrical polymers assembled from 13 protofilaments formed by the head-to-tail addition of αβ-heterodimers. They grow by the addition of GTP-tubulin at their ends. Once incorporated into the microtubule, the GTP is hydrolyzed to GDP. Microtubules cycle between periods of growth and depolymerization. The transition from growth to depolymerization is called catastrophe. The transition from depolymerization to growth is called rescue. Microtubule dynamics are characterized by four parameters: rate of polymerization and depolymerization and frequency of catastrophe and rescue events.
The genetic and chemical complexity of tubulin
Humans have nine isoforms of both α- and β-tubulin. Certain isoforms, like α1A (TUBA1A) and βI (TUBB) are ubiquitous. Others have cell- or tissue-specific expression and are important for the function of specialized microtubule arrays in sperm, platelets and neurons (reviewed in (Ludueña, 2013) and (Breuss et al., 2017)). Early evidence for isoform-specific tubulin function, from studies in Drosophila showed that βIV-tubulin, the dominant β-isoform in flagella and cilia, is important for specifying correct axonemal structure. A sequence motif conserved in this isoform is important for this activity (Kemphues et al., 1979; Nielsen et al., 2001). The first mechanosensation screen revealed that the mec-12 and mec-7 α- and β-tubulin genes are required for the formation of 15-protofilament microtubules found in the mechanosensory neurons of Caenorhabditis elegans (Fukushige et al., 1999; Savage et al., 1989), providing further support for the specialized functions of tubulin isoforms. We now know that mutations in various tubulin isoforms are associated with a broad spectrum of human pathologies (Hussey and Fritz‐Laylin, 2019) ranging from bleeding disorders (Kunishima et al., 2014) and susceptibility to pathogens, (Salinas et al., 2014) to neurodevelopmental and neurodegenerative disorders (reviewed in (Breuss et al., 2017)). The molecular mechanisms behind these physiological responses remain largely unknown and will provide fertile ground for investigations for a new generation of cell biologists.
Tubulin is further diversified through many chemically distinct posttranslational modifications: acetylation of α- and β-tubulin, methylation of α-tubulin, phosphorylation of α- and β-tubulin, palmytoylation of α-tubulin, polyamination of α- and β-tubulin, detyrosination of α-tubulin, removal of the two C-terminal glutamates in α-tubulin to generate Δ2 tubulin, glutamylation and glycylation of α or β-tubulin (Figure 2; reviewed in (Janke and Bulinski, 2011; Yu et al., 2015)).
Figure 2. Sequence variability among tubulin isoforms concentrates on the intrinsically disordered C-terminal tails and at polymerization interfaces.
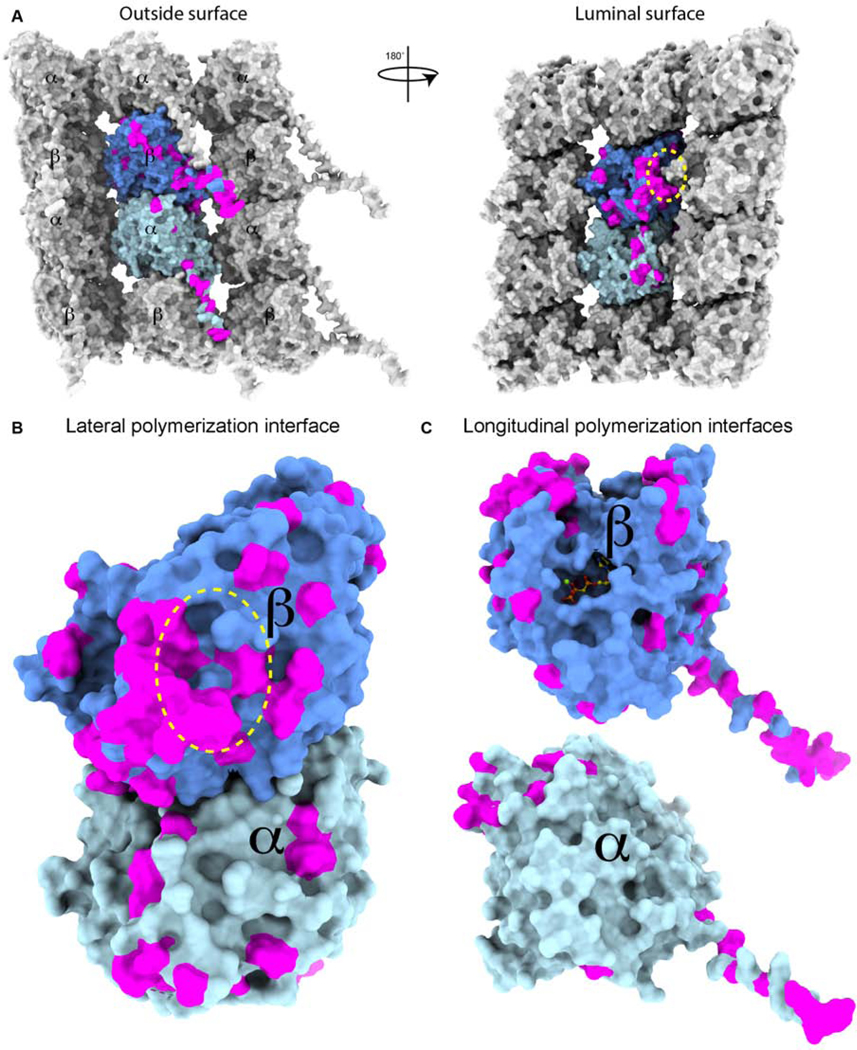
(A) Molecular surface of three protofilaments from a microtubule showing lattice contacts of the central αβ-tubulin dimer within a microtubule. α- and β-tubulin colored as in Figure 1. Sequences from Homo sapiens α- and β-tubulin isoforms were aligned and conservative sequence substitutions mapped in magenta on the atomic model (PDB ID: 6dpu). The M-loop position is highlighted by a yellow ellipse.
(B) Molecular surface of the αβ-tubulin dimer colored as in (A) illustrating the sequence variability on the C-terminal tails and lateral (between tubulin heterodimers in neighboring protofilaments) and longitudinal (between tubulin heterodimers within the same protofilament) polymerization interfaces.
Tubulin posttranslational modifications vary widely between cell types and are developmentally and spatially regulated (reviewed in (Yu et al., 2015)). Many studies have now documented the selective transport of various cargo on subsets of microtubules with specific modifications. For example, lysosomes switch between tyrosinated and acetylated microtubules as they move from the periphery to the cell interior (Guardia et al., 2016). Vesicles in epithelial cells exit the Trans-Golgi network along polyglutamylated microtubules functionalized with septin-2 fibers and aligned with the apicobasal cell axis (Spiliotis et al., 2008). In flagella, cargo movement is efficiently segregated for anterograde and retrograde transport by moving on the adjoining tracks of the microtubule doublet which are differentially modified (Stepanek and Pigino, 2016). Consistent with their involvement in basic cell biological processes, perturbations in tubulin modifications and the enzymes that introduce them are associated with cancers, neurodevelopmental and neurodegenerative disorders (reviewed in (Garnham and Roll‐Mecak, 2012; Magiera et al., 2018b)). The molecular mechanisms that give rise to the formation and readout of microtubule modification patterns in cells are largely unknown.
Historical Overview
Despite knowledge of the chemical complexity of tubulin and its importance for cell physiology dating as far back as the 1970s, our understanding of the cellular and developmental mechanisms behind this diversity and its effects on microtubule functions is still in its infancy. Why do we know so little? Arguably, this is due to (1) the historical development of the microtubule field which emphasized other aspects of tubulin biology after the initial realization of the diversity of microtubule structures and tubulin heterogeneity; (2) the challenges in studying the tubulin code because of the lack of an in vitro reconstitution system, antibodies for tubulin isoforms and tools for the specific and quantitative detection of tubulin modifications in vivo, and the difficulty with fluorescently tagging tubulin in vivo without adverse effects.
The isolation of tubulin (Borisy and Taylor, 1967; Shelanski and Taylor, 1967) ushered in a new biochemical era in microtubule research. Biochemical studies soon revealed that organisms have multiple tubulin forms (Feit et al., 1971; Witman et al., 1972) and that some are posttranslationally modified (Arce et al., 1975). This tubulin heterogeneity gave rise to the “multi-tubulin hypothesis” (Fulton and Simpson, 1976) which postulated that different tubulin isotypes were segregated in cells and required for the biogenesis and function of specialized microtubule structures. The large tubulin diversity became evident with the identification of multigene tubulin families in the 1980s (Cleveland et al., 1980; Lopata et al., 1983; Sullivan and Cleveland, 1986). A chemically diverse array of posttranslational modifications was identified in subsequent decades. Certain tubulin isoforms were found to be ubiquitous while other exhibited cell-type specific expression (reviewed in (Ludueña, 2013)). Likewise, tubulin posttranslational modifications were stereotyped and distributed asymmetrically in various cells (reviewed in (Janke and Bulinski, 2011; Yu et al., 2015)). These results generated a lot of excitement. Landmark studies documented the increase in tubulin modifications during cellular differentiation (Gundersen et al., 1989) and the different susceptibility to drug or temperature-induced depolymerization of cellular microtubules with different posttranslational modifications (Kreis, 1987; Schulze et al., 1987; Schulze and Kirschner, 1987; Webster et al., 1987). However, because many different tubulin isotypes could co-assemble both in vitro and in cells and because the few loss-of-function studies attempted failed to give easily interpretable phenotypes, there was no unifying theory of the roles of tubulin isoforms. As a result, the multi-tubulin hypothesis lost its sheen and most of the field directed its attention elsewhere.
The decades after the identification of tubulin isoforms saw an explosion in the discovery of factors that interact with tubulin and microtubules to build morphologically and dynamically diverse microtubule structures in cells: proteins that stabilize microtubules such as tau (Cleveland et al., 1977), motors that transport cargo (Brady, 1985; Vale et al., 1985a) or end-depolymerize microtubules (Walczak et al., 1996), proteins that localize to microtubule plus ends and modulate their dynamics (Rickard and Kreis, 1990) and enzymes that sever microtubules (Vale, 1991). Crucial to understanding the mechanism of these newly discovered molecular players was the introduction of in vitro reconstitution assays for motility (Vale et al., 1985b) and microtubule dynamics (Horio, 1986; Kinoshita et al., 2001; Walker, 1988). However, throughout all of this exciting time of building the microtubule cytoskeleton from the ground-up, tubulin itself was left out, its chemical and genetic diversity largely ignored and swept under the rug. This was in large part because many tubulin modifying enzymes remained unknown, making the manipulation of modifications difficult, and because there was no biochemical platform to investigate the consequences of tubulin diversity.
Tubulin diversity has returned into focus largely for three reasons: (1) identification of many of the enzymes that posttranslationally modify tubulin (Aillaud et al., 2017; Akella et al., 2010; Janke et al., 2005; Nieuwenhuis et al., 2017; Rogowski et al., 2009; Rogowski et al., 2010; van Dijk et al., 2007), (2) the development of biochemical tools for the production of naïve and recombinant tubulin (Minoura et al., 2013; Ti et al., 2018; Vemu et al., 2016; Widlund et al., 2012), and (3) the generation of tubulin with quantitatively defined posttranslational modifications (Valenstein and Roll-Mecak, 2016; Vemu et al., 2014) for in vitro reconstitution. The “multi-tubulin hypothesis” has resurfaced as the more expansive “tubulin code” hypothesis which postulates that tubulin isoforms and their chemically diverse posttranslational modifications regulate microtubule structure and dynamics and function as cellular GPS signals recognized and interpreted by cellular effectors for localized microtubule-based functional outputs (Verhey and Gaertig, 2007). Here we concentrate on recent advances that have returned the genetic and chemical complexity of tubulin to the fore of the microtubule field.
The tubulin problem
Why do we still understand so little about the effects of tubulin diversity, either genetically encoded or arising from posttranslational modifications? One reason is that tubulin did not succumb to standard molecular biology approaches until recently. While most other fields take recombinant tools for granted, tubulin has resisted attempts at recombinant production. One of the main difficulties is that the generation of microtubule-assembly-competent αβ-tubulin dimers requires a complex collection of chaperones and downstream factors (Lewis et al., 1997). Moreover, eukaryotic cells regulate overall tubulin levels tightly (Burke et al., 1989) and tubulin isoforms copolymerize. Because of a lack of a recombinant source, the field resorted to purifying bulk tubulin from mammalian brain, as tubulin is highly abundant in this tissue. This is problematic, however, because tubulin from this source contains multiple isoforms and is abundantly and diversely modified. In fact, most of the tubulin in the adult nervous system is heavily posttranslationally modified, with glutamylation being most abundant (Wolff et al., 1992; Wolff et al., 1982). Tens of different α– and β-tubulin species are present in these preparations (Vemu et al., 2017; Wolff et al., 1982). Tubulin purification from brain entails repeated cycles of temperature and GTP-driven polymerization and depolymerization (Weisenberg, 1972). This procedure has a critical unintended consequence for deciphering the roles of tubulin diversity: the topographical information originally encoded in the patterns of tubulin isoforms and modifications in cells is lost by tissue homogenization; modifications present in the original cellular microtubules become incorporated in random mosaic patterns when the tubulin is repolymerized in vitro. Thus, microtubules assembled from this tubulin are comprised of a randomized mixture of isoforms bearing multiple, chemically distinct and quantitatively varied posttranslational modifications. As a result, the substrates used to study motors and MAPs have jumbled codes; all the spatial information encoded in them by the cell has been irretrievably scrambled. This tubulin purification procedure also discards tubulin subpopulations that do not depolymerize robustly when subjected to cold, as microtubules with certain posttranslational modifications do (Brady et al., 1984). Lastly, this source also limits investigations to a fixed repertoire of tubulin isoforms found in neuronal tissue. To decipher the tubulin code, novel biochemical and biophysical methods were required for in vitro reconstitution to probe the functions of tubulin diversity.
Initial attempts to express tubulin in bacteria and then refold it using chaperones from reticulate lysate gave very small yields (Shah et al., 2001). Yeast tubulin could be purified, albeit at low yields (Barnes et al., 1992; Kilmartin, 1981), and offered the hope of purifying engineered variants (Gupta Jr et al., 2002). Since then, S. cerevisiae has been used to overexpress and purify tubulin mutants suitable for structural studies (Ayaz et al., 2012; Johnson et al., 2011) or mechanistic work into polymer dynamics (Geyer et al., 2015). More recently, it became possible to express and purify recombinant, engineered, and isotypically pure human tubulin (Minoura et al., 2013; Ti et al., 2018; Vemu et al., 2016).
The second major reason for the limited progress in understanding the code is that in vivo tubulin is chemically complex, and modifications are posttranslational. Thus, making a certain type of tubulin requires knowing, understanding and harnessing the properties of the enzymes involved in modifying it. The ability to make recombinant or unmodified tubulin coupled with the knowledge garnered from recent biochemical and structural characterization of tubulin modifying enzymes (Garnham et al., 2015; Garnham et al., 2017; Li et al., 2019; Liao et al., 2019; Mukai et al., 2009; Prota et al., 2013; Shida et al., 2010; Szyk et al., 2011; Szyk et al., 2014; Wang et al., 2019) allows now the generation of tubulin with quantitatively defined posttranslational modifications to be used in deciphering the direct effects of the tubulin code on microtubule behavior and the factors that interact with microtubules.
Tubulin isoforms specialize and diversify microtubules
Multicellular organisms contain multiple tubulin genes. Each tubulin protomer consists of a compactly folded “body” and an intrinsically disordered, negatively charged C-terminal tail (Nogales et al., 1998). The tubulin body participates in α-tubulin-β-tubulin intradimer and interdimer (microtubule lattice) interactions. The C-terminal tails (two per dimer) decorate the microtubule exterior (Figure 1), where they participate in interactions with molecular motors and MAPs. Since the tails are evolutionarily unconstrained by the need to preserve dimer folding and functional polymerization (microtubule lattice) interfaces, they are hotspots for tubulin sequence variation. The remaining sequence variation concentrates at lateral and longitudinal polymerization interfaces where it can modulate microtubule structure and dynamics (Figure 3). A hotspot for sequence variation is around the M-loop, which participates in inter-protofilament interactions (Figures 3A and B). The M-loop conformation is modulated by microtubule-targeting agents such as taxol (Kellogg et al., 2017), one of the most widely used anti-cancer chemotherapy drugs. The response of the different tubulin isoforms to microtubule targeting agents is of great interest because drug-resistant tumors change their tubulin isoform expression profiles (reviewed in (Gan et al., 2010)).
Figure 3. αβ-tubulin dimers are functionalized with a diverse arrays of chemically distinct posttranslational modifications.

Schematic of a neuron (microtubules, blue; actin, pink) with a close-up of a microtubule decorated with diverse posttranslational modifications. Microtubule colored as in Figure 1. α-tubulin tyrosination, orange; monoglutamaylation, light red; polyglutamylation red; monoglycylation, light green; polyglycylation, green; acetylation or methylation on α-tubulin Lys40, magenta; phosphorylation on Ser172, yellow; polyamination, purple. Microtubule cross-section shows the luminal position of Lys40. Boxes show the chemical structure of selected posttranslational modifications. We note that the linkage of the polyglutamate chain is still not clear with studies reporting both α-linked glutamate chains (Redeker et al., 1991), shown here (where the elongating glutamate forms a peptide bond with the branch glutamate residue) and γ-linked chains (where the elongating glutamate forms an isopeptide bond with the branch glutamate residue (Wolff et al., 1994)).
Both α– and β-tubulin protomers bind to GTP. The GTP site on α-tubulin is at the αβ intradimer interface and not accessible for exchange. The GTP bound to β-tubulin, is hydrolyzed to GDP when tubulin incorporates into the microtubule lattice, and exchanges back to GTP on the depolymerized tubulin dimer (reviewed in (Desai and Mitchison, 1997)). Amino acids in the structural core and GTP-binding sites of the tubulin dimer are largely invariant (Figure 3D), suggesting either a unique solution to the requirement for tubulin to act as its own GTPase activating protein during tubulin polymerization, or that the need for the different nucleotide states of tubulin to interact with multiple partners has limited evolutionary drift (i.e., “the principle of continuity” (Orgel, 1968)). Interestingly, there is at present no evidence for a tubulin GTP exchange factor.
Remarkably, we do not know the exact stoichiometry of tubulin isoforms in most cell types, or even how α- and β-tubulin isoforms pair with each other. Are all combinations equally favorable? Or are there preferred pairings of α and β isoforms? Some evidence exists for tubulin sorting in vivo (Hoyle et al., 2001), as well as for α- and β-tubulin isoform coevolution (Nielsen et al., 2010). More recent studies identified differences in dimerization affinity between α–β-tubulin protomer pairs from different sources (Montecinos-Franjola et al., 2018), but so far, our understanding of the rules for tubulin heterodimerization is rudimentary. We also have limited understanding of the transcriptional, translational and degradation programs that regulate tubulin isoform levels throughout the lifetime of an organism. Early work showed that an increase in soluble tubulin triggers tubulin mRNAs degradation, providing a quick feedback mechanism to achieve tubulin homeostasis, a process termed “tubulin autoregulation” (Cleveland et al., 1981; Pachter et al., 1987). Tubulin targeting agents as well as nutrient deprivation regulate tubulin mRNA levels (Gasic et al., 2019). Recent work finally shed light on the molecular mechanism for this regulation by identifying tetratricopeptide protein 5 (TTC5) as the factor that recognizes the amino terminus of the nascent tubulin peptide on the translating ribosome, targeting it for degradation (Lin et al., 2020).
Early studies supported the idea that microtubule dynamics can be changed in cells by varying the relative levels of tubulin isoforms (Lu and Luduena, 1994; Panda et al., 1994). Recent studies using recombinant tubulin have shown that, indeed, tubulin isoforms have different dynamic parameters (Pamula et al., 2016; Vemu et al., 2017; Vemu et al., 2016) and that titrating tubulin isoform composition can proportionally tune microtubule dynamics and elicit changes of magnitude comparable to those induced by MAPs. For example, microtubules assembled from non-neuronal specific isoforms βI, II, or IVb polymerize faster and are less dynamic than those containing neuronal βIII tubulin (Pamula et al., 2016; Vemu et al., 2017), indicating that changes in tubulin isoform expression during neurogenesis directly impact microtubule turnover in addition to possibly recruiting isoform-specific regulators. Interestingly, βIII tubulin is only a minor isoform in the brain of Antarctic fish which live in temperatures as low as −1.8 C° (Detrich et al., 2000), compared to mammals where it comprises 25% of total tubulin (Banerjee et al., 1992). Tubulin polymerization is finely tuned to organism temperature. Mammalian tubulin depolymerizes rapidly at lower temperatures. Thus, temperature homeostasis of microtubule function could be achieved through differential expression of tubulin isoforms triggered by changes in environmental temperatures. This promises to be an interesting area of future exploration.
Structures of microtubules composed of isotypically pure α1AβIII tubulin revealed structural changes at lateral polymerization interfaces which are a hotspots of sequence variability between tubulin isoforms, and thus can modulate microtubule dynamics parameters (Vemu et al., 2016). More recently, structural studies of C. elegans microtubules also revealed the structural plasticity of lateral polymerization interfaces and proposed that the structuring of these lateral contacts is rate-limiting in microtubule growth (Chaaban et al., 2018). Interestingly, early structure-based sequence analysis of Antarctic fish tubulin isoforms revealed key mutations at lateral interfaces when compared to tubulins from species that live at warmer temperatures (Detrich et al., 2000). Thus, tuning of lateral interfaces could be a general evolutionary mechanism for biochemical adaptation of the tubulin dimer for physiologically optimal dynamic parameters.
The ability to make recombinant tubulin or isolate it from varied sources opens the way to the complete structural and functional characterization of tubulin isoforms. The coming years will likely bring a comprehensive understanding of the basic biophysical properties of tubulin isoforms from a variety of organisms: their polymerization properties, their effect on microtubule structure, dynamics and mechanics and how these relate to cell physiology. The spatial organization of tubulin isoforms is largely unexplored: do tubulin isoforms incorporate into patterns in microtubule arrays and are such patterns used by cells to recruit specific cellular effectors? Such patterns could be generated through different rates of incorporation and relative expression levels of tubulin isoforms or could result from sophisticated regulatory mechanisms such as local translation and degradation. The microtubule plus-end-tracking protein adenomatous polyposis coli (APC) associates with the 3’ UTR of βIIb (TUBB2B) to control its local translation at the growth cone periphery where it incorporates into microtubules (Preitner et al., 2014). Unlike βI and βIII, βII-tubulin localizes to neurites and not cell bodies (Guo et al., 2011). Thus, local translation can support local remodeling of microtubule isoform composition. Enrichment of certain tubulin isoforms at other subcellular locations such as cilia, cell cortex and centrosome has been also documented (reviewed in (Ludueña, 2013)). The mechanism for this selective incorporation of tubulin isoforms at subcellular locations is an interesting area of exploration.
A characterization of tubulin isoform patterns in cells and tissues has been hampered by a limited repertoire of isoform specific antibodies and difficulties with systematic tagging of tubulin without altering the tubulin isoform balance in the organism (e.g., the adverse effects of GFP tagging on microtubule function (Jenkins et al., 2017)). Recent progress in developing better tags for live-cell imaging and electron microscopy will greatly aid these efforts. The renewed interest in the tubulin code, coupled with advances in recombinant tubulin production, should also catalyze the isolation of high-affinity affimers and antibodies specific for different tubulin isoforms and suitable for high-resolution imaging to obtain comprehensive maps of tubulin isoforms in microtubules from different cell types and organisms.
Tubulin modifications: effects on microtubule structure, mechanics and dynamics
The posttranslational modifications most strongly associated with tubulin involve the covalent addition of amino acids with distinctly different physio-chemical properties (glutamate, glycine and tyrosine). Glutamates and glycines are added to internal glutamates in the intrinsically disordered C-terminal tails of α- or β-tubulin (Figure 2). Tyrosine is added at the carboxy-end of α-tubulin (Figure 2), after being first removed posttranslationally as part of the detyrosination/tyrosination cycle. Most of the tubulin purified from brain has three to six added glutamates, and as many as eleven and seven (on α- and β-tubulin, respectively) (Redeker, 2010) and is also detyrosinated. Tubulin in axonemes has as many as 21 added glutamates (Geimer et al., 1997; Schneider et al., 1998). Larger numbers of posttranslationally added glutamates are likely found but are not detected by mass spectrometry because of the challenge in ionizing highly-charged species. Axonemal tubulin is also abundantly glycylated with one to 40 and two to 26 glycines detected on α– and β-tails, respectively (Wall et al., 2016). Thus, the posttranslationally added amino acid chains can be longer than the genetically-encoded tubulin tails, which are ten to 15 residues long and thus can significantly affect the access and recruitment of cellular factors to the microtubule surface. In vivo, glutamylated as well as detyrosinated microtubules turn over slower and are more resistant to cold or drug induced depolymerization (Schulze and Kirschner, 1987; Webster et al., 1987). Glycylation is primarily found in axonemes where it is important for their long-term stability (Bosch Grau et al., 2013). It is not yet clear whether the different behaviors of these microtubule subpopulations in cells reflect different intrinsic polymer properties or are a result of specific interactions with cellular effectors. This question will only be answered by measuring the effects of detyrosination, glutamylation or glycylation on microtubules in vitro with well-defined tubulin preparations.
In addition to the flexible tails, residues in the tubulin body are also modified through acetylation, methylation, phosphorylation, palmytoylation, sumoylation, phosphorylation and polyamination (Figure 2, reviewed in (Yu et al., 2015)). Several of these modifications affect microtubule properties. Acetylation at Lys40 in the microtubule lumen weakens interactions between protofilaments leading to an increase in resistance against buckling (Xu et al., 2017). In the absence of acetylation, microtubules in C. elegans mechanosensory neurons splay apart and have frequent protofilament number changes (Cueva et al., 2012; Topalidou et al., 2012). Acetylated α-tubulin is also essential for touch sensation in mice and Drosophila (Morley et al., 2016; Yan et al., 2018). Acetylation of β-tubulin Lys 252 impairs tubulin polymerization, likely by affecting the α-β intradimer interface and promoting a conformation that disfavors incorporation into microtubules (Chu et al., 2011). Phosphorylation of β-tubulin Ser172 (Figure 2) inhibits tubulin dimer incorporation into microtubules (Fourest-Lieuvin et al., 2006; Ori-McKenney et al., 2016). Several other phosphorylation sites were identified in α and β-tubulin (Liu et al., 2015), however their effects are unknown. Polyamination (Figure 2), primarily on residues at polymerization interfaces renders microtubules resistant to cold-induced depolymerization (Song et al., 2013).
Tubulin code readers: effects of tubulin diversity on microtubule regulators
The microtubule surface, as seen by cellular effectors, is conformationally and chemically complex and not the compact, uniform surface that we have been accustomed to seeing in textbooks (Figure 2). In addition to the changes in electrostatic character introduced by modifications, the flexibility and increased search space of the tubulin tails modified with posttranslationally added amino acids also increase the odds of productive interactions with rigid-body binding partners such as the ATPase domains of molecular motors. The velocity and processivity of kinesin-1, 2 and 3 are all upregulated by polyglutamylation (Sirajuddin, 2014)(Figure 4A, B). Kinesin-1 processivity on βIII microtubules is three-fold lower than on microtubules assembled from βII tubulin because of a positively charged residue in the βIII-tail. A negative charge increase through glutamylation increases run length, restoring it to that on βII microtubules (Sirajuddin, 2014). This illustrates the potential combinatorial effects of tubulin isoform diversity and posttranslational modifications. A striking example of microtubule track specialization is found in flagella where kinesin-II powered intraflagellar trains move on the B-tubule enriched in detyrosination, glutamylation and glycylation while dynein-II powered IFT move on the mainly tyrosinated, unmodified A-tubule modified (Figure 4C; (Johnson, 1998; Lechtreck and Geimer, 2000; Multigner et al., 1996; Stepanek and Pigino, 2016).
Figure 4. The tubulin code regulates interactions with microtubule effectors.
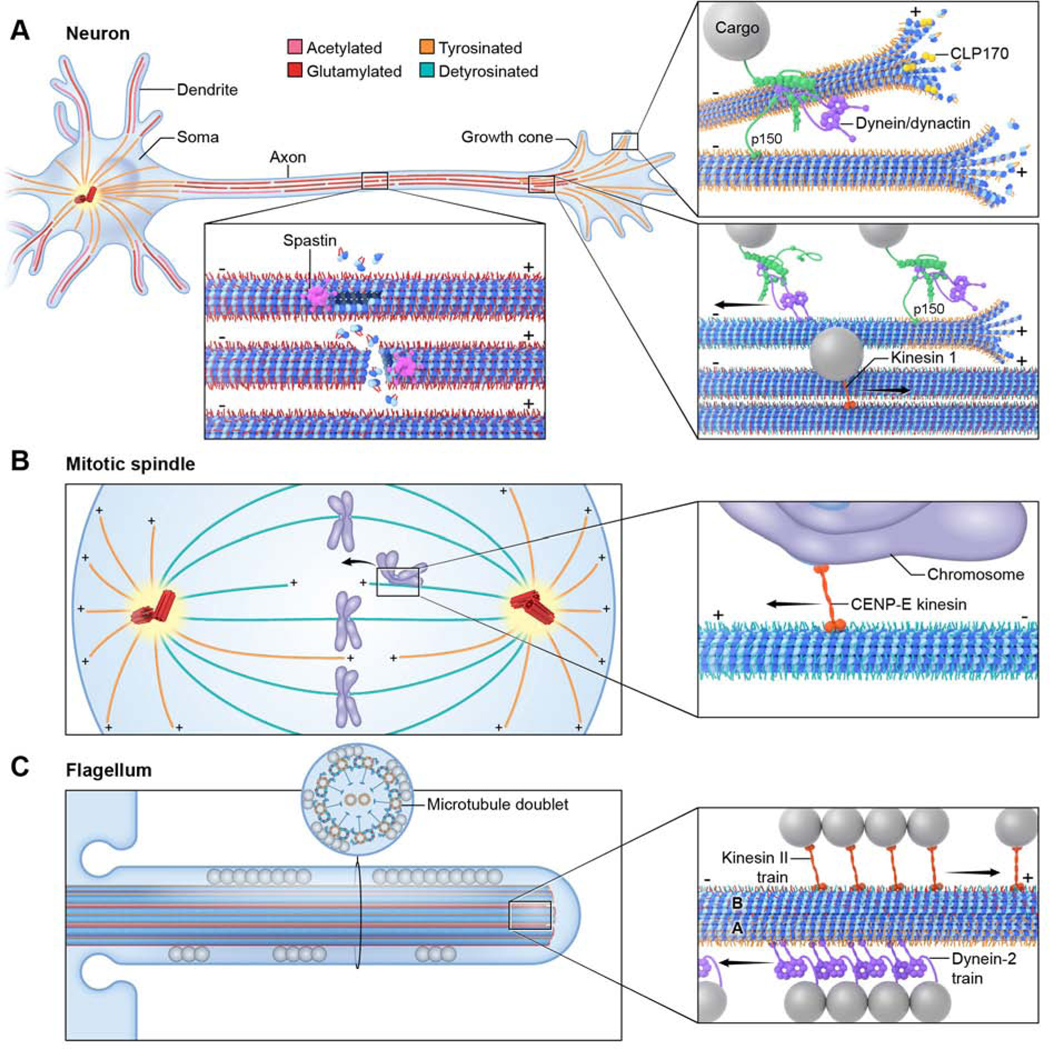
(A) Schematic of a neuron with microtubule and posttranslational modifications colored as in Figure 3. Insets show in clockwise direction, spastin recruited preferentially to glutamylated microtubules, CLIP170 and dynein-dynactin recruited to the plus ends of microtubules enriched in tyrosination, and dynein/dynactin-initiated motility at tyrosinated microtubule sections through the Cap-Gly domain in p150/dynactin; once initiated, motility is independent of the microtubule tyrosination status.
(B) Schematic of the mitotic spindle with microtubule and posttranslational modifications colored as in Figure 3. Inset shows preferential recruitment of chromosome-associated CENP-E to detyrosinated microtubules.
(C) Schematic of a flagellum with microtubules and posttranslational modifications colored as in Figure 3. Spheres denote cargo trains. Inset shows designated transport lanes on the microtubule doublet with kinesin anterograde trains running on the glutamylated, glycylated- and detyrosinated-enriched B-tubule and dynein retrograde trains running on the tyrosinated A-tubule.
Because glutamylation involves the addition of variable glutamate numbers, it can elicit graded responses. The microtubule severing activity of spastin is regulated by the number of glutamates on tubulin tails, providing precise, substrate driven control of microtubule stability (Valenstein and Roll-Mecak, 2016)(Figure 4A). Thus, tubulin modifications can be highly information-rich. Given the high abundance of glutamylated tubulin in neurons and axonemes, it is not surprising that perturbation of glutamylation levels leads to broad neuronal and ciliary phenotypes (reviewed in (Magiera et al., 2018b)). The effects of glycylation on motor or MAP behavior has so far not been investigated, largely because of a lack of a biochemical platform to generate microtubules with controlled glycylation levels.
The detyrosination/tyrosination cycle of α-tubulin plays important roles in many biological processes ranging from cell division to neuronal trafficking and cardiomyocyte beating. Early studies identified detyrosination as a signal for the recruitment of vimentin intermediate filaments by kinesin (Gurland and Gundersen, 1995; Kreitzer et al., 1999). Detyrosination also guides chromosomes towards the cell equator during mitosis by increasing the time the chromosome-associated CENP-E motor remains attached to microtubules under load (Barisic et al., 2015)(Figure 4B). Asymmetric detyrosination of microtubules in the female meiotic spindle, achieved through CDC42-dependent signaling from the cortex, drives non-Mendelian segregation of selfish elements (Akera et al., 2017) through a yet unknown molecular mechanism. The preferential interaction of the depolymerizing kinesin-13 MCAK with tyrosinated microtubules contributes to the higher turnover of tyrosinated microtubules in cells and an imbalance between detyrosination and tyrosination results in spindle positioning defects (Peris et al., 2009). The detyrosination/tyrosination cycle also affects microtubule dynamics by regulating the recruitment of Cap-Gly containing proteins such as CLIP170 to growing microtubule ends (Peris et al., 2006) (Figure 4A). CLIP170 promotes fast actin polymerization from microtubule ends via recruitment of the formin mDia1 (Henty-Ridilla et al., 2016). Thus, tyrosination dependent localization of CLIP170 could mediate communication between actin and microtubule arrays. Tyrosinated microtubules also recruit the dynein-dynactin complex through the Cap-Gly domain of the p150Glued subunit of dynactin (McKenney et al., 2016; Nirschl et al., 2016; Peris et al., 2006). This interaction is used synergistically with CLIP170 to load retrograde vesicles in the distal axon, enriched in tyrosinated microtubules (Nirschl et al., 2016); Figure 4A). The tyrosination dependent recruitment of dynein-dynactin is also important for centrosome positioning (Barbosa et al., 2017).
Desmin intermediate filaments couple the sarcomere to the microtubule. The binding to the microtubule is strongly dependent on detyrosination. (Robison et al., 2016). Because microtubules provide mechanical resistance during cardiac contraction, an increase in detyrosination negatively regulates contractility. Strikingly, cardiomyocytes from failing hearts have heavily detyrosinated microtubules. Reduction in detyrosination reduces stiffness and partially restores contractile function (Chen et al., 2018), revealing a potential point of therapeutic intervention. Motor traffic can also be controlled indirectly by the tubulin code through its effects on microtubule organization. For example, Lys40 acetylation has not been shown so far to have a direct effect on motors (Kaul et al., 2014; Walter et al., 2012), but it promotes the formation of microtubule bundles on which kinesin-1 traffics preferentially (Balabanian et al., 2017; Tas et al., 2017). This preference is important for kinesin-1 to drive the entry of simian virus 40 into the cytosol (Ravindran et al, 2017).
Recent years have seen a growing number of examples of microtubule effectors regulated by individual posttranslational modifications. We will likely soon have a comprehensive characterization of the response to the tubulin code of major cytoskeletal regulators through targeted as well as proteomic approaches. The effects of the microtubule track on motor stalling forces have been largely unexplored, with the exception of CENP-E (Barisic et al., 2015) and this is an area of physiological importance since motors carry large cargo in constrained environments in cells. Multiple posttranslational modifications occur on the same tubulin dimer or on the same microtubule in vivo and it is not yet known whether tubulin modifications act combinatorially. What is the minimal mosaic block on a microtubule enriched in a particular modification that can elicit a response from a cellular effector? What are the effects of different modification types and levels on neighboring microtubules? The physiological output of the tubulin code can be highly sensitive to the concentration of the microtubule effectors and the spatial distribution of microtubules with different modifications. For example, spastin-mediated microtubule stability can be regulated in trans by neighboring microtubules depending on their glutamylation levels. Glutamylated microtubules sequester the enzyme and protect nearby unmodified microtubules from severing (Valenstein and Roll-Mecak, 2016). Thus, cell types with different concentrations of an effector will respond differently to the same changes in the levels of a tubulin modification. Here the microtubule field may learn from the analytical and system approaches in the cell signaling field.
Pattern generation, maintenance and plasticity: the tubulin code writers and erasers
The in vivo chemical complexity of tubulin is generated by the collective action of tubulin modifying enzymes, the code writers and erases. These enzymes belong to diverse protein families ranging from acetyltransferases and kinases to transglutaminases, ATP-dependent ligases and carboxypeptidases. They have roles in cell cycle regulation, neuronal development and cilia function (reviewed in (Yu et al., 2015). Their mutation has been linked to cancers, neurodevelopmental and neurodegenerative disorders (reviewed in (Magiera et al., 2018b)).
The genetically encoded tyrosine on α-tubulin is removed by the vasohibins VASH1 and 2 (Aillaud et al., 2017; Nieuwenhuis et al., 2017), although these are likely not the only enzymes with this activity (Nieuwenhuis et al., 2017). Vasohibins are angiogenesis regulators; however, the role of detyrosination in this pathway is not yet known. Detyrosination is reversed by tubulin tyrosine ligase (TTL) (Ersfeld et al., 1993; Raybin and Flavin, 1977). TTL knockout mice die shortly after birth due to disorganized neuronal arrays (Erck et al., 2005). Higher levels of microtubule detyrosination are associated with tumor aggressiveness and poor prognosis, and TTL inactivation is common in carcinomas and sarcomas (Whipple et al., 2010). Interestingly, VASH2 is expressed in cancer and promotes tumor growth by stimulating angiogenesis (Sato, 2012). Thus, tumors upregulate detyrosination by dysregulation of both TTL and vasohibins.
While the first tubulin modification enzyme, TTL, was isolated in 1977 (Raybin and Flavin, 1977), many tubulin modification enzymes were identified in the last fifteen years. These include the enzymes responsible for the most abundant and evolutionarily widespread tubulin modifications, glutamylation (Janke et al., 2005; van Dijk et al., 2007) and glycylation (Wloga et al., 2009). Glutamylases and glycylases belong to the tubulin tyrosine ligase-like (TTLL) family and share a core structurally homologous to TTL (Garnham et al., 2015; Garnham et al., 2017), consistent with the fact that they all catalyze the RNA-independent ligation of amino acids. Mammals encode thirteen TTLLs. TTLL1, 2, 4, 5, 6, 7, 9, 11 and 13 are glutamylases, while TTLL3, 8, and 10 are glycylase (reviewed in (Yu et al., 2015)). So far, no enzymatic activity has been reported for TTLL12. TTLL enzymes are specialized either to initiate the glutamate or glycine chains by catalyzing the formation of a branch point formed by the addition of a glutamate or glycine to the γ-carboxyl group of an internal glutamate in the tubulin tail, or for elongating these chains once the branch point is formed (Figure 2). Recent studies using purified enzymes and well-defined tubulin substrates demonstrated these segregated activities for TTLL enzymes and defined the active site signatures that specialize TTLLs into glutamate and glycine elongases and initiases (Mahalingan et al., 2020). It is not yet clear whether the exact position of the modification on the tubulin tails is important, or just its presence. Tubulin isolated from brain tissue or axonemes has only a subset of the glutamates in the tails glutamylated or glycylated (Redeker et al., 1998; Wall et al., 2016) and modification enzymes (TTLL6, 7 and 3) have a preference for certain modification sites (Mahallingan et al., 2020; Garnham et al., 2017), indicating some substrate specificity. Intriguingly, experiments in Tetrahymena using tubulin mutants suggest that what is critical is that a tubulin tail carry the modification, even if it is grafted on the body of the other protomer (Xia et al., 2000). Studies in other systems that systematically mutate tubulin modification sites will be needed to gain a clearer understanding of the positional effects of modifications on the tubulin tails.
The action of TTLLs is reversed by the M14D subfamily of metallo-dependent carboxypeptidases, CCPs (Berezniuk et al., 2012; Kimura et al., 2010; Rogowski et al., 2010). Mammals express six CCPs (CCP1–6) with different substrate specificities. Overexpression experiments and use of modification-specific antibodies indicate that CCP1, 2, 3, 4, and 6 shorten polyglutamate chains and produce Δ2-tubulin, while CCP5 removes branching point glutamates (Rogowski et al., 2010). To date, no enzyme with tubulin deglycylating activity has been identified. Both TTLL and CCP enzymes are linked to defects in cilia function and neuronal development. In C. elegans, glutamylation by TTLL11 controls the architecture of the AB doublet in the cilia of sensory neurons, regulates the velocity of kinesin-2 OSM-3/KIF17 and kinesin-3 KLP-6 and is required for ciliary extracellular vesicle release (O’Hagan et al., 2017). TTLL11 loss is epistatic to CCP1 loss in this system. Excessive glutamylation in the mouse cerebellum due to CCP1 loss causes Purkinje cell degeneration and impairs axonal transport (Magiera et al., 2018a; Rogowski et al., 2010). The neurodegenerative phenotype can be rescued by downregulating the glutamylase TTLL1. CCP1 mutations are also linked to infantile-onset neurodegeneration (Shashi et al., 2018). Thus, perturbation of tubulin glutamylation is likely an important mechanism contributing to neurodegeneration.
Even though acetylation of α-tubulin Lys40 was described in 1986 (Maruta et al., 1986), it took more than twenty years to identify the acetyltransferase responsible for this modification, TAT (Akella et al., 2010; Shida et al., 2010). Tubulin is deacetylated by histone deacetylase 6 and NAD-dependent deacetylase sirtuin2, enzymes extensively studied in the context of histone modifications (reviewed in (Garnham and Roll‐Mecak, 2012)). Lys40 was also found recently to be methylated by the Set2 histone methyltransferase (Park et al., 2016), thus providing another common regulator between the tubulin and histone codes.
Tubulin phosphorylation is poorly characterized and many phosphorylation sites have no identified kinases so far. β-tubulin is phosphorylated on Ser172 by cyclin-dependent kinase 1 in mitotic cells (Fourest-Lieuvin et al., 2006) and by the down syndrome minibrain/DYRK1a kinase in neurons where it regulates dendritic arborization (Ori-McKenney et al., 2016). Phosphorylation of α-tubulin Ser165 by protein kinase C enhances motility by promoting microtubule growth and suppressing catastrophe (De et al., 2014). Polyamination is catalyzed by transglutaminases (Figure 2) Transglutaminases catalyze the polyamination of both soluble tubulin and tubulin incorporated in microtubules (Song et al., 2013). They have functions in neuronal differentiation and degeneration (Jeitner et al., 2009), but it is not clear whether these functions are executed only through the modification of tubulin or also other substrates.
For many of the cases examined so far, there is an asymmetry in the free tubulin dimer versus microtubule preference for the code writers and erases. Early experiments established that detyrosination occurs post-polymerization. Once the microtubule depolymerizes, the detyrosinated tubulin is converted to the tyrosinated form (Gundersen et al., 1987). The recently identified tyrosine carboxypeptidase VASH1/2 does indeed prefer the microtubule polymer (Nieuwenhuis et al., 2017), while TTL tyrosinates the tubulin dimer (Raybin and Flavin, 1975). TTLLs also prefer the polymer substrate (Garnham et al., 2015; Mukai et al., 2009), although this has not been experimentally demonstrated for all TTLLs. TAT, despite having to access the microtubule lumen to acetylate Lys40, also prefers the polymeric state of tubulin (Kormendi et al., 2012; Shida et al., 2010) by recognizing the luminal interface between protofilaments (Szyk et al., 2014). Recent structural and molecular dynamics studies indicate that acetylation reduces the conformational flexibility of the acetylation loop, and possibly affects lateral contacts between protofilaments (Eshun-Wilson et al., 2019). The eraser HDAC6 prefer the soluble tubulin dimer (Miyake et al., 2016). Thus, the tubulin code can be reset by tubulin depolymerization if the catalytic rates of the writers and erasers are so balanced. In addition, recent work showing the exchange of lattice tubulin dimers with the soluble tubulin pool as a result of buckling (Schaedel et al., 2015) or the action of severing enzymes (Vemu et al., 2018), raises the question of whether the modifications on the lattice can be edited without depolymerization since the tubulin dimers incorporated from the soluble tubulin pool will have a different history and thus different tubulin modifications than those on the pre-existing microtubule lattice. Such an editing mechanism would create islands within a microtubule that have different lattice structures, as newly incorporated tubulin is bound to GTP, while the old microtubule lattice contains GDP-tubulin (Aumeier et al., 2016; Vemu et al., 2018), but also different posttranslational modifications. Soluble tubulin, which would be incorporated at repair sites is mostly unmodified, while older microtubules accumulate glutamylation and acetylation, the former being a regulator of microtubule severing (Lacroix et al., 2010; Valenstein and Roll-Mecak, 2016).
The complex and diverse spatial and temporal microtubule modification patterns in cells are a function of the expression levels, subcellular distributions, substrate specificity (tubulin versus microtubules; α- versus β-tubulin) and kinetic parameters of tubulin posttranslational modification enzymes. In addition to these first order factors, pre-existing modifications as well as MAPs can influence the addition of new modifications since many MAPs interact with the microtubule C-terminal tails. For example, cilia and spindle-associated protein (CSAP) has been recently identified as a positive regulator of TTLL glutamylases (Bompard et al., 2018). Thus, the tubulin code likely coexists in an intimate relation with a MAP code. Moreover, the intersection between the lifetime of the microtubule, which can be influenced by MAPs, and the kinetic rate of the modification enzymes can generate microtubule arrays where microtubules with different modifications are in close juxtaposition even when the enzymes are broadly distributed. Once the unitary response to each individual modification is quantitatively characterized, deciphering this multilevel regulatory complexity will require in vitro reconstitution experiments with multiple modification enzymes and chemically defined tubulin substrates, coupled with optogenetic and chemical perturbations in cells that will allow quick inactivation or redistribution of modification enzymes to interrogate effects on pattern formation and functional outputs.
Concluding remarks and perspectives
More than 50 years after microtubules were characterized and named (Ledbetter and Porter, 1963; Slautterback, 1963) and tubulin was isolated (Borisy and Taylor, 1967; Shelanski and Taylor, 1967), we still have much to uncover about the physical principles that govern the dynamic behavior and mechanical properties of this cytoskeletal polymer and how these properties are modulated by isoform-specific sequence variations and posttranslational modifications. Deciphering the tubulin code is a perfect problem for the new interdisciplinary era in biology. It will require physical chemistry to understand non-covalent polymer properties and their modulation by strongly polyanionic intrinsically disordered regions (IDRs); forward genetics to discover signaling pathways that regulate tubulin diversity and reverse genetics to interrogate the contribution of tubulin diversity to cell autonomous, tissue and organism behavior. CRISPR technology in non-model organisms that have pushed their microtubule architectures to extremes for specialized functions promises to yield insights into the key evolutionary levers in tubulin sequence and modifications. Cell biology and proteomics coupled with new faster high-resolution light microscopy and high-resolution ultrastructural characterization using cryo-focused ion beam milling scanning electron microscopy (cryo-FIB SEM) will provide detailed cellular maps of microtubule modifications and isoform usage and how they change with stress, developmental stage or synaptic activity. Quantitative in vitro reconstitution will continue to be key to these efforts because cells have likely devised combinatorial, complex means to regulate microtubule behavior due to their centrality to the survival of the eukaryotic cell.
Acknowledgements
We thank E. Tyler (NIH Medical Arts) and E.A Zehr (National Institute of Neurological Disorders and Stroke) for help with illustrations and R. McIntosh (University of Colorado, Boulder) for useful discussions. A.R.M. is supported by the intramural programs of the National Institute of Neurological Disorders and Stroke (NINDS) and the National, Heart, Lung and Blood Institute (NHLBI).
Footnotes
Microtubule structure, dynamics, and mechanics are modulated by a cell’s choice of tubulin isoforms and posttranslational modifications, which then influence microtubule arrays’ function across cell types, cell cycle and developmental stages. Roll-Mecak presents a Review discussing tubulin diversity and highlighting progress in uncovering the mechanistic underpinnings of the tubulin code.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Aillaud C., Bosc C., Peris L., Bosson A., Heemeryck P., Van Dij J., Le Friec J., Boulan B., Vossier F., and Sanman LE. (2017). Vasohibins/SVBP are tubulin carboxypeptidases (TCPs) that regulate neuron differentiation. Science 358, 1448–1453. [DOI] [PubMed] [Google Scholar]
- Akella JS, Wloga D, Kim J, Starostina NG, Lyons-Abbott S, Morrissette NS, Dougan ST, Kipreos ET, and Gaertig J (2010). MEC-17 is an alpha-tubulin acetyltransferase. Nature 467, 218–222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akera T, Chmátal L, Trimm E, Yang K, Aonbangkhen C, Chenoweth DM, Janke C, Schultz RM, and Lampson MA (2017). Spindle asymmetry drives non-Mendelian chromosome segregation. Science 358, 668–672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arce CA, Rodriguez JA, Barra HS, and Caputo R (1975). Incorporation of L-tyrosine, L-phenylalanine and L-3,4-dihydroxyphenylalanine as single units into rat brain tubulin. Eur J Biochem 59, 145–149. [DOI] [PubMed] [Google Scholar]
- Aumeier C, Schaedel L, Gaillard J, John K, Blanchoin L, and Théry M (2016). Self-repair promotes microtubule rescue. Nature cell biology 18, 1054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayaz P, Ye X, Huddleston P, Brautigam CA, and Rice LM (2012). A TOG: αβ-tubulin complex structure reveals conformation-based mechanisms for a microtubule polymerase. Science 337, 857–860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balabanian L, Berger CL, and Hendricks AG (2017). Acetylated microtubules are preferentially bundled leading to enhanced kinesin-1 motility. Biophysical journal 113, 1551–1560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banerjee A., Roach MC., Trcka P., and Luduena RF. (1992). Preparation of a monoclonal antibody specific for the class IV isotype of beta-tubulin. Purification and assembly of alpha beta II, alpha beta III, and alpha beta IV tubulin dimers from bovine brain. Journal of Biological Chemistry 267, 5625–5630. [PubMed] [Google Scholar]
- Barbosa DJ, Duro J, Prevo B, Cheerambathur DK, Carvalho AX, and Gassmann R (2017). Dynactin binding to tyrosinated microtubules promotes centrosome centration in C. elegans by enhancing dynein-mediated organelle transport. PLoS genetics 13, e1006941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barisic M, e Sousa RS, Tripathy SK, Magiera MM, Zaytsev AV, Pereira AL, Janke C, Grishchuk EL, and Maiato H (2015). Microtubule detyrosination guides chromosomes during mitosis. Science 348, 799–803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes G, Louie KA, and Botstein D (1992). Yeast proteins associated with microtubules in vitro and in vivo. Molecular Biology of the Cell 3, 29–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berezniuk I, Vu HT, Lyons PJ, Sironi JJ, Xiao H, Burd B, Setou M, Angeletti RH, Ikegami K, and Fricker LD (2012). Cytosolic carboxypeptidase 1 is involved in processing α- and β-tubulin. Journal of Biological Chemistry 287, 6503–6517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bompard G, Van Dijk J, Cau J, Lannay Y, Marcellin G, Lawera A, Van Der Laan S, and Rogowski K (2018). CSAP Acts as a Regulator of TTLL-Mediated Microtubule Glutamylation. Cell reports 25, 2866–2877. e2865. [DOI] [PubMed] [Google Scholar]
- Borisy GG., and Taylor E. (1967). The mechanism of action of colchicine: binding of colchincine-3H to cellular protein. The Journal of cell biology 34, 525–533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosch Grau M, Gonzalez Curto G, Rocha C, Magiera MM, Marques Sousa P, Giordano T, Spassky N, and Janke C (2013). Tubulin glycylases and glutamylases have distinct functions in stabilization and motility of ependymal cilia. J Cell Biol 202, 441–451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brady ST (1985). A novel brain ATPase with properties expected for the fast axonal transport motor. Nature 317, 73. [DOI] [PubMed] [Google Scholar]
- Brady ST, Tytell M, and Lasek RJ (1984). Axonal tubulin and axonal microtubules: biochemical evidence for cold stability. The Journal of cell biology 99, 1716–1724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breuss MW, Leca I, Gstrein T, Hansen AH, and Keays DA (2017). Tubulins and brain development–The origins of functional specification. Molecular and Cellular Neuroscience. [DOI] [PubMed] [Google Scholar]
- Burke D, Gasdaska P, and Hartwell L (1989). Dominant effects of tubulin overexpression in Saccharomyces cerevisiae. Molecular and cellular biology 9, 1049–1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaaban S, Jariwala S, Hsu C-T, Redemann S, Kollman JM, Müller-Reichert T, Sept D, Bui KH, and Brouhard GJ (2018). The structure and dynamics of C. elegans tubulin reveals the mechanistic basis of microtubule growth. Developmental cell 47, 191–204. e198. [DOI] [PubMed] [Google Scholar]
- Chen CY, Caporizzo MA, Bedi K, Vite A, Bogush AI, Robison P, Heffler JG, Salomon AK, Kelly NA, and Babu A (2018). Suppression of detyrosinated microtubules improves cardiomyocyte function in human heart failure. Nature medicine 24, 1225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu C-W., Hou F., Zhang J., Phu L., Loktev AV., Kirkpatrick DS., Jackson PK., Zhao Y., and Zou H. (2011). A novel acetylation of β-tubulin by San modulates microtubule polymerization via down-regulating tubulin incorporation. Molecular biology of the cell 22, 448–456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cleveland DW, Hwo S-Y, and Kirschner MW (1977). Physical and chemical properties of purified tau factor and the role of tau in microtubule assembly. Journal of molecular biology 116, 227–247. [DOI] [PubMed] [Google Scholar]
- Cleveland DW, Lopata MA, MacDonald RJ, Cowan NJ, Rutter WJ, and Kirschner MW (1980). Number and evolutionary conservation of α-and β-tubulin and cytoplasmic β-and γ-actin genes using specific cloned cDNA probes. Cell 20, 95–105. [DOI] [PubMed] [Google Scholar]
- Cleveland DW, Lopata MA, Sherline P, and Kirschner MW (1981). Unpolymerized tubulin modulates the level of tubulin mRNAs. Cell 25, 537–546. [DOI] [PubMed] [Google Scholar]
- Cueva JG, Hsin J, Huang KC, and Goodman MB (2012). Posttranslational acetylation of α-tubulin constrains protofilament number in native microtubules. Current Biology 22, 1066–1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De S, Tsimounis A, Chen X, and Rotenberg SA (2014). Phosphorylation of α‐tubulin by protein kinase C stimulates microtubule dynamics in human breast cells. Cytoskeleton 71, 257–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desai A., and Mitchison TJ. (1997). Microtubule polymerization dynamics. Annual review of cell and developmental biology 13, 83–117. [DOI] [PubMed] [Google Scholar]
- Detrich HW, Parker SK, Williams RC, Nogales E, and Downing KH (2000). Cold adaptation of microtubule assembly and dynamics structural interpretation of primary sequence changes present in the α-and β-tubulins of antarctic fishes. Journal of Biological Chemistry 275, 37038–37047. [DOI] [PubMed] [Google Scholar]
- Erck C, Peris L, Andrieux A, Meissirel C, Gruber AD, Vernet M, Schweitzer A, Saoudi Y, Pointu H, Bosc C, et al. (2005). A vital role of tubulin-tyrosine-ligase for neuronal organization. Proc Natl Acad Sci U S A 102, 7853–7858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ersfeld K, Wehland J, Plessmann U, Dodemont H, Gerke V, and Weber K (1993). Characterization of the tubulin-tyrosine ligase. J Cell Biol 120, 725–732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eshun-Wilson L, Zhang R, Portran D, Nachury MV, Toso DB, Löhr T, Vendruscolo M, Bonomi M, Fraser JS, and Nogales E (2019). Effects of α-tubulin acetylation on microtubule structure and stability. Proceedings of the National Academy of Sciences 116, 10366–10371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feit H, Slusarek L, and Shelanski ML (1971). Heterogeneity of tubulin subunits. Proceedings of the National Academy of Sciences 68, 2028–2031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fourest-Lieuvin A, Peris L, Gache V, Garcia-Saez I, Juillan-Binard C, Lantez V, and Job D (2006). Microtubule regulation in mitosis: tubulin phosphorylation by the cyclin-dependent kinase Cdk1. Molecular biology of the cell 17, 1041–1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukushige T., Siddiqui ZK., Chou M., Culotti JG., Gogonea CB., Siddiqui SS., and Hamelin M. (1999). MEC-12, an alpha-tubulin required for touch sensitivity in C. elegans. Journal of cell science 112, 395–403. [DOI] [PubMed] [Google Scholar]
- Fulton C, and Simpson PA (1976). Selective synthesis and utilization of flagellar tubulin. The multi-tubulin hypothesis. Cell motility 3, 987–1005. [Google Scholar]
- Gan PP, McCarroll JA, Po’uha ST, Kamath K, Jordan MA, and Kavallaris M (2010). Microtubule dynamics, mitotic arrest, and apoptosis: drug-induced differential effects of betaIII-tubulin. Mol Cancer Ther 9, 1339–1348. [DOI] [PubMed] [Google Scholar]
- Garnham CP, and Roll‐Mecak A (2012). The chemical complexity of cellular microtubules: Tubulin post‐translational modification enzymes and their roles in tuning microtubule functions. Cytoskeleton 69, 442–463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garnham CP, Vemu A, Wilson-Kubalek EM, Yu I, Szyk A, Lander GC, Milligan RA, and Roll-Mecak A (2015). Multivalent Microtubule Recognition by Tubulin Tyrosine Ligase-like Family Glutamylases. Cell 161, 1112–1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garnham CP, Yu I, Li Y, and Roll-Mecak A (2017). Crystal structure of tubulin tyrosine ligase-like 3 reveals essential architectural elements unique to tubulin monoglycylases. Proceedings of the National Academy of Sciences, 201617286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gasic I, Boswell SA, and Mitchison TJ (2019). Tubulin mRNA stability is sensitive to change in microtubule dynamics caused by multiple physiological and toxic cues. PLoS biology 17, e3000225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geimer S., Teltenkötter A., Plessmann U., Weber K., and Lechtreck KF. (1997). Purification and characterization of basal apparatuses from a flagellate green alga. Cell motility and the cytoskeleton 37, 72–85. [DOI] [PubMed] [Google Scholar]
- Geyer EA, Burns A, Lalonde BA, Ye X, Piedra F-A, Huffaker TC, and Rice LM (2015). A mutation uncouples the tubulin conformational and GTPase cycles, revealing allosteric control of microtubule dynamics. Elife 4, e10113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guardia CM, Farías GG, Jia R, Pu J, and Bonifacino JS (2016). BORC functions upstream of kinesins 1 and 3 to coordinate regional movement of lysosomes along different microtubule tracks. Cell reports 17, 1950–1961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gundersen GG, Khawaja S, and Bulinski JC (1987). Postpolymerization detyrosination of alpha-tubulin: a mechanism for subcellular differentiation of microtubules. J Cell Biol 105, 251–264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gundersen GG, Khawaja S, and Bulinski JC (1989). Generation of a stable, posttranslationally modified microtubule array is an early event in myogenic differentiation. J Cell Biol 109, 2275–2288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo J, Qiang M, and Ludueña RF (2011). The distribution of β-tubulin isotypes in cultured neurons from embryonic, newborn, and adult mouse brains. Brain research 1420, 8–18. [DOI] [PubMed] [Google Scholar]
- Gupta ML Jr, Bode CJ, Thrower DA, Pearson CG, Suprenant KA, Bloom KS, and Himes RH (2002). β-Tubulin C354 mutations that severely decrease microtubule dynamics do not prevent nuclear migration in yeast. Molecular biology of the cell 13, 2919–2932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gurland G., and Gundersen GG. (1995). Stable, detyrosinated microtubules function to localize vimentin intermediate filaments in fibroblasts. The Journal of cell biology 131, 1275–1290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henty-Ridilla JL, Rankova A, Eskin JA, Kenny K, and Goode BL (2016). Accelerated actin filament polymerization from microtubule plus ends. Science 352, 1004–1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horio TH, H. (1986). Visualization of the dynamic instability of individual microtubules by dark-field microscopy. Nature 321, 605–607. [DOI] [PubMed] [Google Scholar]
- Hoyle HD, Turner FR, Brunick L, and Raff EC (2001). Tubulin sorting during dimerization in vivo. Molecular biology of the cell 12, 2185–2194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hussey SP, and Fritz‐Laylin LK (2019). “The Missing Link”: The Tubulin Mutation Database Connects Over 1500 Missense Mutations With Phenotypes Across Eukaryotes. Cytoskeleton 76, 175–176. [DOI] [PubMed] [Google Scholar]
- Janke C, and Bulinski JC (2011). Post-translational regulation of the microtubule cytoskeleton: mechanisms and functions. Nat Rev Mol Cell Biol 12, 773–786. [DOI] [PubMed] [Google Scholar]
- Janke C, Rogowski K, Wloga D, Regnard C, Kajava AV, Strub J-M, Temurak N, van Dijk J, Boucher D, van Dorsselaer A, et al. (2005). Tubulin polyglutamylase enzymes are members of the TTL domain protein family. Science 308, 1758–1762. [DOI] [PubMed] [Google Scholar]
- Jeitner TM, Pinto JT, Krasnikov BF, Horswill M, and Cooper AJ (2009). Transglutaminases and neurodegeneration. Journal of neurochemistry 109, 160–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkins BV., Saunders HA., Record HL., Johnson-Schlitz DM., and Wildonger J. (2017). Effects of mutating α-tubulin lysine 40 on sensory dendrite development. J Cell Sci 130, 4120–4131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson KA (1998). The axonemal microtubules of the Chlamydomonas flagellum differ in tubulin isoform content. Journal of Cell Science 111, 313–320. [DOI] [PubMed] [Google Scholar]
- Johnson V, Ayaz P, Huddleston P, and Rice LM (2011). Design, overexpression, and purification of polymerization-blocked yeast alphabeta-tubulin mutants. Biochemistry 50, 8636–8644. [DOI] [PubMed] [Google Scholar]
- Kaul N, Soppina V, and Verhey KJ (2014). Effects of α-tubulin K40 acetylation and detyrosination on kinesin-1 motility in a purified system. Biophysical journal 106, 2636–2643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kellogg EH, Hejab NM, Howes S, Northcote P, Miller JH, Díaz JF, Downing KH, and Nogales E (2017). Insights into the distinct mechanisms of action of taxane and non-taxane microtubule stabilizers from cryo-EM structures. Journal of molecular biology 429, 633–646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemphues KJ, Raff RA, Kaufman TC, and Raff EC (1979). Mutation in a structural gene for a beta-tubulin specific to testis in Drosophila melanogaster. Proceedings of the National Academy of Sciences 76, 3991–3995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilmartin JV (1981). Purification of yeast tubulin by self-assembly in vitro. Biochemistry 20, 3629–3633. [DOI] [PubMed] [Google Scholar]
- Kimura Y., Kurabe N., Ikegami K., Tsutsumi K., Konishi Y., Kaplan OI., Kunitomo H., Iino Y., Blacque OE., and Setou M. (2010). Identification of tubulin deglutamylase among Caenorhabditis elegans and mammalian cytosolic carboxypeptidases (CCPs). J Biol Chem 285, 22936–22941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kinoshita K, Arnal I, Desai A, Drechsel DN, and Hyman AA (2001). Reconstitution of physiological microtubule dynamics using purified components. Science 294, 1340–1343. [DOI] [PubMed] [Google Scholar]
- Kormendi V, Szyk A, Piszczek G, and Roll-Mecak A (2012). Crystal structures of tubulin acetyltransferase reveal a conserved catalytic core and the plasticity of the essential N terminus. Journal of Biological Chemistry 287, 41569–41575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreis TE (1987). Microtubules containing detyrosinated tubulin are less dynamic. The EMBO Journal 6, 2597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreitzer G, Liao G, and Gundersen GG (1999). Detyrosination of tubulin regulates the interaction of intermediate filaments with microtubules in vivo via a kinesin-dependent mechanism. Mol Biol Cell 10, 1105–1118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunishima S, Nishimura S, Suzuki H, Imaizumi M, and Saito H (2014). TUBB 1 mutation disrupting microtubule assembly impairs proplatelet formation and results in congenital macrothrombocytopenia. European journal of haematology 92, 276–282. [DOI] [PubMed] [Google Scholar]
- Lacroix B, van Dijk J, Gold ND, Guizetti J, Aldrian-Herrada G, Rogowski K, Gerlich DW, and Janke C (2010). Tubulin polyglutamylation stimulates spastin-mediated microtubule severing. The Journal of cell biology 189, 945–954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechtreck KF., and Geimer S. (2000). Distribution of polyglutamylated tubulin in the flagellar apparatus of green flagellates. Cell motility and the cytoskeleton 47, 219–235. [DOI] [PubMed] [Google Scholar]
- Ledbetter M, and Porter K (1963). A” microtubule” in plant cell fine structure. The Journal of cell biology 19, 239–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis SA, Tian G, and Cowan NJ (1997). The α-and β-tubulin folding pathways. Trends in cell biology 7, 479–484. [DOI] [PubMed] [Google Scholar]
- Li F, Hu Y, Qi S, Luo X, and Yu H (2019). Structural basis of tubulin detyrosination by vasohibins. Nature Structural & Molecular Biology, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao S, Rajendraprasad G, Wang N, Eibes S, Gao J, Yu H, Wu G, Tu X, Huang H, and Barisic M (2019). Molecular basis of vasohibins-mediated detyrosination and its impact on spindle function and mitosis. Cell research, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Z, Gasic I, Chandrasekaran V, Peters N, Shao S, Mitchison TJ, and Hegde RS (2020). TTC5 mediates autoregulation of tubulin via mRNA degradation. Science 367, 100–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu N, Xiong Y, Ren Y, Zhang L, He X, Wang X, Liu M, Li D, Shui W, and Zhou J (2015). Proteomic profiling and functional characterization of multiple post-translational modifications of tubulin. Journal of proteome research 14, 3292–3304. [DOI] [PubMed] [Google Scholar]
- Lopata MA, Havercroft JC, Chow LT, and Cleveland DW (1983). Four unique genes required for β tubulin expression in vertebrates. Cell 32, 713–724. [DOI] [PubMed] [Google Scholar]
- Lu Q., and Luduena RF. (1994). In vitro analysis of microtubule assembly of isotypically pure tubulin dimers. Intrinsic differences in the assembly properties of alpha beta II, alpha beta III, and alpha beta IV tubulin dimers in the absence of microtubule-associated proteins. J Biol Chem 269, 2041–2047. [PubMed] [Google Scholar]
- Ludueña RF (2013). A hypothesis on the origin and evolution of tubulin. In International review of cell and molecular biology (Elsevier), pp. 41–185. [DOI] [PubMed] [Google Scholar]
- Magiera MM, Bodakuntla S, Žiak J, Lacomme S, Sousa PM, Leboucher S, Hausrat TJ, Bosc C, Andrieux A, and Kneussel M (2018a). Excessive tubulin polyglutamylation causes neurodegeneration and perturbs neuronal transport. The EMBO journal 37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magiera MM, Singh P, Gadadhar S, and Janke C (2018b). Tubulin posttranslational modifications and emerging links to human disease. Cell 173, 1323–1327. [DOI] [PubMed] [Google Scholar]
- Mahalingan KK, Keenen EK, Strickland M, Li Y, Liu Y, Ball HL, Tanner ME, Tjandra N and Roll-Mecak A Structural basis for polyglutamate chain initiation and elongation by TTLL family enzymes. Nature Struct. & Molec. Bio. In press. [DOI] [PubMed] [Google Scholar]
- Maruta H, Greer K, and Rosenbaum JL (1986). The acetylation of alpha-tubulin and its relationship to the assembly and disassembly of microtubules. The Journal of cell biology 103, 571–579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKenney RJ, Huynh W, Vale RD, and Sirajuddin M (2016). Tyrosination of α‐tubulin controls the initiation of processive dynein–dynactin motility. The EMBO journal, e201593071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minoura I, Hachikubo Y, Yamakita Y, Takazaki H, Ayukawa R, Uchimura S, and Muto E. (2013). Overexpression, purification, and functional analysis of recombinant human tubulin dimer. FEBS letters 587, 3450–3455. [DOI] [PubMed] [Google Scholar]
- Mitchison TK,M (1984). Dynamic instability of microtubule growth. Nature 312, 237–242. [DOI] [PubMed] [Google Scholar]
- Miyake Y, Keusch JJ, Wang L, Saito M, Hess D, Wang X, Melancon BJ, Helquist P, Gut H, and Matthias P (2016). Structural insights into HDAC6 tubulin deacetylation and its selective inhibition. Nature chemical biology 12, 748. [DOI] [PubMed] [Google Scholar]
- Montecinos-Franjola F, Chaturvedi S, Schuck P, and Sackett DL (2018). All Tubulins are Not Alike: Dimer Dissociation and Monomer Exchange Differ Depending on the Biological Source of Tubulin. Biophysical Journal 114, 504a. [Google Scholar]
- Morley SJ, Qi Y, Iovino L, Andolfi L, Guo D, Kalebic N, Castaldi L, Tischer C, Portulano C, and Bolasco G (2016). Acetylated tubulin is essential for touch sensation in mice. Elife 5, e20813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mukai M, Ikegami K, Sugiura Y, Takeshita K, Nakagawa A, and Setou M (2009). Recombinant mammalian tubulin polyglutamylase TTLL7 performs both initiation and elongation of polyglutamylation on β-tubulin through a random sequential pathway. Biochemistry 48, 1084–1093. [DOI] [PubMed] [Google Scholar]
- Multigner L, Pignot-Paintrand I, Saoudi Y, Job D, Plessmann U, Rüdiger M, and Weber K (1996). The A and B tubules of the outer doublets of sea urchin sperm axonemes are composed of different tubulin variants. Biochemistry 35, 10862–10871. [DOI] [PubMed] [Google Scholar]
- Nielsen MG., Gadagkar SR., and Gutzwiller L. (2010). Tubulin evolution in insects: gene duplication and subfunctionalization provide specialized isoforms in a functionally constrained gene family. BMC evolutionary biology 10, 113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen MG, Turner FR, Hutchens JA, and Raff EC (2001). Axoneme-specific β-tubulin specialization: a conserved C-terminal motif specifies the central pair. Current Biology 11, 529–533. [DOI] [PubMed] [Google Scholar]
- Nieuwenhuis J, Adamopoulos A, Bleijerveld OB, Mazouzi A, Stickel E, Celie P, Altelaar M, Knipscheer P, Perrakis A, and Blomen VA (2017). Vasohibins encode tubulin detyrosinating activity. Science, eaao5676. [DOI] [PubMed] [Google Scholar]
- Nirschl JJ, Magiera MM, Lazarus JE, Janke C, and Holzbaur EL (2016). α-Tubulin tyrosination and CLIP-170 phosphorylation regulate the initiation of dynein-driven transport in neurons. Cell reports 14, 2637–2652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nogales E (2001). Structural insights into microtubule function. Annual review of biophysics and biomolecular structure 30, 397–420. [DOI] [PubMed] [Google Scholar]
- Nogales E, Wolf SG, and Downing KH (1998). Structure of the alpha beta tubulin dimer by electron crystallography. Nature 391, 199–203. [DOI] [PubMed] [Google Scholar]
- O’Hagan R, Silva M, Nguyen KCQ, Zhang W, Bellotti S, Ramadan YH, Hall DH, Barr MM (2017). Glutamylation Regulates Transport, Specializes Function, and Sculpts the Structure of Cilia. Curr. Biol. 27(22):3430–3441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orgel LE. (1968). Evolution of the genetic apparatus. Journal of molecular biology 38, 381–393. [DOI] [PubMed] [Google Scholar]
- Ori-McKenney KM, McKenney RJ, Huang HH, Li T, Meltzer S, Jan LY, Vale RD, Wiita AP, and Jan YN (2016). Phosphorylation of β-tubulin by the down syndrome kinase, Minibrain/DYRK1a, regulates microtubule dynamics and dendrite morphogenesis. Neuron 90, 551–563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pachter JS, Yen TJ, and Cleveland DW (1987). Autoregulation of tubulin expression is achieved through specific degradation of polysomal tubulin mRNAs. Cell 51, 283–292. [DOI] [PubMed] [Google Scholar]
- Pamula MC, Ti S-C, and Kapoor TM (2016). The structured core of human β tubulin confers isotype-specific polymerization properties. The Journal of cell biology 213, 425–433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panda D, Miller HP, Banerjee A, Luduena RF, and Wilson L (1994). Microtubule dynamics in vitro are regulated by the tubulin isotype composition. Proc Natl Acad Sci U S A 91, 11358–11362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park IY, Powell RT, Tripathi DN, Dere R, Ho TH, Blasius TL, Chiang Y-C, Davis IJ, Fahey CC, and Hacker KE (2016). Dual chromatin and cytoskeletal remodeling by SETD2. Cell 166, 950–962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peris L, Thery M, Fauré J, Saoudi Y, Lafanechère L, Chilton JK, Gordon-Weeks P, Galjart N, Bornens M, Wordeman L, et al. (2006). Tubulin tyrosination is a major factor affecting the recruitment of CAP-Gly proteins at microtubule plus ends. J Cell Biol 174, 839–849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peris L., Wagenbach M., Lafanechere L., Brocard J., Moore AT., Kozielski F., Job D., Wordeman L., and Andrieux A. (2009). Motor-dependent microtubule disassembly driven by tubulin tyrosination. J Cell Biol 185, 1159–1166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preitner N, Quan J, Nowakowski DW, Hancock ML, Shi J, Tcherkezian J, Young-Pearse TL, and Flanagan JG (2014). APC is an RNA-binding protein, and its interactome provides a link to neural development and microtubule assembly. Cell 158, 368–382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prota AE, Magiera MM, Kuijpers M, Bargsten K, Frey D, Wieser M, Jaussi R, Hoogenraad CC, Kammerer RA, Janke C, et al. (2013). Structural basis of tubulin tyrosination by tubulin tyrosine ligase. J Cell Biol 200, 259–270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravindran MA, Engelke MF, Verhey KJ and Tsai B (2017). Exploiting the kinesin-1 molecular motor to generate a virus membrane penetration site. Nature Comm. 8:15496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raybin D, and Flavin M (1975). An enzyme tyrosylating alpha-tubulin and its role in microtubule assembly. Biochem Biophys Res Commun 65, 1088–1095. [DOI] [PubMed] [Google Scholar]
- Raybin D, and Flavin M (1977). Enzyme which specifically adds tyrosine to the alpha chain of tubulin. Biochemistry 16, 2189–2194. [DOI] [PubMed] [Google Scholar]
- Redeker V (2010). Mass spectrometry analysis of C-terminal posttranslational modifications of tubulins. Methods in cell biology 95, 77–103. [DOI] [PubMed] [Google Scholar]
- Redeker V, Le Caer JP, Rossier J, and Prome JC (1991). Structure of the polyglutamyl side chain posttranslationally added to alpha-tubulin. J Biol Chem 266, 23461–23466. [PubMed] [Google Scholar]
- Redeker V., Rossier J., and Frankfurter A. (1998). Posttranslational modifications of the C-terminus of α-tubulin in adult rat brain: α4 is glutamylated at two residues. Biochemistry 37, 14838–14844. [DOI] [PubMed] [Google Scholar]
- Rickard JE, and Kreis TE (1990). Identification of a novel nucleotide-sensitive microtubule-binding protein in HeLa cells. The Journal of Cell Biology 110, 1623–1633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robison P, Caporizzo MA, Ahmadzadeh H, Bogush AI, Chen CY, Margulies KB, Shenoy VB, and Prosser BL (2016). Detyrosinated microtubules buckle and bear load in contracting cardiomyocytes. Science 352, aaf0659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogowski K, Juge F, van Dijk J, Wloga D, Strub JM, Levilliers N, Thomas D, Bre MH, Van Dorsselaer A, Gaertig J, et al. (2009). Evolutionary divergence of enzymatic mechanisms for posttranslational polyglycylation. Cell 137, 1076–1087. [DOI] [PubMed] [Google Scholar]
- Rogowski K, van Dijk J, Magiera MM, Bosc C, Deloulme JC, Bosson A, Peris L, Gold ND, Lacroix B, Bosch Grau M, et al. (2010). A family of protein-deglutamylating enzymes associated with neurodegeneration. Cell 143, 564–578. [DOI] [PubMed] [Google Scholar]
- Salinas RE, Ogohara C, Thomas MI, Shukla KP, Miller SI, and Ko DC (2014). A cellular genome-wide association study reveals human variation in microtubule stability and a role in inflammatory cell death. Mol Biol Cell 25, 76–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato Y. (2012). The vasohibin family: a novel family for angiogenesis regulation. The Journal of Biochemistry 153, 5–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savage C, Hamelin M, Culotti J, Coulson A, Albertson DG, and Chalfie M (1989). mec-7 is a beta-tubulin gene required for the production of 15-protofilament microtubules in Caenorhabditis elegans. Genes & development 3, 870–881. [DOI] [PubMed] [Google Scholar]
- Schaedel L, John K, Gaillard J, Nachury MV, Blanchoin L, and Théry M (2015). Microtubules self-repair in response to mechanical stress. Nature materials 14, 1156–1163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider A, Plessmann U, Felleisen R, and Weber K (1998). Posttranslational modifications of trichomonad tubulins; identification of multiple glutamylation sites. FEBS letters 429, 399–402. [DOI] [PubMed] [Google Scholar]
- Schulze E, Asai DJ, Bulinski JC, and Kirschner M (1987). Posttranslational modification and microtubule stability. The Journal of Cell Biology 105, 2167–2177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulze E, and Kirschner M (1987). Dynamic and stable populations of microtubules in cells. The Journal of cell biology 104, 277–288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shah C, Xu CZ-Q, Vickers J, and Williams R (2001). Properties of microtubules assembled from mammalian tubulin synthesized in Escherichia coli. Biochemistry 40, 4844–4852. [DOI] [PubMed] [Google Scholar]
- Shashi V., Magiera MM., Klein D., Zaki M., Schoch K., Rudnik‐Schöneborn S., Norman A., Neto OLA., Dusl M., and Yuan X. (2018). Loss of tubulin deglutamylase CCP1 causes infantile‐onset neurodegeneration. The EMBO journal 37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shelanski M, and Taylor E (1967). Isolation of a protein subunit from microtubules. The Journal of cell biology 34, 549–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shida T, Cueva JG, Xu Z, Goodman MB, and Nachury MV (2010). The major α-tubulin K40 acetyltransferase αTAT1 promotes rapid ciliogenesis and efficient mechanosensation. Proceedings of the National Academy of Sciences 107, 21517–21522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sirajuddin M, Rice LM & Vale RD (2014). Regulation of microtubule motors by tubulin isotypes and post-translational modifications. Nat Cell Bio 16, 335–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slautterback DB (1963). Cytoplasmic microtubules: I. Hydra. The Journal of cell biology 18, 367–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Y, Kirkpatrick LL, Schilling AB, Helseth DL, Chabot N, Keillor JW, Johnson GV, and Brady ST (2013). Transglutaminase and polyamination of tubulin: posttranslational modification for stabilizing axonal microtubules. Neuron 78, 109–123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spiliotis ET, Hunt SJ, Hu Q, Kinoshita M, and Nelson WJ (2008). Epithelial polarity requires septin coupling of vesicle transport to polyglutamylated microtubules. The Journal of cell biology 180, 295–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stepanek L., and Pigino G. (2016). Microtubule doublets are double-track railways for intraflagellar transport trains. Science 352, 721–724. [DOI] [PubMed] [Google Scholar]
- Sullivan KF, and Cleveland DW (1986). Identification of conserved isotype-defining variable region sequences for four vertebrate beta tubulin polypeptide classes. Proceedings of the National Academy of Sciences 83, 4327–4331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szyk A, Deaconescu AM, Piszczek G, and Roll-Mecak A (2011). Tubulin tyrosine ligase structure reveals adaptation of an ancient fold to bind and modify tubulin. Nature structural & molecular biology 18, 1250–1258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szyk A, Deaconescu AM, Spector J, Goodman B, Valenstein ML, Ziolkowska NE, Kormendi V, Grigorieff N, and Roll-Mecak A (2014). Molecular basis for age-dependent microtubule acetylation by tubulin acetyltransferase. Cell 157, 1405–1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tas RP, Chazeau A, Cloin BM, Lambers ML, Hoogenraad CC, and Kapitein LC (2017). Differentiation between oppositely oriented microtubules controls polarized neuronal transport. Neuron 96, 1264–1271. e1265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ti S-C, Alushin GM, and Kapoor TM (2018). Human β-tubulin isotypes can regulate microtubule protofilament number and stability. Developmental cell 47, 175–190. e175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tilney LG, Bryan J, Bush DJ, Fujiwara K, Mooseker MS, Murphy DB, and Snyder DH (1973). Microtubules: evidence for 13 protofilaments. The Journal of cell biology 59, 267–275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Topalidou I., Keller C., Kalebic N., Nguyen KC., Somhegyi H., Politi KA., Heppenstall P., Hall DH., and Chalfie M. (2012). Genetically separable functions of the MEC-17 tubulin acetyltransferase affect microtubule organization. Curr Biol 22, 1057–1065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vale RD (1991). Severing of stable microtubules by a mitotically activated protein in Xenopus egg extracts. Cell 64, 827–839. [DOI] [PubMed] [Google Scholar]
- Vale RD, Reese TS, and Sheetz MP (1985a). Identification of a novel force-generating protein, kinesin, involved in microtubule-based motility. Cell 42, 39–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vale RD, Schnapp BJ, Reese TS, and Sheetz MP (1985b). Organelle, bead, and microtubule translocations promoted by soluble factors from the squid giant axon. Cell 40, 559–569. [DOI] [PubMed] [Google Scholar]
- Valenstein ML, and Roll-Mecak A (2016). Graded control of microtubule severing by tubulin glutamylation. Cell 164, 911–921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Dijk J, Rogowski K, Miro J, Lacroix B, Edde B, and Janke C (2007). A targeted multienzyme mechanism for selective microtubule polyglutamylation. Mol Cell 26, 437–448. [DOI] [PubMed] [Google Scholar]
- Vemu A, Atherton J, Spector JO, Moores CA, and Roll-Mecak A (2017). Tubulin isoform composition tunes microtubule dynamics. Molecular biology of the cell 28, 3564–3572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vemu A, Atherton J, Spector JO, Szyk A, Moores CA, and Roll-Mecak A (2016). Structure and dynamics of single-isoform recombinant Neuronal Human Tubulin. Journal of Biological Chemistry, jbc. C116. 731133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vemu A., Garnham CP., Lee D-Y., and Roll-Mecak A. (2014). Generation of differentially modified microtubules using in vitro enzymatic approaches. Methods Enzymol 540, 149–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vemu A, Szczesna E, Zehr EA, Spector JO, Grigorieff N, Deaconescu AM, and Roll-Mecak A (2018). Severing enzymes amplify microtubule arrays through lattice GTP-tubulin incorporation. Science 361, eaau1504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verhey KJ, and Gaertig J (2007). The tubulin code. Cell cycle 6, 2152–2160. [DOI] [PubMed] [Google Scholar]
- Walczak CE, Mitchison TJ, and Desai A (1996). XKCM1: a Xenopus kinesin-related protein that regulates microtubule dynamics during mitotic spindle assembly. Cell 84, 37–47. [DOI] [PubMed] [Google Scholar]
- Walker RA, O’Brien ET, Pryer NK, Soboeiro MF, Voter WA, Erickson HP & Salmon ED (1988). Dynamic instability of individual microtubules analyzed by video light microscopy: rate constants and transition frequencies. J Cell Biol 107, 1437–1448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wall KP, Pagratis M, Armstrong G, Balsbaugh JL, Verbeke E, Pearson CG, and Hough LE (2016). Molecular determinants of tubulin’s C-terminal tail conformational ensemble. ACS chemical biology 11, 2981–2990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walter WJ, Beranek V, Fischermeier E, and Diez S (2012). Tubulin acetylation alone does not affect kinesin-1 velocity and run length in vitro. PLoS One 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang N, Bosc C, Choi SR, Boulan B, Peris L, Olieric N, Bao H, Krichen F, Chen L, and Andrieux A (2019). Structural basis of tubulin detyrosination by the vasohibin–SVBP enzyme complex. Nature Structural & Molecular Biology, 1. [DOI] [PubMed] [Google Scholar]
- Webster DR., Gundersen GG., Bulinski JC., and Borisy GG. (1987). Differential turnover of tyrosinated and detyrosinated microtubules. Proceedings of the National Academy of Sciences 84, 9040–9044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weisenberg RC (1972). Microtubule formation in vitro in solutions containing low calcium concentrations. Science 177, 1104–1105. [DOI] [PubMed] [Google Scholar]
- Whipple RA, Matrone MA, Cho EH, Balzer EM, Vitolo MI, Yoon JR, Ioffe OB, Tuttle KC, Yang J, and Martin SS (2010). Epithelial-to-mesenchymal transition promotes tubulin detyrosination and microtentacles that enhance endothelial engagement. Cancer Research 70, 8127–8137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widlund PO, Podolski M, Reber S, Alper J, Storch M, Hyman AA, Howard J, and Drechsel DN (2012). One-step purification of assembly-competent tubulin from diverse eukaryotic sources. Molecular biology of the cell 23, 4393–4401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witman GB, Carlson K.t., and Rosenbaum JL (1972). Chlamydomonas flagella: II. The distribution of tubulins 1 and 2 in the outer doublet microtubules. The Journal of cell biology 54, 540–555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wloga D, Webster DM, Rogowski K, Bre MH, Levilliers N, Jerka-Dziadosz M, Janke C, Dougan ST, and Gaertig J (2009). TTLL3 Is a tubulin glycine ligase that regulates the assembly of cilia. Developmental cell 16, 867–876. [DOI] [PubMed] [Google Scholar]
- Wolff A., de Nechaud B., Chillet D., Mazarguil H., Desbruyeres E., Audebert S., Edde B., Gros F., and Denoulet P. (1992). Distribution of glutamylated alpha and beta-tubulin in mouse tissues using a specific monoclonal antibody, GT335. Eur J Cell Biol 59, 425–432. [PubMed] [Google Scholar]
- Wolff A, Denoulet P, and Jeantet C (1982). High level of tubulin microheterogeneity in the mouse brain. Neurosci Lett 31, 323–328. [DOI] [PubMed] [Google Scholar]
- Wolff A, Houdayer M, Chillet D, de Nechaud B, and Denoulet P (1994). Structure of the polyglutamyl chain of tubulin: occurrence of alpha and gamma linkages between glutamyl units revealed by monoreactive polyclonal antibodies. Biol Cell 81, 11–16. [DOI] [PubMed] [Google Scholar]
- Xia L, Hai B, Gao Y, Burnette D, Thazhath R, Duan J, Bre MH, Levilliers N, Gorovsky MA, and Gaertig J (2000). Polyglycylation of tubulin is essential and affects cell motility and division in Tetrahymena thermophila. J Cell Biol 149, 1097–1106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu Z, Schaedel L, Portran D, Aguilar A, Gaillard J, Marinkovich MP, Théry M, and Nachury MV (2017). Microtubules acquire resistance from mechanical breakage through intralumenal acetylation. Science 356, 328–332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan C, Wang F, Peng Y, Williams CR, Jenkins B, Wildonger J, Kim H-J, Perr JB, Vaughan JC, and Kern ME (2018). Microtubule acetylation is required for mechanosensation in Drosophila. Cell reports 25, 1051–1065. e1056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu I, Garnham CP, and Roll-Mecak A (2015). Writing and reading the tubulin code. Journal of Biological Chemistry 290, 17163–17172. [DOI] [PMC free article] [PubMed] [Google Scholar]