

Abstract
Objectives
The objective of this study was to develop a quantitative 5' nuclease real-time polymerase chain reaction (PCR) assay to diagnose infections caused by Bartonella species.
Methods
Between January and April 2013 whole blood samples were collected by convenience from 151 cats (86 domiciled and 65 stray cats). The feline blood samples were subjected to a novel quantitative 5' nuclease real-time PCR (qPCR) for Bartonella species targeting the nictonamide adenine dinucleotide dehydrogenase gamma subunit (nuoG) and conventional PCR assays targeting intergenic transcribed spacer, ribC, gltA, pap31 and rpoB, followed by sequencing and basic local alignment search tool analysis.
Results
The qPCR assay detected as few as 10 copies of plasmid per reaction. Forty-six (54.4% domiciled and 45.6% stray cats) of 151 sampled cats showed positive results in nuoG qPCR for Bartonella species. The absolute quantification of nuoG Bartonella DNA in sampled cats ranged from 1.1 × 104 to 1.3 × 104. Eighteen (39.1%) of 46 positive samples in the qPCR were also positive in conventional PCR assays. The sequencing confirmed that Bartonella henselae and Bartonella clarridgeiae circulate in cats in midwestern Brazil.
Conclusions and relevance
The present work provides details of a novel qPCR assay to diagnose infections caused by Bartonella species.
Introduction
Owing to ecological and climatic changes, better surveillance programs, increased medical interest and new diagnostic techniques – especially advanced molecular tools – several vector-borne pathogens have recently been incriminated as important emerging and re-emerging etiological agents for diseases. 1 Rapid and accurate detection of Bartonella species is essential in the diagnosis of bartonellosis in animals and humans. Although bacterial culture remains the preferred method for identification of Bartonella species infections, this technique is time consuming. Furthermore, conventional PCR (cPCR) assays show limited sensitivity and, owing to the significant genetic diversity within the genus, are not able to detect all species. 2
Bartonella species are fastidious hemotropic gram-negative bacteria, and are mainly transmitted by fleas, ticks, lice, flies and mites. Thirteen of about 33 described Bartonella species or subspecies have been incriminated as disease agents for the following diseases in humans: Carrion’s disease, cat scratch disease, endocarditis, myocarditis, encephalitis, lymphadenopathy, hepatic peliosis, uveitis, fever and weakness.3–5
Among many animal species infected by Bartonella species, cats have been identified as reservoirs for Bartonella henselae, Bartonella koehlerae and Bartonella clarridgeiae, which are human pathogens. 4 Recently, in Brazil, Bartonella species have been molecularly detected in cats in the states of Rio Grande do Sul, 6 Maranhão, 7 Mato Grosso, 8 São Paulo9,10 and Mato Grosso do Sul. 11 These studies emphasized the need for further research to assess the occurrence and geographical distribution of bartonellosis in Brazil.
A broad-range Bartonella species molecular diagnostic tool would be useful in epidemiologic and clinical approaches, providing advantages in the evaluation of vertebrate and invertebrate organisms as hosts, reservoirs and vectors, and promoting a rapid and efficient diagnosis for treatment. To address this, the present work aimed to develop a quantitative 5' nuclease nuoG real-time polymerase chain reaction (PCR) assay to diagnose infections caused by Bartonella species. Additionally, we tested blood samples from domiciled and stray cats for Bartonella DNA using real-time and conventional PCR assays in midwestern Brazil.
Materials and methods
Sampling area, blood collection, DNA extraction and internal control using the 28S rRNA gene
Between January and April 2013, whole blood samples were collected by convenience from 151 cats (54 males, 95 females, two without gender registration) in the city of Campo Grande, which is the capital of the state of Mato Grosso do Sul, Brazil. Free-roaming non-domiciled cats (n = 65) were caught by technical staff from the local zoonosis control center (CCZ). Domiciled cats (n = 86) were sampled during pre-surgical procedures for a castration project at the CCZ; these animals were returned to their homes after surgery. Blood samples were collected in EDTA and immediately stored at −20°C until the moment of DNA extraction (1 month later). The project was approved by the university’s ethics committee (protocol number 004987/13).
Blood samples were defrosted at room temperature and DNA was extracted from 200 µl each whole blood sample using the QIAamp DNA Mini Kit (Qiagen), in accordance with the manufacturer’s instructions. DNA concentration and quality was measured using the absorbance ratio between 260 and 280 nm (Nanodrop). In order to confirm the presence of amplifiable DNA in samples, an internal control PCR targeting the 28S rRNA gene primers feline 28S rDNAFw (5'-AGCAGGAGGT GTTGGAAGAG-3') and feline 28S rDNARv (5'-AGG GAGAGCCTAAATCAAAGG-3') was used. 12
Quantitative 5' nuclease real-time PCR for Bartonella species
Acquisition of sequence data
The sequences of nuoG (nicotinamide adenine dinucleotide [NADH] dehydrogenase gamma subunit), and DNA sequences of Bartonella species (Bartonella quintana [BX897700], B henselae [BX897699], Bartonella grahamii [EF659941], Bartonella tribocorum [AM260525], Bartonella bacilliformis [CP000524], B clarridgeiae [EF659939], Bartonella rochalimae [FN645459], Bartonella washoensis [EF659945], Bartonella koehlerae [EF659942], Bartonella vinsonii subspecies arupensis [EF659936], B vinsonii subspecies vinsonii [EF659944], B vinsonii subspecies berkhofii [EF659937], Bartonella taylori [EF659943], Bartonella elizabethae [EF659940] and Bartonella bovis [EF659938]) were retrieved from the National Center for Biotechnology Information’s Entrez database (www.ncbi.nlm.nih.gov). The sequences for Bartonella birttlesii, Bartonella alsatica, Bartonella chomelli, Bartonella species Th239 and Bartonella schoenbuchensis nuoG were determined by PCR amplification using the primers described here. Purified PCR amplicons (QIAquick Gel Extraction kit; Qiagen) were ligated into a pGEM-T Easy vector (Promega) followed by transformation of JM109 Escherichia coli competent cells (Promega) using the pGEM cloning kit (Promega), according to manufacturer’s instructions. The resulting clones underwent blue/white colony screening. Plasmid DNA of positive clones was isolated using the QIAprep Miniprep Kit (Qiagen) and submitted for sequence determination.
Primers and hydrolysis probe design
The software AlleleID6 (Premier Biosoft) was used to design the TaqMan primers and probe. The gene target included a consensus nuoG Bartonella sequence (alignment of B quintana [BX897700], B henselae [BX897699], B grahamii [EF659941], B tribocorum [AM260525], B bacilliformis [CP000524], B clarridgeiae [EF659939], B rochalimae [FN645459], B washoensis [EF659945], B koehlerae [EF659942], B vinsonii subspecies arupensis [EF659936], B vinsonii subspecies vinsonii [EF659944], B vinsonii subspecies berkhofii [EF659937], B taylori [EF659943], B elizabethae [EF659940] and B bovis [EF659938]).
Real-time detection was enabled with the use of SYBR green fluorescent dye and optimized with the integration of labeled probe. The result was a TaqMan (5'-nuclease) assay for the CFX PCR Detection System (BioRad). Texas Red was used to detect nuoG common to all Bartonella species. Primers and probes were synthesized by Integrated DNA Technologies (IDT).
Primers and probes that met the following conditions were identified (default parameters according AlleleID): primer and probe length of 16–18 nucleotides, primer melting temperature of 50.0 ± 2ºC, probe melting temperature of 7.0 ± 2ºC, amplicon size of 17–120 base pairs (bp). Primer and hydrolysis probe specificity were tested in silico using the basic local alignment search tool (BLAST) algorithms. Aiming for the best amplification conditions, both the annealing temperature and concentration of primers and hydrolysis probe in the reaction, were determined experimentally. The annealing temperature tested ranged from 50°C to 57°C and the concentration of primers and hydrolysis probe ranged from 0.2 to 1.2 µM (Figure 1).
Figure 1.

Optimization of the annealing temperature and concentrations of the primers and hydrolysis probe. The best annealing temperature and concentrations were detected through a range of gradients. Analysis of the results showed that 52.8°C and 1.2 µM were the best annealing temperature and concentration of primers and hydrolysis probe, respectively.
RFU = relative fluorescence units
The sequence of primers and hydrolysis probe obtained using AlleleID6 were F-Bart (5'-CAATCTTCT TTTGCTTCACC-3'), R-Bart (5'- TCAGGGCTTTAT GTGAATAC-3') and TexasRed-5'-TTYGTCATTTGAACA CG-3'[BHQ2a-Q]3', respectively.
The standardization of real-time PCR (qPCR) followed the minimum information for publication of quantitative real-time PCR experiments. 13
Specificity analysis
The specificity of the assay was assessed using DNA from several genomic bacterial (Ehrlichia canis, Ehrlichia chaffeensis, Ehrlichia muris, Ehrlichia species [Anan strain, IOE agent], Anaplasma phagocytophilum, Anaplasma platys, Anaplasma marginale, Anaplasma centrale, Anaplasma ovis, Anaplasma bovis, B henselae, B quintana, B elizabethae, B vinsonii berkhofii, Bartonella phoceensis, Bartonella schoenbuchii, B chomelli, B bovis, Bartonella alsaticae, Bartonella birtlesii e Bartonella Th239, Rickettsia rickettsii, Rickettsia slovaca, Rickettsia typhi, Rickettsia prowazekii, Neorickettsia risticii, Neorickettsia helminthoeca, Neorickettsia sennetsu and Orientia tsutsugamushi) and protozoan (Plasmodium falciparum, Plasmodium malariae, Trypanosoma brucei rodhesiense) species. Bartonella species DNA samples were kindly supplied by Dr Bruno Chomel (University of California, Davis, CA, USA) and Dr Edward Breitschwerdt (North Carolina State University, Raleigh, NC, USA). The T brucei DNA sample was kindly supplied by Dr Dennis John Grab (Johns Hopkins School of Medicine, Baltimore, MD, USA) (Figure 2).
Figure 2.
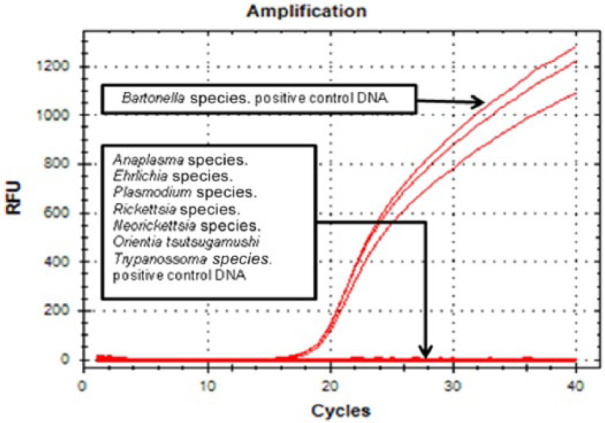
Specificity analysis of the nuoG real-time polymerase chain reaction assay. Amplification curve was observed for only Bartonella species. Positive controls were DNA. No amplification was verified for Ehrlichia species, Anaplasma species, Neorickettsia species, Orientia species, Rickettsia species, Trypanosoma species or Plasmodium species. RFU = relative fluorescence units
Sensitivity analysis
The sensitivity of real-time reactions was determined using 10-fold serial dilutions of pIDTSMART plasmids (Integrated DNA Technologies) encoding the nuoG Bartonella consensus sequence (insert containing 83 bp). The number of plasmid copies was determined according to the formula (X g/µl DNA/[plasmid length in bp × 660]) × 6.022 × 1023 × plasmid copies/µl. Plasmid copy number was adjusted and serially diluted in both Tris-EDTA (TE; 10 mmol/l, Tris-HCl, 0.1 mmol/l, EDTA) (pH 8.0) and TE buffer combined with 30 µg/ml of herring sperm DNA (Sigma-Aldrich) with the aim of imitating the presence of host DNA in the sample.
Amplification efficiency and limit of detection
Amplification efficiency (E) was calculated from the slope of the standard curve in each run using the following formula (E = 10–1/slope). To determine the limit of detection of the TaqMan assay, the standard curves generated by 10-fold dilutions were used to determine the amount of DNA that could be detected with 95% sensitivity. 13
SYBR green and TaqMan PCR reactions
For SYBR green PCR reactions, 10 µl PCR mixtures contained 2 µl of IQ SYBR Green Supermix (BioRad) with a final concentration of 2 µM of each primer and 1 µl of DNA sample. For Taq Man qPCR reactions, the 10 µl PCR mixtures contained 5 µl Go Taq Probe qPCR Master Mix, dTTP (Promega) with a final concentration of 1.2 µM of each primer and hydrolysis probe, and 1 µl DNA sample. PCR amplifications were conducted in low-profile multiplate unskirted PCR plates (BioRad) using a CFX96 Thermal Cycler (BioRad). The amplification conditions were 95°C for 3 mins followed by 40 cycles at 95°C for 10 mins and 52.8°C for 30 s.
Repeatability and reproducibility
The intra-assay variance (repeatability or short-term precision) was determined by running five replicates of the plasmid dilutions (107 to 1 copy of plasmid/reaction) in the same run. The standard curve was then generated using CFX Manager software version 2.1 (BioRad). The inter-assay variance (reproducibility or long-term precision) was determined by running triplicates of the same plasmid dilution in five different runs, on separate days. These replicates were used to determine the mean, SD and coefficient of variation in quantification cycle (Cq) values for each plasmid dilution.
cPCR for Bartonella species
For further molecular characterization and species differentiation, positive samples in nuoG Bartonella amplification were tested using cPCR for other genes: intergenic transcribed spacer (ITS),14,15 ribC, 16 gltA,17,18pap31 19 and rpoB. 15 PCR products were separated by electrophoresis on a 1% agarose gel stained with ethidium bromide. In order to prevent PCR contamination, DNA extraction, reaction set up, PCR amplification and electrophoresis were performed in separate rooms. The gels were imaged under ultraviolet light using Image Lab Software version 4.1 (BioRad).The reaction products were purified using the Silica Bead DNA gel extraction kit (Fermentas). Purified amplified DNA fragments from positive samples were subjected to sequencing for confirmation in an automatic sequencer (ABI Prism 310 genetic analyzer; Applied Biosystems/Perkin-Elmer) and were used for subsequent phylogenetic analysis. Consensus sequences were obtained through analysis of the products from sequencing, from both the forward and the reverse oligonucleotides, using the CAP3 program (http://mobyle.pasteur.fr/cgi-bin/MobylePortal/portal.py). 20 Comparisons with sequences deposited in GenBank were made using BLAST.
Statistical analysis
The point biserial correlation coefficient was used to measure the association between the results obtained in qPCR, expressed in a log-transformed scale L10 = log10 (NuoG+1), and positive results in cPCR. Significance was assessed with the unpaired Student’s t-test with N-2 degrees of freedom (where N is the number of samples).
Aiming to further assess the relationship between the results of cPCR and qPCR, logistic regression models were fitted considering L10 as the explanatory variable and the cPCR result as the response variable (for cases in which point biserial correlation was higher than 0.4). Results were expressed in terms of odds ratio. All analyses were carried using R software. 21
Results
All DNA samples amplified the predicted product for feline 28S rDNA, which indicates a successful DNA extraction.
The efficiency mean of qPCR assays was E = 94.8% (slope = −3.443; r2 = 0.985). As few as 10 copies of plasmid/reaction when using the plasmid standards diluted in TE were detected in this assay. When the plasmid dilutions contained 30 µg/ml of herring sperm, the efficiency was E = 96.5% (slope = −3.380; r2 = 0.989) and the assay also detected as few as 10 copies of plasmid/reaction in all. The intra- and inter-assay repeatabilities of the qPCR are shown in Table 1.
Table 1.
Intra- and inter-assay repeatability of the real-time PCR assay for Bartonella species
| Plasmid copies | Intra-assay repeatability |
Inter-assay repeatability |
||||||
|---|---|---|---|---|---|---|---|---|
| Plasmid diluted in 1 × TE |
Plasmid diluted in DNA |
Plasmid diluted in 1 × TE |
Plasmid diluted in DNA |
|||||
| Mean crossing point (Cq ± SD) | CV (%) | Mean crossing point (Cq ± SD) | CV (%) | Mean crossing point (Cq ± SD) | CV (%) | Mean crossing point (Cq ± SD) | CV (%) | |
| 107 | 13.86 ± 0.13 | 0.94 | 14.31 ± 0.27 | 1.91 | 14.43 ± 0.08 | 0.6 | 13.90 ± 0.55 | 3.96 |
| 106 | 17.31 ± 0.17 | 1.01 | 17.92 ± 0.47 | 2.60 | 18.00 ± 0.24 | 1.3 | 17.31 ± 0.55 | 3.20 |
| 105 | 21.05 ± 0.18 | 0.86 | 21.89 ± 0.16 | 0.74 | 21.95 ± 0.50 | 2.1 | 21.40 ± 0.53 | 2.50 |
| 104 | 24.41 ± 0.30 | 1.24 | 25.44 ± 0.25 | 0.98 | 25.52 ± 0.38 | 1.4 | 25.07 ± 0.43 | 1.74 |
| 10³ | 28.32 ± 0.31 | 1.1 | 29.03 ± 0.27 | 0.94 | 28.84 ± 0.33 | 1.1 | 28.73 ± 0.45 | 1.58 |
| 10² | 31.39 ± 0.14 | 0.44 | 31.76 ± 0.17 | 0.56 | 32.95 ± 0.49 | 1.5 | 31.70 ± 0.45 | 1.43 |
| 10 | 34.99 ± 0.53 | 1.52 | 34.78 ± 0.63 | 1.80 | 35.51 ± 1.41 | 3.9 | 34.14 ± 0.69 | 2.03 |
TE = Tris-EDTA; CV = coefficient of variation; Cq = quantification cycle
The designed primers and probe amplified all genomic DNA from 20 Bartonella species. No amplification signal was obtained from the no template control or from Ehrlichia, Anaplasma, Neorickettsia, Orientia, Rickettsia, Trypanosoma and Plasmodium species.
Forty-six (54.4% domiciled and 45.6% stray cats) of 151 sampled cats showed positive results in nuoG qPCR for Bartonella species. The absolute quantification of nuoG Bartonella DNA in sampled cats ranged from 1.1 × 104 to 1.3 × 104 copies/µl. Eighteen (39.1%) of 46 positive samples in the nuoG Bartonella qPCR were also positive in cPCR assays for Bartonella targeting different genes: 12 cats were positive for ITS, three for rpoB, two for pap-31 and one for gltA. The sequencing confirmed the presence of B clarridgeiae and B henselae circulating in sampled cats (Table 2).
Table 2.
Positive samples for Bartonella species with their respective quantifications in the real-time polymerase chain reaction (qPCR) assay, and the Bartonella species identified by conventional PCR (cPCR) assay and sequencing in cats from Campo Grande, Mato Grosso do Sul, Brazil
| Cat number | Sex | Area of activity | qPCR |
cPCR |
||||
|---|---|---|---|---|---|---|---|---|
| nuoG | ITS | rpoB | pap 31 | ribC | gltA | |||
| 2 | Female | Domiciled | 2.75 × 10¹ | – | – | – | – | – |
| 5 | Female | Domiciled | 2.79 × 10¹ | – | – | – | – | – |
| 7 | Female | Domiciled | 9.70 × 10¹ | – | – | – | – | – |
| 8 | Male | Stray | 2.20 × 10¹ | – | – | – | – | – |
| 13 | Male | Stray | 1.40 × 10² | – | – | – | – | – |
| 14 | Male | Domiciled | 9.50 × 10¹ | – | – | – | – | – |
| 15 | Female | Domiciled | 1.50 × 10¹ | – | – | – | – | – |
| 17 | Female | Domiciled | 1.50 × 10² | – | – | – | – | – |
| 20 | Female | Stray | 3.60 × 10¹ | – | – | – | – | – |
| 21 | Male | Stray | 6.70 × 10¹ | – | – | – | – | – |
| 22 | Male | Stray | 3.90 × 10¹ | – | – | – | – | – |
| 24 | Male | Stray | 2.00 × 10² | – | – | – | – | – |
| 25 | Male | Domiciled | 9.50 × 10¹ | NS | – | – | – | – |
| 27 | Male | Domiciled | 3.80 × 10¹ | – | – | – | – | – |
| 28 | Female | Domiciled | 3.80 × 10¹ | – | – | – | – | – |
| 29 | Female | Domiciled | 3.80 × 10¹ | – | – | – | – | – |
| 30 | Female | Domiciled | 1.70 × 10² | NS | – | – | – | – |
| 31 | Female | Domiciled | 2.60 × 10¹ | – | – | – | – | – |
| 32 | Female | Domiciled | 4.60 × 10¹ | – | – | – | – | – |
| 34 | Female | Domiciled | 2.40 × 10¹ | – | – | – | – | – |
| 37 | Female | Domiciled | 6.80 × 10¹ | – | – | – | – | – |
| 39 | Male | Domiciled | 1.30 × 10² | – | – | – | – | – |
| 50 | Male | Domiciled | 1.90 × 10¹ | – | – | – | – | – |
| 68 | Male | Domiciled | 3.30 × 10¹ | – | – | – | – | – |
| 69 | Male | Domiciled | 1.60 × 10¹ | – | – | – | – | – |
| 74 | Female | Domiciled | 3.80 × 10¹ | – | – | – | – | – |
| 79 | Male | Domiciled | 1.30 × 104 | Bc | NS | – | – | – |
| 86 | Female | Domiciled | 1.90 × 10² | – | – | – | – | – |
| 88 | Female | Domiciled | 4.70 × 10² | – | – | – | – | – |
| 97 | Male | Domiciled | 4.30 × 10¹ | NS | – | – | – | – |
| 100 | Male | Domiciled | 2.40 × 10³ | NS | – | – | – | – |
| 101 | Male | Stray | 2.40 × 10¹ | – | – | – | – | – |
| 102 | Male | Stray | 5.60 × 10³ | NS | – | Bh | – | Bh |
| 108 | Female | Stray | 1.60 × 10² | NS | – | – | – | – |
| 110 | Male | Stray | 2.60 × 10¹ | – | – | – | – | – |
| 111 | Female | Stray | 1.90 × 10¹ | – | – | – | – | – |
| 121 | Female | Stray | 4.40 × 10¹ | NS | NS | – | – | – |
| 124 | Female | Stray | 5.40 × 10¹ | NS | – | – | – | – |
| 125 | Female | Stray | 3.30 × 10¹ | NS | NS | – | – | – |
| 127 | Female | Stray | 4.40 × 10² | – | – | – | – | – |
| 129 | Male | Stray | 1.00 × 10² | NS | – | Bh | – | – |
| 138 | Male | Stray | 1.40 × 10¹ | – | – | – | – | – |
| 139 | Female | Stray | 9.50 × 10¹ | NS | – | – | – | – |
| 143 | Female | Stray | 2.40 × 10¹ | – | – | – | – | – |
| 144 | Female | Stray | 1.10 × 10¹ | – | – | – | – | – |
| 146 | Male | Stray | 1.80 × 10² | – | – | – | – | – |
ITS = intergenic transcriber spacer; NS = positive sample in cPCR but showing weak bands, which precluded sequencing; Bc = Bartonella clarridgeiae; Bh = Bartonella henselae
We verified positive and significant correlations (P <0.001) between the absolute quantification of nuoG Bartonella DNA (L10) and positive results in cPCR for ITS (0.567), pap31 (0.291), gltA (0.282) and rpoB (0.289), two or more genes simultaneously (0.414), and at least one gene (0.570) (Table 3). Logistic regression results revealed a significant association between the absolute quantification of nuoG Bartonella DNA and and positive results in cPCR (P <0.001). An increase of one unit in L10 was associated with increases of about 10-, seven- and 11-fold in the odds for a positive result in cPCR for at least one gene, two or more genes, and ITS, respectively (Table 4).
Table 3.
Point biserial correlation between absolute quantification of nuoG Bartonella DNA L10* and positive results in conventional PCR
| Situation | Estimated coefficient | P value |
|---|---|---|
| ITS | 0.56722 | <0.00001 |
| Pap31 | 0.29100 | 0.00014 |
| gltA | 0.28223 | 0.00021 |
| rpoB | 0.28919 | 0.00015 |
| Two or more genes | 0.41496 | <0.00001 |
| At least one gene | 0.57082 | <0.00001 |
Log10 (NuoG + 1)
ITS = intergenic transcribed spacer
Table 4.
Estimated odds ratio (OR) obtained from the analysis of the real-time and and conventional polymerase chain reaction results, considering L10* as the explanatory variable and the conventional PCR result as the response variable (for cases in which point biserial correlation was >0.4)
| At least one gene vs L10 | |||||
| Parameter | Estimate | SE | z-value | Pr(>|z|) | OR |
| Intercept | –5.585 | 1.172 | –4.764 | <0.001 | 0.004 |
| L10 | 2.332 | 0.609 | 3.827 | <0.001 | 10.296 |
| Two or more genes vs L10 | |||||
| Parameter | Estimate | SE | z-value | Pr(>|z|) | OR |
| Intercept | –6.917 | 1.708 | –4.049 | <0.001 | 0.001 |
| L10 | 2.052 | 0.672 | 3.052 | 0.002 | 7.782 |
| ITS vs L10 | |||||
| Parameter | Estimate | SE | z-value | Pr(>|z|) | OR |
| Intercept | –5.982 | 1.335 | –4.482 | <0.001 | 0.003 |
| L10 | 2.46 | 0.679 | 3.621 | <0.001 | 11.702 |
Log10 (NuoG + 1). SE = standard error; Pr = positive result
Discussion
In this study we constructed a qPCR assay for the detection and quantification of Bartonella species. We chose to develop a broad-range qPCR for Bartonella species as this test may be used for screening in clinical and epidemiologic studies. Vector-borne diseases show a clinical presentation similar to many other infections, and quick treatment is sometimes crucial to a favorable outcome. The screening of invertebrates as vectors, and vertebrates as hosts or reservoirs for vector-borne agents in a genus-specific molecular assay may be very useful.
Although Bartonella species culture is considered ideal, the difficult and time-consuming nature of it make it impractical for large-scale use. 22 Molecular tests based on real-time Bartonella species diagnosis have been developed.23–29 Herein, several primer sets (based on nuoG, gltA and 16SrRNA) were used to amplify reference DNAs from 11 Bartonella species, chosen for their distant phylogenetic relationships (data not shown). nuoG showed specificity and sensitivity as a target gene for use in the detection of Bartonella species. Recently, the superior performance of nuoG over ftsZ and gltA as a target in a qPCR was reported. 22 While the discrimination of Bartonella species was not the goal of the present study, previous studies have done it using a high resolution melt amplification system or by analyzing mutations in an internal region of gltA using direct hybridization of LCRed and fluorescein-labeled probes coupled with melting curve analysis with the use of fluorescence resonance energy transfer technology.24,25 Recently, a SYBR green qPCR was used to detect B henselae and B clarridgeiae among cats sampled in animal shelters in Rio Grande do Sul state, Brazil, inferring a higher sensitivity of this method than cPCR. 26
A broad-range Bartonella PCR has the advantage of allowing the detection of different species, which is particularly important when epidemiologic studies are conducted. Our assay may be used in screening of populations in clinical and epidemiologic surveys, saving time and costs. It detected few DNA copies of Bartonella species, distinguished them from other closely related agents (Ehrlichia species, Anaplasma species, Neorickettsia species and Rickettsia species) and was conducted in <3 h. Validation of the test should be undertaken with the collection of clinical samples (from humans, domestic animals, livestock and wild animals) and vector-suspected arthropods (ticks, flies, fleas, mites, etc) followed by testing with other methods (serology, culture, conventional PCR and high-resolution melting analysis). Besides, qPCR may be useful in the evaluation of the efficacy of treatment protocols for Bartonella species that infect animals and humans.
The new TaqMan assay developed herein showed high sensitivity and specificity, detecting as few as 10 copies of plasmid/reaction and showed no cross-reaction with the DNA of other pathogens analyzed. Although both qPCR and cPCR were able to detect Bartonella species DNA in the cats’ blood samples, the former showed a higher number of positive samples than the latter. Even though the goal of the present study was not compare qPCR and cPCR, our results suggest that this new assay could be useful when assessing samples showing low bacteremia. Logistic regression analysis showed that a 10-fold increase in copy numbers assessed by qPCR was associated with the odds of positive results in cPCR, mainly for ITS, demonstrating the better performance of qPCR over cPCR in detecting low Bartonella DNA copy numbers. The lack of gold-standard techniques for Bartonella species diagnosis along with the fact that the blood samples from the cats were not submitted for culturing precluded the inference of analytical sensitivity of the proposed qPCR described here. Moreover, quantitative data may be used in epidemiologic studies aiming to determine a possible correlation between the number of copies and bacterial factors (species/genotypes more virulent), host factors (age, genetic susceptibility, development of immunity), environmental factors (high or low exposure levels to arthropod vectors) and response to antibiotic therapy.
Although qPCR assays to detect Bartonella species have been recently developed by other researchers, some those studies simultaneously detect other pathogens (Mycobacteria species, Francisella tularensis and Tropheryma whipplei) in addition of Bartonella species, 27 or, when specific to Bartonella, detect few and only some specific species, such those for both B henselae and B clarridgeiae, 26 or those specific for B quintana and B henselae, 29 or only for B vinsonii subspecies berkhoffii. 28 Therefore, the development of new and broad-range qPCR detection and quantification of Bartonella species is much needed.
Although nuoG has been previously used as a target in a qPCR for Bartonella species, 22 in this study we proposed a new TaqMan-based assay, additionally showing parameters for validation. Besides, parameters of linear standard curve (R²), amplification efficiency, limit of detection and consistency across replicate reactions in intra- (repeatability) and inter-assays (reproducibility) are indispensable to the validation of the new qPCR protocol. In our assay, results obtained through SD and coefficients of variation showed highly accurate and reproducible results. Moreover, this new protocol allows absolute quantification when compared with the previously described protocol. 22
Cats are the main reservoirs for B henselae, B clarridgeiae and B koehlerae. 30 Bartonella henselae is incriminated as the causative agent of cat scratch disease, ocular manifestations, encephalopathy, osteomyelitis and hepatic peliosis in humans. 31 In Rio de Janeiro, HIV-infected individuals and breeding cats are at higher risk of Bartonella species infection. 32 According to the present results, cats, through scratches contaminated by flea feces or bites contaminated with infected blood of cats, in the city of Campo Grande may act as a source of infection of B henselae or B clarridgeiae for humans.3,4,30
Conclusions
The present work presents a novel quantitative 5' nuclease qPCR assay to diagnose infections caused by Bartonella species. Future studies aiming to validate this new protocol for molecular diagnosis of Bartonella species in blood and tissue samples from human beings and animals presenting clinical signs suggestive of bartonellosis, arthropods (ticks, fleas and mosquitoes) and wildlife reservoirs are much needed for clinical and epidemiological studies.
Acknowledgments
We thank Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) for the financial support (number #2013/09889-0), and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for MR André’s sandwich-scholarship #1483-10-2). We are also thankful to Dr Bruno Chomel (University of California, Davis, CA, USA), Dr Edward Breitschwerdt (North Carolina State University, Raleigh, NC, USA) and Dr Dennis John Grab (Johns Hopkins School of Medicine, Baltimore, MD, USA). We would like to thank Dr Haroldo Henrique de Rezende Neves (Faculdade de Ciências Agrárias e Veterinárias – FCAV/UNESP Jaboticabal) for assistance with the statistical analysis.
Footnotes
The authors do not have any potential conflicts of interest to declare.
Funding: This work was supported by funds from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (number #2013/09889-0) and NIAID HPTN-Network Laboratory U01 AI068613 (to JSD; PI S. Eshleman).
Accepted: 8 June 2015
References
- 1. Doudier B, Olano J, Parola P, et al. Factors contributing to emergence of Ehrlichia and Anaplasma spp. as human pathogens. Vet Parasitol 2010; 167: 149–154. [DOI] [PubMed] [Google Scholar]
- 2. Diaz MH, Bai Y, Malania L, et al. Development of a novel genus-specific real-time PCR assay for detection and differentiation of Bartonella species and genotypes. J Clin Microbiol 2012; 50: 1645–1649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Breitschwerdt EB, Kordick DL. Bartonella infection in animals: carriership, reservoir potential, pathogenicity, and zoonotic potential for human infection. Clin Microbiol Rev 2000; 13: 428–438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Chomel BB, Kasten RW. Bartonellosis, an increasingly recognized zoonosis. J Appl Microbiol 2010; 109: 743–750. [DOI] [PubMed] [Google Scholar]
- 5. Buffet J-P, Kosoy YM, Vayssier-Taussat M. Natural history of Bartonella-infecting rodents in light of new knowledge on genomics, diversity and evolution. Future Microbiol 2013; 8: 1117–1128. [DOI] [PubMed] [Google Scholar]
- 6. Staggemeier R, Venker CA, Klein DH, et al. Prevalence of Bartonella henselae and Bartonella clarridgeiae in cats in the south of Brazil: a molecular study. Mem Inst Oswaldo Cruz 2010; 105: 873–878. [DOI] [PubMed] [Google Scholar]
- 7. Braga MSCO, Diniz PPVP, André MR, et al. Molecular characterisation of Bartonella species in cats from São Luís, state of Maranhão, north-eastern Brazil. Mem Inst Oswaldo Cruz 2012; 107: 772–777. [DOI] [PubMed] [Google Scholar]
- 8. Miceli NG, Gavioli FA, Gonçalves LR, et al. Molecular detection of feline arthropod-borne pathogens in cats in Cuiabá, state of Mato Grosso, central-western region of Brazil. Rev Bras Parasitol Vet 2013; 22: 385–390. [DOI] [PubMed] [Google Scholar]
- 9. De Bortoli CP, André MR, Seki MC, et al. Detection of hemoplasma and Bartonella species and co-infection with retroviruses in cats subjected to a spaying/neutering program in Jaboticabal, SP, Brazil. Rev Bras Parasitol Vet 2012; 21: 219–223. [DOI] [PubMed] [Google Scholar]
- 10. André MR, Denardi NCB, Sousa KCM, et al. Arthropod-borne pathogens circulating in free-roaming domestic cats in a zoo environment in Brazil. Ticks Tick Borne Dis 2014; 5: 545–551. [DOI] [PubMed] [Google Scholar]
- 11. Santis AC, Herrera HM, Sousa KCM, et al. Molecular detection of hemotrophic mycoplasmas among domiciled and free-roaming cats in Campo Grande, state of Mato Grosso do Sul, Brazil. Rev Bras Parasitol Vet 2014; 23: 231–236. [DOI] [PubMed] [Google Scholar]
- 12. Helps C, Reeves N, Egan K, et al. Detection of Chlamydophila felis and feline herpesvirus by multiplex real-time PCR analysis. J Clin Microbiol 2003; 41: 2734–2736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Bustin SA, Benes V, Garson JA, et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem 2009; 55: 611–622. [DOI] [PubMed] [Google Scholar]
- 14. Maggi RG, Breitschwerdt EB. Potential limitations of the 16S-23S rRNA intergenic region for molecular detection of Bartonella species. J Clin Microbiol 2005; 43: 1171–1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Diniz PPVP, Maggi RG, Schwartz DS, et al. Canine bartonellosis: serological and molecular prevalence in Brazil and evidence of co-infection with Bartonella henselae and Bartonella vinsonii subsp. berkhoffii. Vet Res 2007; 38: 697–710. [DOI] [PubMed] [Google Scholar]
- 16. Johnson G, Ayers M, McClure SCC, et al. Detection and identification of Bartonella species pathogenic for humans by PCR amplification targeting the riboflavin synthase gene (ribC). J Clin Microbiol 2003; 41: 1069–1072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Norman AF, Regnery R, Jamenson NP, et al. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J Clin Microbiol 1995; 33: 1797–1803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Winoto IL, Goethert H, Ibrahim IN, et al. Bartonella species in rodents and shrews in the greater Jakarta area. Southeast Asian J Trop Med Public Health 2005; 36: 1523–1529. [PubMed] [Google Scholar]
- 19. Maggi RG, Breitschwerdt EB. Isolation of bacteriophages from Bartonella vinsonii subsp. berkhoffii and the characterization of Pap31 gene sequences from bacterial and phage DNA. J Mol Microbiol Biotechnol 2005; 9: 44–51. [DOI] [PubMed] [Google Scholar]
- 20. Altschul SF, Gish W, Miller W, et al. Basic local alignment search tool. J Mol Biol 1990; 215: 403–410. [DOI] [PubMed] [Google Scholar]
- 21. R Core Team. R: A language and environment for statistical computing. http://www.R-project.org/ (accessed May 18, 2015).
- 22. Colborn JM, Kosoy MY, Motin VL, et al. Improved detection of Bartonella DNA in mammalian hosts and arthropod vectors by real-time PCR using the NADH dehydrogenase gamma subunit (nuoG). J Clin Microbiol 2010; 48: 4630–4633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Molia S, Chomel BB, Kasten RW, et al. Prevalence of Bartonella infection in wild African lions (Panthera leo) and cheetahs (Acinonyx jubatus). Vet Microbiol 2004; 100: 31–41. [DOI] [PubMed] [Google Scholar]
- 24. Morick D, Baneth G, Avidor B, et al. Detection of Bartonella spp. in wild rodents in Israel using HRM real-time PCR. Vet Microbiol 2009; 139: 293–297. [DOI] [PubMed] [Google Scholar]
- 25. Ciervo A, Ciceroni L. Rapid detection and differentiation of Bartonella spp. by a single-run real-time PCR. Mol Cell Probes 2004; 18: 307–312. [DOI] [PubMed] [Google Scholar]
- 26. Staggemeier R, Pilger DA, Spilki FR, et al. Multiplex SYBR® green-real time PCR (qPCR) assay for the detection and differentiation of Bartonella henselae and Bartonella clarridgeiae in cats. Rev Inst Med Trop Sao Paulo 2014; 56: 93–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Angelakis E, Roux V, Raoult D, et al. JM. Real-time PCR strategy and detection of bacterial agents of lymphadenitis. Eur J Clin Microbiol Infect Dis 2009; 28: 1363–1368. [DOI] [PubMed] [Google Scholar]
- 28. Li D, Song X, Wang J, et al. Real-Time PCR-based detection of Bartonella vinsonii subsp. berkhoffii by TaqMan minor groove binder probe. Wei Sheng Wu Xue Bao 2013; 53: 976–983. [PubMed] [Google Scholar]
- 29. Liberto MC, Lamberti AG, Marascio N, et al., Molecular identification of Bartonella quintana infection using species-specific real-time PCR targeting transcriptional regulatory protein (bqtR) gene. Mol Cell Probes 2011; 25: 238–242. [DOI] [PubMed] [Google Scholar]
- 30. Breitschwerdt EB, Maggi RG, Chomel BB, et al. Bartonellosis: an emerging infectious disease of zoonotic importance to animals and human beings. J Vet Emerg Crit Care 2010; 20: 8–30. [DOI] [PubMed] [Google Scholar]
- 31. Lamas C, Curi A, Bóia M, et al. Human bartonellosis: seroepidemiological and clinical features with an emphasis on data from Brazil – a review. Mem Inst Oswaldo Cruz 2008; 103: 221–235. [DOI] [PubMed] [Google Scholar]
- 32. Lamas CC, Mares-Guia MA, Rozental T, et al. Bartonella spp. infection in HIV positive individuals, their pets and ectoparasites in Rio de Janeiro, Brazil: serological and molecular study. Acta Trop 2010; 115: 137–141. [DOI] [PubMed] [Google Scholar]