

SUMMARY
CD226 plays a vital role in natural killer (NK) cell cytotoxicity, interacting with its ligands CD112 and CD155 to initiate immune synapse formation, primarily through leukocyte function-associated-1 (LFA-1). Our study examined the role of CD226 in NK cell surveillance of acute myeloid leukemia (AML). NK cells in patients with AML had lower expression of CD226. CRISPR-Cas9 deletion of CD226 led to reduced LFA-1 recruitment, poor synapse formation, and decreased NK cell anti-leukemic activity. Engineering NK cells to express a chimeric antigen receptor targeting the AML antigen CD38 (CAR38) could overcome the need for CD226 to establish strong immune synapses. LFA-1 blockade reduced CAR38 NK cell activity, and this depended on the CD38 expression levels of AML cells. This suggests parallel but potentially cooperative roles for LFA-1 and CAR38 in synapse formation. Our findings suggest that CAR38 NK cells could be an effective therapeutic strategy to overcome CD226-mediated immune evasion in AML.
In brief
Melo Garcia et al. demonstrate that CD38-directed CAR NK cells can overcome acute myeloid leukemia (AML)-induced immune evasion by compensating for CD226 downregulation. They show how synthetic CARs can synergize with and substitute for natural NK cell receptors to sustain cytotoxicity against AML cells.
Graphical Abstract

INTRODUCTION
Acute myeloid leukemia (AML) is the most common acute leukemia in adults. Despite recent new therapies, AML-related death rates have remained unchanged for the past 40 years.1 Moreover, patients with refractory/relapsed disease rarely achieve remission,2 highlighting the need for alternative treatments such as adoptive therapy with natural killer (NK) cells.
NK cells are members of the innate lymphoid family and participate in the immunosurveillance of cancers, including AML.3 Their cytotoxicity against tumor targets depends on the balance between activating and inhibitory inputs from their surface receptors. Activating receptors invoke NK cell cytotoxicity against tumor cells to eradicate them, while inhibitory receptors oppose this anti-tumor action.4 A variety of genetic manipulation approaches are being pursued to decrease inhibitory and enhance activating receptor signaling to increase NK cell potency for cancer immunotherapy.5
CD226 (DNAX accessory molecule-1 [DNAM-1]) is an important activating receptor on NK cells.6,7 It interacts with its ligands CD155 and CD112 on tumor targets to increase NK cell effector function, leading to NK cell anti-tumor activity.8,9 CD226 also facilitates immune synapse formation between NK and tumor cells10 by promoting inside-out signaling of the leukocyte function-associated-1 (LFA-1) receptor,11 critical for NK cell binding to targets and tumor cell killing.12 CD226 expression on NK cells is reduced in myeloid malignancies,13–15 suggesting that CD226 downregulation may contribute to the evasion of AML cells from NK cell immune surveillance.
Chimeric antigen receptors (CARs) are synthetic receptors introduced into NK cells to enhance their anti-tumor activity.16 NK cells equipped with CARs represent a promising approach for the treatment of AML.17–20 This technology integrates the directed cytotoxicity derived from the CAR engagement and its target with the innate activity of NK cells against AML through their endogenous receptors. Yet, the dynamic interplay between CARs and native NK cell receptors remains under-researched. Understanding how CARs interact and cooperate with NK cell innate receptors, such as CD226 and LFA-1, might shed light on ways to overcome AML resistance and improve NK cell-based therapy.
Here, we investigated how AML cells evade immune surveillance by downregulating CD226 on NK cells. To delineate the interaction between the synthetic CAR and the natural receptors CD226 and LFA-1, we engineered NK cells to express a CAR against CD38 while deleting CD226 by CRISPR-Cas9 knockout (KO). We showed that CAR38-modified NK cells can bypass the need for CD226 activation, operating with less dependence on LFA-1, and thus may overcome the immune escape mechanism of AML linked to CD226 downregulation.
RESULTS
CD226 ligands are differentially expressed in AML subpopulations and correlate with clinically relevant genomic subgroups
The anti-tumor cytotoxicity of NK cells depends on the interaction between their surface receptors and cognate ligands on tumor targets.5 To examine the expression of NK cell ligands on leukemic blasts, we analyzed RNA sequencing data from three publicly available cohorts for patients with AML (TCGA, BEAT-AML, and Leucegene). Our analysis focused on the expression of 19 NK cell ligands in AML cells (as described in the STAR Methods), which are associated with over 20 key NK cell receptors, and we identified three distinct NK ligand (NKL) clusters. Each of the three cohorts was evenly represented across the clusters (Figures 1A and 1B). The principal difference between these clusters was the differential expression of ligands from the HLA and CD226/TIGIT families. NKL cluster A was enriched for classical and non-classical HLA genes and the CD226 ligand genes NECTIN2 (CD112) and PVR (CD155) (Figures 1C–1E). This cluster was also associated with a higher occurrence of complex karyotypes and mixed lineage leukemia (MLL) fusion gene anomalies (Figure 1F). NKL cluster B, with intermediate HLA gene and reduced CD226 ligand gene expression levels (Figures 1C–1E), was enriched for RUNX1 and CBFB genomic subgroups (Figure 1F). Finally, NKL cluster C was characterized by high PVR, intermediate NECTIN2, and low levels of classical and non-classical HLA genes (Figures 1C–1E) and was associated with normal cytogenetics and AML genomic subgroups with NPM1 or FLT3-ITD mutations (Figure 1F). The expression of genes for natural cytotoxicity receptors and NKG2D ligands did not vary markedly across different leukemic cells within the NKL clusters (Figure 1C). Although this exploratory analysis does not allow for definitive conclusions about the biological significance of these ligands, their distinct expression patterns may serve as clinically relevant biomarkers to help identify AML subgroups with greater susceptibility to NK cell cytotoxicity. Overall, these findings suggest that leukemic cells present a specific pattern of surface ligands to NK cells, predominantly consisting of HLA family members and CD226 ligands.
Figure 1. AML cells differentially express CD226 ligands.

(A) Strategy used for the single-cell RNA sequencing (scRNA-seq) analysis.
(B) Distribution plots showing the spatial cluster organization (top left), cluster allocations according to their hierarchy subtype (top right), and their cohort of origin (bottom).
(C) Heatmap showing the relative expression of NK cell ligands across the clusters.
(D and E) Distribution plots displaying the allocation of CD226 ligands (D) and HLA-A/B/C/E/F/G (E) across NKL clusters and boxplots quantitatively comparing their expression among the clusters.
(F) Distribution of AML genomic subgroups across NKL clusters.
(G) Violin plots comparing the levels of CD226 ligands between leukemic stem cells (LSC+) and leukemic non-stem cells (LSC−) by scRNA-seq (n = 110 samples) and microarray (n = 227 samples). LSCs were categorized based on their transcriptomic profile (STAR Methods and Zeng et al. 202221).
(H) Histograms showing the expression of CD155 and CD112 across genomic subgroups. The histograms display the genomic subgroups in the analysis of the AML cohort combined from BEAT-AML (n = 173), TCGA (n = 281), and Leucegene (n = 410). Each vertical line in the histograms represents an individual patient in this cohort.
(I) Boxplots comparing the expression of CD226 ligands by bulk RNA-seq in 44 patients with AML at diagnosis (Dx) and relapse (Rel).
Statistics: Wilcoxon rank-sum text (D–G and I).
Since CD226 is an essential NK cell-activating receptor22 and its ligands are differentially expressed in AML blasts, we focused our analysis on the patterns of CD226 ligand expression in AML. We observed higher expression of NECTIN2 and PVR in leukemic stem cells compared to their leukemic non-stem cell counterparts (Figure 1G). This was not the case with other NK cell ligands (data not shown). Most genomic subgroups expressed similar levels of both ligands, with the inv(16) subgroup displaying lower PVR and moderate NECTIN2 levels (Figure 1H). When comparing AML cells to normal hematopoietic stem cells, we found that AML cells were enriched for CD226 ligands (Figure S1). Furthermore, we observed no significant differences in their expression with disease progression (Figure 1I). Collectively, our data suggest that the expression of CD226 ligands, although varied among leukemic cells, is stable through different stages and across genomic AML subgroups.
CD155-induced CD226 downregulation modulates NK cell function in AML
Next, we used mass cytometry to characterize the expression levels of different receptors and markers of effector function in NK cells from AML samples. When compared to NK cells from healthy controls, AML NK cells had significantly lower CD226 levels (Figures 2A, S2A, and S2B) without a corresponding increase in TIGIT (an inhibitory receptor that also binds CD155 and CD11223; Figures S2C and S2D). We then examined the association between CD226 expression and NK cell cytotoxicity potential. NK cells from patients with AML were divided into CD226-high (CD226high) and CD226-negative/low (CD226−/low) groups (Figure S2E). The CD226−/low subpopulation was characterized by a marked reduction in critical activating receptors and cytotoxic granules compared to the CD226high population. Despite this, both groups displayed similar levels of TIGIT and other NK cell inhibitory receptors like NKG2A and KLRG1 (Figures 2B and S2F).
Figure 2. CD226 downregulation on NK cells is dependent on CD155 expression in AML cells.

(A) Bar graphs showing percentage of CD226+ and CD226 MMI in NK cells from healthy controls (HCs) (n = 4) and patients with AML (n = 17).
(B) Dot plots comparing the percentage of positive cells for each NK cell marker gated on CD56+CD226high and CD56+CD226low/− subpopulations (n = 17 AML samples).
(C) UMAP plots showing NK cell clusters based on their expression of NK cell markers by mass cytometry (18,850 NK cells from 13 AML samples). The percentage of NK cells per cluster is shown.
(D) Heatmap displaying the expression level of NK cell markers across NK cell clusters.
(E) Representative histogram showing the CD226 levels in clusters 1 and 2.
(F and G) Violin plots comparing single-cell-level expression of NK cell markers between clusters 1 (n = 4,564 cells) and 2 (n = 4,492 cells).
(H) Percentage of CD226+ NK cells after 24 h coculture with AML cells by flow cytometry. Baseline (gray dots): NK cells not exposed to AML cells (n = 10 donors; three independent experiments).
(I) Fold reduction in CD226 expression in NK cells after 96 h coculture with AML cell lines by flow cytometry following rechallenge with tumor cells. Each symbol represents one cord blood donor (n = 3).
(J) Representative images displaying the CD226 expression on the cell surface (blue) and the internalized CD226 (red). Yellow numbers denote median area of internalized CD226. Violin plots showing median area of internalized CD226 after coculture with AML cells by imaging flow cytometry. For gating strategy, see Figures S2I and S2J. Each circle represents one NK cell. NK cells alone (n = 324); NK cells in coculture (n = 482).
(K) Dot plots depicting percentage of CD226+ NK cells and the fold change in MFI relative to the baseline after coculture with U937 (n = 11 donors; four independent experiments).
Statistics: t test with Welch’s correction (A and B), two-tailed Student’s t test (F–H), two-way ANOVA (I), Wilcoxon test (J), and one-way ANOVA (K). **p < 0.01, ***p < 0.001, and ****p < 0.0001. Mean ± SD (A); means (B); mean ± SEM (K). MMI, median metal intensity; MFI, median fluorescence intensity; E:T ratio, effector-to-target ratio.
To allow for their unsupervised study, we concatenated the NK cells from individuals with AML and explored their immunophenotype based on uniform manifold approximation and projection (UMAP) bidimensional clustering. We observed a total of eight clusters expressing different levels of NK cell markers (Figures 2C and 2D). Overall, the clusters expressing higher levels of CD226 (such as clusters 1, 4, 6, and 7) also tended to have higher levels of NK cell-activating receptors, cytotoxic granules, and maturation makers compared to those expressing lower CD226 levels (clusters 2, 5, and 8). Among these, the two largest clusters, cluster 1 (26.7%) and cluster 2 (26.4%), expressed distinct levels of CD226 (Figure 2E), with cluster 1 showing significantly higher CD226 expression than cluster 2 (Figure 2F). Furthermore, cluster 1 expressed higher levels of activating receptors, cytotoxic granules, and maturation markers, with lower levels of the inhibitory receptor NKG2A (Figure 2G).
Next, we investigated whether AML cells regulate CD226 expression on NK cells. Coculture experiments with a variety of AML cell lines resulted in a significant decrease in CD226 expression on NK cells within 24 h, except for the THP-1 cell line (Figure 2H). Additionally, multiple challenges with AML cells resulted in a significant reduction in CD226 levels (Figures 2I and S2G), suggesting the ability of AML cells to modulate CD226 expression on NK cells.
To determine the mechanisms by which AML cells induce the downregulation of CD226 in NK cells, we first verified if direct cell-cell contact was necessary between NK cells and AML cells. Transwell experiments indicated that in the absence of direct contact, CD226 levels on NK cells remained unchanged (Figure S2H). Furthermore, NK cells cultured in the presence of AML cells displayed a significant increase in intracellular CD226 expression compared to NK cells cultured alone (median area of internalized CD226: 4.09 vs. 0.89, respectively; p < 0.001) (Figures 2J, S2I, and S2J), suggesting receptor internalization as a potential mechanism for CD226 downregulation.
Interestingly, THP-1 cells did not alter CD226 levels on NK cells. This cell line had lower CD155 and higher CD112 levels compared to the other cell lines (Figure S2K), suggesting a differential impact on CD226 modulation by these ligands. To identify which CD226 ligand might be responsible for the observed CD226 downregulation, we engineered AML cells with deletions of PVR or NECTIN2. Deletion of one gene did not affect the expression of the other (Figure S2L). Expression of CD226 on NK cells decreased significantly following coculture with NECTIN2 KO AML cells but remained stable with PVR KO AML cells, indicating that CD155 is the critical mediator of CD226 downregulation in NK cells (Figure 2K). Together, these data suggest that AML cells can evade immune surveillance by inducing CD226 downregulation in NK cells, primarily through interactions with CD155.
Lower CD226 levels lead to impaired NK cell function against AML cells due to inadequate immune synapse formation
To investigate the contribution of CD226 to NK cell cytotoxicity against AML cells, we deleted CD226 in NK cells, achieving a KO efficiency of 75%–98% (Figures 3A and 3B). CD226 KO showed no significant effect on the expression of TIGIT, CD96 (an inhibitory receptor that also binds to CD155 and CD112), LFA-1, or 2B4 (a costimulatory receptor that cooperates with CD226) (Figure 3C). Additionally, CD226 KO did not alter the levels of activating and inhibitory receptors or cytotoxic granules or the proliferative capacity of NK cells (Figure S3A).
Figure 3. CD226 and LFA-1 are essential for cytotoxicity and immunological synapse formation against AML cells.

(A) Dot plot showing the percentage of CD226+ NK cells (n = 17 donors).
(B) Representative histograms depicting CD226 levels.
(C) t-distributed stochastic neighbor embedding (tSNE)-cuda maps showing the distribution of the NK cell receptors in CD226 KO (n = 75,454 NK cells) and CTRL (Cas9 control) groups (n = 73,832 NK cells) (n = 3 donors). Scales represent the expression of NK cell receptors. Violin plots showing MMI of the specific receptors.
(D–F) Flow cytometric quantification of CD107a (D), interferon (IFN)-γ (E), and tumor necrosis factor alpha (TNF-α) (F) expression by CD226 KO and CTRL NK cells in coculture with MOLM14 cells (n = 7–10 donors; three independent experiments). Representative zebra plots are shown.
(G) Live-image assay showing the killing of CD226 KO and CTRL NK cells against MOLM14 cells (E:T ratio 1:4; n = 9 donors; four independent experiments). Areas under the curve are presented in the bar graph.
(H) Percentage and fold change in tagged synapse number for CD226 KO and CTRL groups by imaging flow cytometry. Each colored symbol represents one donor (n = 5; three independent experiments).
(I and J) Representative images of F-actin (I) and open LFA-1 (J) within immunological synapses. Bright-field images display the threshold mask (red arrow) used to measure F-actin and open LFA-1 within the synapses. Dot plots show F-actin and open LFA-1 median area and MFI within tagged immunological synapses in CD226 KO (n = 418 synapses) and CTRL (n = 1136 synapses) groups. The same experiments were used as in (H). Symbols represent selected images. For the gating strategy, see Figure S3B and S3C.
(K) Percentage of open LFA-1 within the synapse in CD226 KO and CTRL. The same experiments were used as in (H).
Statistics: one-way ANOVA with Dunnett’s correction for multiple comparisons (A), two-tailed Student’s t test (C–H), two-way ANOVA with Bonferroni’s correction for multiple comparisons (G), and Mann-Whitney test (I–K). Mean ± SD (A and H), mean ± SEM (G), and median with 95% confidence interval (CI) (I–K). *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001; ns, not significant.
Notably, CD226 KO NK cells had significantly lower degranulation (Figure 3D) and effector cytokine production (Figures 3E and 3F) compared to Cas9 control (CTRL) NK cells when in coculture with AML cells. We also observed a partial but significant reduction in NK cell-mediated cytotoxicity (Figure 3G), highlighting the importance of CD226 in AML targeting. Despite the low levels of CD155 on THP-1 cells, CD226-related NK cell cytotoxicity remained unaffected. This suggests that the higher CD112 expression on THP-1 cells (Figure S2K) was likely sufficient to engage CD226 and activate NK cells, compensating for the reduced CD155 levels.
The structural integrity of the immune synapse is critical to NK cell function.24 Given the important role of CD226 in activating LFA-1, an integrin critical for immune synapse formation,25 a reduction or loss of CD226 expression in NK cells could impair immune synapse formation with AML cells. To test this, we co-cultured CD226 KO or CTRL NK cells with MOLM14 AML cells and evaluated their ability to form effective immune synapses. The gating strategy to identify conjugate and tagged synapses is shown in Figures S3B and S3E.
As expected, CD226 levels were significantly lower in the CD226 KO NK-AML cell conjugates compared to CTRL NK cells (Figure S3F). CD226 KO NK cells were less capable of forming proper/tagged immune synapses compared to control cells (Figure 3H). Finally, we quantified the accumulation of F-actin inside the synapse, a surrogate marker of immune synapse quality or strength.26 CD226 KO NK cells displayed impaired F-actin accumulation, with CD226 KO cells showing a 10%–20% reduction in the levels of F-actin within the synapse (p = 0.034) (Figure 3I), suggesting that CD226 enhances immune synapse quality.
CD226 activates the integrin LFA-1 to its open conformation, which results in F-actin accumulation at the synapse.27,28 Therefore, CD226 depletion could impact LFA-1 activation and recruitment to the synapse.29 Open LFA-1 was more abundantly found in NK cell-AML conjugates than in unconjugated singlet NK cells (Figure S3G). In contrast, CD226 KO NK cells had significantly lower levels of open LFA-1 (up to 45% less) at the immune synapses and were less efficient in directing LFA-1 into the synapse compared to wild-type (WT) NK cells (Figures 3J and 3K). These data suggest that CD226 may play an important role in NK cell activity against leukemia, in part by priming LFA-1 to form robust immune synapses.
NK cells expressing a CD38-targeting CAR can overcome the effects of CD226 deficiency in AML
Next, we investigated whether introducing a CAR into NK cells to direct their activity toward target cells could compensate for the loss of CD226. We thus developed a CAR construct targeting CD38, an antigen expressed in AML (Figures S4A–S4C). The CAR construct incorporated the inducible caspase-9 suicide gene, the CD38-targeting scFv extracellular portion, CD28 costimulatory and CD3ζ signaling domains, and interleukin (IL)-15 (hereafter CAR38.3ζ.IL-15; Figure S4D depicts the CAR design). The transduction efficiency of CAR38.3ζ.IL-15 in NK cells is shown in Figure 4A.
Figure 4. CAR38 NK cells can mediate effective cytotoxicity against CD38+ AML targets in the absence of CD226.

(A) Bar graph showing the CAR38 transduction efficiency (donors = 27). Representative flow cytometry plot showing CAR38 expression in NT and CAR38.3ζ.IL-15 NK cells.
(B–D) Flow cytometric quantification of CD107a (B), IFN-γ (C), and TNF-α (D) expression by NT and CAR38 NK cells in coculture with primary AML cells. Each dot represents a combination of an NK cell donor with a CD38+ AML sample (n = 8; two independent experiments).
(E) Fold change in Annexin V in NK-AML cocultures. Each dot represents a combination of NK donor:CD38+ AML sample (n = 11; two independent experiments). The patient and disease characteristics are detailed in Table S1.
(F) Contour plot showing the expression of CD226 and CAR38 in NK cell groups.
(G–I) Flow cytometric quantification of CD107a (G), IFN-γ (H), and TNF-α (I) expression comparing NT ± CD226 KO and CAR38 NK cells ± CD226 KO following coculture with THP-1 and MOLM14 cells. Each dot represents an NK cell donor (n = 6 donors; three independent experiments).
(J) Live-image cytotoxicity assay comparing the percentage of killing between CAR38 NK cells ± CD226 KO against MOLM14 (E:T ratio 1:4; n = 4; two independent experiments).
(K) Number of tumor cells per well (red count per image) over 16 days performed by real-time imaging analysis (n = 3 donors; three independent experiments). Rechallenges with 50,000 AML cells were performed every 48–72 h (arrows).
(L) Representative images showing MOLM14 during the rechallenge assay cocultured with CTRL NT, CD226 KO NT, CTRL CAR38, and CD226 KO CAR38 NK cells.
(M) Bioluminescence imaging (BLI) at days 0 and 21 was used to monitor the growth of firefly luciferase (FFluc)-labeled MOLM14 in NSG mice infused with NT, CTRL CAR38.3ζ.IL15, and CD226 KO CAR38.3ζ.IL15 NK cells (n = 5 mice per group). Mice engrafted with MOLM14 only were used as a control.
(N) Individual BLI data for the four groups of mice shown in (M).
(O) Kaplan-Meyer curves showing the percentage of survival of NSG mice from (M).
Statistics: two-tailed Student’s t test (A–E and G–I), two-way ANOVA with Bonferroni’s correction for multiple comparisons (J and K), and log rank (Mantle-Cox) test (O). Mean ± SD (A, E, J, and K). *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
In coculture with CD38-expressing primary AML cells, CAR38+ NK cells showed a significant increase in degranulation (Figure 4B) and cytokine production (Figures 4C and 4D) compared to non-transduced (NT) NK cells. CAR38 NK cells also induced significantly higher levels of apoptosis in CD38-expressing primary AML cells than NT NK cells (difference between means = 0.69 ± 0.13; p < 0.0001) (Figures 4E and S4E), confirming their direct anti-leukemic cytotoxicity. Furthermore, we observed a significantly lower percentage of CD38+ primary AML blasts when they were cocultured with CAR38 NK cells compared with NT NK cells (Figure S4F). As expected, CAR38 NK cells also eliminated CD38-expressing hematopoietic stem cells from healthy donors (Figure S4G).
Next, we explored whether the absence of CD226 expression would influence CAR NK cell activity. First, we transduced NK cells with CAR38.3ζ.IL-15, followed by CD226 KO, and tested their functionality. We achieved a mean combined CAR38.3ζ.IL-15 transduction and CD226 KO efficiency of 88.7% (range: 86%–97.6%; n = 7 donors), with comparable CD226 KO levels in both NT and CAR38.3ζ.IL-15-transduced NK cells, as well as similar CAR38.3ζ.IL-15 transduction efficiency in the CD226 KO and CTRL groups (Figure 4F). Although CD226 KO significantly reduced the activity of NT NK cells, it did not affect degranulation or cytokine production by CAR38.3ζ.IL-15 NK cells (Figures 4G–4I). Additionally, we confirmed that CAR38.3ζ.IL-15 NK cells retained their ability to kill leukemic cells effectively, independent of CD226 expression (Figure 4J), even against cell lines that express lower CD38 levels, such as U937 and MV411 (Figure S4H). Moreover, NT NK cells lacking CD226 lost tumor control significantly earlier than their non-KO controls, while CAR38.3ζ.IL-15 NK cells sustained their potent anti-tumor response regardless of CD226 status (Figures 4K and 4L).
To test if CAR38.3ζ.IL-15 NK cells could compensate for the absence of CD226 in vivo, we administered CTRL and CD226 KO CAR38.3ζ.IL-15 NK cells into an AML NOD/SCID IL-2 Rγnull (NSG) mouse model. CTRL and CD226 KO CAR38.3ζ.IL-15 NK cells demonstrated comparable anti-leukemic activity, with both showing enhanced anti-leukemic activity and prolonged survival compared to NT NK cells (Figures 4M–4O). Thus, CAR38.3ζ.IL-15 NK cells showed similar tumor control in both the presence and absence of CD226, suggesting that CAR38.3ζ.IL-15 may overcome the need for CD226 expression. These findings indicate that CAR38.3ζ.IL-15-armed NK cells maintain their anti-leukemic properties even in the absence of CD226, suggesting that directing the specificity of NK cells toward a target antigen with a CAR could potentially circumvent this immune escape mechanism in AML.
CAR38 NK cells form effective immunological synapses with their targets independently of CD226 and LFA-1
Next, we investigated whether CAR38 NK cells maintain their effector function against AML targets by forming effective immune synapses without CD226. To dissect the individual contributions of the CD38-directed scFv portion, the CD3ζ signaling domain, and IL-15 to synapse formation, we generated two additional CD38-targeting CAR constructs: one without the IL-15 transgene (CAR38scFv.3ζ) and another lacking both IL-15 and the CD3ζ domain (CAR38scFv) (Figure 5A). NK cells transduced with the different CAR constructs showed comparable CAR expression (Figure S5A). We confirmed the expression of CD3ζ in full-length CAR38.3ζ.IL-15 and CAR38scFv.3ζ (Figure S5B), and IL-15 was exclusively released by cells transduced with the full-length CAR38.3ζ.IL-15 construct (Figure S5C). In cocultures with AML cells, all CAR38-transduced NK cells displayed higher degranulation (Figure 5B), cytokine production (Figures 5C, S5D, and S5E), and cytotoxicity (Figure S5F) than NT NK cells. Of note, this was also observed with CAR38scFv in the absence of the CD3ζ signaling endodomain. These data suggest that the improvement in NK cell effector function facilitated by the CAR could be in part due to the binding of the scFv-transmembrane moiety to its cognate ligand.
Figure 5. CAR38 NK cells build effective immune synapses in the absence of CD226 and reduced LFA-1 recruitment.
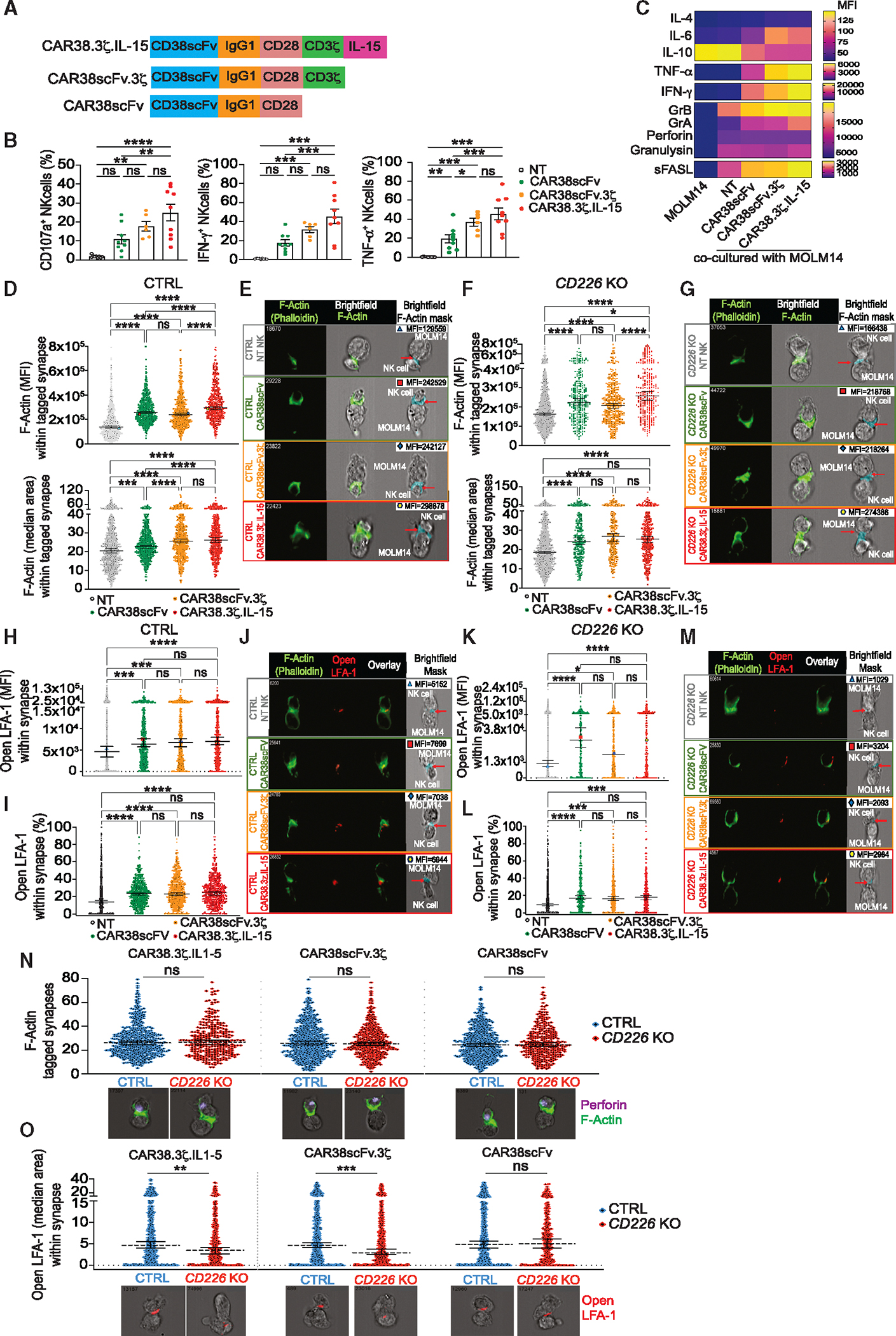
(A) Schematic representation of CAR38 constructs.
(B) Flow cytometric quantification of CD107a, IFN-γ, and TNF-α expression by NT and CD38 CAR-transduced NK cells cocultured with MOLM14 (E:T ratio 1:1) (n = 6–8 donors; three independent experiments).
(C) Heatmap displaying cytokine profile in the supernatants of NK cell-MOLM14 cocultures (E:T ratio 1:1) after 16 h by multiplex bead assay for flow cytometry (n = 5 donors; two independent experiments).
(D, H, and I) Dot plots showing the quantification of F-actin (D) and open LFA-1 (H and I) in the tagged synapses in CTRL subgroups. Groups: NT (1,014 synapses), CAR38scFv (1,654 synapses), CAR38scFv.3ζ (1,284 synapses), and CAR38.3ζ.IL-15 (1,320 synapses) (n = 3 donors; two independent experiments).
(E and G) Representative images of F-actin accumulation within the synapses. Bright-field images display the threshold mask (red arrow) used to quantify F-actin.
(F, K, and L) Dot plots showing the quantification of F-actin (F) and open LFA-1 (K and L) in the tagged synapses in CD226 KO subgroups. Groups: NT (1,401 synapses), CAR38scFv (427 synapses), CAR38scFv.3ζ (544 synapses), and CAR38.3ζ.IL-15 (456 synapses) (n = 2 donors).
(J and M) Representative images of open LFA-1 accumulation within the synapses. Bright-field images display the phalloidin threshold mask intersection with open LFA-1 (red arrow) used to quantify LFA-1.
(N and O) Dot plots comparing the levels of F-actin (N) and open LFA-1 (O) in the tagged synapses between CTRL and CD226 KO groups in the context of CD38 CAR transduction (n = 2 donors; two independent experiments). Analyzed synapses in CTRL groups: CAR38scFv (1,790 synapses), CAR38scFv.3ζ (1,178 synapses), and CAR38.3ζ.IL-15 (982 synapses). Analyzed synapses in CD226 KO groups: CAR38scFv (427 synapses), CAR38scFv.CD3ζ (544 synapses), and CAR38.3ζ.IL-15 (457 synapses). Below the plots are respective representative images. The top right number is the phalloidin (N) or perforin (O) area of the represented synapse.
Statistics: ordinary two-way ANOVA with Bonferroni’s correction for multiple comparisons (B), Kruskal-Wallis test with Dunn’s multiple correction test (D, F, H, I, K, and L), and Mann-Whitney test (N and O). Mean ± SD (B), median fluorescence intensity (MFI) (C), and median with 95% CI (D, F, H, K, L, N, and O). *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
The scFv-transmembrane portion of CAR38 could facilitate the interaction between NK cells and AML cells, leading to the formation of robust immunological synapses. All CAR38-directed NK cells, irrespective of the presence or absence of CD226, showed higher accumulation of F-actin at the immune synapse compared to NT NK cells (Figures 5D–5G), indicating that CD38-targeting CAR NK cells mobilize F-actin more efficiently than their NT counterparts. Furthermore, the scFv-transmembrane portion alone (in the absence of signaling) led to an improvement in F-actin accumulation within the immunological synapse, suggesting how this moiety may at least partially support NK cell effector function against AML cells.
Improved synapse formation by CAR38 NK cells could be related to their ability to recruit LFA-1 into the immunological synapse. We observed that, while overall LFA-1 levels remained similar (Figure S5G), CAR38 NK cells showed greater recruitment of active LFA-1 to the immunological synapse compared to NT NK cells (Figures 5H–5M, S5H, and S5I), even in the absence of CD3ζ signaling. However, CAR38 NK cells did not enhance the colocalization of CD226 and LFA-1 within the immune synapse (Figure S5J), suggesting that CARs targeting CD38 can interact with LFA-1 independently of both CD226 and CD3ζ signaling pathways.
We also found that the deletion of CD226 did not affect F-actin accumulation in CD38 CAR NK cell-AML cell synapses (Figure 5N), even in the absence of the CD3ζ signaling domain, indicating robust immune synapse formation through the CD38 scFv independent of CD226. However, CAR38 NK cells lacking CD226 and bearing the CD3ζ endodomain showed a 25%–55% reduction in their ability to recruit open LFA-1 to the immune synapses (p < 0.0001 for CAR38.3ζ.IL-15 and CAR38scFv.3ζ), while CAR38scFv lacking CD226 showed similar levels of LFA-1 inside the immune synapse (Figure 5O). These data suggest that while CAR38 recruits and cooperates with LFA-1 for optimal immune synapse formation, the intrinsic activity of the CAR38 construct may allow these cells to retain functionality even with reduced LFA-1 recruitment. This suggests a potential compensatory mechanism by the CAR’s engagement with its cognate ligand, enabling effective immune synapse formation and function by NK cells despite CD226 downregulation.
Conjugate formation by CAR38 NK cells is independent of LFA-1 but depends on the density of CD38 antigen on target cells
To investigate whether LFA-1 is essential for immune synapse formation between CAR NK cells and their targets, we blocked the open, active LFA-1 using an inhibitor, BIRT 377.30 We first tested the effect of LFA-1 inhibition on NT NK cells cocultured with K562 cells, which are intrinsically sensitive to NK cell cytotoxicity. NT NK cells expressed lower levels of open LFA-1 in the presence of BIRT 377 (Figure 6A) without alterations to total LFA-1 levels (Figure S6). This was associated with a significant reduction in the number of conjugates between NT NK and K562 cells (Figures 6B and 6C), highlighting the importance of LFA-1’s active configuration in the formation of these conjugates. We then tested the role of LFA-1 in the context of CAR38 NK cells. NK cells expressing CAR38.3ζ.IL-15, CAR38scFv.3ζ, or CAR38scFv maintained their ability to form conjugates with CD38-expressing cells in the presence of BIRT377. This was not observed with NT or NK cells expressing a CAR targeting an irrelevant antigen, CD5 (CAR5), not expressed on MOLM14 cells (Figure 6D). Furthermore, the cytotoxicity of NT NK and CAR5 NK cells against MOLM14 was reduced by LFA-1 inhibition, underscoring the importance of LFA-1 in their cytotoxic function (Figure 6E). Remarkably, inhibition of LFA-1 did not affect conjugate formation by CD38 CAR NK cells, not even in the absence of CD3ζ signaling. This suggests that the scFv component of CAR38 may overcome the need for LFA-1 in synapse formation.
Figure 6. The levels of CD38 on AML cells determine LFA-1 independent of CD38 CAR-targeting NK cells for conjugate formation and effector function.

(A) Open LFA-1 expression in NK cells cocultured with K562 (E:T ratio 1:1) ± BIRT 377 (20 μM) assessed by flow cytometry (n = 4 donors; two independent experiments).
(B) Representative flow cytometry plots displaying conjugate formation between NK and K562 cells ± BIRT 377.
(C) Bar plots showing the percentage of conjugates among live cells in NK cell-K562 cocultures in the presence or absence of BIRT 377. The same experiments were used as in (B).
(D) Dot plots showing the frequencies of conjugates between MOLM14 and NK cells transduced with the different CD38-targeting CARs vs. CD5 CAR vs. NT NK cells in the presence or absence of BIRT 377 by flow cytometry. Each dot represents one donor (n = 5–6; three independent experiments).
(E) Dot plots comparing MOLM14 killing by NT or NK cells expressing different CD38-targeting CARs in the presence and absence of BIRT 377 during a chromium-release assay (20:1 E:T ratio). Each dot represents one donor (n = 7; three independent experiments).
(F) Representative flow cytometry plot showing the CD38 expression by AML cell lines.
(G) Bar graph comparing the average number of CD38 molecules per AML cell line estimated by flow cytometry.
(H–J) Dot plots showing AML killing by NT and CD38 CAR-transduced NK cells (20:1 E:T ratio) when treated with BIRT 377 during a chromium-release assay (n = 7 donors; three independent experiments). Bar graphs display the relative change in the killing of AML cell lines by NT and CD38-targeting NK cells in the presence or absence of BIRT 377.
Statistics: ordinary two-way ANOVA with Bonferroni’s correction for multiple comparisons (A and C), two-tailed paired t Student’s test (D, E, and H–J). Mean ± SD are shown (A, C, D, H, I and J). *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
The effectiveness of CARs in targeting tumors closely correlates with the expression levels of the cognate ligand on tumor cells.31 We hypothesized that the dependence of CAR38 NK cells on LFA-1 activity is inversely proportional to CD38 expression levels on AML targets, with higher CD38 expression levels reducing LFA-1 dependence, while lower CD38 levels necessitate increased LFA-1 activity by CAR38 NK cells for effective immunological synapse formation and cytotoxicity. To explore this relationship, we assessed the levels of CD38 in several AML cell lines (Figures 6F and 6G). U937 and MV411 had, respectively, 125 and 160 times fewer CD38 molecules on their surface compared to MOLM14 cells; MOLM13 cells displayed intermediate levels of CD38 (20 times fewer CD38 molecules per cell than MOLM14). While the cytotoxicity of CD38-targeting CAR NK cells against MOLM14 was not affected by LFA-1 inhibition (Figure 6E), AML cell lines with lower CD38 levels, such as MOLM13, U937, and MV411, showed a significant decrease in susceptibility to CAR38 NK cell killing when LFA-1 was blocked (Figures 6H–6J). These observations suggest that the ability of CAR38 NK cells to kill was independent of LFA-1 when CD38 was expressed at high levels, while intermediate or low CD38 expression required active LFA-1. This indicates a flexibility in the killing mechanism of CAR NK cells, suggesting that they are capable of leveraging both the engagement of the scFv with its cognate antigen and their innate receptor signaling to maintain their anti-leukemic activity.
DISCUSSION
The inherent ability of NK cells to recognize and kill AML cells depends on signals from their germline-encoded receptors, including the activating receptor CD226. Here, we showed that the expression of CD155 on AML cells induces the downregulation of CD226 on NK cells, serving as a mechanism of immune evasion. This downregulation impaired the ability of NK cells to recruit LFA-1 and form robust immune synapses. Engineering NK cells to express a CAR against CD38 overcame the need for CD226 by enabling NK cells to form strong immune synapses with AML blasts. Thus, CAR38 NK cells may overcome immune escape mechanisms secondary to CD226 downregulation by bypassing the need for LFA-1 in synapse formation.
Our results support the crucial role of LFA-1 in NK cell cytotoxicity against AML. CD226 engagement initiates an inside-out signal that primes LFA-1, changing its conformation to an open active form, thus enabling its binding to ICAM-1 on target cells.32–36 These events culminate in a cascade of LFA-1-driven F-actin polymerization, cytoskeleton rearrangement, and immunological synapse formation, building a scaffold for other surface-activating receptors and cytoplasmic-activatable enzymes.37–40 Moreover, this mechanical cell-cell interaction facilitates the delivery of lytic granules to the immune synapse, enhancing NK cell cytotoxic function.41–44 We thus showed the essential role of CD226/LFA-1 interactions for the cytolytic potential of NK cells against AML since the absence of CD226 led to a decrease in LFA-1 recruitment, less actin polymerization, and suboptimal immunological synapse formation between NK and AML cells.
The formation of immunological synapses is also pivotal to the function of CARs.45,46 Our study showed that even with CD226 KO and reduced LFA-1 recruitment into the synapse, NK cells expressing CAR38 preserved their ability for actin polymerization, resulting in strong immunological synapses and potent anti-leukemic cytotoxicity. Remarkably, CAR NK cells maintained their effector function to some extent, even in the absence of CD3ζ signaling, highlighting the importance of the scFv within the CAR construct for initiating cell-to-cell contact, actin remodeling, and NK cell activation. Moreover, the anti-leukemic activity of CAR38 NK cells was partially independent of LFA-1, depending instead on the levels of CD38 antigen on leukemia cells. Consequently, if the AML cells express the target antigen in sufficient quantities, then the cytolytic activity of NK cells can be driven by the synthetic CAR, which simulates the function of the native receptor LFA-1.
Low CD226 levels in effector cells are associated with unfavorable cancer outcomes, including after treatment with immune checkpoint inhibitors.47–52 In patients with myeloid malignancies, NK cell function is impaired, with markedly reduced CD226 levels and poor NK cell cytotoxicity.13,15,53–57 Our data support the notion that CD226 downregulation serves as a mechanism of immune evasion and NK cell dysfunction in AML. However, the partial reduction in cytotoxicity observed with CD226 KO suggests that other activating receptors may compensate for the loss of CD226, maintaining some level of NK cell activity. This indicates that while CD226 plays an important role in NK cell-mediated cytotoxicity, it is not the only pathway involved, and other mechanisms likely contribute to NK cell function against AML.
Our study also showed that CD226 downregulation in NK cells is largely attributed to its interaction with CD155 on AML cells. These data are in keeping with a study by Braun et al. reporting that CD226 is downregulated on murine T cells when they are exposed to CD155-coated beads, leading to internalization, ubiquitination, and subsequent degradation of CD226.51 This phenomenon is not unique to AML, and in other cancers, the direct interaction between CD226 and CD155 is essential for CD226 downregulation in NK cells.49,58 Our study, therefore, showed that the mechanism of CD155-driven NK cell immune escape equally exists in AML.
Patients with AML typically have a poor prognosis, making NK cell-based therapies an appealing therapeutic approach. Insights into the interplay between synthetic receptors such as CARs and NK cell natural receptors could lead to advances in NK cell-based cellular therapies. Our study showed for the first time how NK cells expressing a CAR can overcome immune evasion mechanisms in AML, such as CD226 downregulation, which affects the recruitment of LFA-1 into the immune synapse. We also observed parallels between CAR engagement and LFA-1 function, with both promoting immune synapse formation and F-actin polymerization, resulting in NK cell activation. Thus, CAR-engineered NK cells offer an attractive option for the treatment of AML due to their capacity to overcome AML-related immune escape mechanisms while retaining their native receptor function. This dual capability allows CAR-engineered NK cells to remain highly effective against AML cells, providing a potent response by engaging both synthetic and innate pathways of immune activation. This innovation in cancer immunotherapy holds promise for improving the outcomes of patients with AML.
Limitations of the study
The scope of our study was to examine the immunological synapse in CAR38 NK cells, focusing on how the loss of CD226 and the associated reduction in LFA-1 recruitment contribute to the immune evasion mechanisms of AML. While we have established that CAR38 NK cells, even in the absence of CD3ζ signaling, can form an immunological synapse AML cell, resulting in intracellular signaling and cytotoxicity, the exact mechanisms remain unclear.
RESOURCE AVAILABILITY
Lead contact
Requests for further information and resources should be directed to and will be fulfilled by the lead contact, Katayoun Rezvani (krezvani@mdanderson.org).
Materials availability
The data and materials generated in this study will be made available upon request, but we may require a completed materials transfer agreement if there is potential for commercial application.
Data and code availability
All data reported in this paper will be shared by the lead contact upon request. This paper analyzes existing, publicly available data, accessible at TCGA (https://doi.org/10.1056/NEJMoa1301689), BeatAML (https://doi.org/10.1038/s41586-018-0623-z), and Leucegene (GEO: GSE67040).
This paper does not report original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
STAR★METHODS
EXPERIMENTAL MODEL AND STUDY PARTICIPANT DETAILS
Cell lines and cell cultures
HL-60 (acute promyelocytic leukemia) and MV-4–11 (biphenotypic B myelomonocytic leukemia) cells were grown in Iscove’s Modified Dulbecco’s Medium (IMDM) supplemented with 10% fetal bovine serum (FBS), 1% glutamine, and 1% Penicillin-Streptomycin. K562 (erythroleukemia), MOLM14 (acute myeloid leukemia FAB M5a), OCI-AML3 (acute myeloid leukemia FAB M4), THP-1 (acute monocytic leukemia), U-937 (acute monocytic leukemia), U-937Cd155−/−, and U-937Cd112−/− cell lines were maintained in Roswell Park Memorial Institute (RPMI) medium supplemented with 10% fetal bovine serum (FBS), 1% glutamine, and 1% Penicillin-Streptomycin. Medium was changed every other day. All cells were maintained in a humidified 37°C incubator with 5% CO2, and regularly tested for mycoplasma contamination using the MycoAlert Mycoplasma Detection Kit (Lonza).
Primary cell cultures
Cord-blood (CB) NK cell isolation, culture, and expansion
Freshly collected CB units were obtained from the MD Anderson CB Bank. CB-NK cells were isolated by density-centrifugation method from CB mononuclear cells using Lymphoprep followed by negative selection performed with NK cell isolation kit. CB-NK cell purity (CD56+CD3−cells) was verified by flow cytometry. On day 0, CB-NK cells were expanded with irradiated (100 Gy) uAPCs (K562 feeder cells retrovirally transduced to co-express membrane-bound IL-21, CD48, and 1–4BBL) in a 2 uAPC: 1 CB-NK cell ratio. CB-NK cells were cultured in 10% fetal bovine serum (FBS) Click’s 50/RPMI 50 medium supplemented with 1% glutamine, 1% Penicillin-Streptomycin and recombinant human IL-2 (final concentration 200 U/ml) in a humidified incubator at 37°C/5%CO2. Medium was changed every other day, and IL-2 was added in each passage. After isolation, CB-NK cells were re-stimulated with irradiated feeder cells on days 7 and 14.
In vivo animal model
NOD/SCID IL-2 Rγnull (NSG) mice purchased from The Jackson Laboratory were kept under pathogen-specific conditions at the Research Animal Support Facility at MD Anderson Cancer Center. All subjects were female and treatment naive. The experiments were performed in accordance with the American Veterinarian Association and National Institute of Health (NIH). The protocol was approved by MD Anderson Cancer Center Animal Care and Use Institutional Committee (protocol no. 00000889-RN02).
Patients and healthy donor specimens
All assays involving human participants were approved by the MD Anderson Cancer Center Ethics Committee and were in accordance with the Declaration of Helsinki. Tissue and blood samples were collected after obtaining written informed consent in accordance with MD Anderson Cancer Center Ethics Committee procedures and guidelines.
AML samples
Peripheral blood samples (stored in 90% FBS and 10% DMSO at liquid nitrogen) from patients with AML were obtained from the Leukemia Department of MD Anderson Cancer Center. Upon reception of the samples, leukocytes were separated using the density centrifugation method (Lymphoprep), and they were cryopreserved in 90% FBS and 10% DMSO. For the immunophenotyping, cryopreserved samples were thawed, followed by staining for flow or mass cytometry (as described below). For functional assays, after thawing the samples, leukemic cells were magnetically selected using CD3, CD14, and CD19 beads (Miltenyi Biotec). In summary, samples were thawed by adding cold RPMI and benzonase (100 U/ml) and then centrifuged at 400g for 5 min. After discarding the supernatant, the cells were resuspended in RPMI for counting, washed and centrifuged again with CliniMACS. After discarding the supernatant, cells were then resuspended in CliniMACS PBS/EDTA buffer (Miltenyi Biotec), followed by the addition of CD3, CD14 and CD19 beads (Miltenyi Biotec) to the cell suspension as per manufacturer’s instructions. After 30-min incubation at room temperature, the cell suspension was added to the magnetic columns for separation. The flow-through, which contained the negatively selected leukemic cells, was collected and centrifuged at 400g for 5 min; the supernatant was then discarded, and the cell pellet was resuspended in RPMI medium. Selected leukemic cells were kept overnight in a humidified incubator at 37°C/5%CO2 in RPMI supplemented with 1% glutamine, 1% Penicillin-Streptomycin and enriched with IL-3 (10 ng/mL), stem cell factor (SCF; 25 ng/mL) and IL-6 (10 ng/mL).
METHOD DETAILS
Single-cell RNA sequencing and analysis of AML samples
AML samples used for analysis included 864 samples collected at disease diagnosis from BEAT-AML (n = 173), TCGA (n = 281), and Leucegene (n = 410), constituting a combined cohort previously analyzed in Zeng et al. 2022. Raw counts from patients within each of the three AML cohorts were subject to VST normalization with DESeq2. VST normalized expression measurements of 19 NK ligands was combined across the three cohorts and batch correction was performed with Combat prior to PCA dimensionality reduction. UMAP dimensionality reduction was performed through the scanpy package, using the 10 nearest neighbors and a minimum distance of 0.1. NK Ligand-based clusters were defined with Leiden clustering at a resolution of 0.3, yielding four clusters. One of the four clusters was comprised nearly entirely of Acute Promyelocytic Leukemia (APL) samples, this was removed from downstream analysis. Gene expression measurements were scaled prior to heatmap visualization. Comparisons between groups were made using a wilcoxon rank-sum test. Combined genomic annotations for these 864 patient samples were obtained from the Zeng et al. 2022 publication, and over-representation analysis of genomic subtypes across the NK-ligand based clusters was performed through chi-squared tests.
LSCs were categorized based on their transcriptomic profile. Briefly, RNA sequencing was performed in 111 AML fractions used for xenotransplants and classified as having an engrafting or a non-engrafting potential. The transcriptomic profiles of the engrafting and non-engrafting cells were used to define the LSC+ and the LSC− groups, respectively. Comparisons between LSC+ and LSC− were made through vst normalized and scaled RNA-seq data from Zeng et al. 2022 (GSE199452), as well as variance stabilized, quantile normalized, and log transformed microarray data from Ng et al. 2016 (GSE76008). Comparisons were made through a wilcoxon rank-sum test. Comparisons between diagnosis and relapse were made using logTPM normalized data from four cohorts comprised of diagnosis-relapse AML pairs. These datasets include Li et al. Nature Medicine 2016, Shlush et al. Nature 2017 (GSE199451), Christopher et al. NEJM 2018, and Cocciardi et al. Nature Communications 2019. Paired comparisons between diagnosis and relapse were made using a wilcoxon signed rank test.
Mass cytometry
Mass cytometry antibody preparation and staining for the characterization of NK cells from AML samples
A summary of mass cytometry antibodies, isotopes, and clones can be found in the key resources table. The antibodies conjugated in-house were purchased in a carrier-free form and conjugated (100 μg) with the corresponding metal tag using the Maxpar X8 conjugation kit according to the manufacturer’s recommended protocol (Standard Biotools). Metal isotopes were acquired from Standard Biotools. All conjugated antibodies were diluted in PBS-based antibody stabilization solution supplemented with 0.05% sodium azide to a final concentration of 0.5 mg/mL. Serial titration experiments determined the concentration with the optimal signal-to-noise ratio for each antibody.
KEY RESOURCES TABLE.
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
|
| ||
| Antibodies | ||
|
| ||
| Alexa Fluor 647 AffiniPure F(ab’)2 Fragment Goat Anti-Human IgG (H + L) | Jackson Immunoresearch Laboratories | Cat# 109-606-088; RRID: AB_2337897 |
| Alexa Fluor 647 anti-human CD11a/CD18 (LFA-1) antibody – Clone m24 | Biolegend | Cat# 363412; RRID: AB_2716175 |
| Anti-human CD2 (TS1/2) – 151Eu | Standard Biotools | Cat# 3151003B; RRID: AB_3106938 |
| Anti-human CD16 (3G8) – 209Bi | Standard Biotools | Cat# 3209002B; RRID: AB_2756431 |
| Anti-human CD25 (2A3) – 149Sm | Standard Biotools | Cat# 3149010B; RRID: AB_2756416 |
| Anti-human CD33 (WM53) – 169Tm | Standard Biotools | Cat# 3169010B; RRID: AB_2802111 |
| Anti-human CD39 (A1) – 160Gd | Standard Biotools | Cat# 3160004B; RRID: AB_2687648 |
| Anti-human CD45 (HI30) – 89Y | Standard Biotools | Cat# 3089003B; RRID: AB_2938863 |
| Anti-human CD56/NCAM (NCAM 16.2)- 149Sm | Standard Biotools | Cat# 3149021B; RRID: 2938638 |
| Anti-human CD69 (FN50) – 144ND | Standard Biotools | Cat# 3144018B; RRID: AB_2687849 |
| Anti-human CD85j (GHI/75) – 156Gd | Standard Biotools | Cat# 3156020B; RRID: AB_2756420 |
| Anti-human CD95/Fas (DX2) – 152Sm | Standard Biotools | Cat# 3152017B; RRID: AB_2920762 |
| Anti-human CD117/c-kit (104D2) – 143ND | Standard Biotools | Cat# 3143001B; RRID: AB_2847864 |
| Anti-human CD123 (6H6) – 151Eu | Standard Biotools | Cat# 3151001B; RRID: AB_3661860 |
| Anti-human CD134/OX40 (ACT35) – 158Gd | Standard Biotools | 3158012B |
| Anti-human CD159a/NKG2A (Z199) – 169Tm | Standard Biotools | Cat# 3169013B; RRID: AB_2756426 |
| Anti-human CD161 (HP-3G10) – 159Tb | Standard Biotools | Cat# 3159004B; RRID: AB_2756421 |
| Anti-human CD196/CCR6 (G034E3) – 141Pr | Standard Biotools | Cat# 3141003A; RRID: 2687639 |
| Anti-human CD223/LAG3 (11C3C65) – 173Yb | Standard Biotools | Cat# 3175033B; RRID: AB_2888932 |
| Anti-human CD279/PD-1 (EH12.2H7) – 165Ho | Standard Biotools | Cat# 3165042B; RRID: AB_2905648 |
| Anti-human CD314/NKG2D (ON72) – 166Er | Standard Biotools | Cat# 3166016B; RRID: AB_2892110 |
| Anti-human CD335/NKp46 (BAB281) – 162Dy | Standard Biotools | 3162021B |
| Anti-human CD352 (NT-7) – 143 Nd | Standard Biotools | 3143017B |
| Anti-human CD371/CLEC12A (50C1) – 173Yb | Standard Biotools | Cat# 3173007B; RRID: AB2892698 |
| Anti-human Granzyme B (GB11) – 171Yb | Standard Biotools | Cat# 3171002B; RRID: AB_2687652 |
| Anti-human Granzyme B (GB11) – 173Yb | Standard Biotools | Cat# 3173006B; RRID: AB_2811095 |
| Anti-human Ki-67 (B56) – 162Dy | Standard Biotools | Cat# 3162012B; RRID: AB_2888928 |
| Anti-human TIGIT (MBSA43) – 154Sm | Standard Biotools | Cat# 3154016B; RRID: AB_2888926 |
| Anti-human TIM-3 (F38-2E2) – 153Eu | Standard Biotools | Cat# 3153008B; RRID: AB_2938868 |
| Anti-human/mouse TBET (4B10) – 160Gd | Standard Biotools | Cat# 3160010B; RRID: AB_2810251 |
| Anti-Perforin Antibody (B-D48) | Abcam | Cat# Ab47225; RRID: AB_2169084 |
| Anti-rabbit IgG, HRP-linked antibody | Cell Signaling Technology | Cat# 7074P2; RRID: AB_2099233 |
| APC anti-human CD13 antibody – Clone WM15 | Biolegend | Cat# 301706: RRID: 314182 |
| APC anti-human CD155 (PVR) antibody - Clone SKII.4 | Biolegend | Cat# 337618; RRID: AB_2565815 |
| APC-H7 mouse anti-human CD3 – Clone SK7 | BD Biosciences | Cat# 560176; RRID: AB_1645475 |
| BD Pharmigen Purified Mouse Anti-Human CD18 – Clone 6.7 | BD Biosciences | Cat # 555922; RRID: AB_396223 |
| Biotin-SP (long-spacer) AffinityPure F(ab’)2 Fragment Goat anti- Human IgG (H + L) | Jackson Immunoresearch Laboratories | Cat# 109-066-088; RRID: AB_2337638 |
| Brilliant Violet 421 anti-human CD38 antibody – Clone HIT2 | Biolegend | Cat# 303526; RRID: AB_10983072 |
| Brilliant Violet 605 anti-human CD56 (NCAM) Antibody – Clone HCD56 | Biolegend | Cat# 318333; RRID: AB_11142683 |
| Brilliant Violet 711 anti-human CD19 antibody - Clone HIB19 | Biolegend | Cat# 302246; RRID: AB_2562065 |
| Brilliant Violet 785 anti-human CD14 antibody - Clone M5E2 | Biolegend | Cat# 301840; RRID: AB_2563425 |
| Brilliant Violet 785 anti-human CD107a (LAMP-1) antibody - Clone H4A3 | Biolegend | Cat # 328644; RRID: AB_2565968 |
| BUV395 Mouse Anti-Human CD33 - Clone WM53 | BD Biosciences | Cat# 740293; RRID: AB_2740032 |
| BV421 mouse anti-human CD123 - Clone 9F5 | BD Biosciences | Cat# 562517; RRID: AB_11153668 |
| CD25 Antibody, anti-human REAfinity – Clone REA570 | Miltenyi Biotec | Cat# 130-122-302; RRID: AB_2801883 |
| CD159c (NKG2C) Antibody, anti-human, REAfinity – Clone REA205 | Miltenyi Biotec | Cat# 130-122-278; RRID: AB_2801859 |
| CD226 (DNAM-1) Antibody, anti-human, PE- Clone DX11 | Miltenyi Biotec | Cat# 130-124-232; RRID: AB_2857678 |
| CD279 (PD1) Antibody, anti-human – Clone PD1.3.1.3 | Miltenyi Biotec | Cat# 130-096-168; RRID: AB_10828445 |
| EOMES monoclonal antibody (WD1928) | Thermo Fischer Scientific | Cat# 50-4877-42; RRID: AB_2574229 |
| FITC anti-human CD226 (DNAM-1) antibody - Clone 11A8 | Biolegend | Cat# 338304; RRID: AB_2228763 |
| Human DAP10/HCST antibody – Clone 982129 | RnD Systems | Cat# MAB9786; RRID: AB_3659541 |
| Human DAP12 Antibody – Clone 406209 | RnD Systems | Cat# MAB52401 ; RRID: AB_3658432 |
| Human KIR/CD158 antibody – Clone 180704 | RnD Systems | Cat# MAB1848; RRID: AB_2111687 |
| KLRG1 Monoclonal antibody (13F12F2), Functional Grade | Thermo Fischer Scientific | Cat# 16-9488-85; RRID: AB_2637116 |
| LAG-3 Antibody (BLR027F) | Novus Biologicals | Cat# NBP2-76402; RRID: AB_3403543 |
| PE anti-human CD15 (SSEA-1) antibody – Clone W6D3 | Biolegend | Cat # 323006; RRID: AB_756012 |
| PE anti-human CD38 Antibody – Clone HIT2 | Biolegend | Cat# 303506; RRID: AB_314358 |
| PE anti-human CD112 (Nectin-2) Antibody - Clone TX31 | Biolegend | Cat# 337409; RRID: AB_2174163 |
| PE anti-human INF-γ Antibody - Clone B27 | Biolegend | Cat# 506507; RRID: AB_315440 |
| PE mouse anti-human CD226 - Clone DX11 | BD Biosciences | Cat# 559789; RRID: AB_397330 |
| PE/Cyanine 7 anti-human TNF-α antibody - Clone Mab11 | Biolegend | Cat# 502930; RRID: AB_2204079 |
| PE-Dazzle 594 anti-human CD226 (DNAM-1) antibody - Clone 11A8 | Biolegend | Cat# 338317; RRID: AB_2721693 |
| Perforin monoclonal antibody (dG9(deltaG9)), PerCP-eFluor 710, eBioscience | Thermo Fischer Scientific | Cat# 46-9994-42; RRID: AB_1944475 |
| Purified anti-human CD3 (Maxpar ready) antibody – Clone UCHT1 | Biolegend | Cat# 300443; RRID: AB_2562808 |
| Purified anti-human CD14 antibody – Clone M5E2 | Biolegend | Cat# 301802; RRID: AB_314184 |
| Purified anti-human CD19 antibody – Clone HIB19 | Biolegend | Cat# 302247; RRID: AB_256815 |
| Purified anti-human CD62L antibody – Clone DREG-56 | Biolegend | Cat# 304802; RRID: AB_314462 |
| Purified anti-human CD96 (TACTILE) Antibody – Clone NK92.39 | Biolegend | Cat# 338402; RRID: AB_1279386 |
| Purified anti-human CD134 antibody – Clone ACT35 | Biolegend | Cat # 350002; RRID: AB_10639951 |
| Purified anti-human CD160 antibody – Clone BY55 | Biolegend | Cat# 341202; RRID: AB_2074411 |
| Purified anti-human CD226 antibody – Clone 11A8 | Biolegend | Cat# 338302; RRID: AB_1279155 |
| Purified anti-human CD244 (2B4) antibody – Clone C1.7 | Biolegend | Cat# 329502; RRID: AB_1279194 |
| Purified anti-human CD253 (TRAIL) antibody – Clone RIK-2 | Biolegend | Cat# 308202; RRID: AB_345288 |
| Purified anti-human CD336 (NKp44) antibody – Clone p44–8 | Biolegend | Cat# 325102; RRID: AB_345288 |
| Purified anti-human CD337 (NKp30) antibody – Clone p30–15 | Biolegend | Cat# 325202; RRID: AB_756106 |
| Purified anti-human CD366 (TIM3) antibody – Clone F38-2E2 | Biolegend | Cat# 345002; RRID: AB_2116574 |
| Purified anti-human Granulysin antibody - Clone DH2 | Biolegend | Cat# 348008; RRID: AB_2563602 |
| Purified anti-human Granzyme A antibody – Clone CB9 | Biolegend | Cat# 507202; RRID: AB_315468 |
| Purified anti-human Ki-67 antibody – Clone Ki-67 | Biolegend | Cat# 350502; RRID: AB_10662385 |
| Purified anti-T-bet antibody – Clone 4B10 | Biolegend | Cat# 644802; RRID: AB_1595503 |
| Rabbit anti-human CD3z antibody | Cell Signaling Technology | Cat# 88083; RRID: AB_3095084 |
|
| ||
| Bacterial and virus strains | ||
|
| ||
| Incucyte Nuclight Red Lentivirus (EF1a, Puro) | Sartorius | 4625 |
| NEB 5-alpha competent E. coli (high efficiency) | NEB | C2987H |
|
| ||
| Biological samples | ||
|
| ||
| Acute myeloid leukemia samples | Leukemia Department, MD Anderson Cancer Center | N/A |
| Cord blood donors | Cord Blood Bank, MD Anderson Cancer Center | N/A |
| Healthy donor PBMCs | Gulf Coast Regional Blood Bank Center | N/A |
|
| ||
| Chemicals, peptides, and recombinant proteins | ||
|
| ||
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 |
| Alt-R S.p. Cas9 Nuclease V3, 500ug | IDT | 1081059 |
| Annexin V binding buffer, 10X concentrate | BD Biosciences | 556454 |
| BIRT 377 | Tocris | 4776 |
| Brefeldin A | Sigma-Aldricth | B7651-5MG |
| Brilliant stain buffer | BD Biosciences | 566349 |
| CellEvent Caspase 3/7 green detection reagent | Invitrogen | C10423 |
| Cell-ID Cisplatin – 198Pt | Standard Biotools | 201198 |
| Clarity Western ECL substrate | BioRad | 1705061 |
| Click’s Medium (EHAA) | Fujifilm | 9195-500ML |
| CliniMACS PBS/EDTA buffer | Miltenyi Biotec | 130-021-201 |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11965 |
| eBioscience Fixable Dye eFluor 660 | Invitrogen | 65-0864-18 |
| Fugene HD Transfection Reagent | Promega | E2312 |
| Ghost dye UV 450 | Tonbo biosciences | 13-0868-T500 |
| Iscove’s Modified Dulbecco’s Medium (IMDM) | Gibco | 12440053 |
| Laemmli sample buffer | BioRad | 1610737 |
| LIVE/DEAD Fixable Aqua Dead Cell Staining Kit, for 405 nm excitation | Invitrogen | L34957 |
| Lymphoprep | StemCell Technologies | 07801 |
| Polyacrylamide NuPAGE Bis-Tris gel | Invitrogen | NP0322BOX |
| Qdot 625 Streptavidin Conjugate | Invitrogen | A10196 |
| Recombinant human IL-3 protein | RnD Systems | 203-IL |
| Recombinant human IL-6 protein | RnD Systems | 206-IL |
| Recombinant human SCF protein | RnD Systems | 255-SC |
| Roswell Park Memorial Institute (RPMI) 1640 Medium Gibco | 12633020 | |
| Retronectin Recombinant Human Fibronectin Fragment | Takara | T100B |
| TBS/Tween | BioRad | BUF028 |
| V500 Annexin V | BD Biosciences | 561501 |
|
| ||
| Critical commercial assays | ||
|
| ||
| 13 multi-analyte LEGENDplex human CD8/NK panel kit | Biolegend | 1610737 |
| BD quantibrite PE Fluorescence quantitation kit | BD Biosciences | 340495 |
| CD3 microbeads, human | Miltenyi Biotec | 130-097-043 |
| CD14 microbeads, human | Miltenyi Biotec | 130-050-201 |
| CD19 microbeads, human | Miltenyi Biotec | 130-050-301 |
| DuoSet ELISA Ancillary reagent kit 2 | RnD Systems | DY008 |
| Fixation/Permeabilization solution Kit | BD Biosciences | 554714 |
| Human IL-15 DuoSet ELISA | RnD Systems | DY24705 |
| iBlot 2 transfer stacks, nitrocellulose, regular size | Invitrogen | IB23001 |
| MycoAlert Mycoplasma Detection kit | Lonza | LT07-318 |
| Neon Transfection System 10ul kit | Invitrogen | MPK1025 |
| NK cell isolation kit, human | Miltenyi Biotec | 130-092-657 |
| PureLink HiPure Plasmid Maxiprep kit | Invitrogen | K210006 |
|
| ||
| Deposited data | ||
|
| ||
| TCGA-LAML cohort | Genomic Data Commons (National Cancer Institute) | https://doi.org/10.1056/NEJMoa1301689 |
| BEAT-AML cohort | Vizome | https://doi.org/10.1038/s41586-018-0623-z |
| Leucegene cohort | GEO | GEO: accession number GSE67040 |
|
| ||
| Experimental models: Cell lines | ||
|
| ||
| 293T | ATCC | CRL-3216 |
| HL-60 (acute promyelocytic leukemia) | ATCC | CCL-240 |
| K562 (erythroleukemia) | ATCC | CRL-3344 |
| MOLM14 (acute myeloid leukemia FAB M5a) | DSMZ | ACC 777 |
| MV-4-11 (biphenotypic B myelomonocytic leukemia) | ATCC | CRL-9591 |
| OCI-AML3 (acute myeloid leukemia FAB M4) | DSMZ | ACC 582 |
| THP-1 (acute monocytic leukemia) | ATCC | TIB-202 |
| U-937 (acute monocytic leukemia) | ATCC | CRL-1593.22 |
| U-937Cd155 −/− | Generated in house | N/A |
| U-937Cd112 −/− | Generated in house | N/A |
|
| ||
| Other | ||
|
| ||
| NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ | The Jackson Laboratory | 005557 |
|
| ||
| Oligonucleotides | ||
|
| ||
| Hs.Cas9.CD226.1.AA Sequence: mG*mU*mU* rArArG rArGrG rUrCrG rArUrC rUrGrA rCrGrG rUrUrU rUrArG rArGrC rUrArG rArArA rUrArG rCrArA rGrUrU rArArA rArUrA rArGrG rCrUrA rGrUrC rCrGrU rUrArU rCrArA rCrUrU rGrArA rArArA rGrUrG rGrCrA rCrCrG rArGrU rCrGrG rUrGrC mU*mU*mU* rU | IDT | 293007027 |
| Hs.Cas9.CD226.1.AC Sequence: mG*mU*mU* rCrArA rGrArU rCrGrG rGrArC rCrCrA rGrCrG rUrUrU rUrArG rArGrC rUrArG rArArA rUrArG rCrArA rGrUrU rArArA rArUrA rArGrG rCrUrA rGrUrC rCrGrU rUrArU rCrArA rCrUrU rGrArA rArArA rGrUrG rGrCrA rCrCrG rArGrU rCrGrG rUrGrC mU*mU*mU* rU | IDT | 293007028 |
| Hs.Cas9.NECTIN2.2.1.AB Sequence:/AltR1/rArCrA rCrCrA rUrCrU rGrCrU rCrGrG rCrCrC rGrArG rUrUrU rUrArG rArGrC rUrArU rGrCrU/AltR2/ | IDT | 303526483 |
| Hs.Cas9.PVR.2.1.AA Sequence:/AltR1/rCrCrU rGrCrG rGrGrA rArCrG rUrGrA rCrGrA rArCrG rUrUrU rUrArG rArGrC rUrArU rGrCrU/AltR2/ | IDT | 303526480 |
|
| ||
| Recombinant DNA | ||
|
| ||
| iC9.CARCD38.CD28-z-2A-IL15 | Generated in-house | N/A |
| iC9.CARCD38.CD28-z | Generated in-house | N/A |
| iC9.CARCD38.CD28 | Generated in-house | N/A |
|
| ||
| Software and algorithms | ||
|
| ||
| Amnis Image Stream | Amnis, EMD Millipore | https://www.luminexcorp.com/imaging-flow-cytometry/ |
| BD LSFortessa™ X-20 cell analyzer | BD Biosciences | – |
| Cytobank platform | Cytobank | – |
| Diva Software | BD Biosciences | – |
| FlowJo software (v.10) | FlowJo LLC | https://www.flowjo.com |
| GraphPad Prism (v.10) | Graph pad Software, Inc | https://www.graphpad.com |
| iBlot 2 transfer device | Invitrogen | – |
| IDEAS Software 6.3 | Amnis, EMD Millipore | https://www.luminexcorp.com/imaging-flow-cytometry/ |
| Incucyte S3 live-cell analysis system | Sartorius | – |
| Living Image v4.4 | Perkinelmer | Perkinelmer.com |
Thawed peripheral blood samples from patients with AML and healthy donors were washed with PBS and incubated for 3 min with 2.5 μM Cisplatin (Fluidigm) for live/dead discrimination. Dead cells were separated from live cells as they acquired the cisplatin. After washing with cell staining buffer (Fluidigm), samples were stained with a freshly prepared surface antibody mixture containing 48 antibodies markers and incubated for 30 min at room temperature. Following washing with cell staining buffer, samples were fixed and permeabilized using the BD Cytofix/Cytoperm kit for 30 min at 4°C. After two additional washes with perm/wash solution, cells were incubated with a mixture of intracellular antibodies for 30 min at 4°C. An additional wash was performed with perm/wash solution, and samples were stored overnight in 500 μL of 1.6% paraformaldehyde solution containing 125 nM iridium nucleic acid intercalator (Fluidigm). Before acquisition on a Helios mass cytometer (Fluidigm) at MD Anderson North Campus Flow Cytometry Core, cells were washed twice with cell MaxPar Cell staining buffer. After discarding supernatant, the pellet was washed with double-deionized water with 0.1% BSA. Supernatant was discarded after centrifugation and the pellet was resuspended in double-deionized water.
Following the acquisition, analysis was performed using FlowJo software for initial clean-up and gating. NK cells were identified sequentially as (1) CD45+, (2) CD14−and CD19−, (3) CD56+ and CD3−, and (4) CD123−. After their identification, NK cells were subdivided into two subpopulations, CD226 positive and CD226 negative/low, based on the CD226 expression of healthy donors. The CD226 positive subpopulation was defined based on the DNAM-1 levels of 90% of NK cells of healthy individuals: CD226 high reflecting a CD226 expression level of at least 90% of the NK cell population of healthy individuals, and CD226 negative/low representing the remaining peripheral blood NK cells. NK cell immunophenotype was determined from 17 AML samples and compared between the CD226 subpopulations using the percentage of expression of each marker by manual gating and their median metal intensity. Subsequently, unsupervised analysis was performed by downsampling of AML samples to 1,450 NK cells per sample to reduce donor induced bias. Exclusion of samples which had less than 500 cells was performed, and NK cells cells from 13 AML samples were concatenated for further analysis. Concatenated NK cells were imported into Cytobank for dimensionality reduction using UMAP algorithm. For further phenotypic characterization of NK cells, clusters were manually identified and compared using the Morpheus software and the measurement of median metal intensity per NK cell marker.
Mass cytometry antibody preparation and staining for the characterization of CD226 knockout CB-NK
Antibody preparation was performed as described above. Serial titration experiments determined the concentration with the optimal signal-to-noise ratio for each antibody. A summary of mass cytometry antibodies, isotopes, and clones can be found in the key resources table. Expanded CB-NK cells were collected between days 14 and 21 after isolation, and antibody staining was performed as described for the AML samples. Data acquisition was performed immediately on a Helios mass cytometer (Fluidigm) at MD Anderson North Campus Flow Cytometry Core. For phenotypic characterization based on the expression of NK cell markers, concatenated CB-NK cells were imported into Cytobank for dimensionality reduction using the t-SNE-cuda algorithm.
CD226 levels measurement using flow cytometry after NK-AML co-culture
For the 24-h assays, NK cells were co-cultured with AML cell lines (1:5 E:T ratio) in a round bottom 96-well plate in triplicates. Cells were collected and stained with antibodies to distinguish NK cells from AML cell lines by flow cytometry. Monoclonal antibodies used for this assay were CD56, CD3, CD33, CD15, CD13, and DNAM-1 (CD226). NK cells were identified as CD56 positive and CD33, CD15, CD13 negative, and CD3 negative. After NK cell discrimination, CD226 levels on NK cells were accessed. CD226 Relative Reduction = [(CD226 baseline expression – CD226 expression after co-culture)/CD226 baseline expression]. Similar co-culture, cell staining, and cell discrimination were used to measure CD226 in NK cells after co-culture with U937 wildtype (WT) and U937 NECTIN2 and PVR knockout (KO) cells.
For the transwell assays, NK cells and AML cell lines were co-cultured in a 1:5 ratio for 24 or 48 h. The transwell inserts had 0.4μm pores. NK cells were cultured in the bottom of the plate, while AML cell lines were cultured inside the transwell insert. The monoclonal antibody panel was the same as in the 24-h assay.
Rechallenge assays were performed by co-culturing NK cells with AML cells in various effector-to-target ratios. Cells were collected for staining every 24 h for a total of 96 h. After cell collection, rechallenges were performed by adding the same number of tumor cells as at the starting point of the co-culture. The monoclonal antibody panel was the same as in the 24-h assay.
Data were acquired using a BD LSRFortessa X20 Flow Cytometer and analyzed using FlowJo (version 10).
Internalization assay using imaging flow cytometry
Expanded CB-NK cells were kept in culture alone or with MOLM14 in a 1:1 E:T ratio in complete RPMI in a round 96-well plate. After 1-h incubation at 37°C, cells were harvested and washed once with PBS. The first-step surface staining for CD226 (FITC; clone 11A8) and CD123 (BV421; clone 9F5) was performed by incubating the cells with the antibody mix for 20 min at room temperature, followed by fixation and permeabilization using BD Cytofix/Cytoperm kit. After permeabilization, anti-CD226 (PE; clone DX11) and anti-perforin antibodies were added, followed by an incubation period of 30 min at 4°C. A preliminary assay using anti-CD226 antibody clone 11A8 was performed to determine the antibody concentration needed to block the anti-CD226 antibody clone DX11, leading to staining saturation of surface CD226. This way, the anti-CD226 antibody clone DX11 could only bind to the intracellular CD226 post-permeabilization. MOLM14 were identified as CD123 positive and perforin negative, while NK cells were perforin-positive and CD123-negative (Figure S2I). Data were then immediately acquired using a four-laser, 12-channel Amnis ImageStream at a 60x fold magnification at low speed. The gating strategy and data analysis workflow can be found in Figure S2I. Focused cells were chosen based on the feature ‘‘gradient RMS’’. Cells with high aspect ratio intensity were selected for further analysis as they were likely singlets. The automated Wizard software was initially used to identify the internalization of CD226, followed by a visual inspection step to confirm the quality of images and the internalized CD226. Data were analyzed using the IDEAS software (version 6.3; Luminex).
Generation of PVR and NECTIN2 knockout U937 cell lines
Stable knockouts (KOs) of the NECTIN2 and PVR genes in U937 AML cell line were created using the targeted-CRISPR-Cas9 technique. Pre-designed single-guide RNAs (sgRNAs) were purchased from Integrated DNA Technologies (IDT). Pre-designed sgRNAs Hs.Cas9.NECTIN2.2.1.AB and Hs.Cas9.PVR.2.1.AA reached a KO efficiency of around 40% for the NECTIN2 and PVR genes, respectively, as measured via flow cytometry. In summary, U937 cells were harvested (2×10E6 cells), washed twice with PBS, and resuspended in R buffer (Invitrogen). Meanwhile, ribonucleoprotein RNP preparation was performed by incubating sgRNA and diluting Cas9 protein (IDT) in a 1:1 ratio with T buffer (5μg Cas9 and 5g sgRNA) for 15 min at room temperature. The incubation product was then used during the electroporation of the cells using the Neon transfection system (Thermo Scientific). Optimized electroporation parameters were 1450 V, 10 ms, and 3 pulses. Cells were then kept in culture, and CD112-and CD155-negative populations were sorted using a BD SORP FACSAria IIu (Aria#1) cell sorter to obtain >98% NECTIN2 and PVR KO populations (Figure S2L).
Generation of CD226 knockout CB-NK cells
On day 6 post-isolation, CB-NK cells were harvested for the CD226 knockout using the CRISPR-Cas9 method. They were washed twice with PBS and resuspended in T buffer (Invitrogen) containing the ribonucleoprotein (RNP) complex. RNP complex was prepared by incubating Cas9 protein and sgRNAs in a 1:1 ratio (5 μg of Cas9 and a total of 5g sgRNAs) for 15 min at room temperature. Pre-designed sgRNAs were purchased from IDT. Two sgRNAs were used to achieve CD226 KO: sgRNA Hs.CAS9.CD226.1.AA and Hs.CAS9.CD226.1.AC. CB-NK cells were then electroporated (1600 V, 10 ms, 3 pulses) in a solution containing T buffer and the RNP complexes. The Cas9 control group consisted of CB-NK cells that underwent electroporation in the presence of Cas9 without sgRNAs. KO efficiency was verified 2–3 days after CD226 KO using flow cytometry. The same procedure was applied for CB-NK cells transduced with CD38 CAR; CD226 KO was performed 48–72 h after CD38 CAR transduction.
NK cell degranulation and intracellular cytokine production
Expanded CB-NK cells (0.2 × 106 cells/well) were harvested and co-cultured with target cells at an effector-to-target ratio of 1:1. Cells were then incubated for 5–6 h in round-bottom 96-well plates at 37°C in the presence of Brefeldin A (BFA, Sigma) and anti-CD107a antibody. In summary, after co-culture, the cells were harvested, washed with PBS, and live/dead staining was performed in PBS for 20 min at room temperature using the UV450 Ghost Tonbo amine-reactive dye, allowing for analysis of dye-negative live cells only. Cells were washed with PBS with 2% fetal bovine serum (FBS). As previously described, surface staining was performed to distinguish NK from AML cells. Anti-human IgG1 Fab fragment was added to the surface antibody mix to identify CAR NK cells. Cells were then incubated for 20 min at room temperature and protected from light. After incubation, cells were washed with PBS 2% FBS. BD Cytofix solution was used to fix and permeabilize the cells at 4°C for 30 min. Following fixation and permeabilization, cells were washed twice with the BD Perm-Wash solution. Intracellular staining was performed using an antibody mix containing anti-IFNγ and anti-TNFα antibodies. Cells were incubated for 30 min at 4°C and washed twice with the BD Perm-Wash solution. Data were acquired up to 16 h following the staining using a BD LSFortessa X-20 cell analyzer (BD Biosciences). NK cells expressing CD38 CARs were gated (CAR38-positive) and compared to NK cells not expressing CD38 CAR molecules (CAR38-negative) within the same group. NK cells alone were used as negative controls. CB-NK cells stimulated with phorbol myristate acetate (PMA, Sigma) and Ionomycin (Sigma) were used as positive controls. Conditions were performed in technical duplicates. Data were analyzed using FlowJo (version 10).
In vitro real-time cytotoxicity assay
To ensure proper identification, AML cell lines were transduced with the Incucyte Nuclight Red Lentivirus (Sartorius), purified using puromycin, and bright mKate2 (λabs/λem = 588/633 nm) AML cells were sorted and kept in culture. On the day of the 24-h assay, AML cells were harvested, washed with PBS, and mixed with NK cells in an effector-to-target ratio of 1:4 in a 96-well plate. CellEvent Caspase 3/7 Green Detection Reagent (λabs/λem = 502/530 nm) was added to detect apoptotic AML cells. Initially, 12,500 NK cells and 50,000 AML cells were mixed in each well of a 48-well plate for the rechallenge assay. Subsequently, 50,000 AML cells were added every 48–72 h to each well of the 48-well plate. Plates were placed in a humidified incubator at 37°C/5%CO2 equipped with an IncuCyte S3 live-cell analysis system (Sartorius). Frames were captured every 3 h for24–72 h from 4 separate 1.75 × 1.29 mm2 regions per well with a 10x objective. Technical duplicates were assessed. For the 24-h assay, the percentage of killing was calculated as (number of mKate2 and Caspase 3/7 overlapped events)/(number of mKate2 events) per well. For the rechallenge assay and Figure S4F, results were based on the mKate2 signal, which reflected the number of AML cells in the wells.
In vivo animal studies
NOD/SCID IL-2 Rγnull (NSG) mice engrafted with MOLM14 cells were used to examine the anti-tumor activity of CAR38.3ζ.IL-15 NK cells. All experiments were performed in accordance with the American Veterinary Medical Association and National Institutes of Health (NIH) recommendations under protocols approved by the MDACC Institutional Animal Care and Use Committee (protocol no. 00000889-RN02). Mice were maintained under specific-pathogen-free conditions, with a 12-h night-day light cycle, and at a stable ambient temperature with 40–70% relative humidity. Ten-week-old female mice were irradiated on day −3 and engrafted with Ffluc-MOLM14 cells (0.5 × 106) on day −2. On day 0, 5 × 106 CAR38.3ζ.IL-15 NK cells with or without CD226 KO were injected. Control groups were injected with NT NK cells or MOLM14 cells only. Mice were randomly assigned to each group and followed for a total of 50 days. Weekly bioluminescence imaging (Xenogen IVIS-200 Imaging System) was performed to monitor tumor growth and data were analyzed using Living Image v4.4.
Immune synapse studies using imaging flow cytometry
NK and AML cells were harvested, washed with PBS, mixed in a 1:1 E:T ratio, and co-cultured in a round-bottom 96-well plate in a humidified incubator at 37°C/5%CO2 for 20 min. The total volume inside the well was 150 μL. The anti-open LFA-1 (clone m24 that binds specifically to LFA-1 in its open conformation29) antibody was added during the co-culture. After incubation, cells were harvested by gently pipetting and washed with PBS. Combined live/dead with fixable dye (excitation: 405nm; emission: 526nm) and the first step of surface staining were performed with an antibody mix containing anti-DNAM-1, anti-CD112, and anti-CD38 antibodies, together with a biotin-conjugated anti-IgG1 Fab fragment for CAR detection. Cells were incubated for 20 min at room temperature with the antibody mix. Following incubation, cells were washed with PBS +1% bovine serum albumin (BSA), and the second step of surface staining was performed with the addition of a QD625-conjugated streptavidin compound. After incubation for 20 min at room temperature, cells were washed with PBS +1% BSA, and fixation and permeabilization were performed with Cytofix solution (BD Biosciences) at room temperature for 15 min. After incubation, cells were washed twice with the BD Perm-Wash solution. A mixture containing anti-human perforin antibody and phalloidin was added, followed by incubation for 20 min at room temperature. Cells were washed once with PBS +1% BSA and left in a concentration of 2×10E6 cells per mL for immediate acquisition using a four-laser, 12-channel Amnis ImageStream at a 60x fold magnification at low speed. The gating strategy and data analysis workflow can be found in Figure S2B. Focused cells were chosen based on the feature ‘‘gradient RMS’’. Events with combined low aspect ratio intensity and area between 200 and 600 μm2 were selected for analysis as they were likely conjugates. During the analysis, thresholds were used to create masks and features. These thresholds were fixed and used across experiments. Synapses were defined as described in Figures S3B and S3C. The valley mask was used to validate the visual inspection mask. The valley mask is a function mask in the IDEAS software that identifies the region of contact between the cells using the brightfield image. The IDEAS software determines a rectangular region in the dimmer portion between two objects,59 the immune synapse. Since the measured phalloidin did not differ between the visual-inspection and the valley mask and the first preserved a higher number of synapses, we pursued the analyses using the visual inspection mask. Threshold masks were established to measure f-actin (phalloidin) and LFA-1. The levels LFA-1 within the synapse were determined based on the intersection between the f-actin (phalloidin) and the LFA-1 threshold masks for each tagged synapse. The percentage of LFA-1 was calculated as [(LFA-1 median area within the synapse)/(total LFA-1 median area)] per conjugate. Data were analyzed using the IDEAS software version 6.3.
Leukemic blast isolation for NK cell functional assays
Leukemic blasts were isolated from the peripheral blood (PB) of patients diagnosed with acute myeloid leukemia (AML) as described above. PB leukocytes were obtained by the density separation method, and aliquots were frozen in liquid nitrogen. PB leukocytes were thawed, and leukemic blasts were isolated by depletion of CD3 positive, CD19 positive, and CD14 positive cells performed with respective magnetic beads. After isolation, leukemic blasts were kept in culture for up to 24 h using IL-3 (10 ng/mL), IL-6 (10 ng/mL), and stem cell factor (SCF; 25 ng/mL) for further NK cell functional studies. Phenotypic characterization of the leukemic cells was performed by flow cytometry to ensure the expression of CD38.
Annexin V assay
On the day of the assay, NK and leukemic cells were harvested, washed with PBS, and co-cultured in a 1:1 E:T ratio for 5–6 h in a round-bottom 96-well plate in a humidified incubator at 37°C/5%CO2. Following the incubation, cells were collected and stained with propidium iodide for 20 min at room temperature in PBS to identify dead cells. Subsequently, cells were washed with PBS +2% FBS, and surface staining for CD56, CD3, and CD38 was performed as described above. Cells were washed twice with annexin V binding buffer, and annexin V was added to the cell suspension. Data were acquired immediately following the staining using a BD LSFortessa X-20 cell analyzer (BD Biosciences). Analysis was performed using FlowJo software (version 10).
Retroviral vectors, viral production, and CB-NK cell transduction
Retroviral vectors encoding the constructs iC9.CARCD38.CD28-CD3ζ−2A-IL15 (CAR38.3ζ.IL-15), iC9.CARCD38.CD28-CD3ζ (CAR38scFv.3ζ), iC9.CARCD38.CD28 (CAR38scFV), and iC9.CARCD5.CD28-CD3ζ−2A-IL15 (CAR5) were produced using 293T cells. In summary, 293T cells were seeded 24 h before transfection to achieve a confluency of 80% on the day of the experiment. A mixture containing Fugene, OptiMEM medium, and the CD38 CAR construct was incubated for 15 min at room temperature and then added to the 293T cell culture, which was kept in culture for 24 h. The following day, the medium containing the retrovirus was discarded, and fresh DMEM medium with 10% FBS and 1% glutamine was added. Forty-eight hours after transfection, the retrovirus was collected and frozen at −80°C.
The day before the CB-NK cell transduction, 24-well non-tissue culture plates were coated with Retronectin and incubated at 4°C overnight. On the day of the transduction, the Retronectin was discarded, DMEM medium was added, and plates were incubated for 15 min at 37°C. After incubation, the medium was discarded, and thawed retroviral supernatant was added to the wells. Plates were spun at 2000g for 2 h at 32°C. Following the spinning, retroviral supernatant was discarded. CB-NK cells at a concentration of 0.5 × 10E6 cells/ml were added to the wells with a medium enriched with IL-2 (200 U/ml). Plates were spun at 400g for 5 min at room temperature. These cells were then incubated for 48 h at 37°C. Transduction efficiency was measured by the expression of CAR on CB-NK cells 48–72 h post-transduction using an AF647 affinity purified fragment (ab’) goat anti-human antibody directed against the CH2-CH3 domain of the IgG hinge of the CAR followed by flow cytometric analysis. Between days 14 and 21 after isolation, CAR-transduced CB-NK cells were harvested for phenotypic and functional studies.
Multiplex cytokine assay for flow cytometry
CD38 CAR-transduced and NT NK cells were harvested and co-cultured in a 1:1 E:T ratio with MOLM14 cells for 16 h in a round-bottom 96-well plate. IL-2 (100 U/ml) was added to the culture. After incubation, supernatant from the culture was collected. Cytokine levels in the supernatant were assessed using the 13 multi-analyte LEGENDplex human CD8/NK panel kit according to the manufacturer’s instructions. Data were acquired immediately following the staining using a BD LSFortessa X-20 cell analyzer (BD Biosciences). Analysis was performed using FlowJo software (version 10).
Western blot for the detection of the CD3ζ domain of the CD38 CAR constructs
CB-NK cells were transduced with CD38 CAR constructs as described above. Forty-eight to 72 h after transduction, NK cells were harvested, washed, and 1×106 cells in 100 μL of PBS pelleted by centrifugation (13,000g for 1 min at 4°C) followed by mechanical lysis. Laemmli sample buffer was added to the cell suspension (200 μL), and the samples were homogenized. Samples were boiled for 10 min at 95°C, cooled on ice for 1 min, and centrifuged at 13,000g for 10 min at 4°C. After preparation, samples were loaded into and run in precast polyacrylamide NuPAGE Bis-Tris gels (Invitrogen). Electrotransfer was performed using the iBlot 2 transfer stacks and the iBlot 2 gel transfer device according to the manufacturer’s instructions (Invitrogen). After transfer, 5% skim milk in tris-buffered saline + Tween 20 (TBST) 0.1% was used to block the membrane for 1 h at room temperature. All washes and incubation steps were performed on a shaker. Rabbit anti-human CD3ζ antibody in 5% skim milk (1:1000 dilution) in TBST was added and incubated with the membrane overnight at 4°C. Following overnight incubation, the antibody mixture was discarded, and the membrane was washed three times with TBST for 10 min at room temperature. HRP-conjugated anti-rabbit IgG antibody in 5% skim milk (1:2000 dilution) was added and incubated with the membrane for 1 h at room temperature. The antibody mixture was discarded, and the membrane was washed three times with TBST. Clarity Western ECL substrate was prepared per the manufacturer’s instructions and used for chemiluminescence detection. Endogenous CD3ζ was used as the loading control.
IL-15 detection in 293T supernatants using ELISA
293T cells were transfected with the CD38 CAR constructs as previously described. Seventy-two hours after transfection, the supernatant was collected, and solid phase sandwich ELISA was performed using the human IL-15 quantikine ELISA kit (RnD) per the manufacturer’s instructions.
Conjugate formation assay by flow cytometry
CD38 CAR-transduced and NT NK cells were harvested and co-cultured in a 1:1 E:T ratio with target cells for 20 min at 37°C/5%CO2 in a round-bottom 96-well plate. After incubation, cells were harvested by gently pipetting, washed once with PBS, and centrifuged. After discarding supernatant, live/dead dye incubation and surface staining were performed as described above. For the conjugate formation assay against K562, NK cells were identified as CD56 positive and CD32 negative, while K562 cells were CD56 negative and CD32 positive. For conjugate formation assay against MOLM14, NK cells were identified as NKG2D positive and mKate2 negative, while MOLM14 cells were NKG2D negative and MKate2 positive. Doublets were not eliminated, and conjugates were identified by their double positivity. To preserve conjugates, vortexing was not performed. Data were acquired immediately following the staining from non-fixed samples using a BD LSFortessa X-20 cell analyzer (BD Biosciences). Analysis was performed using FlowJo software (version 10).
51Chromium (Cr) release assay
In summary, 0.5 × 106 target cells (T) were resuspended in 1 mL RPMI medium and a 100 μL of 51Cr was added for tumor labeling. Tumor cells were then incubated in this solution for 2 h in a humidified incubator at 37°C/5%CO2. After incubation, target cells were washed twice with PBS. After centrifugation, supernatant was discarded and the cells were resuspended in Click’s 50/RPMI 50 medium, followed by co-culture with NT or CAR-transduced NK cells as effector cells (E) for 4 h in v-bottomed 96-well plates at 37°C/5% CO2. Co-culture supernatant was then collected, and 51Cr release was measured. Conditions were tested in triplicates, and the means of the test wells were used for percent-specific lysis calculation. Percent-specific lysis was defined as [(mean of the test wells)−(mean of the spontaneous release wells)/(mean of maximal release wells)−(mean of the spontaneous release wells)] × 100.
To block the activation of LFA-1, we used the compound BIRT 377, an antagonist that keeps LFA-1 in its closed, inactive conformation.30 For these assays, NK cells were treated with either DMSO or BIRT 377 (20 μM in DMSO). Relative change in the killing was defined as [(% killing in the presence of DMSO CD38 CAR NK cell) − (% killing in the presence of BIRT 377 CD38 CAR NK cell)]/[(% killing in the presence of DMSO NT NK cell) − (% killing in the presence of BIRT 377 NT NK cell)]. These reagents were kept during the whole co-culture period. No cytokines were added.
Assessment of CD38 molecules on AML cells by flow cytometry
To quantify the number of CD38 molecules on target AML cells, the BD quantibrite PE Fluorescence quantitation kit was used as per manufacturer’s instructions. In summary, AML cells (1 × 106 cells) were harvested, washed twice with PBS, centrifuged, and the supernatant was discarded. Single marker surface staining was performed by adding PE-conjugated anti-human CD38 antibody (clone HIT2) followed by a 20-min incubation at room temperature. Cells were washed and kept in PBS. Lyophilized tubes containing PE-conjugated beads were prepared and run in parallel with CD38-stained cells. Data were acquired immediately following the staining from non-fixed samples using a BD LSFortessa X-20 cell analyzer (BD Biosciences). Analysis was performed using FlowJo software (version 10).
QUANTIFICATIONS AND STATISTICAL ANALYSIS
Two-tailed Student’s t- or Mean-Whitney tests were used to compare quantitative variables. One-way or two-way repeated measures analysis of variance (two-way ANOVA) was used to compare samples and concentration or samples and longitudinal time points, followed by Bonferroni’s multiple comparisons. Statistical significance was assessed with the Prism 9.0 software (GraphPad Software, Inc.). Two-sided p values below 0.05 were considered to be significant.
Supplementary Material
Highlights.
CD226 and LFA-1 are essential for immune synapse formation between NK and AML cells
CAR38 compensates for the loss of CD226 to preserve NK cell activity against AML
CAR38 cooperates with LFA-1 to maintain NK cell cytotoxicity against AML cells
ACKNOWLEDGMENTS
This work was supported in part by the generous philanthropic contributions to The University of Texas MD Anderson Cancer Center Moon Shots Program and The Sally Cooper Murray Endowment; by grants (1 R01 CA211044-01, 5 P01CA148600-03, and R01 CA280827-01) from the National Institutes of Health (NIH), a Leukemia Specialized Program of Research Excellence (SPORE) grant (P50CA100632), and a grant (P30 CA016672) from the NIH to the MD Anderson Cancer Center. We thank the MD Anderson Cancer Center Flow Cytometry and Cellular Imaging Facility – North Campus for their assistance with mass cytometry assays. Y.L. is supported by CPRIT Research Training Award RP210028.
Footnotes
DECLARATION OF INTERESTS
P.B., Y.L., P.L., M.D., H.R., R.B., E.L., E.J.S., K.R., and The University of Texas MD Anderson Cancer Center have an institutional financial conflict of interest with Takeda Pharmaceutical. R.B., E.L., and K.R. have an institutional financial conflict of interest with Affimed GmbH. K.R. participates on the scientific advisory boards of Avenge Bio, Virogin Biotech, Navan Technologies, Caribou Biosciences, Bit Bio Limited, Replay Holdings, oNKo Innate, The Alliance for Cancer Gene Therapy ACGT, Innate Pharma, and Shinobi Therapeutics. K.R. is the scientific founder of Syena. E.J.S. has served on the scientific advisory boards of Adaptimmune, Axio, Celaid, FibroBiologics, Navan Technologies, New York Blood Center, and Novartis. M.D. participates on the scientific advisory board of Cellsbin.
SUPPLEMENTAL INFORMATION
Supplemental information can be found online at https://doi.org/10.1016/j.celrep.2024.115122.
REFERENCES
- 1.Shallis RM, Wang R, Davidoff A, Ma X, and Zeidan AM (2019). Epidemiology of acute myeloid leukemia: Recent progress and enduring challenges. Blood Rev. 36, 70–87. 10.1016/j.blre.2019.04.005. [DOI] [PubMed] [Google Scholar]
- 2.Breems DA, Van Putten WLJ, Huijgens PC, Ossenkoppele GJ, Verhoef GEG, Verdonck LF, Vellenga E, De Greef GE, Jacky E, Van der Lelie J, et al. (2005). Prognostic index for adult patients with acute myeloid leukemia in first relapse. J. Clin. Oncol. 23, 1969–1978. 10.1200/JCO.2005.06.027. [DOI] [PubMed] [Google Scholar]
- 3.Herberman RB, Nunn ME, and Lavrin DH (1975). Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic acid allogeneic tumors. I. Distribution of reactivity and specificity. Int. J. Cancer 16, 216–229. 10.1002/ijc.2910160204. [DOI] [PubMed] [Google Scholar]
- 4.Morvan MG, and Lanier LL (2016). NK cells and cancer: you can teach innate cells new tricks. Nat. Rev. Cancer 16, 7–19. 10.1038/nrc.2015.5. [DOI] [PubMed] [Google Scholar]
- 5.Shimasaki N, Jain A, and Campana D (2020). NK cells for cancer immunotherapy. Nat. Rev. Drug Discov. 19, 200–218. 10.1038/s41573-019-0052-1. [DOI] [PubMed] [Google Scholar]
- 6.Shibuya A, Campbell D, Hannum C, Yssel H, Franz-Bacon K, McClanahan T, Kitamura T, Nicholl J, Sutherland GR, Lanier LL, and Phillips JH (1996). DNAM-1, a novel adhesion molecule involved in the cytolytic function of T lymphocytes. Immunity 4, 573–581. 10.1016/s1074-7613(00)70060-4. [DOI] [PubMed] [Google Scholar]
- 7.El-Sherbiny YM, Meade JL, Holmes TD, McGonagle D, Mackie SL, Morgan AW, Cook G, Feyler S, Richards SJ, Davies FE, et al. (2007). The requirement for DNAM-1, NKG2D, and NKp46 in the natural killer cell-mediated killing of myeloma cells. Cancer Res. 67, 8444–8449. 10.1158/0008-5472.CAN-06-4230. [DOI] [PubMed] [Google Scholar]
- 8.Bottino C, Castriconi R, Pende D, Rivera P, Nanni M, Carnemolla B, Cantoni C, Grassi J, Marcenaro S, Reymond N, et al. (2003). Identification of PVR (CD155) and Nectin-2 (CD112) as cell surface ligands for the human DNAM-1 (CD226) activating molecule. J. Exp. Med. 198, 557–567. 10.1084/jem.20030788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tahara-Hanaoka S, Shibuya K, Onoda Y, Zhang H, Yamazaki S, Miyamoto A, Honda SI, Lanier LL, and Shibuya A (2004). Functional characterization of DNAM-1 (CD226) interaction with its ligands PVR (CD155) and nectin-2 (PRR-2/CD112). Int. Immunol. 16, 533–538. 10.1093/intimm/dxh059. [DOI] [PubMed] [Google Scholar]
- 10.Hou S, Ge K, Zheng X, Wei H, Sun R, and Tian Z (2014). CD226 protein is involved in immune synapse formation and triggers Natural Killer (NK) cell activation via its first extracellular domain. J. Biol. Chem. 289, 6969–6977. 10.1074/jbc.M113.498253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Shibuya K, Lanier LL, Phillips JH, Ochs HD, Shimizu K, Nakayama E, Nakauchi H, and Shibuya A (1999). Physical and functional association of LFA-1 with DNAM-1 adhesion molecule. Immunity 11, 615–623. 10.1016/s1074-7613(00)80136-3. [DOI] [PubMed] [Google Scholar]
- 12.Castriconi R, Dondero A, Cantoni C, Della Chiesa M, Prato C, Nanni M, Fiorini M, Notarangelo L, Parolini S, Moretta L, et al. (2007). Functional characterization of natural killer cells in type I leukocyte adhesion deficiency. Blood 109, 4873–4881. 10.1182/blood-2006-08-038760. [DOI] [PubMed] [Google Scholar]
- 13.Pende D, Spaggiari GM, Marcenaro S, Martini S, Rivera P, Capobianco A, Falco M, Lanino E, Pierri I, Zambello R, et al. (2005). Analysis of the receptor-ligand interactions in the natural killer-mediated lysis of freshly isolated myeloid or lymphoblastic leukemias: evidence for the involvement of the Poliovirus receptor (CD155) and Nectin-2 (CD112). Blood 105, 2066–2073. 10.1182/blood-2004-09-3548. [DOI] [PubMed] [Google Scholar]
- 14.Carlsten M, Baumann BC, Simonsson M, Jädersten M, Forsblom AM, Hammarstedt C, Bryceson YT, Ljunggren HG, Hellström-Lindberg E, and Malmberg KJ (2010). Reduced DNAM-1 expression on bone marrow NK cells associated with impaired killing of CD34+ blasts in myelodysplastic syndrome. Leukemia 24, 1607–1616. 10.1038/leu.2010.149. [DOI] [PubMed] [Google Scholar]
- 15.Sanchez-Correa B, Gayoso I, Bergua JM, Casado JG, Morgado S, Solana R, and Tarazona R (2012). Decreased expression of DNAM-1 on NK cells from acute myeloid leukemia patients. Immunol. Cell Biol. 90, 109–115. 10.1038/icb.2011.15. [DOI] [PubMed] [Google Scholar]
- 16.June CH, and Sadelain M (2018). Chimeric Antigen Receptor Therapy. N. Engl. J. Med. 379, 64–73. 10.1056/NEJMra1706169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dong H, Ham JD, Hu G, Xie G, Vergara J, Liang Y, Ali A, Tarannum M, Donner H, Baginska J, et al. (2022). Memory-like NK cells armed with a neoepitope-specific CAR exhibit potent activity against NPM1 mutated acute myeloid leukemia. Proc. Natl. Acad. Sci. USA 119, e2122379119. 10.1073/pnas.2122379119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Caruso S, De Angelis B, Del Bufalo F, Ciccone R, Donsante S, Volpe G, Manni S, Guercio M, Pezzella M, Iaffaldano L, et al. (2022). Safe and effective off-the-shelf immunotherapy based on CAR.CD123-NK cells for the treatment of acute myeloid leukaemia. J. Hematol. Oncol. 15, 163. 10.1186/s13045-022-01376-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Albinger N, Pfeifer R, Nitsche M, Mertlitz S, Campe J, Stein K, Kreyenberg H, Schubert R, Quadflieg M, Schneider D, et al. (2022). Primary CD33-targeting CAR-NK cells for the treatment of acute myeloid leukemia. Blood Cancer J. 12, 61. 10.1038/s41408-022-00660-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gurney M, Stikvoort A, Nolan E, Kirkham-McCarthy L, Khoruzhenko S, Shivakumar R, Zweegman S, Van de Donk NWCJ, Mutis T, Szegezdi E, et al. (2022). CD38 knockout natural killer cells expressing an affinity optimized CD38 chimeric antigen receptor successfully target acute myeloid leukemia with reduced effector cell fratricide. Haematologica 107, 437–445. 10.3324/haematol.2020.271908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zeng AGX, Bansal S, Jin L, Mitchell A, Chen WC, Hussein AA, Chan-Seng-Yue M, Voisin V, van Galen P, Tierens A, et al. (2022). A cellular hyerarchy framework for understanding heterogeneity and predicting drug response in acute myeloid leukemia. Nat. Med. 28, 1212–1223. [DOI] [PubMed] [Google Scholar]
- 22.Conner M, Hance KW, Yadavilli S, Smothers J, and Waight JD (2022). Emergence of the CD226 Axis in Cancer Immunotherapy. Front. Immunol. 13, 914406. 10.3389/fimmu.2022.914406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Banta KL, Xu X, Chitre AS, Au-Yeung A, Takahashi C, O’Gorman WE, Wu TD, Mittman S, Cubas R, Comps-Agrar L, et al. (2022). Mechanistic convergence of the TIGIT and PD-1 inhibitory pathways necessitates co-blockade to optimize anti-tumor CD8(+) T cell responses. Immunity 55, 512–526.e9. 10.1016/j.immuni.2022.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mace EM, Dongre P, Hsu HT, Sinha P, James AM, Mann SS, Forbes LR, Watkin LB, and Orange JS (2014). Cell biological steps and checkpoints in accessing NK cell cytotoxicity. Immunol. Cell Biol. 92, 245–255. 10.1038/icb.2013.96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Comrie WA, Babich A, and Burkhardt JK (2015). F-actin flow drives affinity maturation and spatial organization of LFA-1 at the immunological synapse. J. Cell Biol. 208, 475–491. 10.1083/jcb.201406121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mukherjee M, Mace EM, Carisey AF, Ahmed N, and Orange JS (2017). Quantitative Imaging Approaches to Study the CAR Immunological Synapse. Mol. Ther. 25, 1757–1768. 10.1016/j.ymthe.2017.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mace EM, Zhang J, Siminovitch KA, and Takei F (2010). Elucidation of the integrin LFA-1-mediated signaling pathway of actin polarization in natural killer cells. Blood 116, 1272–1279. 10.1182/blood-2009-12-261487. [DOI] [PubMed] [Google Scholar]
- 28.Hogg N, Patzak I, and Willenbrock F (2011). The insider’s guide to leukocyte integrin signalling and function. Nat. Rev. Immunol. 11, 416–426. 10.1038/nri2986. [DOI] [PubMed] [Google Scholar]
- 29.Weulersse M, Asrir A, Pichler AC, Lemaitre L, Braun M, Carrié N, Joubert MV, Le Moine M, Do Souto L, Gaud G, et al. (2020). Eomes-Dependent Loss of the Co-activating Receptor CD226 Restrains CD8(+) T Cell Anti-tumor Functions and Limits the Efficacy of Cancer Immunotherapy. Immunity 53, 824–839.e10. 10.1016/j.immuni.2020.09.006. [DOI] [PubMed] [Google Scholar]
- 30.Schürpf T, and Springer TA (2011). Regulation of integrin affinity on cell surfaces. EMBO J. 30, 4712–4727. 10.1038/emboj.2011.333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Leick MB, Silva H, Scarfo I, Larson R, Choi BD, Bouffard AA, Gallagher K, Schmidts A, Bailey SR, Kann MC, et al. (2022). Non-cleavable hinge enhances avidity and expansion of CAR-T cells for acute myeloid leukemia. Cancer Cell 40, 494–508.e495. 10.1016/j.ccell.2022.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Barber DF, Faure M, and Long EO (2004). LFA-1 contributes an early signal for NK cell cytotoxicity. J. Immunol. 173, 3653–3659. 10.4049/jimmunol.173.6.3653. [DOI] [PubMed] [Google Scholar]
- 33.Bryceson YT, Ljunggren HG, and Long EO (2009). Minimal requirement for induction of natural cytotoxicity and intersection of activation signals by inhibitory receptors. Blood 114, 2657–2666. 10.1182/blood-2009-01-201632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Urlaub D, Höfer K, Müller ML, and Watzl C (2017). LFA-1 Activation in NK Cells and Their Subsets: Influence of Receptors, Maturation, and Cytokine Stimulation. J. Immunol. 198, 1944–1951. 10.4049/jimmunol.1601004. [DOI] [PubMed] [Google Scholar]
- 35.Junich Takagi BMP, Thomas W, and Timothy AS (2002). Global conformational rearrangements in integrin extracellular domains in out-in and inside-out signaling. Cell 110, 599. [DOI] [PubMed] [Google Scholar]
- 36.Nishida N, Xie C, Shimaoka M, Cheng Y, Walz T, and Springer TA (2006). Activation of leukocyte beta2 integrins by conversion from bent to extended conformations. Immunity 25, 583–594. 10.1016/j.immuni.2006.07.016. [DOI] [PubMed] [Google Scholar]
- 37.Vyas YM, Mehta KM, Morgan M, Maniar H, Butros L, Jung S, Burkhardt JK, and Dupont B (2001). Spatial organization of signal transduction molecules in the NK cell immune synapses during MHC class I-regulated noncytolytic and cytolytic interactions. J. Immunol. 167, 4358–4367. 10.4049/jimmunol.167.8.4358. [DOI] [PubMed] [Google Scholar]
- 38.Wulfing C, and Davis MM (1998). A receptor/cytoskeletal movement triggered by costimulation during T cell activation. Science 282, 2266–2269. 10.1126/science.282.5397.2266. [DOI] [PubMed] [Google Scholar]
- 39.Orange JS, Harris KE, Andzelm MM, Valter MM, Geha RS, and Strominger JL (2003). The mature activating natural killer cell immunologic synapse is formed in distinct stages. Proc. Natl. Acad. Sci. USA 100, 14151–14156. 10.1073/pnas.1835830100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Liu D, Bryceson YT, Meckel T, Vasiliver-Shamis G, Dustin ML, and Long EO (2009). Integrin-dependent organization and bidirectional vesicular traffic at cytotoxic immune synapses. Immunity 31, 99–109. 10.1016/j.immuni.2009.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wülfing C, Sjaastad MD, and Davis MM (1998). Visualizing the dynamics of T cell activation: intracellular adhesion molecule 1 migrates rapidly to the T cell/B cell interface and acts to sustain calcium levels. Proc. Natl. Acad. Sci. USA 95, 6302–6307. 10.1073/pnas.95.11.6302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Rak GD, Mace EM, Banerjee PP, Svitkina T, and Orange JS (2011). Natural killer cell lytic granule secretion occurs through a pervasive actin network at the immune synapse. PLoS Biol. 9, e1001151. 10.1371/journal.pbio.1001151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Phatarpekar PV, and Billadeau DD (2020). Molecular regulation of the plasma membrane-proximal cellular steps involved in NK cell cytolytic function. J. Cell Sci. 133, jcs240424. 10.1242/jcs.240424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Basu R, Whitlock BM, Husson J, Le Floc’h A, Jin W, Oyler-Yaniv A, Dotiwala F, Giannone G, Hivroz C, Biais N, et al. (2016). Cytotoxic T Cells Use Mechanical Force to Potentiate Target Cell Killing. Cell 165, 100–110. 10.1016/j.cell.2016.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Xiong W, Chen Y, Kang X, Chen Z, Zheng P, Hsu YH, Jang JH, Qin L, Liu H, Dotti G, and Liu D (2018). Immunological Synapse Predicts Effectiveness of Chimeric Antigen Receptor Cells. Mol. Ther. 26, 963–975. 10.1016/j.ymthe.2018.01.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Davenport AJ, Cross RS, Watson KA, Liao Y, Shi W, Prince HM, Beavis PA, Trapani JA, Kershaw MH, Ritchie DS, et al. (2018). Chimeric antigen receptor T cells form nonclassical and potent immune synapses driving rapid cytotoxicity. Proc. Natl. Acad. Sci. USA 115, E2068–E2076. 10.1073/pnas.1716266115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Viot J, Abdeljaoued S, Vienot A, Seffar E, Spehner L, Bouard A, Asgarov K, Pallandre JR, Renaude E, Klajer E, et al. (2023). CD8(+) CD226(high) T cells in liver metastases dictate the prognosis of colorectal cancer patients treated with chemotherapy and radical surgery. Cell. Mol. Immunol. 20, 365–378. 10.1038/s41423-023-00978-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Huang H, Huang Z, Ge J, Yang J, Chen J, Xu B, Wu S, Zheng X, Chen L, Zhang X, and Jiang J (2023). CD226 identifies functional CD8(+)T cells in the tumor microenvironment and predicts a better outcome for human gastric cancer. Front. Immunol. 14, 1150803. 10.3389/fimmu.2023.1150803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Briukhovetska D, Suarez-Gosalvez J, Voigt C, Markota A, Giannou AD, Schübel M, Jobst J, Zhang T, Dörr J, Märkl F, et al. (2023). T cell-derived interleukin-22 drives the expression of CD155 by cancer cells to suppress NK cell function and promote metastasis. Immunity 56, 143–161.e11. 10.1016/j.immuni.2022.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zhang Y, Zhao ZX, Gao JP, Huang YK, and Huang H (2023). Tumor-infiltrating CD226(+)CD8(+) T cells are associated with postoperative prognosis and adjuvant chemotherapeutic benefits in gastric cancer patients. J. Cancer Res. Clin. Oncol. 149, 4381–4389. 10.1007/s00432-022-04346-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Braun M, Aguilera AR, Sundarrajan A, Corvino D, Stannard K, Krumeich S, Das I, Lima LG, Meza Guzman LG, Li K, et al. (2020). CD155 on Tumor Cells Drives Resistance to Immunotherapy by Inducing the Degradation of the Activating Receptor CD226 in CD8(+) T Cells. Immunity 53, 805–823.e15. 10.1016/j.immuni.2020.09.010. [DOI] [PubMed] [Google Scholar]
- 52.Tan W, Liu M, Wang L, Guo Y, Wei C, Zhang S, Luo C, and Liu N (2021). Novel immune-related genes in the tumor microenvironment with prognostic value in breast cancer. BMC Cancer 21, 126. 10.1186/s12885-021-07837-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Epling-Burnette PK, Bai F, Painter JS, Rollison DE, Salih HR, Krusch M, Zou J, Ku E, Zhong B, Boulware D, et al. (2007). Reduced natural killer (NK) function associated with high-risk myelodysplastic syndrome (MDS) and reduced expression of activating NK receptors. Blood 109, 4816–4824. 10.1182/blood-2006-07-035519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kiladjian JJ, Bourgeois E, Lobe I, Braun T, Visentin G, Bourhis JH, Fenaux P, Chouaib S, and Caignard A (2006). Cytolytic function and survival of natural killer cells are severely altered in myelodysplastic syndromes. Leukemia 20, 463–470. 10.1038/sj.leu.2404080. [DOI] [PubMed] [Google Scholar]
- 55.Szczepanski MJ, Szajnik M, Welsh A, Foon KA, Whiteside TL, and Boyiadzis M (2010). Interleukin-15 enhances natural killer cell cytotoxicity in patients with acute myeloid leukemia by upregulating the activating NK cell receptors. Cancer Immunol. Immunother. 59, 73–79. 10.1007/s00262-009-0724-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Fauriat C, Just-Landi S, Mallet F, Arnoulet C, Sainty D, Olive D, and Costello RT (2007). Deficient expression of NCR in NK cells from acute myeloid leukemia: Evolution during leukemia treatment and impact of leukemia cells in NCRdull phenotype induction. Blood 109, 323–330. 10.1182/blood-2005-08-027979. [DOI] [PubMed] [Google Scholar]
- 57.Régis T, Costello SS, Marcenaro E, Lafage-Pochitaloff M, Mozziconacci M-J, Reviron D, Gastaut J-A, Pende D, Olive D, and Moretta A (2002). Defective expression and function of natural killer cell–triggering receptors in patients with acute myeloid leukemia. Blood 99, 3661–3667. [DOI] [PubMed] [Google Scholar]
- 58.Carlsten M, Norell H, Bryceson YT, Poschke I, Schedvins K, Ljunggren HG, Kiessling R, and Malmberg KJ (2009). Primary human tumor cells expressing CD155 impair tumor targeting by down-regulating DNAM-1 on NK cells. J. Immunol. 183, 4921–4930. 10.4049/jimmunol.0901226. [DOI] [PubMed] [Google Scholar]
- 59.Dominical V, Samsel L, and McCoy JP Jr. (2017). Masks in imaging flow cytometry. Methods 112, 9–17. 10.1016/j.ymeth.2016.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data reported in this paper will be shared by the lead contact upon request. This paper analyzes existing, publicly available data, accessible at TCGA (https://doi.org/10.1056/NEJMoa1301689), BeatAML (https://doi.org/10.1038/s41586-018-0623-z), and Leucegene (GEO: GSE67040).
This paper does not report original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.