

Abstract
The microcirculation exemplifies the mesoscale in physiological systems, bridging larger and smaller scale phenomena. Microcirculatory research represents an example of a ‘middle-out,’ rather than ‘top-down’ or ‘bottom-up,’ approach to the study of biological function. Computational and mathematical approaches can be used to analyse the functioning of the microcirculation and to establish quantitative relationships between microvascular processes and phenomena occurring on larger and smaller scales, leading to insights which could not be obtained solely by reductionist biological experiments. Given its integrative approach to processes occurring on disparate scales and its emphasis on theoretical as well as experimental approaches, microcirculatory research belongs within current definitions of systems biology.

Timothy W. Secomb (left) received his PhD in Applied Mathematics from the University of Cambridge. Since 1981, he has been at the University of Arizona, where he is Professor of Physiology and Mathematics. His research is on theoretical modeling of biological systems, with emphasis on the microcirculation. Axel R. Pries, MD, FESC (right) is Professor of Physiology and Director of the Institute for Physiology at the Charité Berlin. His main research interests are in the field of microcirculation including vascular adaptation and remodelling, endothelial function, microvascular networks and blood rheology, and combining intravital microscopy and molecular approaches with mathematical modelling.
Introduction
The term ‘systems biology’ came into frequent use around the year 2000 to describe efforts to synthesize and interpret the enormous amount of data generated by techniques of molecular biology, including the sequence of the human genome (Strange, 2005). While frequently understood to refer to the goal of understanding biological processes based on genomic, proteomic and molecular data, with an emphasis on networks of interacting cellular processes, systems biology can also be defined more broadly as ‘a comprehensive quantitative analysis of the manner in which all the components of a biological system interact functionally over time’ (Aderem, 2005). This definition recognizes that the goals of systems biology ultimately require integration of biological information at all structural levels from the molecule to the cell to the tissue to the whole organism. According to this definition, systems biology is in essence synonymous with physiology (Strange, 2005).
In some cases, the relationship between molecular-level phenomena and systems behaviour is direct. An example is the role of connexin-26 mutations in hereditary non-syndromic sensorineural deafness (Kelsell et al. 1997). However, this situation, as illustrated in Fig. 1 (‘Ideal’), is atypical. A more typical situation is that multiple biological entities and processes on each structural scale interact with processes occurring on larger and smaller scales, as indicated in Fig. 1 (‘Reality’). This implies that there is no unique ‘right’ level at which to start analysing biological systems. Both ‘bottom-up’ and ‘top-down’ approaches have limitations. For example, knowledge of the molecular basis of cardiac muscle contraction does not by itself allow prediction of the heart's pumping efficiency, which depends critically on large scale structural features. On the other hand, some top-down approaches to cardiac mechanics utilize phenomenological descriptions of muscle contraction, which may not adequately reflect the actual muscle biophysics. A ‘middle-out’ approach, which starts at an intermediate level of scale and reaches out to link with larger and smaller scale phenomena, may be advantageous (Noble, 2006). Such an approach to cardiac mechanics might for instance focus initially on the mechanical properties and arrangement of muscle fibres in the myocardium.
Figure 1. Schematic illustration of the relationship between the biological phenomena occurring at multiple scales.
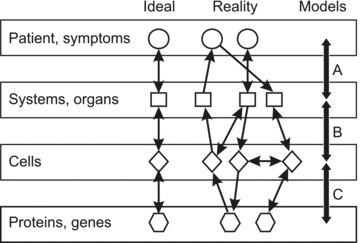
In some cases, a direct one-to-one link can be established between molecular information and functions or diseases (‘Ideal’). In general, however, interactions between biological processes and mechanisms at a given level of scale or organization influence events occurring on smaller and larger scales (‘Reality’). Theoretical models provide a framework for integrating information within and across biological scales (‘Models’). For example, classical cardiovascular system models can be used to relate the behaviour of the heart and blood vessels to systemic parameters such as blood pressure (A). In neurobiology, theoretical models to predict behaviour of populations of neurons based on the properties of individual neurons are well advanced (B). The development of models for genetic regulatory networks is currently an active field (C). From Secomb et al. (2008), reproduced by permission of John Wiley and Sons.
For such complex systems, intuitive or qualitative approaches are often insufficient for gaining an integrated understanding of their operation. Biological systems frequently involve integration of multiple inputs and contain feedback loops, so that the system's behaviour is determined by the balance between several competing factors. In a qualitative description of such a system, the relative importance of each factor is not known and the overall behaviour may therefore be unpredictable. Therefore, quantitative theoretical approaches are an essential and integral part of systems biology. They are particularly valuable in providing a framework that can be used to bridge the disparate scales of biological systems (Fig. 1, ‘Models’).
In the microcirculation, processes occurring at intermediate scales have direct interactions with phenomena occurring on larger and smaller scales. Microvascular functions such as vascular tone and local perfusion are determined by processes occurring at cellular and molecular levels, and the functional status of the microcirculation strongly influences tissue and organ behaviour. Conversely, systemic parameters such as blood pressure and fluid balance affect the function of the microcirculation, which in turn determines the environment in which cellular and molecular processes operate. Because the microcirculation involves a combination of physical processes, such as blood flow, blood vessel mechanics and diffusive mass transport, and biological processes, such as active cellular responses to physical and biochemical signals, research on the microcirculation requires an interdisciplinary approach. Physicists, engineers and mathematicians have naturally been drawn to the study of microcirculation, beginning almost a century ago (Krogh, 1919). Computational and mathematical approaches now have a well established role in the field (Secomb et al. 2008). As a field involving integration of phenomena across multiple scales and requiring theoretical as well as experimental approaches, investigation of the microcirculation has characteristics typical of systems biology in a broad sense. In the following paragraphs, we review several areas of research on the microcirculation from a systems biology perspective.
Structure of the microcirculation
The main function of the circulatory system is to transport materials by convection throughout the body. Molecular diffusion is effective only over very short distances. For instance, the diffusion distance of oxygen in oxygen-consuming tissue is typically less than 100 μm. Convection, i.e. transport by a flowing fluid, has no such limitation, and is utilized for mass transport by all but the smallest living organisms. Convection over large distances is most efficient if relatively large tubes are used, since resistance to fluid flow increases rapidly with decreasing tube diameter. However, the transport of substances to all parts of the organism requires a large number of small tubes finely distributed throughout the tissues. This is the microcirculation, generally understood to refer to the small branches of the circulatory system with diameters ranging from a few hundred micrometres down to a few micrometres (Pries & Secomb, 2008). The total number of capillaries in the human body is of the order of 109 or more. On the other hand, a single red blood cell within a capillary contains about 3 × 108 haemoglobin molecules. Therefore, research on the microcirculation is indeed a ‘middle-out’ approach to the circulatory system.
Microvessel networks have complex structures. In some tissues, the microcirculation may be approximated as a diverging tree of arterioles which feed the capillaries, which in turn drain into a converging tree of venules. An example is the mesentery, which, being a thin sheet, allows almost complete visualization of its microvascular structure. The microcirculation is often idealized as a set of vessels of different categories connected in series, where the parallel vessels in each category exhibit identical properties (Pries & Secomb, 2008). In reality, however, the structure is highly heterogeneous. For example, the generation number (number of branch points between the main feeding arteriole and a given vessel) of capillaries in mesentery networks varies between 2 and 25 (Pries et al. 1995), as illustrated in Fig. 2. As a consequence, the capillaries experience widely varying haemodynamic and functional conditions and cannot be regarded as a set of equivalent vessels. The same applies to arterioles and venules. This heterogeneity has important effects on the functional behaviour of microcirculation (Duling & Damon, 1987). Given the complexity of microvessel network structures and the resulting flow patterns, it may not be possible to analyse such effects adequately by qualitative arguments, and theoretical approaches using mathematical or computational models are essential (Secomb et al. 2008). An example is the occurrence of local tissue hypoxia, as discussed in the following section.
Figure 2. Computer-generated image of a microvascular network in rat mesentery, illustrating the characteristic structural heterogeneity of the microcirculation.

The diverging arteriolar and capillary network fed by the main feeding arteriole (A) is colour-coded according to the generation number, defined as the number of branch points between the main feeding arteriole and a particular vessel. All other vessels are assigned a dark blue colour. The main draining venule is labelled V. The network contains 546 distinct segments (between branch points). Generation numbers of terminal branches (capillaries) range from 3 to 21 in this network. Vessel diameters are doubled in the image, for clarity. The experimental methods used to obtain the structure were described previously (Pries et al. 1995).
Mass transport in the microcirculation
As already pointed out, the circulatory system exists primarily to meet requirements for mass transport. Of the many substances that must be distributed and removed, oxygen is one of the most critical. Many tissues, notably brain and heart, have a high and continuous demand for oxygen, and suffer damage rapidly if the supply fails. Because oxygen has a relatively low solubility in water and in tissues, ensuring an adequate oxygen supply places heavy demands on the system, including the necessity to bring oxygen by convection within a very small distance of all oxygen-consuming cells. Then, very steep gradients between blood and cells are generated, sufficient to drive adequate diffusive fluxes. As a consequence, oxygen levels in tissue are sensitively dependent on the microvascular architecture and flow distribution. For example, the maximal oxygen consumption rate that can be sustained by skeletal muscle is limited by capillary-level diffusive processes as well as by convective supply (Krogh, 1919; McGuire & Secomb, 2001). Heterogeneous vascular network structure can lead to regions of hypoxia even in otherwise well-oxygenated tissue. Such hypoxic regions may affect normal tissue function and compromise the effectiveness of therapy, as in the case of radiation treatment of tumours (Secomb et al. 1995). This is an example of the tissue-level consequences of microcirculation-scale phenomena. Available experimental approaches do not allow detailed mapping of tissue oxygen fields on the scale of micrometres, particularly in three dimensions, necessitating the use of theoretical approaches in this field of investigation.
Regulation of blood flow
The circulatory system must not only supply adequate blood flow to all regions of the body, but also control the flow according to requirements that vary with time and position on a variety of scales (Secomb, 2008). Moreover, the system is capable of wide variations in perfusion in blood flow in response to changing demands (metabolic regulation), while maintaining flow almost constant over a range of perfusion pressures (autoregulation) (Goodman et al. 1978; Johnson, 1986). To achieve this, the vascular system responds to many different stimuli, including flow-generated mechanical forces of pressure and shear stress on vessel walls, and levels of oxygen and other metabolites and vasoactive agents. On short time scales, the local control of blood flow is achieved through the contraction and relaxation of smooth muscle cells in the walls of arterioles. The endothelial cells and the smooth muscle cells are largely responsible for processing, integrating and reacting to these multiple stimuli. Axial cell–cell communication via gap junctions, particularly between endothelial cells, is a crucial component in the ability of the vasculature to respond in a coordinated way to changing demands (Segal, 2005).
Because flow in any given vessel depends on the behaviour of both upstream and downstream segments, the analysis of this system requires consideration of multiple interacting biological processes and feedback loops within a network structure, as illustrated in Fig. 3 (Arciero et al. 2008). The understanding of blood flow regulation presents challenges that are analogous to those involved in studying cell signalling processes in single cells and in multi-cell structures. Development of theories and models in which systems biology approaches to cell signalling are integrated with network-level analyses of blood flow regulations represents a logical direction for future development.
Figure 3. Schematic diagram illustrating interactions between factors involved in blood flow regulation.

Lines with arrowheads show positive effects and blunt-ended lines show negative effects. Heavy lines show the primary mechanism of metabolic regulation. Pathway a indicates vasodilatation invoked by increased metabolic demand. Pathways b and c indicate secondary effects resulting from increased diameter, causing decreased wall shear stress and increased wall tension. Both of these effects tend to increase arteriolar smooth muscle tone, counteracting the metabolic vasodilatation. Vertical arrows show effects of increasing metabolic demand in the presence of metabolic, shear-dependent and myogenic responses. (+) and (−) indicate the increase or decrease in tone, respectively, generated by each mechanism as a result of increased metabolic demand. A theoretical model based on this scheme was used to obtain quantitative estimates of the effects of these mechanisms on flow regulation. From Arciero et al. (2008), reproduced by permission of the American Physiological Society.
Structural adaptation of the vasculature and angiogenesis
Over time scales of hours, days and longer, the vascular system undergoes structural changes including growth of new vessels and remodelling or regression of existing vessels. These processes respond to changing demands during growth and development, normal physiological fluctuations, and disease processes. As in the case of acute flow regulation, the system must respond to multiple signals and stimuli, within the context of a complex network structure where changes in flow distribution require coordinated responses of multiple vessels forming flow pathways.
Early efforts at understanding the design of the vascular system were based on the concept of optimality (Murray, 1926). In this approach, the ‘cost’ of a blood vessel is assumed to consist of the work required to drive blood through the vessel, plus a term proportional to blood volume which represents the cost of maintaining the blood. The minimization of cost for a network of vessels then requires that the flows are proportional to the cubes of the diameters, a condition known as Murray's law. From haemodynamic considerations, it can be shown that this is approximately satisfied if wall shear stress is uniform throughout the network. Endothelial cells respond to changes in wall shear stress, and hence Murray's law could be satisfied if the endothelial cells sense levels of shear stress and drive changes in vessel diameters so as to achieve a ‘set point’ level of wall stress in each segment.
This concept has been influential, but as pointed out by Rodbard (1975), such a mechanism leads to instability of parallel flow pathways and cannot therefore account for actual network structures. Progress has come from theoretical approaches aimed at understanding how responses to multiple interacting stimuli combine to produce functionally adequate network structures that are consistent with experimental observations. Pries et al. (1998) developed a model in which responses to wall shear stress, intravascular pressure, local metabolic conditions, and signals propagated along vessel walls determine the diameters of vascular segments. In further development of this approach, a model containing multiple feedback loops (Fig. 4) was shown to be capable of predicting vessel wall thickness in addition to diameter (Pries et al. 2005).
Figure 4. Schematic diagram illustrating interactions between factors involved in structural adaptation of blood vessel diameter and wall thickness.
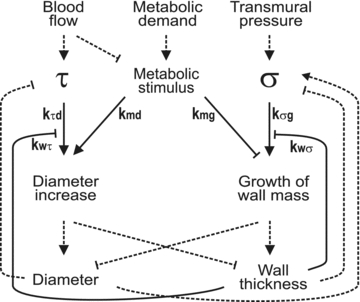
Structural adaptation occurs in response to the haemodynamic forces of wall shear stress (τ) and circumferential wall stress (σ), and a metabolic stimulus, dependent on local oxygen level. Dashed lines indicate relationships dictated by physical laws. Continuous lines indicate biological reactions and associated coefficients show the strengths of these reactions. For example, kτd and kmd quantify the effects of wall shear stress and the metabolic stimulus on diameter increase. The coefficient kwτ represents a reduction in sensitivity of vessel diameter change to wall shear stress, with increasing wall thickness. Lines with arrowheads show positive effects and blunt-ended lines show negative effects. A theoretical model based on this scheme predicts distributions of vessel diameters and ratios of vessel diameter to wall thickness consistent with experimental observations and allows estimation of the various coefficients. This model was found to be minimal in the sense that satisfactory agreement with experimental observations could not be obtained if any of the indicated effects were omitted. From Pries et al. (2005), reproduced by permission of Wolters Kluwer Health.
Angiogenesis, the growth of new vessels, has received great attention both for its relevance to normal physiological processes and for its potential as a target for therapeutic approaches. While much work has focused on molecular targets, the success of pro- or anti-angiogenic approaches depends on their effects on network structure and flow distribution. For example, microvascular density is not a good indicator of therapeutic efficacy (Kerbel & Folkman, 2002). Redistribution of flows within a network of given structure can have significant effects on function (Pries et al. 2010). For growth factors including VEGF (vascular endothelial growth factors), interactions between microvascular-level transport processes and signalling processes must be considered (Mac Gabhann & Popel, 2008). Therefore, progress in understanding and controlling angiogenesis requires multi-scale approaches, combining theoretical and experimental methods (Peirce, 2008).
Conclusions
The behaviours of biological systems emerge as a result of events occurring on a wide range of physical scales, from molecular to the whole body. Progress in understanding normal functions and diseases requires consideration of all these scales and their interactions. The microcirculation represents a mesoscale in the circulatory system, bridging smaller and larger scale phenomena. Theoretical and computational approaches are well established complements to experimental studies of microcirculation. Microcirculatory research provides an example and perhaps a paradigm for the development of systems biology.
Acknowledgments
This work was supported by NIH grants HL034555 and HL070657.
References
- Aderem A. Systems biology: Its practice and challenges. Cell. 2005;121:511–513. doi: 10.1016/j.cell.2005.04.020. [DOI] [PubMed] [Google Scholar]
- Arciero JC, Carlson BE, Secomb TW. Theoretical model of metabolic blood flow regulation: roles of ATP release by red blood cells and conducted responses. Am J Physiol Heart Circ Physiol. 2008;295:H1562–H1571. doi: 10.1152/ajpheart.00261.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duling BR, Damon DH. An examination of the measurement of flow heterogeneity in striated muscle. Circ Res. 1987;60:1–13. doi: 10.1161/01.res.60.1.1. [DOI] [PubMed] [Google Scholar]
- Goodman AH, Einstein R, Granger HJ. Effect of changing metabolic rate on local blood flow control in the canine hindlimb. Circ Res. 1978;43:769–776. doi: 10.1161/01.res.43.5.769. [DOI] [PubMed] [Google Scholar]
- Johnson PC. Autoregulation of blood flow. Circ Res. 1986;59:483–495. doi: 10.1161/01.res.59.5.483. [DOI] [PubMed] [Google Scholar]
- Kelsell DP, Dunlop J, Stevens HP, Lench NJ, Liang JN, Parry G, Mueller RF, Leigh IM. Connexin 26 mutations in hereditary non-syndromic sensorineural deafness. Nature. 1997;387:80–83. doi: 10.1038/387080a0. [DOI] [PubMed] [Google Scholar]
- Kerbel R, Folkman J. Clinical translation of angiogenesis inhibitors. Nat Rev Cancer. 2002;2:727–739. doi: 10.1038/nrc905. [DOI] [PubMed] [Google Scholar]
- Krogh A. The number and the distribution of capillaries in muscle with the calculation of the oxygen pressure necessary for supplying the tissue. J Physiol. 1919;52:409–515. doi: 10.1113/jphysiol.1919.sp001839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mac Gabhann F, Popel AS. Systems biology of vascular endothelial growth factors. Microcirculation. 2008;15:715–738. doi: 10.1080/10739680802095964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGuire BJ, Secomb TW. A theoretical model for oxygen transport in skeletal muscle under conditions of high oxygen demand. J Appl Physiol. 2001;91:2255–2265. doi: 10.1152/jappl.2001.91.5.2255. [DOI] [PubMed] [Google Scholar]
- Murray CD. The physiological principle of minimum work. I. The vascular system and the cost of blood volume. Proc Natl Acad Sci U S A. 1926;12:207–214. doi: 10.1073/pnas.12.3.207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noble D. The Music of Life. Biology Beyond Genes. Oxford: Oxford University Press; 2006. pp. 79–80. [Google Scholar]
- Peirce SM. Computational and mathematical modeling of angiogenesis. Microcirculation. 2008;15:739–751. doi: 10.1080/10739680802220331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pries AR, Hopfner M, le Noble F, Dewhirst MW, Secomb TW. The shunt problem: control of functional shunting in normal and tumour vasculature. Nat Rev Cancer. 2010;10:587–593. doi: 10.1038/nrc2895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pries AR, Reglin B, Secomb TW. Remodeling of blood vessels: responses of diameter and wall thickness to hemodynamic and metabolic stimuli. Hypertension. 2005;46:725–731. doi: 10.1161/01.HYP.0000184428.16429.be. [DOI] [PubMed] [Google Scholar]
- Pries AR, Secomb TW. Blood flow in microvascular networks. In: Tuma RF, Duran WN, Ley K, editors. Handbook of Physiology: section 2, The Cardiovascular System, vol. IV, Microcirculation. 2nd edn. San Diego: Academic Press; 2008. pp. 3–36. [Google Scholar]
- Pries AR, Secomb TW, Gaehtgens P. Structure and hemodynamics of microvascular networks: heterogeneity and correlations. Am J Physiol Heart Circ Physiol. 1995;269:H1713–H1722. doi: 10.1152/ajpheart.1995.269.5.H1713. [DOI] [PubMed] [Google Scholar]
- Pries AR, Secomb TW, Gaehtgens P. Structural adaptation and stability of microvascular networks: theory and simulations. Am J Physiol Heart Circ Physiol. 1998;275:H349–H360. doi: 10.1152/ajpheart.1998.275.2.H349. [DOI] [PubMed] [Google Scholar]
- Rodbard S. Vascular caliber. Cardiology. 1975;60:4–49. doi: 10.1159/000169701. [DOI] [PubMed] [Google Scholar]
- Secomb TW. Theoretical models for regulation of blood flow. Microcirculation. 2008;15:765–775. doi: 10.1080/10739680802350112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Secomb TW, Beard DA, Frisbee JC, Smith NP, Pries AR. The role of theoretical modeling in microcirculation research. Microcirculation. 2008;15:693–698. doi: 10.1080/10739680802349734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Secomb TW, Hsu R, Ong ET, Gross JF, Dewhirst MW. Analysis of the effects of oxygen supply and demand on hypoxic fraction in tumors. Acta Oncol. 1995;34:313–316. doi: 10.3109/02841869509093981. [DOI] [PubMed] [Google Scholar]
- Segal SS. Regulation of blood flow in the microcirculation. Microcirculation. 2005;12:33–45. doi: 10.1080/10739680590895028. [DOI] [PubMed] [Google Scholar]
- Strange K. The end of “naive reductionism”: rise of systems biology or renaissance of physiology? Am J Physiol Cell Physiol. 2005;288:C968–C974. doi: 10.1152/ajpcell.00598.2004. [DOI] [PubMed] [Google Scholar]