

Abstract
Hemibiotrophic phytopathogenic fungi cause devastating diseases in agronomically important crops. These fungal pathogens exploit a stealth bi-phasic infection strategy to colonize host plants. Their morphological and nutritional transition from biotrophy (characterized by voluminous intracellular primary hyphae) to necrotrophy (characterized by thin secondary hyphae) known as the biotrophy-necrotrophy switch (hemibiotrophy) is critical in symptom and disease development. To establish successful hemibiotrophic parasitism, pathogens likely secrete suites of proteins at the switch that constitute the biotrophy-necrotrophy switch secretome. To catalog such proteins, a directional cDNA library was constructed from mRNA isolated from infected Lens culinaris leaflet tissues displaying the switch of Colletotrichum truncatum, and 5,000 expressed sequence tags (ESTs) were generated. Four potential groups [hydrolytic enzymes, cell envelope-associated proteins (CEAPs), candidate effectors and proteins with diverse functions] were identified from pathogen-derived ESTs. Expression profiling of transcripts encoding CEAPs and candidate effectors in an infection time-course revealed that the majority of these transcripts were expressed or induced during the necrotrophic phase and repressed during the biotrophic phase of in planta colonization, indicating the massive accumulation of proteins at the switch. Taken together, our data suggest that the hemibiotrophic mode of fungal proliferation entails complex interactions of a pathogen with its host wherein the pathogen requires live host cells prior to switching to the necrotrophic phase. The microbial proteins employed during pathogenesis are likely to have defined roles at specific stages of pathogenesis.
Key words: hemibiotrophy, anthracnose, Colletotrichum truncatum, lentil, secretory proteins, effectors
Hemibiotrophic fungal phytopathogens (hereafter referred to as hemibiotrophs), pose a threat to world food supplies by causing notorious diseases like anthracnose and rice blast. Hemibiotrophs, such as Colletotrichum truncatum (causal agent of lentil anthracnose), C. higginsianum (causal agent of crucifer anthracnose) and Magnaporthe oryzae (causal agent of rice blast) exploit a sequential biotrophic- and necrotrophic-infection strategy to colonize their host plants. After the development of melanized appressoria from germinating conidia and breaching of the host cuticle and cell wall with the help of penetration pegs, the fungus initially grows biotrophically characterized by voluminous primary hyphae inside the lumen of living epidermal cells. Once biotrophic growth has been successfully established, the fungus switches to a necrotrophic phase associated with the production of thin secondary hyphae (Fig. 1A) that kill and macerate host tissues. The transition from the asymptomatic biotrophic phase to the symptomatic necrotrophic phase also known as the biotrophy-necrotrophy switch (BNS), is critical in symptom and disease development. Hemibiotrophs likely secrete repertoires of protein at this switch, collectively known as the BNS secretome. Some of these proteins may explicitly be transition specific, whereas others play a variety of roles in establishing successful hemibiotrophy. So far, only a few studies have addressed this critical BNS. The protein CLTA1 (C. lindemuthianum transcriptional activator 1) from C. lindemuthianum has been identified as a regulator of hemibiotrophic parasitism.1 Mutants harboring disrupted CLTA1 were unable to switch to the necrotrophic phase. This transcriptional factor is indispensable for pathogenicity on common bean. In a recently published paper, we constructed a directional cDNA plasmid library from mRNA isolated from infected lentil (Lens culinaris Medik.) leaflets displaying the BNS of C. truncatum and 5,000 ESTs were generated (Fig. 1B). One hundred sixty-two pathogen-derived ESTs (assembled into 122 tentative unigenes) were predicted to encode putative secretory proteins as they contained N-terminal signal peptides in their deduced open reading frames. Sequence analysis of unigenes revealed four potential groups, i.e., hydrolases (proteases and CWDEs), cell envelope-associated proteins, effectors and other proteins (Fig. 1C).2 Hemibiotrophs orchestrate a physiological switch from the asymptomatic biotrophic infection to the massive cell death and tissue maceration, presumably resulting from the coordinated secretion of cell-death elicitors and hydrolases (CWDEs, cell wall degrading enzymes).3 Fungal pathogens secrete a wide range of CWDEs capable of hydrolyzing host cell wall components (cellulose, hemicellulose, pectin and structural proteins) to accommodate a necrotrophic life style. The BNS secretome of C. truncatum contained pectinases and glycosyl hydrolases (glycosidases), which the fungal pathogens utilize to degrade the host plant cell wall for nutrition.2
Figure 1.
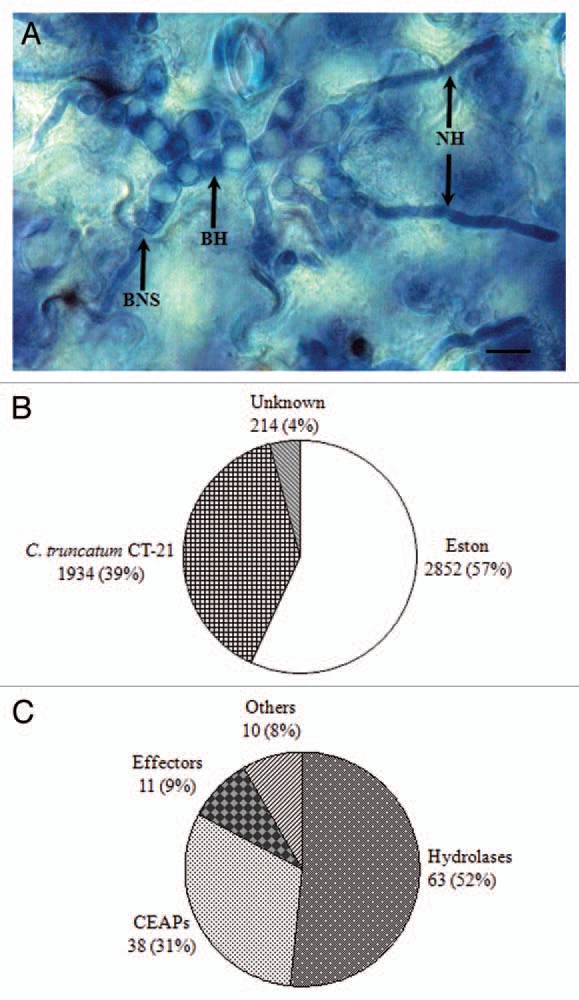
Colletotrichum truncatum biotrophy-necrotrophy switch library. (A) Trypan blue-stained in planta (Lens culinaris Medik. cv. Eston) fungal structures of C. truncatum. Excized lentil leaflets were droplet-inoculated with conidial suspension (1 × 104 conidia/ml) of C. truncatum and harvested at 48 h post-inoculation. BH, biotrophic hyphae; BNS, biotrophy-necrotrophy switch; NH, necrotrophic hyphae. Bar = 10 µM. (B) A directional cDNA library was constructed from mRNA isolated from lentil leaflet tissues displaying the switch from fat BH to thin NH and 5,000 ESTs were generated. Proportion of fungal and plant ESTs is illustrated. (C) One hundred sixty-two ESTs (assembled into 122 unique sequences or unigenes) were likely to encode proteins with putative signal peptide sequence at their N-terminus. Pie chart shows potential functional groups identified from unigenes encoding putative secretory proteins. CEAPs, cell envelope-associated proteins.
The outcome of host-pathogen interaction (resistance or susceptibility) is to a large extent governed by two classes of proteins: The first class of proteins that are secreted or localized to or at the cell membrane or cell wall of the pathogen, are known as CEAPs, and the second class of microbial proteins that are delivered into host plants are known as effectors. Some CEAPs lack a transmembrane domain and possess a C-terminal glycosyl-phosphatidylinositol (GPI) anchor. The majority of elicitin [extracellular proteins capable of inducing a hypersensitive response (HR) in tobacco] identified from Phytophthora spp. possess a GPI anchor.4 Expression profiling in an infection time-course revealed that the transcripts encoding GPI proteins were abrogated during the biotrophic phase and abruptly expressed during the necrotrophic phase, suggesting their potential role in eliciting cell death. Hemibiotrophs deliver effector proteins into the host plants to manipulate the host defense surveillance machinery in order to avert host defense responses, including the associated HR cell death during the biotrophic phase, and induce large scale necrosis during the necrotrophic phase. Similar to necrotrophic phytopathogens, hemibiotrophs also employ host selective toxins (secondary metabolites) to elicit plant cell death in order to derive nutrition from dead plant tissue.5 However, little is currently known about the implication of effector proteins in switching the life style of hemibiotrophs. Reports published to date have indicated that hemibiotrophs possess large repertoires of effector proteins. On the basis of their in planta functions, effectors can be classified into three distinctive groups: host defense suppressors, cell death elicitors and effectors with dual functions. Host defense suppressors subvert both layers of inducible host innate defense i.e., pathogen associated molecular pattern-triggered immunity or basal immunity and effector triggered immunity. Effector proteins that are explicitly meant to promote systemic cell death for facilitating the necrotrophic growth of a pathogen are known as elicitors or cell death inducers. In addition to suppressors and elicitors, hemibiotrophs also likely deliver certain effectors into the host milieu that may possess dual function. These effectors suppress the host defense responses and associated HR cell death during the biotrophic phase and promote necrosis during the necrotrophic phase. In addition to host defense manipulator, effectors are also shown to be involved in the penetration of host.6 We retrieved 11 candidate effectors from the hemibiotrophic secretome of C. truncatum on the basis of their small size (<300 amino acid residues), soluble nature (lacking transmembrane domain) and richness in cysteine residues (4–14). Among the functionally annotated effector proteins were lysin motif containing extracellular protein 6-like proteins, a cerato-platanin domain containing eliciting plant response-like protein (CP protein) and a HR inducing protein. Transient expression of genes encoding CP proteins from C. truncatum (Ct) and M. oryzae (Mo) in tobacco leaves using potato virus X-based agroinfiltration revealed that CtCP was unable to cause any micro- or macroscopic cell death, whereas MoCP (MgSM1),7 elicitated HR. This indicates a species-specific function of effector proteins.2
Several studies have pointed out that pathogens secrete effectors to avert the HR cell death during the biotrophic phase, and elicitors to promote necrosis during the necrotrophic phase. A HR averting effector protein, CgDN3 was identified from C. gloeosporioides. This effector is expressed during early stages of infection (infection vesicles), and mutants harboring the disrupted CgDN3 locus were unable to infect Stylosanthes guianensis. The ΔCgDN3 mutant elicited the localized hypersensitive like response at attempted infection sites, which prevented further in planta progress of the pathogen, suggesting its role as HR suppressor. Mutants were unable to grow biotrophically but could grow necrotrophically when conidia were inoculated directly onto wound sites.8 Kelley and colleagues (2010) identified a SNE1 suppressor of necrosis 1) protein from the fungus-like pathogen Phytophthora infestans, the causative agent of potato late blight that was specifically expressed during the biotrophic colonization of tomato. Transient expression of SNE1 in tobacco and tomato by agroinfiltration suppressed the HR induced by necrosis- and ethylene-inducing proteins like PiNPP1.1,9 from P. infestans and PsojNIP10 from P. sojae. The authors concluded that SNE1 proteins perturbed the activity of cell death elicitors secreted by P. infestans to promote the necrotrophic phase.3
In summary, it appears that hemibiotrophs are sophisticated manipulator of the host innate immunity. A picture is emerging suggesting that hemibiotrophs camouflage their conserved molecular signatures during the biotrophic phase to subvert HR cell death, and unmask them in the necrotrophic phase to induce large scale cell death and necrosis, but further experimental evidence is required to confirm this hypothesis.
Acknowledgments
This work was supported by NSERC-CRD, NSERC-Discovery and the Saskatchewan Pulse Growers grants to Drs. S.B., A.V. and Y.W.
Short communication to: Bhadauria V, Banniza S, Vandenberg A, Selvarai G, Wei Y. EST mining identifies proteins putatively secreted by the anthracnose pathogen Colletotrichum truncatum. BMC Genom. 2011;12:327. doi: 10.1186/1471-2164-12-327.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- 1.Dufresne M, Perfect S, Pellier AL, Bailey JA, Langin T. A GAL4-like protein is involved in the switch between biotrophic and necrotrophic phases of the infection process of Collectotrichum lindemuthianum on common bean. Plant Cell. 2000;12:1579–1590. doi: 10.1105/tpc.12.9.1579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bhadauria V, Banniza S, Vandenberg A, Selvaraj G, Wei Y. EST mining identifies proteins putatively secreted by the anthracnose pathogen Colletotrichum truncatum. BMC Genomics. 2011;12:327. doi: 10.1186/1471-2164-12-327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kelley BS, Lee SJ, Damasceno CMB, Chakravarthy S, Kim BD, Martin GB, et al. A secreted effector protein (SNE1) from Phytophthora infestans is a broadly acting suppressor of programmed cell death. Plant J. 2010;62:357–366. doi: 10.1111/j.1365-313X.2010.04160.x. [DOI] [PubMed] [Google Scholar]
- 4.Jiang RHY, Tyler BM, Whisson SC, Hardham AR, Govers F. Ancient origin of elicitin gene clusters in Phytophthora genomes. Mol Biol Evol. 2006;23:338–351. doi: 10.1093/molbev/msj039. [DOI] [PubMed] [Google Scholar]
- 5.Howlett BJ. Secondary metabolite toxins and nutrition of plant pathogenic fungi. Curr Opin Plant Biol. 2006;9:371–375. doi: 10.1016/j.pbi.2006.05.004. [DOI] [PubMed] [Google Scholar]
- 6.Doehlemann G, van der Linde K, Aßmann D, Schwammbach D, Hof A, Mohanty A, et al. Pep1, a secreted effector protein of Ustilago maydis, is required for successful invasion of plant cells. PLoS Pathog. 2009;5:1000290. doi: 10.1371/journal.ppat.1000290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yang Y, Zhang H, Li G, Li W, Wang X, Song F. Ectopic expression of MgSM1, a Cerato-platanin family protein from Magnaporthe grisea, confers broad-spectrum disease resistance in Arabidopsis. Plant Biotech J. 2009;7:763–777. doi: 10.1111/j.1467-7652.2009.00442.x. [DOI] [PubMed] [Google Scholar]
- 8.Stephenson SA, Hatfield J, Rusu AG, Maclean DJ, Manners JM. CgDN3: an essential pathogenicity gene of Colletotrichum gloeosporioides necessary to avert a hypersensitive-like response in the host Stylosanthes guianensis. Mol Plant-Microbe Interact. 2000;13:929–941. doi: 10.1094/MPMI.2000.13.9.929. [DOI] [PubMed] [Google Scholar]
- 9.Kanneganti TD, Huitema E, Cakir C, Kamoun S. Synergistic interactions of the plant cell death pathways induced by Phytophthora infestans Nep1-like protein PiNPP1.1 and INF1 elicitin. Mol Plant-Microbe Interact. 2006;19:854–863. doi: 10.1094/MPMI-19-0854. [DOI] [PubMed] [Google Scholar]
- 10.Qutob D, Kamoun S, Gijzen M. Expression of a Phytophthora sojae necrosis inducing protein occurs during transition from biotrophy to necrotrophy. Plant J. 2002;32:361–373. doi: 10.1046/j.1365-313x.2002.01439.x. [DOI] [PubMed] [Google Scholar]