

Abstract
Insect-derived baculoviruses have emerged as versatile and safe workhorses of biotechnology. Baculovirus expression vectors (BEVs) have been applied widely for crop and forest protection, as well as safe tools for recombinant protein production in insect cells. However, BEVs ability to efficiently transduce noninsect cells is still relatively poorly recognized despite the fact that efficient baculovirus-mediated in vitro and ex vivo gene delivery into dormant and dividing vertebrate cells of diverse origin has been described convincingly by many authors. Preliminary proof of therapeutic potential has also been established in preclinical studies. This review summarizes the advantages and current status of baculovirus-mediated gene delivery. Stem cell transduction, preclinical animal studies, tissue engineering, vaccination, cancer gene therapy, viral vector production, and drug discovery are covered.
Introduction
Gene therapy has matured to the point whereby the high potential of this therapeutic modality can now be realized for treating diseases with unmet needs, such as severe immune deficiencies,1,2 ocular diseases,3 and cancer.4 Several virus-derived vectors and nonvirus gene transfer agents have been used to address these applications. However, the safety, generation, immune response, duration of expression, and gene delivery capacity, still limits the use of these gene drugs.5 Thus, better vectors are still needed. Since first developed for overexpression of heterologous proteins,6 baculoviruses have been applied widely in biotechnology due to their versatility and the close resemblance between insect cell and mammalian cell protein posttranslational modification systems.7 Studies conducted over the last decade have also established the ability of baculoviruses to deliver genes into a wide range of species and types of vertebrate cells.7,8,9
The Baculoviridae are large, oblong-shaped (30–60 × 250–300 nm) enveloped particles with a circular, double-stranded DNA genome, approximately 80–180 kb, that naturally infect mainly Lepidoptera (butterflies and moth) larvae hosts.10 The family consists of about 700 known members of which the genome is characterized for almost 60 species.11 Baculoviruses are common in nature and in our food crops and are well known as pesticides and convenient tools for recombinant protein production in insect cells.6 No diseases have been linked to these viruses in any organism outside the phylum Arthropoda.12,13 Classically, baculoviruses were divided into two morphologically distinct genera: nuclear polyhedrosis viruses (NPVs) and granulosis viruses. However, baculovirus phylogeny follows the classification of the hosts more closely than the morphological traits, and a new classification divides baculoviruses into four genera: α baculovirus (lepidopteran-specific NPV), β baculovirus (lepidopteran-specific granulosis viruses), γ baculovirus (hymenopteran-specific NPV) and Δ baculovirus (dipteran-specific NPV).14 α baculoviruses are further divided into group I and II by their essential envelope glycoproteins gp64 and F-protein, respectively. NPVs obtain their envelope from the nuclear membrane and are occluded within a polyhedrin protein matrix, forming large (1–15 µm) polyhedral inclusion bodies. NPVs can be further distinguished on the basis of whether they contain in the polyhedral inclusion bodies a single nucleocapsid or multiple nucleocapsids. Unlike NPVs, granulosis viruses are embedded only as a single virion into a small inclusion body.11
Baculoviruses have many attractive features. They are easy to manipulate,11 able to carry large (at least 38 kbp)15 and multiple DNA inserts,16 and can be readily produced and purified at high titers.11 The inherent inability of baculoviruses to replicate in mammalian cells and low cytotoxicity make them potentially safe candidates for therapeutic gene delivery.12,13 Hepatocytes were the first noninsect origin cell type shown to be susceptible to baculovirus transduction in the mid 1990s.17,18 However, today it is evident, with only a few known exceptions, virtually all types and species of cells, including primary and stem cells, can be transduced efficiently.7,8,9,19 Baculovirus expression vectors (BEVs) can also be applied for the safe and efficient generation of induced pluripotent stem cells.20 Gene delivery efficacy can be boosted by enhancing vector design and optimizing transduction conditions.19 Translation of this technology to therapeutic use has begun and is supported not only by the availability of the latest generation vectors,21,22 but also by membrane technology-based virus purification protocols amenable to high-scale and good manufacturing practice quality virus generation.23,24 Baculovirus-mediated gene transfer has already demonstrated therapeutic efficacy in ex vivo and in vivo gene therapy studies.25,26,27,28 The approval of baculovirus insect cell produced human vaccine components (Cervarix, GlaxoSmithKline, Rixensart, Belgium and Provenge, Dendreon, Seattle, WA) by the European Medicines Agency and US Food and Drug Administration sets an important precedent for future regulatory approval29 of baculovirus-based gene therapy.
Autographa californica multiple nucleopolyhedrovirus (AcMNPV), often considered the prototypic baculovirus, has a circular double-stranded DNA genome (~134 kb) inside a rod-shaped nucleocapsid (25 × 260 nm).10 The genome of AcMNPV has been sequenced, whereas the structure of virion is only known by protein composition (no 3D structure available).30 The transgene capacity of AcMNPV in theory is unlimited as the capsid can freely extend.11 Multiple high-titer viruses can be generated and titered in parallel in a relatively short time frame in insect cells.11,31 Insect cells are easy to cultivate as adherent cells or in suspension and adapt well to growth in serum-free media.11 The recombinant viruses may be stored for long periods at 4 °C or within cryopreserved infected cells (titerless infected-cells preservation and scale-up).32,33 AcMNPV can be manipulated in biosafety level 1 facilities because it is strictly an insect pathogen and not known to cause disease in vertebrates.12,13
Progeny production of baculoviruses is biphasic (Figure 1).34 The budded virus (BuV) spreads the infection within the host and the occlusion-derived virus between hosts. The two forms of virus differ only in their envelopes. The envelope of BuV derives from the cell membrane, whereas the occlusion-derived virus derives its envelope from the nuclear membrane.35 The BuV form is the most widely used in biotechnology and enters insect and other host cells by endocytosis. The portal into mammalian cells has been shown recently to resemble phagocytosis more closely than pinocytosis.36,37,38,39 However, the receptor(s) or cell-signaling events mediating attachment and entry are largely unknown, and consensus as to the exact endocytic pathway(s) involved is lacking. Common cell surface ligands consisting of heparan sulphate proteoglycan, phospholipid, integrin, or lectin, are likely receptor candidates.9 Receptor binding and early steps of endocytosis are followed by an acid-induced fusion event, which releases the nucleocapsid into the cytoplasm.40,41,42 The capsids travel through the cytosol towards the nucleus by actin polymerization behind the nucleocapsid.43,44 AcMNPV equipped with a gene under the control of a promoter active in the target cell is able to transduce both nondividing and dividing mammalian cells40,45 which implies that nucleocapsid is able to transport the genome across the intact nuclear membrane through the nuclear pore complex.46 Indeed, electron microscopy has revealed that nucleocapsids dock at the nuclear pore complex and apparently intact DNA filled nucleocapsids localize to the nucleoplasmic space45,47 where the genome is released. However, the detailed molecular basis of transduction remains poorly understood in vertebrate cells and further studies are needed to resolve the pathway involved in efficient gene delivery.
Figure 1.

Life cycle and applications of baculoviruses. Baculoviruses have a biphasic life cycle in which two different forms of the virus spread through vertical and horizontal infection. Infection between hosts is mediated by virions occluded in PIBs late in infection (1). PIBs subsist in nature for years awaiting larva to ingest them (2). PIBs dissolve and release the ODVs only in the unique alkaline environment of the larval midgut (3). ODVs then penetrate the peritrophic membrane and infect epithelial cells by direct fusion of the virion and cell membrane (4). Nucleocapsids travel to the nucleus and initiate replication (5). To evade host defense mechanisms, some nucleocapsids traverse the cell directly to speed up the infection. The infected midgut cells produce the BuV form of the virus which spreads the infection within the larva through the tracheal system and hemolymph (6). The BuV can be used as a versatile tool for a plethora of biotechnological applications (7). BEVS, baculovirus expression vector system; BuV, budded virus; ODV, occlusion-derived virus; PIB, polyhedral inclusion body; PM, peritrophic membrane.
Gene Delivery Into Vertebrate Cells
The first strong evidence of AcMNPV's capability to enter a wide range of cells was gained while its safety for pest control was studied in the early 1980's.48 The experiments showed that many vertebrate cells internalized AcMNPV particles; however, no viral replication occurred. The lack of replication indicates that only the population of initial cells exposed to virus are transduced, with no cell-to-cell spread of the virus, and nontarget cells remaining unaffected. The early experiments were carried out with a wild-type virus lacking a vertebrate-compatible expression cassette. When Hofmann17 and Boyce,18 with their coworkers, in the mid 1990's outfitted AcMNPV with an expression cassette driving a marker gene via a strong viral promoter (active in the target cells), they could detect transgene expression, especially in hepatocytes. These pioneering studies boosted interest in baculoviruses as potential vectors for gene therapy. Many subsequent studies have confirmed the safety and feasibility of AcMNPV for efficient gene delivery in vertebrate cells, including cells other than those of hepatic origin.8,9,25 The list of permissive cells is extensive, and still expanding.7,9 It includes cell lines, primary cells, progenitor, and stem cells. Cells derived from human, monkey, porcine, rabbit, rat, feline, mouse, hamster, fish, avian, frog, and shrimp tissues have been transduced successfully.7,9 Many cell types can be transduced by outfitting the AcMNPV genome with a suitable expression cassette.49 These modified vectors are often referred to as BacMam viruses. In those instances in which the transduction efficiency is low or higher rates of transgene expression are desirable, modified viruses along with optimized transduction conditions can be used.19,50 Culture medium can have a role in transduction efficiency. Dulbecco's modified Eagle medium has been reported to support transduction poorly in some cell types and should be replaced with media such as RPMI1640, HyClone Classical Media, or Freestyle 293 Expression Medium.51,52,53 Transduction in phosphate-buffered saline buffer has also been shown to be effective with cell types which can tolerate short-term removal from complete medium.50 Transduction time is also important and virus treatment for at least 8 hours is recommended. Transient gene expression can also be extended by supertransduction20 or applying episomal replication and transposon systems, such as Sleeping Beauty or PiggyBac, within baculovirus hybrid viruses.54 The Woodchuck hepatitis virus posttranscriptional element provides significant benefit for baculovirus-mediated gene delivery by boosting substantially transgene expression.55 This can reach efficacy comparable to enhancement achievable with histone deacetylate inhibitors,56 such as a sodium butyrate, trichostatin A, or valproic acid, but without HDAI associated toxicity. Finally, the virus surface can be modified to augment AcMNPV effectiveness in gene delivery.57 Vesicular stomatitis virus G (VSV-G) transmembrane glycoprotein58,59 or surface display of its truncated transmembrane fragment (VSV-GED)60 are examples of successful pseudotyping. Surface display of avidin, biotin, and a lymphatic homing peptide or polymer coating with polyethyl glycol or polyethylenimine have also been used to improve the in vitro and in vivo transduction efficiency of AcMNPV.19,61
Preclinical Studies
AcMNPV is a promising vector not only for in vitro gene delivery, but also for ex vivo and in vivo gene delivery (Figure 2).9,25,26,27,28 The studies carried out in mice, rat, and rabbit models have revealed the central nervous system, eye (see Figure 2d–f), and testis as good targets (for a recent comprehensive review see).25 These targets represent immunoprivileged areas with an intrinsic ability to prevent the activation of adaptive and innate immune responses to antigens.61 Indeed, although pre-existing immunity does not restrict baculovirus-mediated gene delivery,62 both alternative and classical complement pathways are restrictive.63,64 This fact delayed the first successful in vivo applications of baculovirus-mediated gene transfer to the beginning of the new millennium,65,66 although the use of these viruses in an ex vivo context in mice was reported soon after the discovery that AcMNPV can efficiently transduce hepatocytes.67 Innate immunity-related problems can be avoided in ex vivo protocols, and promising results have been observed in cartilage and bone tissue engineering (see below). The low and transient immune response in the ex vivo context could also be useful in organ transplantation.68
Figure 2.
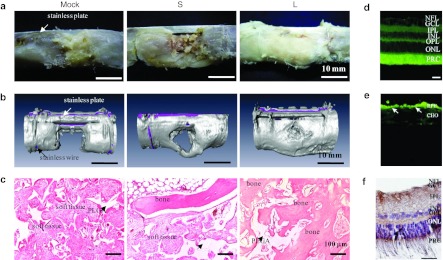
AcMNPV shows a high transduction efficacy ex vivo and in vivo. (a–c) ASCs genetically modified by the hybrid baculovirus ameliorate the healing of segmental bone defects in rabbits. The ASCs derived from the subcutaneous fat pad of NZW rabbits were transduced with hybrid baculovirus vectors conferring sustained expression of BMP-2 or VEGF, mixed with cells at a 4:1 ratio, loaded into cylindrical poly(l-lactide-co-glycolide) scaffolds (1.5 × 106 cells/scaffold) and transplanted into the critical-sized segmental defects at the femora of NZW rabbits (two scaffolds/defect, designated L group). The S group comprised PLGA scaffolds and ASCs that were transduced with the conventional baculoviruses transiently expressing BMP-2 or VEGF and transplanted in an identical manner. The mock group consisted of PLGA scaffolds and mock-transduced ASCs as the negative control. After 12 weeks transplantation, (a) gross appearance examination, (b) µCT analyses and hematoxylin & (c) eosin (H&E) staining collectively demonstrated that the L group (persistently expressing BMP-2 and VEGF) gave rise to significant new bone formation and bridging of bone defect in comparison with the S group (transiently expressing BMP-2 and VEGF) and mock group. (d–f) Intravitreal transduction leads to expression of (d,e) green fluorescent protein and (f) vascular endothelial growth factor D also in the deeper layers of a rabbit eye. The arrows in e indicate RPE layer. ASCs, Adipose-derived stem cells; BMP-2, bone morphogenetic protein-2; CHO, choroidea; GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; NFL, nerve fiber layer; NZW, New Zealand White; ONL, outer nuclear layer; OPL, outer plexiform layer; PLGA, poly(l-lactide-co-glycolide); PRC, photoreceptor cells; RPE, retinal pigment epithelium; VEGF, vascular endothelial growth factor. Scale bar = 20 µm. a and b were adapted from Lin et al.,95 c shows C.Y. Lin and Y.-C. Hu; unpublished data, and d–f were adapted from Kinnunen et al. (2009).80
Despite the complement-mediated inactivation of AcMNPV, a meaningful angiogenic response has recently been reported in immunocompetent rabbit skeletal muscle22 and rat heart69 after intramuscular and intramyocardial delivery, respectively. Conversely, the immunogenicity of AcMNPV can be taken advantage of in cancer therapy27 and vaccination (see below).26 Suppression of human epidermal-derived tumor growth in mice by programmed cell death gene delivery70 and induction of lung cancer and melanoma antitumor immunity using baculovirus-infected bone marrow-derived dendritic cells,71 represent the most recent examples. Other examples of preclinical efficacy include glioma treatment with p53 bearing AcMNPV in combination with sodium butyrate72 and the use of AcMNPV as an antiangiogenic agent in model systems to treat prostate and human ovarian cancer.54
To overcome complement-related problems, complement inhibitors including antibody to the complement component 5, cobra venom factor,63 a soluble complement receptor type 1,73,74 inhibitors for protease activating the complement, FUT-17558 and compstatin,64 and positively charged polyethylenimine75 have been used. Direct inoculation of baculovirus into various tissues such as rodent brain,58,66,76,77 rodent skeletal muscle,78 rodent testis,58 rodent eye,79 and rabbit eye80 or mouse cancerous tissue including melanoma, lung cancer, brain cancer,81 and hepatoma82 have been reported. In some cases, baculoviruses possessing the VSV-G protein showed significantly higher gene expression than the unmodified virus in mice.58,60,78 By using a silastic collar to avoid exposure to complement, rabbit carotid artery65 was also amenable to baculovirus transduction. Furthermore, baculoviruses bearing chimeric GP64 fused with CD55/decay-accelerating factor (which block complement) showed some resistance to complement inactivation.74 It has also been shown that GP64 can interact with decay-accelerating factor in a lipid raft and confer resistance to serum inactivation.83 More recently, recombinant baculovirus displaying several complement regulatory proteins (decay-accelerating factor, factor H-like protein-1, C4b binding protein, and membrane cofactor protein) fused with the membrane anchor of VSV-G protein induced lower levels of inflammatory cytokines interleukin 1β (IL-1β), IL-6, and IL-12p40 in macrophages resulting in attenuation of liver inflammation in mice.84 These results indicate that decay-accelerating factor display confers protection to baculovirus against complement inactivation and alleviates complement-mediated inflammation injury. Although baculoviruses bearing VSV-G protein have increased resistance to complement inactivation compared with the unmodified virus,58,84 GP64 bearing lentiviral vectors, including feline immunodeficiency virus, exhibited more resistant to human and mouse complements than those bearing VSV-G protein.85,86
Vaccination
Numerous efforts have been made to develop baculoviruses as viral vaccine vectors possessing adjuvant activity.26,87,88 One of the advantages of using a recombinant baculovirus to carry a foreign antigen is that this method allows efficient gene delivery into immune competent cells and confers protective immunity in vivo without severe cytotoxicity. The absence of pre-existing antibodies to baculovirus is also an advantage of using baculoviruses for foreign gene transduction in vivo.62 Although recombinant baculoviruses are capable of inducing antigen-specific humoral immune responses in mice, the induction of cellular immune responses are crucial for elimination of pathogens that cause chronic disease, such as hepatitis C virus and human immunodeficiency virus, because these viruses escape from humoral immune responses.89 Further studies are needed to improve induction of the cellular immune response by baculoviral vectors to confer sufficient protective immunity in vivo.
Ligands for pattern recognition receptors, including toll-like receptor (TLR), retinoic-acid-inducible protein I (RIG-I)-like receptor (RLR), and the nucleotide binding oligomerization domain, are capable of stimulating innate and adaptive immune cells and have been proposed as promising adjuvant candidates. It has previously been shown that intranasal inoculation of mice with AcMNPV induces protective immunity from a lethal challenge with influenza virus90 and that AcMNPV produces type I interferons in the immune cells of mice via a TLR-independent pathway.91 Recent studies have further confirmed that AcMNPV induces a host antiviral immune response through a TLR/RLR-independent pathway.92 These observations suggest that a novel innate immune signaling pathway in addition to the TLR/RLR signaling pathway may contribute to the induction of the host antiviral immune response induced by baculoviruses. Further studies are needed to clarify the precise mechanisms involved, such as that of the DNA-sensing machinery, for the induction of type I interferons. The ability of baculoviruses to induce innate immunity makes them promising candidates for future adjuvant-containing vaccine vehicles. Benefits of AcMNPV as an adjuvant are discussed below in the context of cancer vaccine therapy.
Baculoviral Transduction of Stem Cells
Advances in the identification, isolation, and derivation of human stem cells, especially the generation of human induced pluripotent stem cells, have raised great enthusiasm for regenerative medicine therapies. Among viral vectors capable of transducing stem cells, baculoviral vectors possess a unique feature in mediating high-level transient expression of transgenes, offering an attractive alternative to chemical or physical transfection methods and lentivirus-based transduction methods for stem cell genetic engineering.
As demonstrated with mesenchymal stem cells, baculoviral vectors can transduce multipotent stem cells obtained from different sources, including umbilical cord tissue,93 bone marrow,93,94 adipose tissue,95 and human embryonic stem cells (hESCs).96 Neural stem cells are another type of multipotent stem cells that can be effectively transduced by baculoviral vectors,97,98 in which baculovirus-mediated transgene expression may last for up to 3 months.97 Baculoviral vectors are also capable of transducing pluripotent stem cells such as hESCs99,100,101 and induced pluripotent stem cells.102,103 Currently, baculovirus transduction has been used to generate stem cells carrying therapeutic payloads of interest suitable for cartilage and bone tissue engineering and targeted glioma gene therapy. In these environments baculoviral transduction provides transgene expression from days to weeks fitting the needed therapeutic window. Baculovirus-mediated transient transduction may be useful as well in controlling the differentiation and stimulating the expansion of stem cells for medical applications.
In addition to providing transient transgene expression, baculoviral vectors can be tailored to permit stable transgene expression in stem cells. As baculovirus is a nonintegrating virus, a mechanism that promotes genomic integration needs to be incorporated into the vectors. In an initial study,99 the rep 78/68 genes and inverted terminal repeat sequences from adeno-associated virus (AAV) were incorporated into a baculoviral vector for hESC transduction to achieve Rep-mediated nonhomologous recombination at the AAVS1 locus of human chromosome 19q13.3-qter,104 a site with open chromatin structure and native insulators that may prevent transgene silencing. Although hESC lines with stable transgene expression can be generated with the hybrid AAV/baculovirus vector and the hESC lines maintain transgene expression during their prolonged self-renewal and differentiation processes, the study was unable to confirm site-specific transgene integration due to the low frequency of such events.99 A recent study has adopted a two-step process to target the AAVS1 locus in hESCs (Figure 3).101 Homologous recombination was first used to introduce heterospecific loxP sites into the locus, followed by baculoviral transduction to deliver Cre recombinase and a floxed transgene. The transgene can be efficiently and specifically inserted into the AAVS1 locus via recombinase-mediated cassette exchange, providing targeting efficiency of up to 100% in the master hESC line containing the loxP-docking sites. In another recent study, baculoviral vectors have been used to deliver zinc-finger nucleases together with a DNA donor template for targeting the CCR5 locus in hESCs,100 although with a relatively low targeting efficiency of 5%. The development of these site-specific integration methods has taken advantage of the large cloning capacity of baculoviral vectors that allows the inclusion of multiple genomic elements. These methods provide safer alternative technologies to modify stem cells for downstream therapeutic applications, for example deriving unlimited numbers of functionally enhanced or genetically corrected adult cells for cell replacement therapy.
Figure 3.

Cre recombinase-mediated cassette exchange in human embryonic stem cells (hESCs) by AcMNPV. Homologous recombination was used to introduce the EGFP gene and heterospecific loxP sites into the AAVS1 locus in hESCs. Two AcMNPV vectors, one to express Cre recombinase and another containing the mCherry gene driven by the EF1α promoter, were used for Cre recombinase-mediated cassette exchange. mCherry-positive cells were visible at week 1 and increased by mechanical selection at the time for normal hESC subculture. After three rounds of selection (week 5), almost the entire hESC colony was mCherry-positive. AcMNPV, Autographa californica multiple nucleopolyhedrovirus; EGFP, enhanced green fluorescent protein; hESCs, human embryonic stem cells; mCherry, red fluorescent protein. Scale bar = 200 µm. (unpublished data).
Baculovirus For Cartilage and Bone Tissue Engineering
Articular cartilage is a weight-bearing tissue that protects bones, but is limited in self-repair capacity. Hu and coworkers first demonstrated that baculovirus effectively transduced rat articular chondrocytes.105 However, chondrocytes tend to de-differentiate during subculture, thus hindering their in vitro expansion and subsequent transplantation back to the host. Sung et al. demonstrated that transduction of de-differentiated chondrocytes with a baculovirus vector (Bac-CB) expressing bone morphogenetic protein-2 (BMP-2) not only restores the differentiation status of passaged chondrocytes in vitro but also increased the cartilage-specific extracellular matrix accumulation.106 These activities were further augmented by baculovirus-mediated coexpression of transforming growth factor β.107 Moreover, when the Bac-CB–transduced chrondrocytes were seeded onto a polymeric scaffold and cultured dynamically for 3 weeks in a rotating-shaft bioreactor, the transduced cell/scaffold constructs grew into cartilage-like tissues.108 Eight weeks after such constructs were implanted into osteochondral defects of New Zealand White (NZW) rabbits, hyaline cartilages had regenerated and integrated well with adjacent host cartilage.109 These studies collectively demonstrated the potential of baculovirus in cartilage tissue engineering.
Baculovirus also efficiently transduces human bone marrow-derived mesenchymal stem cells (BMSCs)93 and progenitors originating from human BMSCs without obstructing their proliferation and differentiation potential. Transduction efficiencies can be elevated to ~95% under optimized conditions.110 As BMSCs can differentiate into osteoblasts and BMP-2 is a potent osteogenic factor, Hu and coworkers genetically engineered human BMSCs with Bac-CB and demonstrated that Bac-CB transduction directed osteogenesis of naïve BMSCs. After implantation, the transduced BMSCs induced ectopic bone formation in nude mice and promoted calvarial bone repair in immunocompetent rats.111
In addition to osteogenesis, massive segmental bone healing necessitates coordinated vascularization to rebuild blood vessels. Lin et al. recently constructed a recombinant baculovirus (Bac-CV) expressing vascular endothelial growth factor (VEGF).112 Implantation of Bac-CV– and Bac-CB–transduced rabbit BMSCs into critical-size segmental bone defects at the femora of NZW rabbits accelerated and ameliorated bone healing, thanks to the in vivo coexpression of BMP-2/VEGF and ensuing improved osteogenesis/angiogenesis.112
Adipose-derived stem cells (ASCs) are another promising cell source for bone regeneration but ASCs were suggested to be inferior to BMSCs in osteogenesis potential. To use ASCs for massive bone repair, Hu and coworkers hypothesized that sustained expression of genes promoting osteogenesis (BMP-2) and angiogenesis (VEGF) is necessary.95 Therefore, they used a dual baculovirus vector system constituting two baculoviruses: one expressing the flippase recombinase, whereas the other accommodating the BMP-2 or VEGF cassette flanked by the flippase recognition target (Frt) sequences. Cotransduction of mammalian cells with the hybrid vectors resulted in gene cassette excision of the baculovirus genome, formation of episomal circles, and prolonged transgene expression.110 Such flippase/Frt-mediated recombination occurred efficiently in the NZW rabbit ASCs, enabling persistent transgene expression for >28 days.95 Allotransplantation of the NZW rabbit ASCs transduced with the hybrid baculoviruses expressing BMP-2/VEGF into the critical-size femoral segmental defects accelerated the healing, improved the bone quality and angiogenesis when compared with transplanting ASCs engineered with the conventional baculoviruses (Figure 2a–c). The same technique was also exploited to promote the healing of critical-size calvarial defects in rabbits.113 These data altogether support the feasibility of using baculovirus for stem cell engineering and bone regeneration.95
The safety issues pertinent to the application of baculovirus-engineered stem cells were also addressed. It was uncovered that baculovirus transduction of BMSCs upregulated certain inflammatory genes (e.g., IL-1β, interferon α, and IL-6) and stimulated transient, low level upregulation of human leukocyte antigen I (HLA-I). However, baculovirus neither altered HLA-II expression nor impaired the immunosuppressive property of BMSCs. Microarray analysis further revealed that baculovirus perturbed the expression of 816 genes in BMSCs and activated the TLR3 pathway, leading to robust secretion of IL-6 and IL-8. Nonetheless, the induction was transient and no other inflammatory cytokines or interferon β were provoked. Furthermore, baculovirus transduction of BMSCs neither integrated the transgene into the host chromosomes nor disrupted the karyotype.114 These studies thus supported the safety of baculovirus-engineered BMSCs for cell therapy.114
Cancer Gene Therapy
Effective transduction and high-level transgene expression mediated by baculoviral vectors are especially suited for cancer gene therapy. Baculoviral vectors armed with suicide genes, tumor suppressor genes, proapoptotic genes, immunopotentiating genes, and antiangiogenesis genes have been tested in vivo in animal tumor models in many different anticancer regimens.27,54,115 Early studies investigated transcriptional targeting and transgene expression in directly transduced glioblastomas in the brain, an “immunoprivileged” site where complement-mediated inactivation is prevented by the blood–brain barrier. Transgenes under the control of a tissue-specific promoter, a tumor-selective promoter and a tissue-specific promoter and microRNA target sequences were evaluated (see for example, Figure 2d–f).27 More recently, ex-vivo gene therapy studies have been conducted using baculoviruses equipped with suicide genes to transduce stem cells.94,96,98 This approach benefits from the tropism exhibited by many adult stem cells for primary, solid, and metastatic tumors.
Cancer immunotherapy through vaccination with baculovirus-expressed tumor-associated antigens has been a promising area of research as well. For example, the majority of mice vaccinated with infected insect cells expressing colon carcinoma-specific antigen/major histocompatibility complex class I complexes were protected when challenged subsequently with colon tumor cells.116 Perhaps more surprising, vaccination with wild-type baculovirus devoid of any foreign genes elicited potent antimelanoma immunity equivalent to that achieved by five doses of immunostimulatory cytokine IL-12.82 This intrinsic immunostimulatory property of baculovirus was attributed primarily to natural killer cell proliferation and associated antitumor activity, but could also involve enhanced tumor-specific cytotoxic T lymphocyte responses and tumor-specific antibody production.71,117 Baculoviral transduction of antigen-presenting dendritic cells yielded promising results for melanoma therapy as well.118 Specifically, baculovirus transduction of dendritic cells stimulated the upregulation of CD1d (a nonclassical class I-like major histocompatibility complex molecule) that, in turn, induced a proinflammatory cytokine profile and enhanced the priming of naïve CD8+ T cells against a melanoma antigen.118
Assay Development and Drug Screening Applications
Viral-based gene delivery systems comprise an important part of the tool box for mammalian cell-based assay development and mechanism of action studies in drug discovery programs.119,120 Baculovirus-infected insects cells are used routinely for producing recombinant proteins for biochemical assay development and structural chemistry efforts.121,122 The development of versatile vector systems such as MultiBac have served to enhance the system for the production of multiprotein complexes.21 More recently, baculoviruses modified to express proteins in mammalian cells have also begun to fulfill an important role in drug discovery.123,124 In particular, drug discovery studies often require the development of robust, reproducible, readily transferable, and automatable cell-based assays for screening chemical libraries to identify starting points for chemical structure activity relationship studies. Historically, stable cell lines have been developed for this purpose. However, many attributes of BacMam-based gene delivery position this technology as an attractive alternative to stable cells and transient transfection methods. BacMam viruses can carry large DNA inserts, transduce a wide variety of mammalian cell types with little to no obvious cytotoxicity and are readily produced in insect cells. The viruses can be stored for long periods of time at 4 °C in the dark with little to no drop in virus titer. Transductions for cell-based assay applications are carried out simply by the addition of virus solution to cells grown either in suspension or attached culture. The transduced cells can be readily dispensed into microtiter culture plates for drug screening in automated drug screening facilities (Figure 4). BacMam viruses cannot replicate in mammalian cells, providing a unique low risk biosafety profile.13 The viruses can be used to support a variety of assay formats for different targets proteins, such as G protein coupled receptors,125,126 transporters,127,128 ion channels,129,130 histones,131 enzyme fragment complementation,132 and others. A particularly appealing aspect of using these viruses for developing assays requiring the expression of multiple proteins is that BacMam can be titrated reproducibly according to the amount of target protein desired simply by adding more or less virus.133,134
Figure 4.
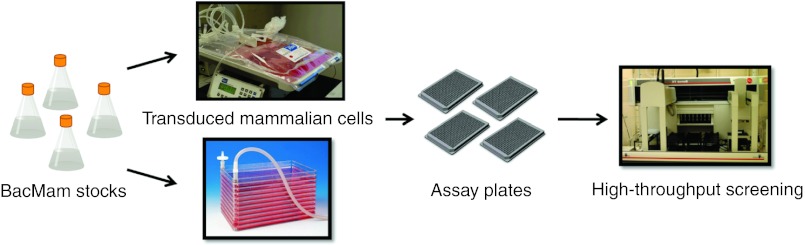
Application of BacMam transduction for assay development and automated high-throughput drug screening. Large quantities of either suspension or anchorage-dependent mammalian cells can be transduced by addition of single or multiple BacMam viruses at various titers. The transduced cells can be either dispensed directly using automated systems into 384 or 1536 well microtiter plates and subsequently incubated for 24–48 hours before assay or incubated for the desired period, harvested and stored frozen in liquid nitrogen for future use. In many instances this eliminates the need to develop stable cell lines for screening assays.
Two other important applications of BacMam gene delivery include the ability to launch viral infections and produce secreted proteins in mammalian cells. The delivery of hepatitis B virus genomes into hepatocytes has proven particularly useful for antiviral studies,135 and recently, the system has been used for the delivery of a Semliki Forest virus replicon136 and a human bocavirus.137 BacMam transduction of suspension culture cells has been shown to yield efficient expression of secreted proteases138 and membrane glycoproteins.139 Further developments in BacMam technology will continue to expand the use of this versatile gene delivery vector within drug discovery programs.
Large-Scale Generation of Other Gene Therapy Vectors
Kotin and coworkers have extended the versatility of the baculovirus insect cell expression system to include the production of faithfully assembled and packaged recombinant AAV (rAAV) vectors suitable for in vivo gene therapy applications.140 To achieve this goal, they developed a set of serotype-specific BEVs that produce stoichiometric amounts of the three AAV capsid proteins, VP1, VP2, and VP3 (Figure 5), which self-assemble within BEV-infected insect cells to form icosahedral virus-like particles. Additional BEVs were constructed that express the major AAV nonstructural genes, Rep78 and Rep52, which are essential for AAV genomic replication and packaging. When cap- and rep-expressing BEVs are introduced into insect cells along with a transgene of interest flanked by AAV inverted terminal repeat elements, efficient replication and encapsidation of linear, single-stranded AAV vector genomes occurs.140,141 The development of insect cell-based rAAV production was predicated on the unexpected finding that AAV vector genomes are efficiently replicated in lepidopteran insect cell lines, such as Sf9 and Sf21, in a Rep-dependent fashion.
Figure 5.

AAV and lentivirus generation by baculoviruses. (a) Schematic diagram of the AAV genome. The prototypical AAV-2 genome is 4.7 kb in length. Two major open reading frames encoded by the rep and cap genes are indicated. The protein coding sequences are flanked at each end of the genome by short (<0.15 kb) ITR elements, which provide cis-acting sequences necessary for replication and packaging of the viral genome. Three separate promoters (at map units 5, 19, and 40) give rise to a nested set of 3′ coterminal transcripts, which share a common polyadenylation signal. The translated portions of the Rep- and Cap-encoding mRNAs are indicated by cylinders. The “V-shape” indicates a differential splicing event. (b) Representation of AAV Rep and Cap coding sequences within a “second-generation” rAAV-producing BEV. (Right) The two major AAV nonstructural proteins, Rep78 and Rep52, are translated from a single species of mRNA via a leaky ribosomal scanning mechanism (see text). Rep78 proteins initiate at a CUG codon, whereas Rep52 proteins initiate at a downstream AUG codon. (Left) The AAV structural proteins, VP1, -2, and -3, are expressed from a single species of mRNA via leaky ribosomal scanning using a combination of AUG and ACG codons as indicated. RNAs are indicated by solid arrows. Expressed polypeptides are indicated by cylinders. VP, virion protein. (c) The constructs for the third generation self inactivating lentivirus production i.e., the transfer construct in which gene of interest is driven by hPKG promoter, the packing construct expressing HIV gag and pol driven by CMV promoter, the envelope construct providing the envelope G glycoprotein of the VSV-G, and HIV Rev under RSV promoter are cloned into AcMNPV genome and baculoviruses are generated in insect cells. Adherent 293T cells or suspension cultures are transduced by hybrid viruses to produce small or larger batches of LV, respectively. High-titer lentiviruses are collected from the medium and purified to homogeneity by ion-exchange chromatography. AcMNPV, Autographa californica multiple nucleopolyhedrovirus; BEV, baculovirus expression vector; BuV, the budded form of baculovirus; CMV, cytomegalovirus; HIV, human immunodeficiency virus; hPKG, human phosphoglycerate kinase; ITR, inverted terminal repeat; LTR, long terminal repeat; LV, Lentiviruses; P10, baculovirus p10 promoter; Ph, baculovirus polyhedrin promoter; RSV, Rous sarcoma virus; UAA, ocher stop codon; VSV-G, vesicular stomatitis virus G; WPRE, Woodchuck hepatitis virus posttranscriptional regulatory element.
Engineering of AAV transcription units was required for efficient baculovirus-mediated rAAV production in insect cells. During wild-type AAV infection, four nonstructural proteins (Rep78, -68, -52, and -40) are produced from the single open reading frame of the AAV rep gene via differential promoter selection and alternative splicing (Figure 5a). Rep78 and -68 participate in AAV genomic replication, whereas Rep52 and -40 have been implicated in packaging of progeny genomes.142,143 Recombinant AAV generation in insect cells requires production of at least one large (Rep78 or -68) and one small (Rep52 or -40) Rep protein, in addition to the three structural AAV capsid proteins. A strategy was developed to achieve expression of Rep78 and Rep52 from a single open reading frame without the need for an internal promoter element or splicing144 (Figure 5b). The AUG initiation codon of the full-length rep open reading frame was altered, along with its immediate flanking sequences, to yield a non-AUG codon presented in the context of a strong translation initiation motif (known as a Kozak consensus sequence). Internal AUG triplets intervening between the Rep78 and Rep52 initiation codons were changed to yield either silent mutations or conservative amino acid substitutions, depending on the reading frame. Intermittent translational initiation at the context-enhanced non-AUG triplet coupled with ribosomal scanning and translational initiation at the AUG codon of Rep52 resulted in efficient expression of both Rep78 and Rep52 from a single mRNA species. Combination of the translationally modified rep expression cassette with a similarly modified AAV coat protein expression cassette into a single recombinant baculovirus facilitated robust, scalable, high-titer production of rAAV in insect cells
Development of a baculovirus-mediated rAAV production method has facilitated large-scale preparation of rAAV vectors. Sf9 insect cells can be readily grown in serum-free suspension culture, thus providing a more cost-effective and easily scalable production platform than adherent mammalian cell-based strategies. In addition, delivery of the cis and trans elements needed for rAAV production via viral infection of producer cells is an efficient and robust process. However, the use of recombinant baculovirus seed stocks presents additional production issues, particularly upon scale-up. First, reliable titering of recombinant baculovirus stocks by plaque assay is a time consuming process. Second, recombinant baculovirus stocks loose infectivity upon prolonged storage in serum-free medium. These issues have been addressed by adapting a process for recombinant baculovirus-mediated protein production developed by Wasilko and colleagues,33 known as titerless infected-cells preservation and scale-up, that uses cryopreserved recombinant baculovirus-infected insect cells in batch production processes. Upon thawing, the number of infectious baculovirus particles released from viable rBEV-infected cells remains relatively unchanged, thus obviating the need for periodic plaque titering of cell-free baculovirus stocks. Inoculating Sf9 cell culture at a defined cell density with a predetermined optimal fractional volume of baculovirus-infected insect cells stocks produced consistent rAAV yields.145 An average yield of approximately 7 × 1013 purified, vector genome-containing rAAV particles per liter of batch culture has been reported for a wide range of process scales, resulting in total rAAV vector yields in excess of 1016 from 200 l scale bioreactor preparations.145 The baculovirus – Sf9 cell system has provided a scalable and economic method for large-scale rAAV production.
Lentiviruses (LVs) have become widely used for gene delivery and gene therapy when sustained gene expression is required.146 The popularity rests on the fact that lentiviruses show better safety and efficacy in gene delivery compared with conventional murine γ retroviruses.146,147 Preclinical and clinical applications, however, are still hampered by the difficulty of producing lentiviruses in sufficient quantity and purity.148 To answer this unmet need, BEVs was recently applied for lentivirus generation.52,53 In contrast to AAV production, human embryonic kidney cells (293T) were used for virus propagation instead of insect cells. The production relies on four novel hybrid baculoviruses (Figure 5c). The BAC-transfer virus expresses a transgene and BAC-gag-pol, BAC-VSVg, and BAC-rev viruses contain the elements required for safe production of the third generation lentiviruses.149 The results show that, in optimized production conditions, AcMNPV can successfully transduce both adherent and suspension cell cultures.52,53 High-titer LV stocks were readily produced and the generated virions performed similarly in vitro and in vivo as the viruses produced by conventional means. The same procedural problems related to seed stocks as with AAV concern baculovirus-based LV generation. However, the purification of LV from the contaminating baculoviruses is less cumbersome because baculoviruses do not replicate in vertebrate cells and they will be thus cleared after transduction by the producer cells. A scalable and cost-effective capture purification based on a diethylaminoethyl monolithic column enables 65% recovery of highly purified lentiviruses.52 BEVs are thus a promising platform for large scale LV production and further development and scale-up is in progress. The baculovirus system has also been exploited for AAV vector production in HEK-293 cells.150 Hu et al. have constructed a baculovirus system that comprises three vectors: Bac-LacZ carries the lacZ gene flanked by AAV inverted terminal repeats, Bac-RC harbors AAV rep and cap genes, and Bac-Helper carries helper genes derived from adenovirus. Cotransduction of HEK-293 cells with these three baculoviruses resulted in successful production of rAAV. Using a packed-bed reactor for cell culture and transduction processes significantly improved the AAV vector yield, thereby rendering this system an economically viable approach for AAV production.
Conclusion
Baculovirus technology has matured to the state in which it can be applied for numerous applications. Transduction of a plethora of mammalian cell types in addition to infection of insect cells establishes these viruses as a versatile gene delivery and protein expression platform. The first studies of therapeutic gene delivery in the context of immunoprivileged tissues, cancer, and ex vivo applications are encouraging and support further development of AcMNPV from preclinical applications to human studies. The application of baculovirus for producing AAV and lentiviruses facilitates the development of the gene therapy field from the bioprocess perspective. A deeper understanding of the molecular basis of target cell transduction and the antigenic properties of AcMNPV will help guide the development of improved vectors further enhancing the use of this unique and powerful gene delivery system.
Acknowledgments
R.H.S., R.M.K. funding was provided by the Division of Intramural Research of the National Heart, Lung, and Blood Institute (National Institutes of Health (NIH)). Additional funding was provided by the International Collaborative Effort (ICE) for Duchenne Muscular Dystrophy comprised of the French Duchenne Parent Project and the Association Monogasque contre les Myopathies. Portions of the technology described in this report are covered by United States and European patents assigned to the Secretary of the Department of Health and Human Services. A fraction of the licensing fees made to the NIH is distributed to the inventors in accordance with U.S. Government and NIH policy. The authors declared no conflict of interest.
References
- Boztug K, Schmidt M, Schwarzer A, Banerjee PP, Díez IA, Dewey RA.et al. (2010Stem-cell gene therapy for the Wiskott-Aldrich syndrome N Engl J Med 3631918–1927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hacein-Bey-Abina S, Hauer J, Lim A, Picard C, Wang GP, Berry CC.et al. (2010Efficacy of gene therapy for X-linked severe combined immunodeficiency N Engl J Med 363355–364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedrich MJ. Seeing is believing: gene therapy shows promise for ocular disorders. JAMA. 2010;304:1543–1545. doi: 10.1001/jama.2010.1412. [DOI] [PubMed] [Google Scholar]
- Immonen A, Vapalahti M, Tyynelä K, Hurskainen H, Sandmair A, Vanninen R.et al. (2004AdvHSV-tk gene therapy with intravenous ganciclovir improves survival in human malignant glioma: a randomised, controlled study Mol Ther 10967–972. [DOI] [PubMed] [Google Scholar]
- Giacca M., and, Zacchigna S. Virus-mediated gene delivery for human gene therapy. J Control Release. 2012;161:377–388. doi: 10.1016/j.jconrel.2012.04.008. [DOI] [PubMed] [Google Scholar]
- Summers MD. Milestones leading to the genetic engineering of baculoviruses as expression vector systems and viral pesticides. Adv Virus Res. 2006;68:3–73. doi: 10.1016/S0065-3527(06)68001-9. [DOI] [PubMed] [Google Scholar]
- Kost TA, Condreay JP., and, Jarvis DL. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat Biotechnol. 2005;23:567–575. doi: 10.1038/nbt1095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Airenne KJ, Mähönen AJ, Laitinen OH., and, Ylä-Herttuala S.2009Baculovirus-mediated gene transfer: An emerging universal concept Templeton NS.ed.). Gene and cell therapy: Therapeutic Mechanisms and Strategies, 3rd ednCRC Press: Boca Raton. 263–307.
- Chen CY, Lin CY, Chen GY., and, Hu YC. Baculovirus as a gene delivery vector: recent understandings of molecular alterations in transduced cells and latest applications. Biotechnol Adv. 2011;29:618–631. doi: 10.1016/j.biotechadv.2011.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theilmann DA, Blissard GW, Bonning B, Jehle J, O'Reilly DR, Rohrmann GF.et al. (2005Family Baculoviridae Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA.eds.). Virus Taxonomy: Eight Report of the International Committee on Taxonomy of Viruses. Elsevier: London; 177–185. [Google Scholar]
- O'Reilly DR, Miller LK., and, Luckov VA. Oxford University Press: New York; 1994. Baculovirus Expression Vectors. A Laboratory Manual. [Google Scholar]
- Burges HD, Croizier G., and, Huger J. A review of safety tests on baculoviruses. Entomaphaga. 1980;25:329–340. [Google Scholar]
- Kost TA., and, Condreay JP. Innovations-Biotechnology: Baculovirus vectors as gene transfer vectors for mammlian cells: Biosafety considerations. J Am Biol Safety Ass. 2002;7:167–169. [Google Scholar]
- Jehle JA, Blissard GW, Bonning BC, Cory JS, Herniou EA, Rohrmann GF.et al. (2006On the classification and nomenclature of baculoviruses: a proposal for revision Arch Virol 1511257–1266. [DOI] [PubMed] [Google Scholar]
- Cheshenko N, Krougliak N, Eisensmith RC., and, Krougliak VA. A novel system for the production of fully deleted adenovirus vectors that does not require helper adenovirus. Gene Ther. 2001;8:846–854. doi: 10.1038/sj.gt.3301459. [DOI] [PubMed] [Google Scholar]
- Bieniossek C, Imasaki T, Takagi Y., and, Berger I. MultiBac: expanding the research toolbox for multiprotein complexes. Trends Biochem Sci. 2012;37:49–57. doi: 10.1016/j.tibs.2011.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofmann C, Sandig V, Jennings G, Rudolph M, Schlag P., and, Strauss M. Efficient gene transfer into human hepatocytes by baculovirus vectors. Proc Natl Acad Sci USA. 1995;92:10099–10103. doi: 10.1073/pnas.92.22.10099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyce FM., and, Bucher NL. Baculovirus-mediated gene transfer into mammalian cells. Proc Natl Acad Sci USA. 1996;93:2348–2352. doi: 10.1073/pnas.93.6.2348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Airenne KJ. Optimization of baculovirus-mediated gene delivery into vertebrate cells. BioProcessing J. 2009;8:54–59. [Google Scholar]
- Takata Y, Kishine H, Sone T, Andoh T, Nozaki M, Poderycki M.et al. (2011Generation of iPS cells using a BacMam multigene expression system Cell Struct Funct 36209–222. [DOI] [PubMed] [Google Scholar]
- Airenne KJ, Peltomaa E, Hytönen VP, Laitinen OH., and, Ylä-Herttuala S. Improved generation of recombinant baculovirus genomes in Escherichia coli. Nucleic Acids Res. 2003;31:e101. doi: 10.1093/nar/gng102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heikura T, Nieminen T, Roschier MM, Karvinen H, Kaikkonen MU, Mähönen AJ.et al. (2012Baculovirus-mediated vascular endothelial growth factor-D(?N?C) gene transfer induces angiogenesis in rabbit skeletal muscle J Gene Med 1435–43. [DOI] [PubMed] [Google Scholar]
- Aucoin MG, Mena JA., and, Kamen AA. Bioprocessing of baculovirus vectors: a review. Curr Gene Ther. 2010;10:174–186. doi: 10.2174/156652310791321288. [DOI] [PubMed] [Google Scholar]
- Vicente T, Peixoto C, Carrondo MJ., and, Alves PM. Purification of recombinant baculoviruses for gene therapy using membrane processes. Gene Ther. 2009;16:766–775. doi: 10.1038/gt.2009.33. [DOI] [PubMed] [Google Scholar]
- Airenne KJ, Makkonen KE, Mähönen AJ., and, Ylä-Herttuala S. In vivo application and tracking of baculovirus. Curr Gene Ther. 2010;10:187–194. doi: 10.2174/156652310791321206. [DOI] [PubMed] [Google Scholar]
- Madhan S, Prabakaran M., and, Kwang J. Baculovirus as vaccine vectors. Curr Gene Ther. 2010;10:201–213. doi: 10.2174/156652310791321233. [DOI] [PubMed] [Google Scholar]
- Wang S., and, Balasundaram G. Potential cancer gene therapy by baculoviral transduction. Curr Gene Ther. 2010;10:214–225. doi: 10.2174/156652310791321251. [DOI] [PubMed] [Google Scholar]
- Lin CY, Lu CH, Luo WY, Chang YH, Sung LY, Chiu HY.et al. (2010Baculovirus as a gene delivery vector for cartilage and bone tissue engineering Curr Gene Ther 10242–254. [DOI] [PubMed] [Google Scholar]
- Lesch HP, Makkonen KE, Laitinen A, Määttä AM, Närvänen O, Airenne KJ.et al. (2011Requirements for baculoviruses for clinical gene therapy applications J Invertebr Pathol 107 SupplS106–S112. [DOI] [PubMed] [Google Scholar]
- Ayres MD, Howard SC, Kuzio J, Lopez-Ferber M., and, Possee RD. The complete DNA sequence of Autographa californica nuclear polyhedrosis virus. Virology. 1994;202:586–605. doi: 10.1006/viro.1994.1380. [DOI] [PubMed] [Google Scholar]
- Kärkkäinen HR, Lesch HP, Määttä AI, Toivanen PI, Mähönen AJ, Roschier MM.et al. (2009A 96-well format for a high-throughput baculovirus generation, fast titering and recombinant protein production in insect and mammalian cells BMC Res Notes 263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarvis DL., and, Garcia A., Jr Long-term stability of baculoviruses stored under various conditions. BioTechniques. 1994;16:508–513. [PubMed] [Google Scholar]
- Wasilko DJ., and, Lee SE. TIPS: Titerless infected-cells preservation and scale up. BioProcessing J. 2006;5:29–32. [Google Scholar]
- Federici BA.1997Baculovirus Pathogenesis Miller LK.ed.). The Baculoviruses, 1 edn Plenum Press: New York; 33–59. [Google Scholar]
- Funk CJ, Braunagel SC., and, Rohrmann GF.1997Baculovirus Structure Miller LK.ed.). The Baculoviruses, 1 edn Plenum Press: New York; 7–32. [Google Scholar]
- Laakkonen JP, Mäkelä AR, Kakkonen E, Turkki P, Kukkonen S, Peränen J.et al. (2009Clathrin-independent entry of baculovirus triggers uptake of E. coli in non-phagocytic human cells PLoS ONE 4e5093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matilainen H, Rinne J, Gilbert L, Marjomäki V, Reunanen H., and, Oker-Blom C. Baculovirus entry into human hepatoma cells. J Virol. 2005;79:15452–15459. doi: 10.1128/JVI.79.24.15452-15459.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Long G, Pan X, Kormelink R., and, Vlak JM. Functional entry of baculovirus into insect and mammalian cells is dependent on clathrin-mediated endocytosis. J Virol. 2006;80:8830–8833. doi: 10.1128/JVI.00880-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kataoka C, Kaname Y, Taguwa S, Abe T, Fukuhara T, Tani H.et al. (2012Baculovirus GP64-mediated entry into mammalian cells J Virol 862610–2620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Loo ND, Fortunati E, Ehlert E, Rabelink M, Grosveld F., and, Scholte BJ. Baculovirus infection of nondividing mammalian cells: mechanisms of entry and nuclear transport of capsids. J Virol. 2001;75:961–970. doi: 10.1128/JVI.75.2.961-970.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blissard GW. Baculovirus–insect cell interactions. Cytotechnology. 1996;20:73–93. doi: 10.1007/BF00350390. [DOI] [PubMed] [Google Scholar]
- Kukkonen SP, Airenne KJ, Marjomäki V, Laitinen OH, Lehtolainen P, Kankaanpää P.et al. (2003Baculovirus capsid display: a novel tool for transduction imaging Mol Ther 8853–862. [DOI] [PubMed] [Google Scholar]
- Ohkawa T, Volkman LE., and, Welch MD. Actin-based motility drives baculovirus transit to the nucleus and cell surface. J Cell Biol. 2010;190:187–195. doi: 10.1083/jcb.201001162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goley ED, Ohkawa T, Mancuso J, Woodruff JB, D'Alessio JA, Cande WZ.et al. (2006Dynamic nuclear actin assembly by Arp2/3 complex and a baculovirus WASP-like protein Science 314464–467. [DOI] [PubMed] [Google Scholar]
- Laakkonen JP, Kaikkonen MU, Ronkainen PH, Ihalainen TO, Niskanen EA, Häkkinen M.et al. (2008Baculovirus-mediated immediate-early gene expression and nuclear reorganization in human cells Cell Microbiol 10667–681. [DOI] [PubMed] [Google Scholar]
- Au S., and, Panté N. Nuclear transport of baculovirus: revealing the nuclear pore complex passage. J Struct Biol. 2012;177:90–98. doi: 10.1016/j.jsb.2011.11.006. [DOI] [PubMed] [Google Scholar]
- Salminen M, Airenne KJ, Rinnankoski R, Reimari J, Välilehto O, Rinne J.et al. (2005Improvement in nuclear entry and transgene expression of baculoviruses by disintegration of microtubules in human hepatocytes J Virol 792720–2728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkman LE., and, Goldsmith PA. In Vitro Survey of Autographa californica Nuclear Polyhedrosis Virus Interaction with Nontarget Vertebrate Host Cells. Appl Environ Microbiol. 1983;45:1085–1093. doi: 10.1128/aem.45.3.1085-1093.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spenger A, Ernst W, Condreay JP, Kost TA., and, Grabherr R. Influence of promoter choice and trichostatin A treatment on expression of baculovirus delivered genes in mammalian cells. Protein Expr Purif. 2004;38:17–23. doi: 10.1016/j.pep.2004.08.001. [DOI] [PubMed] [Google Scholar]
- Hsu CS, Ho YC, Wang KC., and, Hu YC. Investigation of optimal transduction conditions for baculovirus-mediated gene delivery into mammalian cells. Biotechnol Bioeng. 2004;88:42–51. doi: 10.1002/bit.20213. [DOI] [PubMed] [Google Scholar]
- Mähönen AJ, Makkonen KE, Laakkonen JP, Ihalainen TO, Kukkonen SP, Kaikkonen MU.et al. (2010Culture medium induced vimentin reorganization associates with enhanced baculovirus-mediated gene delivery J Biotechnol 145111–119. [DOI] [PubMed] [Google Scholar]
- Lesch HP, Laitinen A, Peixoto C, Vicente T, Makkonen KE, Laitinen L.et al. (2011Production and purification of lentiviral vectors generated in 293T suspension cells with baculoviral vectors Gene Ther 18531–538. [DOI] [PubMed] [Google Scholar]
- Lesch HP, Turpeinen S, Niskanen EA, Mähönen AJ, Airenne KJ., and, Ylä-Herttuala S. Generation of lentivirus vectors using recombinant baculoviruses. Gene Ther. 2008;15:1280–1286. doi: 10.1038/gt.2008.76. [DOI] [PubMed] [Google Scholar]
- Luo WY, Shih YS, Hung CL, Lo KW, Chiang CS, Lo WH.et al. (2012Development of the hybrid Sleeping Beauty: baculovirus vector for sustained gene expression and cancer therapy Gene Ther 19844–851. [DOI] [PubMed] [Google Scholar]
- Mähönen AJ, Airenne KJ, Purola S, Peltomaa E, Kaikkonen MU, Riekkinen MS.et al. (2007Post-transcriptional regulatory element boosts baculovirus-mediated gene expression in vertebrate cells J Biotechnol 1311–8. [DOI] [PubMed] [Google Scholar]
- Condreay JP, Witherspoon SM, Clay WC., and, Kost TA. Transient and stable gene expression in mammalian cells transduced with a recombinant baculovirus vector. Proc Natl Acad Sci USA. 1999;96:127–132. doi: 10.1073/pnas.96.1.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mäkelä AR., and, Oker-Blom C. The baculovirus display technology–an evolving instrument for molecular screening and drug delivery. Comb Chem High Throughput Screen. 2008;11:86–98. doi: 10.2174/138620708783744525. [DOI] [PubMed] [Google Scholar]
- Tani H, Limn CK, Yap CC, Onishi M, Nozaki M, Nishimune Y.et al. (2003In vitro and in vivo gene delivery by recombinant baculoviruses J Virol 779799–9808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barsoum J, Brown R, McKee M., and, Boyce FM. Efficient transduction of mammalian cells by a recombinant baculovirus having the vesicular stomatitis virus G glycoprotein. Hum Gene Ther. 1997;8:2011–2018. doi: 10.1089/hum.1997.8.17-2011. [DOI] [PubMed] [Google Scholar]
- Kaikkonen MU, Räty JK, Airenne KJ, Wirth T, Heikura T., and, Ylä-Herttuala S. Truncated vesicular stomatitis virus G protein improves baculovirus transduction efficiency in vitro and in vivo. Gene Ther. 2006;13:304–312. doi: 10.1038/sj.gt.3302657. [DOI] [PubMed] [Google Scholar]
- Kaikkonen MU, Ylä-Herttuala S., and, Airenne KJ. How to avoid complement attack in baculovirus-mediated gene delivery. J Invertebr Pathol. 2011;107 Suppl:S71–S79. doi: 10.1016/j.jip.2011.05.007. [DOI] [PubMed] [Google Scholar]
- Strauss R, Hüser A, Ni S, Tuve S, Kiviat N, Sow PS.et al. (2007Baculovirus-based vaccination vectors allow for efficient induction of immune responses against plasmodium falciparum circumsporozoite protein Mol Ther 15193–202. [DOI] [PubMed] [Google Scholar]
- Hofmann C., and, Strauss M. Baculovirus-mediated gene transfer in the presence of human serum or blood facilitated by inhibition of the complement system. Gene Ther. 1998;5:531–536. doi: 10.1038/sj.gt.3300607. [DOI] [PubMed] [Google Scholar]
- Georgopoulos LJ, Elgue G, Sanchez J, Dussupt V, Magotti P, Lambris JD.et al. (2009Preclinical evaluation of innate immunity to baculovirus gene therapy vectors in whole human blood Mol Immunol 462911–2917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Airenne KJ, Hiltunen MO, Turunen MP, Turunen AM, Laitinen OH, Kulomaa MS.et al. (2000Baculovirus-mediated periadventitial gene transfer to rabbit carotid artery Gene Ther 71499–1504. [DOI] [PubMed] [Google Scholar]
- Sarkis C, Serguera C, Petres S, Buchet D, Ridet JL, Edelman L.et al. (2000Efficient transduction of neural cells in vitro and in vivo by a baculovirus-derived vector Proc Natl Acad Sci USA 9714638–14643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandig V, Hofmann C, Steinert S, Jennings G, Schlag P., and, Strauss M. Gene transfer into hepatocytes and human liver tissue by baculovirus vectors. Hum Gene Ther. 1996;7:1937–1945. doi: 10.1089/hum.1996.7.16-1937. [DOI] [PubMed] [Google Scholar]
- Murguía-Meca F, Plata-Muñoz JJ, Hitchman RB, Danquah JO, Hughes D, Friend PJ.et al. (2011Baculovirus as delivery system for gene transfer during hypothermic organ preservation Transpl Int 24820–828. [DOI] [PubMed] [Google Scholar]
- Paul A, Binsalamah ZM, Khan AA, Abbasia S, Elias CB, Shum-Tim D.et al. (2011A nanobiohybrid complex of recombinant baculovirus and Tat/DNA nanoparticles for delivery of Ang-1 transgene in myocardial infarction therapy Biomaterials 328304–8318. [DOI] [PubMed] [Google Scholar]
- Kim YK, Kwon JT, Choi JY, Jiang HL, Arote R, Jere D.et al. (2010Suppression of tumor growth in xenograft model mice by programmed cell death 4 gene delivery using folate-PEG-baculovirus Cancer Gene Ther 17751–760. [DOI] [PubMed] [Google Scholar]
- Suzuki T, Oo Chang M, Kitajima M., and, Takaku H. Induction of antitumor immunity against mouse carcinoma by baculovirus-infected dendritic cells. Cell Mol Immunol. 2010;7:440–446. doi: 10.1038/cmi.2010.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo H, Choudhury Y, Yang J, Chen C, Tay FC, Lim TM.et al. (2011Antiglioma effects of combined use of a baculovirual vector expressing wild-type p53 and sodium butyrate J Gene Med 1326–36. [DOI] [PubMed] [Google Scholar]
- Hoare J, Waddington S, Thomas HC, Coutelle C., and, McGarvey MJ. Complement inhibition rescued mice allowing observation of transgene expression following intraportal delivery of baculovirus in mice. J Gene Med. 2005;7:325–333. doi: 10.1002/jgm.671. [DOI] [PubMed] [Google Scholar]
- Hofmann C, Hüser A, Lehnert W., and, Strauss M. Protection of baculovirus-vectors against complement-mediated inactivation by recombinant soluble complement receptor type 1. Biol Chem. 1999;380:393–395. doi: 10.1515/BC.1999.052. [DOI] [PubMed] [Google Scholar]
- Yang Y, Lo SL, Yang J, Yang J, Goh SS, Wu C.et al. (2009Polyethylenimine coating to produce serum-resistant baculoviral vectors for in vivo gene delivery Biomaterials 305767–5774. [DOI] [PubMed] [Google Scholar]
- Li Y, Wang X, Guo H., and, Wang S. Axonal transport of recombinant baculovirus vectors. Mol Ther. 2004;10:1121–1129. doi: 10.1016/j.ymthe.2004.08.018. [DOI] [PubMed] [Google Scholar]
- Lehtolainen P, Tyynelä K, Kannasto J, Airenne KJ., and, Ylä-Herttuala S. Baculoviruses exhibit restricted cell type specificity in rat brain: a comparison of baculovirus- and adenovirus-mediated intracerebral gene transfer in vivo. Gene Ther. 2002;9:1693–1699. doi: 10.1038/sj.gt.3301854. [DOI] [PubMed] [Google Scholar]
- Pieroni L, Maione D., and, La Monica N. In vivo gene transfer in mouse skeletal muscle mediated by baculovirus vectors. Hum Gene Ther. 2001;12:871–881. doi: 10.1089/104303401750195845. [DOI] [PubMed] [Google Scholar]
- Haeseleer F, Imanishi Y, Saperstein DA., and, Palczewski K. Gene transfer mediated by recombinant baculovirus into mouse eye. Invest Ophthalmol Vis Sci. 2001;42:3294–3300. [PMC free article] [PubMed] [Google Scholar]
- Kinnunen K, Kalesnykas G, Mähönen AJ, Laidinen S, Holma L, Heikura T.et al. (2009Baculovirus is an efficient vector for the transduction of the eye: comparison of baculovirus- and adenovirus-mediated intravitreal vascular endothelial growth factor D gene transfer in the rabbit eye J Gene Med 11382–389. [DOI] [PubMed] [Google Scholar]
- Kim CH, Yoon JS, Sohn HJ, Kim CK, Paik SY, Hong YK.et al. (2007Direct vaccination with pseudotype baculovirus expressing murine telomerase induces anti-tumor immunity comparable with RNA-electroporated dendritic cells in a murine glioma model Cancer Lett 250276–283. [DOI] [PubMed] [Google Scholar]
- Kitajima M, Abe T, Miyano-Kurosaki N, Taniguchi M, Nakayama T., and, Takaku H. Induction of natural killer cell-dependent antitumor immunity by the Autographa californica multiple nuclear polyhedrosis virus. Mol Ther. 2008;16:261–268. doi: 10.1038/sj.mt.6300364. [DOI] [PubMed] [Google Scholar]
- Kaname Y, Tani H, Kataoka C, Shiokawa M, Taguwa S, Abe T.et al. (2010Acquisition of complement resistance through incorporation of CD55/decay-accelerating factor into viral particles bearing baculovirus GP64 J Virol 843210–3219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaikkonen MU, Maatta AI, Ylä-Herttuala S., and, Airenne KJ. Screening of complement inhibitors: shielded baculoviruses increase the safety and efficacy of gene delivery. Mol Ther. 2010;18:987–992. doi: 10.1038/mt.2010.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schauber CA, Tuerk MJ, Pacheco CD, Escarpe PA., and, Veres G. Lentiviral vectors pseudotyped with baculovirus gp64 efficiently transduce mouse cells in vivo and show tropism restriction against hematopoietic cell types in vitro. Gene Ther. 2004;11:266–275. doi: 10.1038/sj.gt.3302170. [DOI] [PubMed] [Google Scholar]
- Sinn PL, Burnight ER, Hickey MA, Blissard GW., and, McCray PB., Jr Persistent gene expression in mouse nasal epithelia following feline immunodeficiency virus-based vector gene transfer. J Virol. 2005;79:12818–12827. doi: 10.1128/JVI.79.20.12818-12827.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu YC, Yao K., and, Wu TY. Baculovirus as an expression and/or delivery vehicle for vaccine antigens. Expert Rev Vaccines. 2008;7:363–371. doi: 10.1586/14760584.7.3.363. [DOI] [PubMed] [Google Scholar]
- Tani H, Abe T, Matsunaga TM, Moriishi K., and, Matsuura Y. Baculovirus vector for gene delivery and vaccine development. Future Virol. 2008;3:35–43. [Google Scholar]
- McMichael AJ, Borrow P, Tomaras GD, Goonetilleke N., and, Haynes BF. The immune response during acute HIV-1 infection: clues for vaccine development. Nat Rev Immunol. 2010;10:11–23. doi: 10.1038/nri2674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abe T, Takahashi H, Hamazaki H, Miyano-Kurosaki N, Matsuura Y., and, Takaku H. Baculovirus induces an innate immune response and confers protection from lethal influenza virus infection in mice. J Immunol. 2003;171:1133–1139. doi: 10.4049/jimmunol.171.3.1133. [DOI] [PubMed] [Google Scholar]
- Abe T, Hemmi H, Miyamoto H, Moriishi K, Tamura S, Takaku H.et al. (2005Involvement of the Toll-like receptor 9 signaling pathway in the induction of innate immunity by baculovirus J Virol 792847–2858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abe T, Kaname Y, Wen X, Tani H, Moriishi K, Uematsu S.et al. (2009Baculovirus induces type I interferon production through toll-like receptor-dependent and -independent pathways in a cell-type-specific manner J Virol 837629–7640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho YC, Chung YC, Hwang SM, Wang KC., and, Hu YC. Transgene expression and differentiation of baculovirus-transduced human mesenchymal stem cells. J Gene Med. 2005;7:860–868. doi: 10.1002/jgm.729. [DOI] [PubMed] [Google Scholar]
- Bak XY, Yang J., and, Wang S. Baculovirus-transduced bone marrow mesenchymal stem cells for systemic cancer therapy. Cancer Gene Ther. 2010;17:721–729. doi: 10.1038/cgt.2010.32. [DOI] [PubMed] [Google Scholar]
- Lin CY, Lin KJ, Kao CY, Chen MC, Lo WH, Yen TC.et al. (2011The role of adipose-derived stem cells engineered with the persistently expressing hybrid baculovirus in the healing of massive bone defects Biomaterials 326505–6514. [DOI] [PubMed] [Google Scholar]
- Bak XY, Lam DH, Yang J, Ye K, Wei EL, Lim SK.et al. (2011Human embryonic stem cell-derived mesenchymal stem cells as cellular delivery vehicles for prodrug gene therapy of glioblastoma Hum Gene Ther 221365–1377. [DOI] [PubMed] [Google Scholar]
- Zeng J, Du J, Lin J, Bak XY, Wu C., and, Wang S. High-efficiency transient transduction of human embryonic stem cell-derived neurons with baculoviral vectors. Mol Ther. 2009;17:1585–1593. doi: 10.1038/mt.2009.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Y, Lam DH, Yang J, Lin J, Tham CK, Ng WH.et al. (2012Targeted suicide gene therapy for glioma using human embryonic stem cell-derived neural stem cells genetically modified by baculoviral vectors Gene Ther 19189–200. [DOI] [PubMed] [Google Scholar]
- Zeng J, Du J, Zhao Y, Palanisamy N., and, Wang S. Baculoviral vector-mediated transient and stable transgene expression in human embryonic stem cells. Stem Cells. 2007;25:1055–1061. doi: 10.1634/stemcells.2006-0616. [DOI] [PubMed] [Google Scholar]
- Lei Y, Lee CL, Joo KI, Zarzar J, Liu Y, Dai B.et al. (2011Gene editing of human embryonic stem cells via an engineered baculoviral vector carrying zinc-finger nucleases Mol Ther 19942–950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramachandra CJ, Shahbazi M, Kwang TW, Choudhury Y, Bak XY, Yang J.et al. (2011Efficient recombinase-mediated cassette exchange at the AAVS1 locus in human embryonic stem cells using baculoviral vectors Nucleic Acids Res 39e107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen GY, Pang DW, Hwang SM, Tuan HY., and, Hu YC. A graphene-based platform for induced pluripotent stem cells culture and differentiation. Biomaterials. 2012;33:418–427. doi: 10.1016/j.biomaterials.2011.09.071. [DOI] [PubMed] [Google Scholar]
- Chen GY, Hwang SM, Su HJ, Kuo CY, Luo WY, Lo KW.et al. (2012Defective antiviral responses of induced pluripotent stem cells to baculoviral vector transduction J Virol 868041–8049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kotin RM, Menninger JC, Ward DC., and, Berns KI. Mapping and direct visualization of a region-specific viral DNA integration site on chromosome 19q13-qter. Genomics. 1991;10:831–834. doi: 10.1016/0888-7543(91)90470-y. [DOI] [PubMed] [Google Scholar]
- Chen HC, Lee HP, Ho YC, Sung ML., and, Hu YC. Combination of baculovirus-mediated gene transfer and rotating-shaft bioreactor for cartilage tissue engineering. Biomaterials. 2006;27:3154–3162. doi: 10.1016/j.biomaterials.2006.01.018. [DOI] [PubMed] [Google Scholar]
- Sung LY, Lo WH, Chiu HY, Chen HC, Chung CK, Lee HP.et al. (2007Modulation of chondrocyte phenotype via baculovirus-mediated growth factor expression Biomaterials 283437–3447. [DOI] [PubMed] [Google Scholar]
- Sung LY, Chiu HY, Chen HC, Chen YL, Chuang CK., and, Hu YC. Baculovirus-mediated growth factor expression in dedifferentiated chondrocytes accelerates redifferentiation: effects of combinational transduction. Tissue Eng Part A. 2009;15:1353–1362. doi: 10.1089/ten.tea.2008.0310. [DOI] [PubMed] [Google Scholar]
- Chen HC, Sung LY, Lo WH, Chuang CK, Wang YH, Lin JL.et al. (2008Combination of baculovirus-expressed BMP-2 and rotating-shaft bioreactor culture synergistically enhances cartilage formation Gene Ther 15309–317. [DOI] [PubMed] [Google Scholar]
- Chen HC, Chang YH, Chuang CK, Lin CY, Sung LY, Wang YH.et al. (2009The repair of osteochondral defects using baculovirus-mediated gene transfer with de-differentiated chondrocytes in bioreactor culture Biomaterials 30674–681. [DOI] [PubMed] [Google Scholar]
- Lo WH, Hwang SM, Chuang CK, Chen CY., and, Hu YC. Development of a hybrid baculoviral vector for sustained transgene expression. Mol Ther. 2009;17:658–666. doi: 10.1038/mt.2009.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuang CK, Lin KJ, Lin CY, Chang YH, Yen TC, Hwang SM.et al. (2010Xenotransplantation of human mesenchymal stem cells into immunocompetent rats for calvarial bone repair Tissue Eng Part A 16479–488. [DOI] [PubMed] [Google Scholar]
- Lin CY, Chang YH, Lin KJ, Yen TC, Tai CL, Chen CY.et al. (2010The healing of critical-sized femoral segmental bone defects in rabbits using baculovirus-engineered mesenchymal stem cells Biomaterials 313222–3230. [DOI] [PubMed] [Google Scholar]
- Lin CY, Chang YH, Kao CY, Lu CH, Sung LY, Yen TC.et al. (2012Augmented healing of critical-size calvarial defects by baculovirus-engineered MSCs that persistently express growth factors Biomaterials 333682–3692. [DOI] [PubMed] [Google Scholar]
- Chen CY, Wu HH, Chen CP, Chern SR, Hwang SM, Huang SF.et al. (2011Biosafety assessment of human mesenchymal stem cells engineered by hybrid baculovirus vectors Mol Pharm 81505–1514. [DOI] [PubMed] [Google Scholar]
- Luo WY, Shih YS, Lo WH, Chen HR, Wang SC, Wang CH.et al. (2011Baculovirus vectors for antiangiogenesis-based cancer gene therapy Cancer Gene Ther 18637–645. [DOI] [PubMed] [Google Scholar]
- Hervas-Stubbs S, Rueda P, Lopez L., and, Leclerc C. Insect baculoviruses strongly potentiate adaptive immune responses by inducing type I IFN. J Immunol. 2007;178:2361–2369. doi: 10.4049/jimmunol.178.4.2361. [DOI] [PubMed] [Google Scholar]
- Kitajima M., and, Takaku H. Induction of antitumor acquired immunity by baculovirus Autographa californica multiple nuclear polyhedrosis virus infection in mice. Clin Vaccine Immunol. 2008;15:376–378. doi: 10.1128/CVI.00285-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng J, Shahbazi M, Wu C, Toh HC., and, Wang S. Enhancing immunostimulatory function of human embryonic stem cell-derived dendritic cells by CD1d overexpression. J Immunol. 2012;188:4297–4304. doi: 10.4049/jimmunol.1102343. [DOI] [PubMed] [Google Scholar]
- Lotze MT., and, Kost TA. Viruses as gene delivery vectors: application to gene function, target validation, and assay development. Cancer Gene Ther. 2002;9:692–699. doi: 10.1038/sj.cgt.7700493. [DOI] [PubMed] [Google Scholar]
- Ames RS., and, Lu Q. Viral-mediated gene delivery for cell-based assays in drug discovery. Expert Opin Drug Dis. 2009;4:243–256. doi: 10.1517/17460440902751599. [DOI] [PubMed] [Google Scholar]
- Vijayachandran LS, Viola C, Garzoni F, Trowitzsch S, Bieniossek C, Chaillet M.et al. (2011Robots, pipelines, polyproteins: enabling multiprotein expression in prokaryotic and eukaryotic cells J Struct Biol 175198–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drugmand JC, Schneider YJ., and, Agathos SN. Insect cells as factories for biomanufacturing. Biotechnol Adv. 2012;30:1140–1157. doi: 10.1016/j.biotechadv.2011.09.014. [DOI] [PubMed] [Google Scholar]
- Kost TA, Condreay JP, Ames RS, Rees S., and, Romanos MA. Implementation of BacMam virus gene delivery technology in a drug discovery setting. Drug Discov Today. 2007;12:396–403. doi: 10.1016/j.drudis.2007.02.017. [DOI] [PubMed] [Google Scholar]
- Kost TA, Condreay JP., and, Ames RS. Baculovirus gene delivery: a flexible assay development tool. Curr Gene Ther. 2010;10:168–173. doi: 10.2174/156652310791321224. [DOI] [PubMed] [Google Scholar]
- Davenport EA, Nuthulaganti P., and, Ames RS. BacMam: versatile gene delivery technology for GPCR assays. Methods Mol Biol. 2009;552:199–211. doi: 10.1007/978-1-60327-317-6_14. [DOI] [PubMed] [Google Scholar]
- Condreay JP., and, Watson CA. Pharmacological applications of baculovirus-mediated protein expression in mammalian cells. Curr Protoc Pharmacol. 2010;Chapter 6:Unit 6.11. doi: 10.1002/0471141755.ph0611s49. [DOI] [PubMed] [Google Scholar]
- Shukla S, Schwartz C, Kapoor K, Kouanda A., and, Ambudkar SV. Use of baculovirus BacMam vectors for expression of ABC drug transporters in mammalian cells. Drug Metab Dispos. 2012;40:304–312. doi: 10.1124/dmd.111.042721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hassan NJ, Pountney DJ, Ellis C., and, Mossakowska DE. BacMam recombinant baculovirus in transporter expression: a study of BCRP and OATP1B1. Protein Expr Purif. 2006;47:591–598. doi: 10.1016/j.pep.2005.12.008. [DOI] [PubMed] [Google Scholar]
- Pfohl JL, Worley JF., 3rd, , Condreay JP, An G, Apolito CJ, Kost TA.et al. (2002Titration of KATP channel expression in mammalian cells utilizing recombinant baculovirus transduction Recept Channels 899–111. [PubMed] [Google Scholar]
- Beacham DW, Blackmer T, O' Grady M., and, Hanson GT. Cell-based potassium ion channel screening using the FluxOR assay. J Biomol Screen. 2010;15:441–446. doi: 10.1177/1087057109359807. [DOI] [PubMed] [Google Scholar]
- Machleidt T, Robers MB, Hermanson SB, Dudek JM., and, Bi K. TR-FRET cellular assays for interrogating posttranslational modifications of histone H3. J Biomol Screen. 2011;16:1236–1246. doi: 10.1177/1087057111422943. [DOI] [PubMed] [Google Scholar]
- Xie W, Pao C, Graham T, Dul E, Lu Q, Sweitzer TD.et al. (2012Development of a cell-based high throughput luciferase enzyme fragment complementation assay to identify nuclear-factor-e2-related transcription factor 2 activators Assay Drug Dev Technol 10514–524. [DOI] [PubMed] [Google Scholar]
- Jenkinson S, McCoy DC, Kerner SA, Ferris RG, Lawrence WK, Clay WC.et al. (2003Development of a novel high-throughput surrogate assay to measure HIV envelope/CCR5/CD4-mediated viral/cell fusion using BacMam baculovirus technology J Biomol Screen 8463–470. [DOI] [PubMed] [Google Scholar]
- Trowitzsch S, Klumpp M, Thoma R, Carralot JP., and, Berger I. Light it up: highly efficient multigene delivery in mammalian cells. Bioessays. 2011;33:946–955. doi: 10.1002/bies.201100109. [DOI] [PubMed] [Google Scholar]
- Isom HC, Abdelhamed AM, Bilello JP., and, Miller TG. Baculovirus-mediated gene transfer for the study of hepatitis B virus. Methods Mol Med. 2004;96:219–237. doi: 10.1385/1-59259-670-3:219. [DOI] [PubMed] [Google Scholar]
- Pan Y, Zhao Q, Fang L, Luo R, Chen H., and, Xiao S. Efficient gene delivery into mammalian cells by recombinant baculovirus containing a hybrid cytomegalovirus promoter/Semliki Forest virus replicon. J Gene Med. 2009;11:1030–1038. doi: 10.1002/jgm.1390. [DOI] [PubMed] [Google Scholar]
- Li J, Sun B, Han H, Ouyang J, Yao H, Yang Y.et al. (2012Application of baculovirus as a delivery vehicle for study of transcription and translation mechanism of parvovirus in non-permissive mammalian cells J Virol Methods 1831–7. [DOI] [PubMed] [Google Scholar]
- Scott MJ, Modha SS, Rhodes AD, Broadway NM, Hardwicke PI, Zhao HJ.et al. (2007Efficient expression of secreted proteases via recombinant BacMam virus Protein Expr Purif 52104–116. [DOI] [PubMed] [Google Scholar]
- Dukkipati A, Park HH, Waghray D, Fischer S., and, Garcia KC. BacMam system for high-level expression of recombinant soluble and membrane glycoproteins for structural studies. Protein Expr Purif. 2008;62:160–170. doi: 10.1016/j.pep.2008.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urabe M, Ding C., and, Kotin RM. Insect cells as a factory to produce adeno-associated virus type 2 vectors. Hum Gene Ther. 2002;13:1935–1943. doi: 10.1089/10430340260355347. [DOI] [PubMed] [Google Scholar]
- Virag T, Cecchini S., and, Kotin RM. Producing recombinant adeno-associated virus in foster cells: overcoming production limitations using a baculovirus-insect cell expression strategy. Hum Gene Ther. 2009;20:807–817. doi: 10.1089/hum.2009.092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chejanovsky N., and, Carter BJ. Mutagenesis of an AUG codon in the adeno-associated virus rep gene: effects on viral DNA replication. Virology. 1989;173:120–128. doi: 10.1016/0042-6822(89)90227-4. [DOI] [PubMed] [Google Scholar]
- King JA, Dubielzig R, Grimm D., and, Kleinschmidt JA. DNA helicase-mediated packaging of adeno-associated virus type 2 genomes into preformed capsids. EMBO J. 2001;20:3282–3291. doi: 10.1093/emboj/20.12.3282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith RH, Levy JR., and, Kotin RM. A simplified baculovirus-AAV expression vector system coupled with one-step affinity purification yields high-titer rAAV stocks from insect cells. Mol Ther. 2009;17:1888–1896. doi: 10.1038/mt.2009.128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cecchini S, Virag T., and, Kotin RM. Reproducible high yields of recombinant adeno-associated virus produced using invertebrate cells in 0.02- to 200-liter cultures. Hum Gene Ther. 2011;22:1021–1030. doi: 10.1089/hum.2010.250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dropulic B. Lentiviral vectors: their molecular design, safety, and use in laboratory and preclinical research. Hum Gene Ther. 2011;22:649–657. doi: 10.1089/hum.2011.058. [DOI] [PubMed] [Google Scholar]
- Lim KI. Retroviral integration profiles: their determinants and implications for gene therapy. BMB Rep. 2012;45:207–212. doi: 10.5483/bmbrep.2012.45.4.207. [DOI] [PubMed] [Google Scholar]
- Merten OW, Charrier S, Laroudie N, Fauchille S, Dugué C, Jenny C.et al. (2011Large-scale manufacture and characterization of a lentiviral vector produced for clinical ex vivo gene therapy application Hum Gene Ther 22343–356. [DOI] [PubMed] [Google Scholar]
- Cockrell AS., and, Kafri T. Gene delivery by lentivirus vectors. Mol Biotechnol. 2007;36:184–204. doi: 10.1007/s12033-007-0010-8. [DOI] [PubMed] [Google Scholar]
- Huang KS, Lo WH, Chung YC, Lai YK, Chen CY, Chou ST.et al. (2007Combination of baculovirus-mediated gene delivery and packed-bed reactor for scalable production of adeno-associated virus Hum Gene Ther 181161–1170. [DOI] [PubMed] [Google Scholar]