

Abstract
Optic ataxia is a high order deficit in reaching to visual goals that occurs with posterior parietal cortex (PPC) lesions. It is a component of Balint’s syndrome that also includes attentional and gaze disorders. Aspects of optic ataxia are misreaching in the contralesional visual field, difficulty preshaping the hand for grasping, and an inability to correct reaches online. Recent research in non-human primates (NHPs) suggests that many aspects of Balint’s syndrome and optic ataxia are a result of damage to specific functional modules for reaching, saccades, grasp, attention, and state estimation. The deficits from large lesions in humans are likely composite effects from damage to combinations of these functional modules. Interactions between these modules, either within posterior parietal cortex or downstream within frontal cortex, may account for more complex behaviors such as hand-eye coordination and reach-to-grasp.
Introduction
Optic ataxia (OA) is a fascinating neurological deficit that has been of considerable interest for the last century. OA patients have difficulty reaching to visually guided goals in peripheral vision, and the deficit leaves voluntary eye movements largely unaffected (Figure 1). The cortical neurologic deficit is a result of a lesion to the superior parietal lobule (SPL) and areas around the intraparietal sulcus (IPS). Interestingly there are no primary sensory or motor deficits involved in lesions to this region, and thus the problem is at a more integrative sensorimotor level. This clinical phenomenon’s specificity and reproducibility provides interesting insights into brain organization and higher level sensorimotor functions.
Figure 1. Patient with optic ataxia.
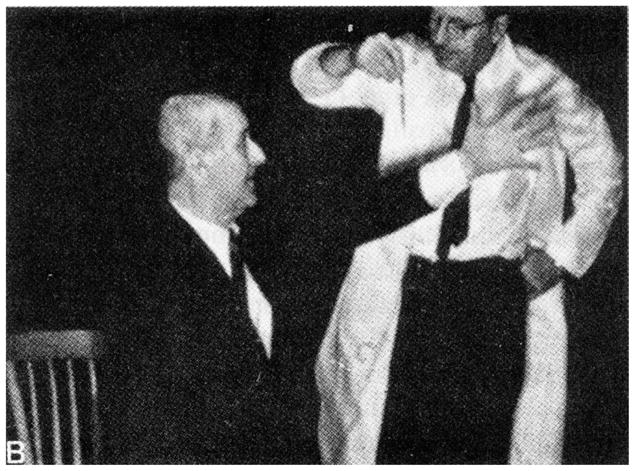
The patient misreaches beyond the pencil when asked to touch it. From Cogan 1965.
In clinical practice, studies of deficits after brain lesions in humans provide powerful insights into the functional roles of the affected brain areas. However, as pointed out by numerous investigators, lesion studies have some drawbacks in terms of interpretation of the behavioral results. Foremost is the fact that these accidents of nature invariably involve more than one functional module. Thus there can be a large degree of individual variability in both the type and severity of defects.
As a case in point, Balint’s syndrome, first described by the Hungarian neurologist Rezso Balint in 1909, has three defining symptoms: psychic paralysis of gaze, spatial disorder of attention, and optic ataxia. Balint coined the term optic ataxia. He believed the deficit was not an apraxia (a disorder in executing learned behaviors), but rather a lack of coordination between visual input and motor outputs, and thus the term ataxia, Greek for “Lack of Order”. Balint drew on the similarity with the contemporary disorder in the early 1900’s of “tabetic ataxia”, a deficit of proprioceptive origin due to dorsal column degeneration seen in patients with tertiary syphilis who are ataxic due to their lack of sensory input. Since Balint’s patient had difficulty in visually driven reach tasks, he defined the deficit as optic ataxia. However, Balint distinguished optic ataxia from simple visual deficits since his patient had a “hand effect” in which misreaching was largely confined to the right hand.
Balint’s patient had bilateral lesions of the posterior parietal cortex. Subsequent studies showed that the deficit could also result from unilateral lesions and that optic ataxia could exist in isolation from the attentional disorders. However, OA usually exists with other disturbances such as misshaping of the hand for grasping (Figure 1) and a deficit in online visuomotor control.
New animal studies are beginning to elucidate a finer grain of functional organization within the PPC. Recording experiments have demonstrated regions for reaching, grasping and eye movements. Early functional magnetic resonance imaging (fMRI) studies in humans showed varying degrees of functional localization. However, recent fMRI studies with advanced data analysis techniques and studies using magnetoencephalography (MEG) and transcranial magnetic stimulation (TMS) are in more agreement with the animal findings of functional localization.
This review will include studies of PPC using pharmacological inactivations in animals. One of the advantages of this approach is confining the inactivation to single functionally and anatomically defined modules. Thus these more punctate inactivations provide a bridge between the human neuropsychological and imaging studies and animal recording studies.
Although much work is still needed, an emerging picture suggests that the various and complex aspects of OA are the result of damage to an array of functional modules. Moreover, deficits in complex behaviors such as reach-to-grasp and eye-hand coordination may be explained by the disruption of modules whose coordinated interactions give rise to these complex behaviors.
Clinical: Human optic ataxia (OA)
Symptoms and Neurologic Exam in Optic Ataxia
Optic ataxia is an ataxia in which patients have difficulty completing visually guided reaching tasks in the absence of other sensory cues. Patients with isolated optic ataxia have intact visual fields, stereoscopic vision, oculomotor control, proprioception, motor abilities and cerebellar function, excluding other causes of ataxia with reaching (Garcin et al., 1967; Perenin & Vighetto, 1988).
On neurologic examination deficits in reaching tasks appear similar to cerebellar ataxias, in that the patients exhibit dysmetria with overshooting and undershooting of the target; however, unlike cerebellar ataxia, once the patients receives proprioceptive or auditory cues, they are able to perform the task smoothly. For example, Holmes described a patient wounded in World War I in 1916 who was able to correct his reaching upon receiving proprioceptive information: “He suffered with an extremely gross disturbance of localization in space by vision, but this diminished after some weeks. When asked to touch or grasp my hand or a pencil held in front of his eyes, he groped wildly for it, and, as a rule, brought his hand beyond it when it was within his reach, but he made errors in every direction in the judgment of its position. If his hand, however, came in contact with my arm, he moved his fingers promptly along this until they reached my hand or the object it held.” (Holmes, 1918). Most of the patients described by Holmes were also able accurately to touch or bring food to their own body, demonstrating that only visually guided tasks were compromised. In addition, these patients lack dysdiadokinesia (impairment of rapid alternating movements) and intention tremor (increased amplitude of tremor as approaching the intended object), both of which can be seen in cerebellar ataxias (Blumenfeld, 2002; Caplan, and van Gijn, 2012).
Causes
In human patients OA is often accompanied by other deficits given the proximity to other important structures and given that many of the disease processes that lead to OA affect large territories expanding past the occipito-parietal junction (Rafal, 2001). OA often appears in the context of Balint’s syndrome with more widespread bilateral parietal insults resulting in the triad of optic ataxia, simultagnosia and ocular apraxia (Balint, 1909; Homes, 1918). Balint’s syndrome can be caused by a variety of disease processes including vascular compromise, neurodegenerative diseases, infections, traumatic brain injury, inflammatory diseases and iatrogenic causes. Vascular infarcts involving the posterior parietal cortex are often caused by watershed infarcts at the location of the terminal branches of the middle cerebral artery and posterior cerebral artery, a location which is particularly vulnerable in cases of hypotension and cardiac arrest (Montero et al., 1982; Rafal, 2001; Caplan and van Gijn, 2012; Cavina-Pratesi et al., 2013). In addition, occlusions of the posterior circulation, such as those caused by atherosclerotic vertebrobasilar disease, can also lead to bilateral hypoperfusion of the posterior parietal cortex at the watershed area between the middle cerebral artery and posterior cerebral artery (Montero et al., 1982).
Progressive neurodegenerative diseases are common in the aging population and can cause damage to varying parts of the brain, but have propensity towards the posterior cortex. These neurodegenerative diseases that can cause Balint’s syndrome include posterior cortical atrophy, Alzheimer’s disease, dementia with Lewy Bodies, Parkinson’s disease and Creutzfeldt Jakob’s disease (McMonagle et al., 2006; Kas et al., 2011, Meek et al., 2013; Mendez et al., 1990; Hof et al., 1990; Kirshner and Lavin, 2006; Moreaud, 2003). In children X-linked adrenoleukodystropy, a disabling progressive genetic lipid storage disorder, may also present with OA in the setting of Balint’s syndrome (Carmant, 1998). Given that these neurodegenerative diseases are usually caused by deposition of unwanted products in the cortex leading to disruption of signal transmission, such as amyloid plaques in Alzheimer’s disease, they are often accompanied by more extensive symptoms, such as memory deficits and movement disorders. In posterior cortical atrophy, a progressive neurodegenerative disease targeting the parieto-occipital and temporo-occipital lobes, Balint’s syndrome may be one of the presenting symptoms, demonstrating the importance of clinicians’ familiarity with Balint’s syndrome and OA. For example, in one case series of 19 patients with posterior cortical atrophy, 5 presented with Balint’s triad, 10 presented with the combination of simultanagnosia and OA, and 2 presented with simultanagnosia alone (McMonagle et al., 2006).
Other less common causes of Balint’s syndrome cited in the literature include traumatic brain injury, brain tumors, infections, inflammatory conditions, and seizures. Reports of traumatic brain injury involving parietal damage were common during World War I when Holmes described 6 patients with penetrating wounds (Holmes, 1918). Brain tumors affecting the bilateral parietal lobes include CNS lymphomas and “butterfly tumors”, which are often caused by high-grade astrocytomas or glioblastoma multiforme, a common malignant tumor that can cross the corpus collosum (Rafal, 2001). Balint’s syndrome has been seen in posterior reversible encephalopathy syndrome (PRES), a syndrome caused by vasogenic edema, most commonly affecting the parietal and occipital cortex and underlying white matter, and may be seen secondary to a variety of medical conditions including malignant hypertension, eclampsia and pre-eclampsia in pregnancy, high dose chemotherapy, post-transplant immunosuppressive drugs and renal disease (Fugate et al., 2010; Kumar et al., 2011; Gurjinder, 1989). Infectious processes have been reported to cause optic ataxia in Balint’s syndrome including cerebral toxoplasmosis and progressive multifocal leukoencephalopathy secondary to the JC Virus in immunocompromised patients with HIV (Ayuso-Peralta et al., 1994; Garcia Guijo et al., 1990), and in herpes encephalitis in an infant (Amin et al., 2012). Iatrogenic causes reported include administration of nitroglycerin leading to hypotension and watershed infarcts and secondary to angiography contrast prior to CT (Mejia et al., 2008; Rafal, 2001; Merchut and Richie, 2002). Finally, Balint’s syndrome has been reported in a case of status epilepticus due to an uncontrolled seizure disorder (Ristic et al., 2012).
Causes of “pure optic ataxia,” that is OA occurring in isolation without the simultagnosia and ocular apraxia of Balint’s syndrome, is a rarer finding due to the fact that pure OA is caused by more localized and discrete lesions affecting the unilateral posterior parietal cortex. Pure optic ataxia was first described by Garcin et al. in 1967. This patient had older lesions of the left hemisphere but presented with a new deficit eight years later that was diagnosed as a lesion to the right parietal cortex using EEG. This newer injury produced OA. Although the authors acknowledged that the patient had bilateral lesions, they believed the later lesion to be the one responsible for OA. Subsequent reports of pure OA include various unilateral brain tumors, isolated infarcts, hematomas and localized traumatic brain injury (Perenin and Vighetto, 1988). Posterior cortical atrophy has also been reported to cause pure optic ataxia (Meek, 2013).
Prognosis
Given that the causes of both Balint’s syndrome and pure OA are vast, the prognosis of recovery from optic ataxia is largely based on the underlying mechanism of disease, the individual’s clinical course and co-morbid conditions. For example, Perenin and Vighetto, 1988 reported a series of 10 patients with pure optic ataxia, and prognosis was described in 7 of the patients with varying outcomes. In two patients with hematomas, one showed improvement of his optic ataxia at 2 years with resolution at 8 years, while the other patient’s OA was partially resolved within 3 months. Their patient with a widespread astrocytoma, status post partial tumor debulking, demonstrated no changes in his symptoms following his surgery until his death 4 months later. On the other hand, their patient with a malignant melanoma survived surgical removal and continued to have OA 2 years following his surgery but in a more mild form. Their patient with a pyogenic abscess obtained partial recovery at 3 years follow up, and the one stroke patient whose long-term clinical course demonstrated recovery from OA at 6 years after the onset of his symptoms.
Balint’s syndrome and OA are rarely described in children, but in one case series of three children, the prognosis appears to be superior to those described in adults. Hutchinson et al’s case series reported a 7 year old boy who had traumatic brain injury affecting the parietal lobes following a motor vehicle accident who presented with Balint’s syndrome and had resolution of his symptoms 6 months after the accident. They also reported a 5 year old girl with a mitochondrial disorder and bilateral parieto-occipital strokes who also presented with Balint’s syndrome and subsequently improved 1 year after her infarcts. Finally they reported a 2 year old girl with encephalitis secondary to herpes simplex virus resulting in bilateral temporal and parieto-occipital damage who showed improvement of her Balint’s syndrome one year after the infection (Amin et al., 2012).
Neuropsychological research in humans
Location of lesion
Initial reports of OA involved individuals with bilateral lesions. Balint’s patient suffered lesions to the posterior aspect of the parietal lobe involving both the inferior and superior parietal lobules. Holmes and Horrax’s 1919 patient had a bilateral lesion they interpreted as involving the supramarginal gyrus of both hemispheres, but based only on position of entrance and exit wounds on the skull. Holmes, 1918 reported on 6 patients with what appeared to be bilateral lesions, centered on the angular and supramarginal gyri but also extending to surrounding occipital, temporal and parietal cortex, and mesial cortex. Lesion locations for two of the patients were verified by autopsy.
Garcin et al., 1967 described a patient who had bilateral lesions but the lesions occurred separated in time and suggested that optic ataxia could result from a unilateral lesion. In a landmark paper Perenin and Vighetto, 1988 studied optic ataxia with unilateral lesions in 10 patients. Not only did they demonstrate that OA can arise from unilateral damage, but they also showed from X-ray computed tomography that the overlap of lesions between subjects included primarily the superior parietal lobule (SPL) and intraparietal sulcus (IPS). A subsequent study further localized common lesion locations in 16 OA subjects using magnetic resonance imaging (MRI) (Karnath and Perenin, 2005). Lesion overlap between subjects was found at the junction of the SPL and occipital cortex and extending medially into the precuneus. In summary, although initial reports indicated that OA resulted from bilateral lesions localized to the supramarginal gyrus, subsequent studies showed that OA can be result with unilateral lesions and the primary lesion sites are the SPL and IPS.
Three frameworks for Optic Ataxia
Three basic frameworks have been proposed for OA—disruption of 1) visuomotor processing, 2) visual orientation or 3) online visuomotor control. The visuomotor framework posits that the deficit is one of sensorimotor integration, in this case specific to vision and reaching. Figure 1 is taken from Cogan 1965, and shows a patient with optic ataxia misreaching to a pencil. The second framework ascribes OA to a disruption of visual orientation in which the patient cannot properly orient or judge the location of objects in space. The third framework holds that OA is due to a deficit in a mechanism which is normally used to correct ongoing movements via visual feedback. A discussion of these 3 concepts follows.
Visuomotor deficit
Balint proposed that optic ataxia was a visuomotor deficit. He reasoned that it was visuomotor because his patient did not have primary visual or motor defect but showed poor reaching to visual goals with the right limb. Thus the deficit appeared to be of a higher order and specific to visually guided reaching. Perenin and Vighetto, 1988, who also interpreted OA in visuomotor terms, found that their unilateral lesion patients had OA that was independent of attention deficits, showing that OA can exist without the other components of Balint’s syndrome. They observed both proximal and distal effects, with misreaching as well as difficulty orienting the wrist. There were “field” effects with greater difficulty in reaching in the contralesional space for either left or right hemisphere damage. “Hand” effects were also present for most left hemisphere damaged patients in which the right hand misreached to either visual field. If the subjects could orient their gaze to the object before reaching, the errors were dramatically reduced, indicating that reaching to targets in the peripheral visual field show the largest effects.
Perenin and Vighetto’s observations later formed the cornerstone for the dorsal pathway of Goodale and Milner’s 1992 two cortical visual systems hypothesis. Patients with lesions to ventral visual areas have agnosia—difficulty in recognizing objects—whereas patients with lesions to the dorsal visual areas have OA—difficulty in properly grasping objects. They proposed that the ventral system was for perception (“what”) and the dorsal system was for action (“how”).
A number of other researchers have also come to the conclusion that OA is a visuomotor disturbance. Rondot et al., 1977 proposed that OA be renamed visuomotor ataxia. Jackson et al., 2005 studied a patient who could reach to foveated targets with either hand but could not reach with the right hand to peripheral targets. The reaches with the right hand were heavily biased toward the direction of gaze, similar to previous reports of magnetic misreaching in which reach is constrained to locations that are fixated (Carey et al., 1997; Buxbaum and Coslett, 1997). The authors proposed that OA is a breakdown in the ability to decouple eye and hand movements for spatially independent movements.
Although the visual aspect of manual control in OA is most apparent on casual inspection, some patients also show a deficit in proprioceptively guided reaching; for instance when the ataxic hand is reaching to the other hand in the dark (Blangero et al., 2007; Jackson et al., 2009; Pisella et al., 2000). Also suggestive of proprioceptive involvement, OA patients show hand effects in addition to field effects that combine to produce the greatest errors for pointing in the contralesional field with the contralesional hand (Blangero, 2007).
Spatial disorientation
Holmes, 1918 and Holmes and Horrax, 1919 reported on soldiers with bilateral penetrating brain injuries of the PPC. The symptoms they described were very similar to those of Balint’s patient. However, rather than a visuomotor interpretation, they put forth the notion that the deficits could all be explained under one perceptual principle, the loss of visual orientation.
Similar to Balint’s patient, the deficits reported by Holmes and Holmes and Horrax could not be attributed to primary visual or motor disturbances and thus were higher level. However, the defects were restricted to the visual modality. The subjects had gaze difficulty in finding and fixating objects, similar to Balint’s psychic paralysis of gaze. Attention disorders similar to spatial neglect and simultagnosia were also reported. Errors in pointing were observed that persisted over the course of recovery for peripheral vision (measured in Figure 3 of Holmes and Horrax). However, the authors focused on disturbances to visual orientation and space perception. These defects included perceiving the relative and absolute distances of objects from the body, their lengths and sizes, and relative positions of objects. Moreover, the patients had extreme difficulty in avoiding obstacles when walking. They interpreted the misreaching and gaze difficulties as being secondary consequences of a loss of visual orientation. Thus, Cogan’s patient’s misreaching in Figure 1 would be attributed to his inability to accurately perceive the spatial location of the pencil.
More recently, McIntosh et al., 2011 showed that an OA subject had correlated deficits in correcting for reaches to jumped targets and perceiving the direction of the jumps, especially when the jumps were toward the periphery. They proposed that OA is a perceptual deficit that results from slowed contralesional orienting of attention. However, this finding also has similarities to spatial disorientation.
Although general spatial disorientation may manifest itself in some OA patients, this idea is at odds with the fact that many OA patients show deficits of specific visuomotor functions. Reach inaccuracies have been reported to be present without saccade inaccuracies (Trillenberg et al., 2007). Also, the hand effect first reported by Balint also does not fit easily into a general perceptual deficit. Finally, if there is a general spatial deficit it would easily explain the often simultaneous occurrence of reach and grasp inaccuracies. However, patient studies show differential effects on proximal/grasp and distal/reach behaviors (Himmelbach et al., 2006; Cavina-Pratesi et al., 2013; see also Online Control).
Online control
Early neuropsychology studies did not have the technical advantage of being able to precisely and repeatedly measure the dynamics of limb and eye movements. Recent studies have shown that the dynamics of these movements are disturbed with PPC lesions. Moreover, the ability to make online adjustments from visual feedback is compromised. These findings have led to an alternative hypothesis to explain OA—that the errors in reaching are due to disruption of an online correction mechanism.
If a visual target is jumped around the time of a reach, normal subjects can modify the movement online, often unconsciously, to account for the perturbation. Pisella et al., 2000 deemed this online control process an “automatic pilot” because normal subjects could not suppress adjusting their movements in response to target jumps. In contrast, their OA patient did not invoke this “automatic pilot” and they proposed that OA interferes with this online correction mechanism. Since the patient could stop executing a movement in response to a change in the target color with similar ability to controls, they reasoned that her deficit could not be due to a general slowing of visual or motor processing. Moreover, they accounted for the greater inaccuracies for reaching to peripheral targets as peripheral vision being less accurate and thus requiring greater on-line control.
The online control hypothesis has also been extended to fast oculomotor control. Gaveau et al., 2008 studied eye-hand coordination in two OA patients. Using the target jump paradigm, but instructing the subjects to make a coordinated eye and hand movement to the target instead of just a hand movement, they found that the OA patients took more time to accurately fixate the jumped target and delayed reaching until the final corrective saccade, thus extending the onset of the reach as well. The authors contrasted the loss of fast ocular control with the previous reports of loss of fast manual control and raised the possibility that a common cortical module is responsible for online control of eye and hand movements. However, the online correction idea does not appear to generalize to grip scaling. Himmelbach et al., 2006 studied the same patient as Pisella et al., 2000. They found that this patient was able to adjust grip size online as well as controls, indicating that the online deficit, at least for this patient, appears to be restricted to reach and not grasp.
Coordinate frames
The study of coordinate frames of OA has the advantage of mapping the deficit onto animal research, in which the coordinate frames of PPC cortical areas have been extensively studied. Also, since one of the roles of the PPC appears to be coordinate transformations, OA deficits have also been framed in the context of a deficit in these transformations.
The transformation required in visually guided reaching is to convert the location of the visual stimulus, coded in eye coordinates, into the appropriate action in motor coordinates. Between these stages can be intermediate representations in head, body and hand coordinates or combinations of these coordinate frames (Andersen and Buneo, 2002; Bremner et al., 2012). Studies of OA patients have examined the coordinate frame of the deficit. It has also been proposed that OA is in part a defect in making transformations between coordinate frames
The parietal reach region (PRR) in the PPC of monkeys has been found to represent space largely in eye-centered coordinates (Batista et al., 1999; Cohen and Andersen, 2002). In experiments in which the eye, head and body position of visual reach targets have been varied, the misreaching of OA patients has been reported to vary only with eye position (Dijkerman et al., 2006; Blangero et al., 2010). This result is indicative of damage to a cortical representation of reach targets in eye coordinates and draws a parallel with the monkey studies of space representation in PRR. This representation in eye coordinates also applied to the internal representation of the target because when a saccade was interleaved between the target presentation and the reach, the field effect of OA misreaching manifested in relation to the new gaze direction, not to the original gaze direction at the time of target presentation (Khan et al., 2005).
Buxbaum and Coslett, 1997, 1998 proposed that the non-foveal reaching errors in OA are the result of the transformation from eye-centered coordinates to other coordinate frames. Khan et al., 2013 examined the effects of head roll on reaching errors in an OA patient with a unilateral lesion. The subject was able to compensate for head roll in the ipsilesional visual field but not the contralesional visual field. They interpret their results as causal evidence that the PPC is involved in coordinate transformations and these results are consistent with the interruption that a disruption in the coordinate transformation process with OA.
In the study of OA for reaching in the dark, it was found that the proprioceptive deficit was also in eye coordinates (Blangero et al., 2007). This dual representation of targets and hand position in eye coordinates is consistent with earlier animal studies suggesting a direct transformation of reach vectors from these variables coded in eye coordinates (Buneo et al., 2002). Observing field effects for both the proprioceptive position of the reaching hand, and the visual location of the target, Jackson et al., 2009 proposed that the OA deficit is one of simultaneously representing the hand, in postural coordinates, and the target, in external coordinates. Moreover, they hypothesize that the deficit may be in the additional step of combining information in these two coordinate frames.
Strengths and limitations of neuropsychological data
A good deal of what we know about the functions of brain areas comes from lesion deficits. However, if we look at the OA patient reaching in Figure 1, his deficit is consistent with all three frameworks of visuomotor, visual orientation and online deficits. Difficulties in distinguishing between these models based only on the study of OA patients include the relative scarcity of OA patients (Jax et al., 2009), the majority of studies using only one or two patients, and many single case reports being made from the same patients (Borchers et al., 2013). Perhaps though, the greatest difficulty is the variability of deficits that arises from rather large lesions of cortex (Glover, 2004; Jax et al., 2009). Figure 2 shows axial MRIs of two of the most studied OA patients in the literature, A.T. (a) and I.G. (b) (Schindler et al., 2004). Both patients have large and extensive bilateral lesions involving large extents of the PPC. Thus many different functional modules are likely involved.
Figure 2. Patient lesions are large and likely involve several functional areas.
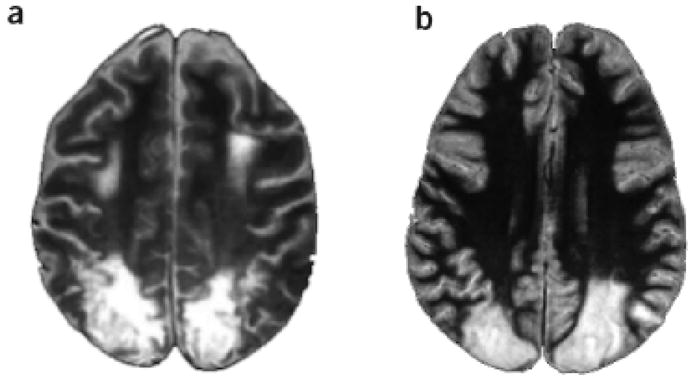
Horizontal magnetic resonance imagining sections through the bilateral parietal lobe lesions of patients A.T. (a) and I.G. (b). From Schindler et al. 2004.
The next sections will review in finer detail the organization of PPC. These studies involve non-human primate (NHP) anatomy and physiology and human non-invasive studies including functional imaging and stimulation. Finally, animal studies in which selective modules are temporarily pharmacologically inactivated will be discussed. NHP inactivations tie together human lesion and human normal studies with animal studies for a clearer picture of Balint’s syndrome—a composite of damage to many modules. However, optic ataxia in isolation can be explained by damage to a very specific module, the parietal reach region (PRR).
Animal studies
In single cell recording experiments in behaving NHPs Mountcastle and colleagues (Mountcastle et al., 1975; Lynch et al., 1977) identified a number of different cell types within the PPC. These included cells active for reach, saccades, smooth pursuit eye movements, hand manipulation, and fixation. Robinson et al., 1978 identified modulation of visual responses in PPC by attention.
In their original formulation Mountcastle et al. proposed that the different cell types were organized into inter-digitated columns within PPC. Later studies have shown a fine parcellation of PPC into different cortical areas with respect to function (Andersen and Buneo, 2002). This review will concentrate on four of these cortical areas that have relevance to Balint’s syndrome and OA.
The lateral intraparietal area (LIP)
A number of studies have implicated LIP in the processing of saccadic eye movements. The cortex of the lateral wall of the posterior half of the intraparietal sulcus (IPS) where LIP is located, is connected more strongly to saccadic eye movement centers than other parts of the PPC. These connections include projections to the intermediate and deep layers of the superior colliculus (Lynch et al., 1985) and connections the frontal eye fields (FEF) in the frontal lobe (Andersen et al., 1985). A subsequent study reported that cells within LIP had responses preceding eye movements whereas adjoining cortex on the convexity of the inferior parietal lobe, area 7a, had mostly post-saccadic responses (Andersen et al., 1987). Gnadt and Andersen, 1988 discovered a previously unreported activity in the PPC of persistent activity in area LIP when monkeys planned a saccade to a remembered location in the dark. This persistent activity was shown to be stronger for planning eye movements than reach movements, indicating that it represents the motor plan for an eye movement and not general memory or attention (Figure 3a, Snyder et al., 1997, Cui et al., 2007). Electrical stimulation of LIP produces saccades (Thier and Andersen, 1998). Taken together, these studies show that LIP plays an important role in saccades. Other studies have also implicated LIP in attention processing (Colby and Goldberg, 1999 for review).
Figure 3. Action specificity in non-human primate posterior parietal areas.

a–b. Cells recorded from the lateral intraparietal area (LIP) and the parietal reach region (PRR) in saccade versus reach trials. The LIP neuron is more active in saccade trials, whereas the PRR neuron is more active in reach trials. Modified from Snyder et al. 1997. c. Cell recorded from the anterior intraparietal area (AIP) for different objects and hand manipulation. This visuomotor neuron is highly selective for both object and grip type. Modified from Murata et al. 2000.
The parietal reach region (PRR)
The same study that examined persistent activity in LIP selective for saccade plans also found cortex more posterior and medial to LIP with persistent activity selective for reach plans (Figure 3b, Snyder et al., 1997). This region was subsequently named the parietal reach region (PRR), (Snyder et al., 1998, 2000). PRR was labeled a region instead of a cortical area because it appears to include a number of reach selective cortical areas. PRR extends from the dorsal aspect of the parieto-occipital area (PO) including V6A and the posterior aspect of the medial bank of the intraparietal sulcus (IPS) including the medial intraparietal area (MIP) and possibly extending anteriorly in the IPS into area 5v. V6A has been found to contain neurons activated by reach, wrist orientation and grasp (Fattori et al., 2001, 2012). Another area where reach selectivity has been reported is area 5d on the convexity of the SPL bordering PRR (covered in more detail below). Thus there appears to be a complex of reach related cortical areas within the IPS, PO, and SPL. Most subsequent studies of PRR have concentrated on the medial bank of the IPS (e.g. Batista et al., 1999; Buneo et al., 2002; Cui and Andersen, 2007, 2011; Hwang et al., 2012). While it is tempting to label these experiments as being performed in MIP, it was defined by myeloarchitecture and covers only a part of the medial bank of the IPS (Colby et al., 1988; Lewis and Van Essen, 2000).
The anterior intraparietal area (AIP)
Murata et al., 2000 recorded activity for grasping objects from the anterior intraparietal area (AIP), a region just anterior to LIP in the lateral bank of the IPS. They found a variety of cell types that were selective for visual, visual-motor or motor aspects of the task. The visual cells were selective for the shape, size and orientation of three dimensional objects whereas the motor cells appeared to be selective for the grip shape, size and orientation. Visual-motor cells were selective both for vision of the object and grip parameters. Figure 3c shows an example of a visual-motor neuron that was selective for a vertical plate. Baumann et al., 2009 studied the context dependence of power and precision grasps in AIP. When the object was shown first, cells coding both grip types became active, but when the grip type was instructed first the signals were weak, implying that AIP cells code context but the context signal is weak if there is not an accompanying visual object.
Whether AIP is the only PPC area selective for grasps has been recently challenged. Fattori et al., 2012 found area V6A, which had previously been reported to have reach selective neurons, also had a substantial number of cells selective for objects and grasp. Similarly, it has recently been reported that within the population of AIP neurons, besides grasp selectivity, there is also reach selectivity (Lehmann and Scherberger, 2013).
Dorsal Brodmann’s area 5 (area 5d)
Buneo and Andersen, 2006 reported a gradient in the transition zone between Brodmann’s area 5 and PRR with area 5 representing targets more toward hand coordinates and PRR more toward eye coordinates. In subsequent studies this area has been referred to as dorsal area 5 (area 5d, Cui and Andersen, 2011). Whereas PRR represents both potential reaches prior to the decision to reach as well as the reach plans after the decision (Klaes et al., 2011, Cui and Andersen, 2007), area 5d represents only the reach plan after the decision (Cui and Andersen, 2011). This result indicates that area 5d is downstream of PRR for the reach decision-making process.
Patient studies, outlined above, point to the PPC as a region important for online control. Online control using just sensory feedback is generally too slow for rapid online adjustments due to sensory delays. Thus it has been hypothesized that PPC may also utilize efference copy signals of ongoing movements, possibly originating from motor and premotor cortex, to produce an accurate estimate of the state of the limb during reaching (figure 4a). Mulliken et al., 2008 examined the timing of information regarding the current state of the limb in a joystick task. Recordings were performed in the general region of area 5d. It was found that some neurons predicted the instantaneous movement vector before the movement, others after the movement, but most indicated the instantaneous direction without any lag (0 msec, Figure 4b). Thus the area 5d neurons appear to represent by and large the current state of the limb with the aid of a forward model derived from efference copy.
Figure 4. State estimation in area 5.

a. Model of posterior parietal cortex sensorimotor integration. Sensory signals arrive with a delay, approximately 90 ms for visual inputs and 30ms for proprioceptive inputs. An efference copy signal of movement commands arrives with no delay. Outputs include a movement-goal location estimate and a dynamic estimate of the current movement state. b. Histogram of optimal lag times for area 5 cells sensitive to the movement angle. The lag times are the times at which the cells best estimate the instantaneous movement angle. Overall cells show a distribution of lag times that are centered around 0 ms, consistent with the forward state estimation hypothesis. Modified from Mulliken et al. 2008.
Corticocortical connections of PRR, LIP, AIP and area 5d
Similar to PPC the frontal cortex has segregated functions with FEF largely involved with saccades, PMd with reaches, and PMv with grasp. This segregation can also be seen in the corticocortical connectivity—for reach the strongest connections are with PRR, V6A, and area 5d in the PPC with PMd in frontal cortex; for saccades LIP and V6A with FEF; and for grasp AIP with PMv (Tanne-Gariepy et al., 2002; Andersen et al., 1990; Bakola et al., 2010; Borra et al., 2008).
Within the superior parietal lobe there are extensive connections between the reach, saccade and grasp selective regions. The medial bank of the intraparietal sulcus, probably overlapping with MIP, is heavily connected with the medial dorsal parietal area (MDP) in the medial bank of the parieto-occipital sulcus, which appears to overlap with area V6Ad (Blatt et al., 1990; Gamberini et al., 2009). LIP projects to a more ventral location in the parieto-occipital area (PO, Colby et al., 1988) which likely overlaps with V6Av (Passarelli et al., 2011). This segregation of connections is consistent with physiological and connection studies which attribute V6Ad being more somatomotor and V6Av more visual sensory (Passarelli et al., 2011). AIP is connected to an area in the caudal aspect of the lateral bank of the intraparietal sulcus (CIP) which integrates perspective and disparity cues of surface orientation and could provide useful visual signals for grasp (Tsutsui et al., 2001; Borra et al., 2008). AIP is connected to the rostral aspect of LIP which is involved in eye movements, as well as two reach related areas in the PPC, MIP and V6A (Borra et al., 2008). The caudal most third of the convexity of the SPL, extending onto the medial aspect of the hemisphere (PEc) has very strong connections with an area just dorsal to MIP (MIPd) (Bakola et al., 2010). PEc appears to overlap considerably with area 5d. It is also connected to V6A (Bakola et al., 2010). In summary, the reach related areas in PPC are heavily interconnected and the reach, grasp and eye movement areas also connect with one another. Thus there is ample anatomical evidence for potential pathways between these areas for the coordination of complex behaviors such as reach-to-grasp and eye-hand coordination.
The sensory inputs to cortical areas within PPC also vary and provide clues to their function. LIP and AIP are unique in having connections with inferotemporal cortical regions and provide a direct link between the ventral visual pathway for object recognition and the dorsal pathway for object-based actions such as grasp or scanning eye movements (Blatt et al., 1990; Borra et al., 2008). Both areas also receive input from a number of extrastriate visual areas, as does V6Av. Area 5d receives direct inputs from somatosensory areas (Area 5 and Area 2) as well as visual inputs from V6A. These inputs, along with connections to PMd, could provide sensory and efference copy signals necessary for area 5d’s proposed role in state estimation (Fig. 4a).
Coordinate frames
The PPC is thought to play a role in coordinate transformations. As mentioned above, visual targets need to be converted from eye coordinates to muscle coordinates to make movements. In between these two ends of the spectrum, there is evidence from recording and behavioral experiments for many other representations including head, shoulder, trunk and hand centered reference frames. Determining the reference frames used by cortical areas in PPC of monkeys provide clues to the level of disruption that occurs with OA.
Since different cortical areas of PPC are uniquely active for different actions, it might be expected that these areas would be encoding stimuli in action coordinates; that is, LIP in eye coordinates for saccades, and PRR, AIP and area 5d in limb coordinates for reach and grasp. Surprisingly LIP, PRR and AIP all appear to encode targets and actions mostly in eye coordinates (Batista et al., 1999; Andersen et al., 1985, 1990; Lehmann and Scherberger, 2013). Only area 5d has been found to code in predominantly limb coordinates (Bremner et al., 2012). Other response fields in these areas are intermediate between coordinate frames; these examples appear to code simultaneously in eye and hand coordinates (Buneo et al., 2002) and may reflect intermediate stages in the coordinate transformation process. “Gain fields” are also present in all these areas. Gain fields describe the multiplicative or non-linear additive interaction between sensory signals (e.g. visual signals) and signals coding body-part positions (e.g. eye position). They have been proposed to provide a mechanism for coordinate transformations (Andersen et al., 1985; Zipser and Andersen, 1988; Salinas and Thier, 2000).
As mentioned above, OA appears to be largely represented in eye coordinates as well (Buxbaum and Coslett, 1997; Khan et al., 2005; Dijkerman et al., 2006, Blangero et al., 2010). Deficits in other coordinate frames (Jax et al., 2009) may involve areas such as area 5d that code in other coordinate frames, or may result from interference with the coordinate transformation process. Buneo et al., 2002 proposed that reaching may often be achieved by a direct method in which the seen location of the target, in eye coordinates, is subtracted from the seen location of the hand, also in eye coordinates. Such a framework would not require transformation into other coordinate frames and thus would be direct. In the case of an unseen hand, then the proprioceptive signals of the unseen hand would need to be transformed into eye coordinates for this direct process. Interestingly, proprioceptive deficits have been reported for OA patients (Blangero et al., 2007 Pisella et al., 2009, Jackson et al., 2009) and the proprioceptive deficits also appear to be in eye coordinates (Blangero et al., 2007).
Human functional anatomy
Functional magnetic resonance (fMRI) studies of PPC
A natural avenue to study functional specialization in human PPC has been to utilize fMRI in healthy human subjects. Experiments have been designed asking the subject to saccade, reach, point, or grasp within the magnet. The results have in some cases emphasized functional specialization, similar to the monkey studies, and in other cases have found less functional specialization than the monkey studies. This literature has been previously reviewed (Crawford and Vesia, 2012; Culham et al., 2006; Filimon, 2010) and only parts that pertain to OA will be discussed here.
Early fMRI studies reported reach, eye movement and grasp specific areas in human PPC (Connolly et al., 2003; Astafiev et al., 2003; Prado et al., 2005; Culham et al., 2003). Other fMRI studies emphasized the overlap in areas active for eye and limb movements (Levy et al., 2007; Beurze et al., 2009; Hinkley et al., 2009) and speculated that PPC might be differently organized in humans and non-human primates (Hinkley et al., 2009). Vesia and Crawford, 2012 entertained two possibilities to explain the apparent discrepancy between animal and human studies: 1) traditional fMRI may be difficult at detecting differences in activation within an area, and 2) since the differences in activity in animals is relative and not absolute, fMRI may not be sensitive enough to distinguish between areas. The more recent use of multivoxel pattern analysis has revealed greater functional specificity than found in earlier studies using signal amplitude (Gallivan et al., 2011a, b). Using this technique, a qualitatively similar level of functional localization within PPC was found for humans when compared to non-human primates. Reach, grasp, and saccade related areas in the human PPC will be described next using results derived largely from fMRI experiments but also from MEG and TMS studies.
Reach
A complex of reach related areas have been found within the PPC (Figure 5). In general these areas are located within two regions. The medial region includes the precuneous (PrC), posterior intraparietal sulcus (pIPS) at the caudal end of the IPS (Gallivan, 2011a), parieto-occipital junction (POJ) located on both banks of the parieto-occipital sulcus (Prado 2005), and superior parietal occipital cortex located medial and directly anterior to the parieto-occipital sulcus (SPOC). The more lateral region is located on the medial bank of the (mIPS, Prado, 2005) and the middle IPS half way along the length of the IPS in the medial bank (midIPS, Gallivan, 2011a). There is some degree of overlap within both the medial and lateral regions defined by different investigators. Activations generally involve both hemispheres (Prado et al., 2005).
Figure 5. Action specificity in human posterior parietal areas.

Lesion sites in optic ataxia and action specific sites for reaching or pointing, eye movements, and grasping. The yellow lines demarcate the central (CS) and intraparietal sulci (IPS). L, R, D, V are left, right, dorsal, ventral. The Talairach coordinates of these sites, reported in Culham et al. 2006, were translated to the Montreal Neurological Institute template, and then visualized using BrainNet Viewer (Xia et al. 2013). Not all sites are clearly visible in the lateral view due to the convolutions of the brain. The numbers in the superior view indicate the original studies that identified these sites: 1) Connolly et al. 2003, 2) Astafiev et al. 2003, 3) Prado, et al. 2005, 4) Grefkes et al. 2004, 5) Petit et al. 1999, 6) Sereno et al. 2001, 7) Medendorp et al. 2003, 8) Binkofski et al. 1998, 9) Culham et al. 2003, 10) Frey et al. 2005, 11) Karnath & Perenin 2005.
Early studies revealed an area on the medial surface of the superior aspect of the PPC that was selective when subjects planned to point as opposed to making saccades and this area was proposed to be the homolog of PRR (Connolly et al., 2003). Astafiev et al., 2003 similarly found a band of pointing related activity from SPL to precuneus, generally in the region reported by Connolly et al. and also proposed the area of activation to be homologous to PRR. Prado et al., 2005 described an area activated for reaching in foveal vision in the medial IPS and a second region that was activated for peripheral reaching in the parieto-occipital junction (POJ), an area roughly similar in location to the region activated by pointing in Connolly et al. and Astafiev et al. The POJ area overlaps with the core site that includes the precuneus and POJ shown to be involved in OA (Figure 5, Karnath and Perenin, 2005, Prado et al., 2005). Since OA largely produces errors in peripheral reaching, it has been proposed that there are two reach systems for reaching, one involved in central reaches and another, more extensive one for peripheral reaches and that OA can result from damage to the peripheral reaching system comprising POJ (Prado et al., 2005). Reach without a grasp component activates two foci within SPOC, one anterior to the POS in the precuneus medial to the IPS and a second posterior to the POS in the cuneus (Cavina-Pratesi et al., 2010). Two regions within the medial PPC also appear to receive different modalities of sensory feedback (Filimon et al., 2009). One is located anteriorly in the precuneus extending into mIPS is active for reaches with or without visual feedback whereas a more posterior region at the superior aspect of POS is more active for reaches when visual feedback is available. It was proposed that the anterior region receives proprioceptive signals to guide reaching whereas the more posterior region relies more on visual feedback. Using a multivoxel pattern analysis on fMRI data Gallivan et al., 2011a were able to decode intended effectors for reaches and saccades and the directions of these intended movements from the PPC. The intent predictions for reaches were made from the superior parieto-occipital cortex (SPOC) posterior IPS and mid IPS. Effector specificity has also been reported with magnetoencephalography (MEG) with a medial aspect of PPC being selective for hand movements and a more central part being selective for eye movements (Van Der Werf et al., 2010). Pitzalis et al., 2013 mapped a retinotopic region in the dorsal aspect of the POS with an over-representation of the contralateral lower visual field and periphery. This area was activated by pointing movements. Based on its location with respect to other proposed homologues of human V3a and V6, and its similar retinotopic and limb movement sensitivity with V6A in monkeys, they proposed this area to be the human homolog of V6A. This region falls on the posterior extent of the human parietal reach region proposed by Connolly et al., 2003 and appears to overlap with some of the medial PPC reach areas described by others.
In an fMRI study Heed et al., 2011 reported a segregation of activation between eye and hand movements but not between hand and foot movements. They hypothesized that PPC is organized with respect to function (look versus reach) rather than effector. Support for this idea comes from the study of an OA patient who showed a hand effect but also a foot effect when reaching with either the hand or foot into the contralesional field (Cavina-Pratesi et al., 2013) and functional imaging of observed motor acts which found clustering according to the type of motor act and but not effector in AIP (Jastorff et al. 2010).
Transcranial magnetic stimulation (TMS) over PPC in humans produces errors in adjusting to jumped targets with the contralateral limb (Desmurget et al., 1999). In fact, this study was one of the first to propose the online visuomotor function for PPC. A subsequent TMS study delivered TMS over SPOC and a site close to the IPS over the angular gyrus (AG) (Vesia et al., 2010). Stimulation of the AG/IPS site produced increased endpoint variability for reaches and decreased accuracy for contralateral saccades. Stimulation of SPOC produced deviation of endpoint errors toward the fixation point and no effect on saccades. Visual feedback of the limb cancelled the effect of AG/IPS stimulation but not SPOC. The investigators interpreted these results as demonstrating the first causal evidence for functional specificity within PPC of human with the AG/IPS region containing overlapping maps for eye and hand movement plans and the SPOC region specialized for peripheral goals. The latter result is consistent with the site of OA lesions (Karnath and Perenin, 2005) and fMRI activation for peripheral reach targets (Prado et al., 2005). Inouchi et al., 2013 recorded Bereitschaftgspotentials (BP) associated with reaching from subdural ECoG grids implanted for clinical evaluation of epilepsy. They found BPs in more medial aspects of the PPC in the IPS, SPL, and precuneus. Interestingly electrical stimulation of some of these BP sites produced imprecision in reaching but no other limb movements and only for peripheral reaches, reminiscent of OA.
Homologies between monkey and human reach areas have been proposed by a number of investigators. As mentioned above, several groups have assigned activation zones to be the human equivalent of PRR. Some have proposed more fine grain assignments with mIPS homologous to MIP (Prado et al., 2005, Filimon, 2010) and the medial PPC complex with V6A (Prado et al. 2005, Cavina-Pratesi et al., 2010, Filimon 2010, Pitzalis et al., 2013). Interestingly OA -like deficits including misreaching to peripheral targets are produced with inactivation within the IPS around MIP/area 5v in monkeys which would argue for MIP being within the more medial complex of reach areas (Karnath and Perenin, 2005; Prado et al., 2005). It would be important to know if inactivations of V6A in monkeys also produce OA. Caution has been raised regarding assigning homologies and some of the proposed homologies have only been tentatively assigned (Culham et al., 2006; Konen et al., 2013). Care is particularly warranted given that subregions of PRR may diverge during evolution and that there is a disproportionate expansion of PPC in humans (Culham et al., 2006).
Grasp
A prominent model for reach-to-grasp movements has proposed 2 channels, one for the transport component (reach) and one for grasp (Jeannerod et al., 1995; Cavina-Pratesi et al., 2010; Vesia et al., 2013). Culham et al., 2003 found greater fMRI activation for grasping than reaching bilaterally within the anterior aspect of the IPS (aIPs) and proposed that this area was the human homologue of AIP (Figure 5). Cavina-Pratesi et al., 2010 designed a task in which the transport (reach) was dissociated from grasp. With grasp they found activity in aIPS and with reach in SPOC. These results provide an anatomical basis for the support of the 2 channel hypothesis. With a more sensitive pattern recognition analysis Gallivan et al. 2011b found that grasps could be decoded from the contralateral dorsomedial PPC including SPOC, precuneus, and midIPS. This finding is consistent with a relative rather than absolute specificity of function and may reflect the need to coordinate reaching and grasping. Consistent with a greater primacy of aIPS for grasp and SPOC for reach/transport Vesia et al., 2013 showed with paired pulse TMS greater functional interaction between M1 and aIPS during grasp and between M1 and SPOC during transport. Multivoxel pattern recognition also found that reaching with the contralateral and ipsilateral limb can be separately decoded (Gallivan et al., 2013) Recently Konen et al., 2013 described a gradient between reaching and grasping activation, with greatest reach activation located in the precuneus and SPL and grasp activity greatest in the anterior IPS near the junction with the postcentral sulcus. They labeled these areas the human parietal grasp region (hPGR) and the human parietal reach region (hPRR) but were careful in not drawing a direct homology to AIP and PRR in monkey.
Saccade
An area active for saccades has been reported within the IPS at its middle of the anterior-posterior extent of the IPS (Petit and Haxby, 1999; Sereno et al., 2001; Medendorp et al., 2003; Figure 5) in the medial bank of the IPS (Culham et al., 2006). Konen and Kastner, 2008 described a gradient for eye movements within the PPC with greatest activity in an area they labeled superior parietal lobule 1 (SPL1). Saccade activations were also reported in topographic intraparietal areas 1 and 2.
Pharmacological inactivation
Areas in PPC can be inactivated pharmacologically, typically with muscimol or other GABA agonists. There are many advantages to this technique. The inactivation area can be relatively well confined to individual anatomically and functionally defined modules. The short time-course of the inactivations avoids long term compensation and the repeatability of inactivations allows for more refined statistical analyses. Pharmacological inactivations do not affect fibers of passage, simplifying the interpretation of what neural substrate is being affected. The results of these studies can also suggest how different modules within PPC work together when more complicated behaviors such as eye-hand coordination are examined.
PRR inactivation produces OA
Hwang et al., 2012 investigated the effect of inactivating PRR in two non-human primates. The PPR in each animal was mapped using single cell recordings prior to commencing with the inactivations. Figure 6a shows the location of inactivation in one of the animals in a coronal MRI slice within the medial bank of the IPS. The inactivation site may have involved MIP and area 5v, adjoining areas within the medial bank of the IPS. The animals exhibited increased errors for reach movements to visual stimuli but not for saccades to the same stimuli (Figure 6b). Similar to human OA the errors were seen for extrafoveal but not foveal targets (Figure 6c). Interestingly, there was a field effect for contralesional targets for one animal but ipsilesional for the other. However, a population analysis of the response fields of the PRR neurons at the injection sites correlated with the field of inactivation; that is the contralateral deficit had largely contralateral response fields at the injection site and vice versa for the ipsilateral deficit.
Figure 6. Optic ataxia after parietal reach region inactivation.

a. Coronal MRI slice through the injection site in PRR in monkey Y. The injection site appears bright due to injection of the contrast agent gadolinium. The dotted blue line indicates the intraparietal sulcus. b. Movement amplitudes for reaches and saccades in control (black) and inactivation (purple) sessions. The reach amplitudes are hypometric but saccades are unaffected. c. Reaches to extrafoveal targets are affected but not reaches to foveal targets. Modified from Hwang et al. 2012.
LIP inactivation produces eye movement and attention deficits
Li et al., 1999 showed effects on saccadic eye movements after muscimol injection. Memory saccades toward the contralesional space became hypometric, and in one animal became slightly hypermetric for the ipsilesional space. The metrics of saccades to visual targets were unaffected. Thus the deficit resembled OA, but for eye movements and only for memory saccades. The latencies to onset of both visual and memory saccades toward the contralesional space were increased and in many instances showed increased variance.
Wardak et al., 2002, using muscimol inactivation, found that monkeys chose ipsilesional over contralesional targets (so-called extinction) and increased search time for contralateral targets in a search task. They did not observe any eye movement deficits for single target visual or memory guided saccades. Liu et al. 2010 reported that inactivations of the dorsal aspect of LIP increased saccade latencies and that inactivations of ventral LIP increased both saccade and search latencies.
Wilke et al., 2012 inactivated LIP (Figure 7a) while measuring fMRI activity during a choice task between bilateral saccade targets—one in the contralesional and one in the ipsilesional field (Figure 7b). Monkeys chose the ipsilesional targets more frequently (Figure 7c). Interestingly, in those cases in which the animals made choices in the contralesional field, the activity in parieto-temporal and frontal areas in both hemispheres increased. This result only partially agrees with a competition model for extinction/neglect, which would predict only an increase in the lesioned hemisphere. Rather the finding suggests an additional component of interhemispheric cooperation. One possibility is an overall increase in general effort that affects both hemispheres. Consistent with this possibility are findings of extinction produced by pulvinar inactivation (Wilke et al., 2013). An increase in the reward associated with the contralesional hemifield greatly reduces the bias toward the ipsilesional field. This effect cannot be explained by increased salience alone since increasing the contrast of the contralesional target had only a modest effect. Since LIP has also been shown to play a role in decision making (Shadlen and Newsome, 1996; Cui and Andersen, 2007) and the contralesional hemifield is the most represented hemifield within LIP (Andersen et al., 1990), it is possible that lesion to LIP may reduce the choice of saccades into the contralesional space due to increased cost/effort as a result of the lesion.
Figure 7. Extinction after lateral intraparietal area inactivation.

a. Coronal MRI slice through the injection site in LIP of monkey F. b. Schematic of the choice task. The animal is free to choose between targets in the ipsilesional and contralesional visual fields. The red square is the fixation point and purple squares the two saccade targets. c. Percent of choices into the ipsilesional field. In the control condition (black) the animal chooses nearly equally between fields but in the inactivation condition (purple) the animal is biased toward the ipsilesional target. Modified from Wilke et al. 2012.
Online control
In order to test online control by PPC, Battaglia-Mayer et al., 2013 made bilateral inactivations in the region of area 5d and tested the effect on the jumped target task. They observed an increase in the time required to make the correction leading to an elongation of the hand path toward the initial target location. They interpreted their finding as a deficit in online control similar to that observed in OA. This result is also consistent with disruption of a forward model or state estimate of the limb (Mulliken et al., 2008).
Hand-eye coordination
With evidence for modules for hand and eye movements (Snyder et al., 1997, Andersen and Buneo, 2002) and deficits for hand-eye coordination after PPC lesions (Gaveau et al., 2008), an obvious possibility is that the hand and eye modules work together through interactions during eye-hand coordination either within the PPC or at a location downstream in frontal cortex. Moreover, LIP and PRR are directly connected with one another and may share connections with other cortical areas (Blatt et al., 1990).
Dean et al., 2012 reported a population of neurons that are active coherent with beta band LFPs, and their activity predicts coordinated eye-hand movements but not eye movements alone. Similar beta band LFP activity is seen in PRR and they proposed that the coherent activity in PPC acts to coordinate eye-hand movements. Yttri et al., 2013a saw increases in reaction times for coordinated eye-hand movements with LIP inactivation but not for reaches alone. They interpreted the findings as LIP being involved in saccade planning and not reach planning, but when saccades are delayed an eye-hand coordination mechanism delays the reach until after the saccade is made. They also found only increases in reaction time for reaching with the contralesional limb after PRR inactivation with no effect on eye-hand coordination (Yttri et al., 2013b). They interpreted these results as PRR being only involved in limb movements and PRR is upstream from any eye-hand coupling mechanism. One cautionary note is that the reaction time deficits in these two studies were very small (under 10 ms). The fact that greater deficits are seen in other studies, i.e. larger reaction times for LIP (Li et al. 1999) and misreaching with PRR inactivation (Hwang et al., 2012), may be due to differences in the size of the inactivation or a difference in location within the IPS (Yttri et al., 2013b). It is possible that a more behaviorally effective inactivation would show coupled eye-hand effects.
AIP inactivation
Gallese et al., 1994 reported that inactivation of AIP interfered with grasp, particularly the preshaping of finger postures. They did not observe reach deficits and they proposed that AIP is important for visually guided grasping. AIP has strong connections to the ventral premotor cortex (PMv, Luppino et al., 1999). Inactivations of PMv produce grasp deficits (Fogassi et al. 2001). Taken together, these results suggest a parietofrontal circuit for grasping (Fogassi et al., 2001; Luppino et al., 1999). Battaglini and colleagues, 2002 found that permanent lesion of V6A produced deficits in reach, wrist orientation, and grasping, consistent with V6A playing a role in reaching and grasping (Fattori et al., 2012).
Synthesis
Balint’s syndrome and OA are the result of disruption of many functional cortical modules
As shown in Figure 8 the visuomotor component of optic ataxia may be traced to PRR at least in monkey (Hwang et al. 2012). The inactivations produce peripheral but not foveal reach deficits and do not produce saccade deficits. This area in monkeys has an eye centered map of visual space (Batista et al. 1999) which agrees with the reference frame most often reported for OA in humans (Dijkerman et al., 2006; Blangero et al., 2010).
Figure 8. Proposed functional segregation within posterior parietal cortex.

Area PRR inactivation produces optic ataxia, LIP inactivation eye movement deficits and extinction, area 5d inactivation online deficits consistent with disruption of state estimation, and AIP grasp deficits. These inactivation results and recording data suggest specialization of PRR for reach, LIP for eye movements and attention, area 5d for state estimation, and AIP for grasp.
The contralesional/neglect component of Balint’s syndrome may include areas like LIP where extinction and visual search deficits are found after LIP inactivation (Wardak et al., 2002, 2004; Wilke et al., 2012, Liu et al., 2010, Fig. 8). The eye movement difficulties found in Balint’s syndrome and reported occasionally for OA may also be a result of lesion to an area like LIP, where saccade deficits are found with inactivation (Li et al., 1999, Liu et al., 2010). In humans, this area may include SPL1 and IPS1,2 (Konen and Kastner, 2008). The simultagnosia seen in Balint’s syndrome may also be an attentional deficit, but at a more object based level which may result from involvement of other cortical areas.
Online estimates of the current state of the limb are found in the activity of area 5d neurons (Mulliken et al., 2008). Lesions in this area produce deficits consistent with online control (Battaglia-Mayer et al., 2013). Thus a module similar to area 5d may be damaged in OA in humans to account for online control deficits.
Area AIP contains cells active for different grasp postures (Murata et al., 2000; Baumann et al., 2009) and lesion of this region in monkey produces a grasp deficit. However, grasp activity (Fattori et al., 2012) and deficits in hand postures after lesion (Battaglini et al., 2002) have been reported also for area V6A. fMRI studies point to the anterior aspect of IPS as an area preferentially activated by grasping (Culham et al., 2003; Konen et al., 2013). Lesions to PPC that produce grasp deficits in combination with other aspects of Balint’s syndrome including OA may be due to lesions that include anterior IPS.
Increased effort as a possible root cause for OA and extinction
The effects of OA often present as hypometria in the affected field in humans (Blangero et al., 2010). Similarly inactivations in monkeys also produce hypometria for reaches for PRR (Hwang et al., 2012) and eye movements for LIP (Li et al., 1999). These deficits may be a result of reduced effort. The extinction effects may also be overcome by increased effort as suggested by increased fMRI activations in both hemispheres with compensation (Wilke et al., 2012). The ability to negate a good deal of the extinction by manipulating reward (Wilke et al., 2013) may reflect overcoming costs (effort) with increased reward. These results suggest that training, reward manipulation, or electrical stimulation may promote repair. For instance, electrical stimulation of the ipsilesional dorsal pulvinar, in a manner similar to deep brain stimulation for movement disorders, may help to alleviate the symptoms of neglect by producing activation in frontal and occipito-temporal circuits connected to PPC reach and eye movement areas.
A complex of reach-related areas in humans and non-human primates
Although homologies within the complex of reach related areas in humans and non-human primates is still not clear, further studies with inactivations in non-human primates may advance understanding of similarities. Based on human lesion and imaging results, Pisella et al. 2009 proposed that the field effect is due to lesions of POJ and the hand effect to lesion of mIPS. Vesia and Crawford, 2012 proposed that SPOC specifies reach goals in visual coordinates and mIPS provides information on hand position to calculate the reach vector. This functional proposal would also predict a hand effect being derived from mIPS and a field effect from SPOC. In the interest of establishing homologies, it would be informative to see if inactivation of PRR would produce a hand effect. Also of interest and related to the field effect, Hwang et al. 2012 showed contralateral and ipsilateral field effects for small inactivations of PRR. However, human lesions likely take out entire cortical areas. Would a larger inactivation of PRR produce a primarily contralateral field effect as is seen with human subjects and be explained as a bias toward contralateral space within the population of PRR neurons?
Interaction between modules for eye-hand coordination and reach-to-grasp
The inactivation studies in animals reviewed here are making important inroads into the functional parcellation and interaction between modules that comprise the PPC. As mentioned above, the inactivation of PRR produces a specific deficit in reach accuracy but not saccades. This deficit is also restricted to reaches to targets in the peripheral visual field, reproducing the peripheral field deficit found in OA patients.
The specificity of the deficit for reaches and not saccades is of interest since PRR, although heavily biased toward reach related activity, also contains some activation for saccades. One view would be that although PRR is reach-specific, reaching is usually coordinated with eye movements. Thus it is not surprising that eye movement information is also present in PRR. Moreover, the hypometria for reaches, and the specificity for peripheral targets is consistent with PRR being important for allowing reaches to be performed independently of saccades, an often overlooked aspect of eye-hand coordination. A similar argument has been made for cases of magnetic misreaching in human OA (Jackson et al., 2005).
This same issue of specificity can be raised for grasping and for saccades. Although a grasp deficit has been reported for AIP (Gallese et al., 1994), this study was rather qualitative and would not necessarily register reach deficits. Since grasp and reach selectivity are found in AIP neural activity (Lehmann and Scherberger, 2013) it would be informative to perform inactivation studies in which the reach-to-grasp movement kinematics are precisely recorded. A considerable literature suggests that reach and grasp are processed separately. However, reach and grasp activity has been recorded in V6A (Fattori et al., 2012) and both reach and grasp deficits have been reported with permanent lesion of V6A.
Another open question is how areas become engaged for coordinated movements. Evidence exists that LIP and PRR interact during eye-hand coordination (Dean et al., 2012; Yttri et al., 2013). Although inactivation of PRR does not affect eye movements alone, would errors be seen for saccades during eye-hand coordination due to interactions between the areas? Would inactivation of AIP produce reach deficits only for reaches in reach-to-grasp movements but not for reaches alone (and vice versa for PRR inactivations)?
In order to understand how the functional modules of the PPC may interact with one another and support complex coordinated behaviors, it is essential to delineate the circuit connections between different PPC modules. With cutting-edge genetic and molecular techniques such as optogenetics, neural circuits have begun to be dissected with unprecedented specificity. These advanced techniques are mainly working in simpler animal models at present, but translation to primate models is gradually taking place (Han et al., 2009; Diester et al., 2011). A particularly useful way to determine the functional role of inputs from one area to another is activating or inactivating projection neurons selectively. Such projection-specific intervention can be achieved by injecting anterograde viruses carrying opsin genes in the upstream area and delivering light in the downstream area or by injecting retrograde viruses in the downstream area and delivering light in the upstream area (Gradinaru et al., 2009; Tye et al., 2011; Lammel et al., 2012). Because opsins can be quickly switched on and off by light, the effect of optogenetic intervention shows a high-temporal precision. Furthermore, projection neurons can be identified by extracellular recording in the upstream area during antidromic photo-stimulation in the downstream area, and thus the response properties of projection neurons can be selectively characterized (Gradinaru et al., 2009; Jennings et al., 2013).
Establishing homologies using similar recording modalities in humans and non-human primates
One method of establishing homologies between humans and non-human primates would be to perform fMRI studies with both species using the identical experimental protocols. This approach was used by Kagan et al., 2010 to examine processing of saccadic eye movements in putatively homologous areas of the parietal, frontal and temporal lobes. They found that even for such a basic movement, there exist species differences with the monkey areas showing much greater contralateral preference than the human areas.
Single cell recordings from electrode arrays in humans will promote a detailed understanding of PPC and help to bridge the research that has been performed in human and non-human primates. Experiments in monkeys show that intentions can be decoded for action and potentially similar signals can be used in neuroprosthetic applications in patients with paralysis (Musallam et al., 2004; Mulliken et al. 2008; Hauschild et al., 2012). Such neuroprosthetic research in humans will be synergistic with understanding the basic science and circuitry of PPC. Moreover, these efforts can establish whether similar areas and circuits exist in both species. For instance, are cells in AIP of humans selective for hand postures for grasping? Such studies can establish the degree of effectors specificity by asking the patient to imagine moving different effectors. Are the cells selective for imagining a reach independent of imaging a reach with the foot or the hand as has been suggested by some and thus it is the concept of reaching that is being encoded in reach selective areas? Or will there be a high degree of specificity for effectors, with imagined movements being selective for left and right arm and left and right leg? Similarly, how detailed will kinematics be encoded? Will there be cells selective for particular degrees-of-freedom, such as pronation-supination and flexion/extension around the wrist?
In summary a rich knowledge of the functional organization of PPC for action planning is emerging in both humans and NHPs. fMRI studies in NHPs and electrophysiological recordings in humans promise to bridge the gap in understanding of similarities and differences between the two primate species.
Highlights.
Balint’s syndrome and optic ataxia result from disruption of many cortical modules.
A complex of reach areas exists in the posterior parietal cortex.
Interaction between modules appears to facilitate coordinated movements.
Homologies of the cortical modules found in non-human primates may exist in humans.
Acknowledgments
We wish to thank the National Eye Institute and the Boswell Foundation for support.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
Richard A. Andersen, Email: andersen@vis.caltech.edu.
Kristen N. Andersen, Email: KNAndersen@mednet.ucla.edu.
EunJung Hwang, Email: eunjunghwang.phd@gmail.com.
Markus Hauschild, Email: markus@vis.caltech.edu.
References
- Amin H, Ehrreich S, Kelly J, Lebby P, Hutchinson H. Balint syndrome in the pediatric population: A case report of three patients. J Neurol. 2012:P02.179. [Google Scholar]
- Andersen RA, Essick GK, Siegel RM. The encoding of spatial location by posterior parietal neurons. Science. 1985;230:456–458. doi: 10.1126/science.4048942. [DOI] [PubMed] [Google Scholar]
- Andersen RA, Essick GK, Siegel RM. Neurons of area 7 activated by both visual stimuli and oculomotor behavior. Exp Brain Res. 1987;67:316–322. doi: 10.1007/BF00248552. [DOI] [PubMed] [Google Scholar]
- Andersen RA, Bracewell RM, Barash S, Gnadt JW, Fogassi L. Eye position effects on visual, memory and saccade-related activity in areas LIP and 7A of macaque. J Neurosci. 1990;10:1176–1196. doi: 10.1523/JNEUROSCI.10-04-01176.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersen RA, Buneo CA. Intentional maps in posterior parietal cortex. Ann Rev Neurosci. 2002;25:189–220. doi: 10.1146/annurev.neuro.25.112701.142922. [DOI] [PubMed] [Google Scholar]
- Astafiev SV, Shulman GL, Stanley CM, Snyder AZ, Van Essen DC, Corbetta M. Functional organization of human intraparietal and frontal cortex for attending, looking, and pointing. J Neurosci. 2003;23(11):4689–4699. doi: 10.1523/JNEUROSCI.23-11-04689.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayuso-Peralta L, Jimenez-Jimenez FJ, Tejeiro J, Vaquero A, Cabrera-Valdivia F, Madero S, Cabello A, Garcia-Albea E. Progressive multifocal leukoencephalopahty in HIV infection presenting as Balint’s syndrome. Neurology. 1994;44(7):1339–40. doi: 10.1212/wnl.44.7.1339. [DOI] [PubMed] [Google Scholar]
- Bakola S, Gamberini M, Passarelli L, Fattori P, Galletti C. Cortical connections of parietal field PEc in the macaque: linking vision and somatic sensation for the control of limb action. Cereb Cortex. 2010;20:2592–2604. doi: 10.1093/cercor/bhq007. [DOI] [PubMed] [Google Scholar]
- Balint R. Seelenlahmung des “Schauens”, optische Ataxie, raumliche Storung der Aufmerksamkeit. Monatsschr Psychiatr Neurol. 1909;25:51–81. [Google Scholar]
- Batista AP, Buneo CA, Snyder LH, Andersen RA. Reach plans in eye-centered coordinates. Science. 1999;285:257–260. doi: 10.1126/science.285.5425.257. [DOI] [PubMed] [Google Scholar]
- Battaglia-Mayer A, Ferrari-Toniolo S, Visco-Comandini F, Archambault PS, Saberi-Moghadam S, Caminiti R. Impairment of online control of hand and eye movements in a monkey model of optic ataxia. Cereb Cortex. 2013;23:2644–2656. doi: 10.1093/cercor/bhs250. [DOI] [PubMed] [Google Scholar]
- Battaglini PP, Murzur A, Galletti C, Skrap M, Brovelli A, Fattori P. Effects of lesions to area V6A in monkeys. Exp Brain Res. 2002;144:419–422. doi: 10.1007/s00221-002-1099-4. [DOI] [PubMed] [Google Scholar]
- Baumann MA, Fluet MC, Scherberger H. Context-specific grasp movement representation in the macaque anterior intraparietal area. J Neurosci. 2009;29:6436–6448. doi: 10.1523/JNEUROSCI.5479-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beurze SM, de Lange FP, Toni I, Medendorp WP. Spatial and effector processing in the human parietofrontal network for reaches and saccades. J Neurophysiol. 2009;101:3053–3062. doi: 10.1152/jn.91194.2008. [DOI] [PubMed] [Google Scholar]
- Binkofski F, Dohle C, Posse S, Stephan KM, Hefter H, Seitz RJ, et al. Human anterior intraparietal area subserve prehension: A combined lesion and functional MRI activation study. Neurology. 1998;50(5):1253–1259. doi: 10.1212/wnl.50.5.1253. [DOI] [PubMed] [Google Scholar]
- Blangero A, Ota H, Delporte L, Revol P, Vindras P, Rose G, Boisson D, Vighetto A, Rosetti Y, Pisella L. Optic ataxia is not only ‘optic’, impaired spatial integration of proprioceptive information. Neuroimage. 2007;36:T61–T68. doi: 10.1016/j.neuroimage.2007.03.039. [DOI] [PubMed] [Google Scholar]
- Blangero A, Ota H, Rosetti Y, Fujii T, Ohtake H, Tabuchi M, Vighetto A, Yamadori A, Vindras P, Pisella L. Systematic retinotopic reaching error vectors in unilateral optic ataxia. Cortex. 2010;46:77–93. doi: 10.1016/j.cortex.2009.02.015. [DOI] [PubMed] [Google Scholar]
- Blatt G, Andersen RA, Stoner G. Visual receptive field organization and cortico-cortical connections of area LIP in the macaque. J Comp Neurol. 1990;299:421–445. doi: 10.1002/cne.902990404. [DOI] [PubMed] [Google Scholar]
- Blumenfeld H. Neuroanatomy through Clinical Cases. Sinauer Associates, Inc; 2002. [Google Scholar]
- Borchers S, Muller L, Synofzik M, Himmelbach M. Guidelines and quality measures for the diagnosis of optic ataxia. Front Hum Neurosci. 2013;7:1–13. doi: 10.3389/fnhum.2013.00324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borra E, Belmalih A, Calzavara R, Gerbella M, Murata A, Rozzi S, Luppino G. Cortical connections of the macaque anterior intraparietal (AIP) area. Cerbral Cortex. 2008;18:1094–1111. doi: 10.1093/cercor/bhm146. [DOI] [PubMed] [Google Scholar]
- Bremner LR, Andersen RA. Coding of the reach vector in parietal area 5d. Neuron. 2012;75:342–351. doi: 10.1016/j.neuron.2012.03.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buneo CA, Jarvis MR, Batista AP, Andersen RA. Direct visuomotor transformations for reaching. Nature. 2002;416:632–636. doi: 10.1038/416632a. [DOI] [PubMed] [Google Scholar]
- Buneo CA, Andersen RA. The posterior parietal cortex: Sensorimotor interface for the planning and online control of visually guided movements. Neuropsychologia. 2006;44:2594–2606. doi: 10.1016/j.neuropsychologia.2005.10.011. [DOI] [PubMed] [Google Scholar]
- Buxbaum LJ, Coslett HB. Subtypes of optic ataxia: reframing the disconnection account. Neurocase. 1997;3:159–166. [Google Scholar]
- Buxbaum LJ, Coslett HB. Spatio-motor representations in reaching: evidence for subtypes of optic ataxia. Cognitive Neuropsychology. 1998;15:279–312. doi: 10.1080/026432998381186. [DOI] [PubMed] [Google Scholar]
- Caplan LR, van Gijn J. Stroke syndromes. 3. Cambridge University Press; 2012. [Google Scholar]
- Carmant L, Decarie JC, Fon E, Shevel MI. Transient visual symptoms as the initial manifestation of childhood adrenoleukodystrophy. Pediatr Neurol. 1998 Jul;19(1):62–4. doi: 10.1016/s0887-8994(98)00015-0. [DOI] [PubMed] [Google Scholar]
- Carey DP, Coleman RJ, Della Sala S. Magnetic misreaching. Cortex. 1997;33:639–652. doi: 10.1016/s0010-9452(08)70722-6. [DOI] [PubMed] [Google Scholar]
- Cavina-Pratesi C, Monaco S, Fattori P, Galletti C, McAdam TD, Quinlan DJ, Goodale MA, Culham JC. Functional magnetic resonance imaging reveals the neural substrates of arm transport and grip formation in reach-to-grasp actions in humans. J Neurosci. 2010;30:10306–10323. doi: 10.1523/JNEUROSCI.2023-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavina-Pratesi C, Connolly JD, Milner AD. Optic ataxia as a model to investigate the role of the posterior parietal cortex in visually guided action: evidence from studies of patient M. H. Front Hum Neurosci. 2013 Jul;16:7, 336. doi: 10.3389/fnhum.2013.00336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cogan DG. Ophthalmic manifestations of bilateral non-occipital cerebral lesions. Brit J Opthhal. 1965;49:281–296. doi: 10.1136/bjo.49.6.281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen YE, Andersen RA. A common reference frame for movement plans in the posterior parietal cortex. Nat Rev Neurosci. 2002;3:553–562. doi: 10.1038/nrn873. [DOI] [PubMed] [Google Scholar]
- Colby CL, Gattass R, Olson CR, Gross CG. Topographical organization of cortical afferent to extrastriate visual area PO in the macaque: a dual tracer study. J Comp Neurol. 1988;269:392–413. doi: 10.1002/cne.902690307. [DOI] [PubMed] [Google Scholar]
- Colby CL, Goldberg ME. Space and attention in parietal cortex. Ann Rev Neurosci. 1999;22:819–349. doi: 10.1146/annurev.neuro.22.1.319. [DOI] [PubMed] [Google Scholar]
- Connolly JD, Andersen RA, Goodale MA. FMRI evidence for a ‘parietal reach region’ in the human brain. Exp Brain Res. 2003;153:140–145. doi: 10.1007/s00221-003-1587-1. [DOI] [PubMed] [Google Scholar]
- Crawford JD, Vesia M. Specialization of reach function in human posterior parietal cortex. Exp Brain Res. 2012;221:1–18. doi: 10.1007/s00221-012-3158-9. [DOI] [PubMed] [Google Scholar]
- Cui H, Andersen RA. Posterior parietal cortex encodes autonomously selected motor plans. Neuron. 2007;56:552–559. doi: 10.1016/j.neuron.2007.09.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui H, Andersen RA. Different representations of potential and selected motor plans by distinct parietal areas. J Neurosci. 2011;31(49):18130–18136. doi: 10.1523/JNEUROSCI.6247-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Culham JC, Danckert SL, DeSouza JFX, Gati JS, Menon RS, Goodale MA. Visually guided grasping produces fMRI activation in dorsal but not ventral stream brain areas. Exp Brain Res. 2003;153:180–189. doi: 10.1007/s00221-003-1591-5. [DOI] [PubMed] [Google Scholar]
- Culham JC, Cavina-Pratesi C, Singhal A. The role of parietal cortex in visuomotor control: what have we learned from neuroimaging? Neuropsychologia. 2006;44:2668–2684. doi: 10.1016/j.neuropsychologia.2005.11.003. [DOI] [PubMed] [Google Scholar]
- Dean HL, Hagan MA, Pesaran B. Only coherent spiking in posterior parietal cortex coordinates looking and reaching. Neuron. 2012;23;73(4):829–41. doi: 10.1016/j.neuron.2011.12.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desmurget M, Epstein CM, Turner RS, Prablanc C, Alexander GE, Grafton ST. Role of the posterior parietal cortex in updating reaching movements to a visual target. Nat Neurosci. 1999;2:6. doi: 10.1038/9219. [DOI] [PubMed] [Google Scholar]
- Diester I, Kaufman MT, Mogri M, Pashaie R, Goo W, Yizhar O, Ramakrishnan C, Deisseroth K, Shenoy KV. An optogenetic toolbox designed for primates. Nat Neurosci. 2011;14:387–397. doi: 10.1038/nn.2749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dijkerman HC, McIntosh RD, Anema HA, de Haan EHF, Kappelle LJ, Milner AD. Reaching errors in optic ataxia are linked to eye position rather than head or body position. Neuropsychologia. 2006;44:2766–2773. doi: 10.1016/j.neuropsychologia.2005.10.018. [DOI] [PubMed] [Google Scholar]
- Fattori P, Gamberini M, Kutz DF, Galletti C. Arm-reaching neurons in the parietal area V6A of the macaque monkey. Eur J Neurosci. 2001;13:2309–2313. doi: 10.1046/j.0953-816x.2001.01618.x. [DOI] [PubMed] [Google Scholar]
- Fattori P, Breveglieri R, Raos V, Bosco A, Galletti C. Vision for action in the macaque medial posterior parietal cortex. J Neurosci. 2012;32(9):3221–3234. doi: 10.1523/JNEUROSCI.5358-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filimon F, Nelson JD, Huang RS, Sereno MI. Multiple parietal reach regions in humans: cortical representations for visual and proprioceptive feedback during on-line reaching. J Neurosci. 2009;29:2961–2971. doi: 10.1523/JNEUROSCI.3211-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filimon F. Human cortical control of hand movements: parietofrontal networks for reaching, grasping and pointing. Neuroscientist. 2010;16:388–407. doi: 10.1177/1073858410375468. [DOI] [PubMed] [Google Scholar]
- Fogassi L, Gallese V, Buccino G, Craighero L, Fadiga L, Rizzolatti G. Cortical mechanism for the visual guidance of hand grasping movements in the monkey: a reversible inactivation study. Brain. 2001;124:571–586. doi: 10.1093/brain/124.3.571. [DOI] [PubMed] [Google Scholar]
- Frey SH, Vinton D, Norlund R, Grafton ST. Cortical topography of human anterior intraparietal cortex active during visually guided grasping. Brain Res Cogn Brain Res. 2005;23(2–3):397–405. doi: 10.1016/j.cogbrainres.2004.11.010. [DOI] [PubMed] [Google Scholar]
- Fugate JE, Claassen DO, Cloft HJ, Kallmes DF, Kozak OS, Rabinstein AA. Posterior reversible encephalopathy syndrome: Associated clinical and radiologic findings. Mayo Clin Proc. 2010;85(5):427–432. doi: 10.4065/mcp.2009.0590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallese V, Murata A, Kaseda M, Nanako N, Sakata H. Deficit of hand preshaping after muscimol injection in monkey parietal cortex. Neuroreport. 1994;5:1525–1529. doi: 10.1097/00001756-199407000-00029. [DOI] [PubMed] [Google Scholar]
- Gallivan JP, McLean DA, Smith FW, Culham JC. Decoding effector-dependent and effector-independent movement intentions from human parieto-frontal brain activity. J Neurosci. 2011a;31:17149–17168. doi: 10.1523/JNEUROSCI.1058-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallivan JP, McLean DA, Valyear KF, Pettypiece CE, Culham JC. Decoding action intentions from preparatory brain activity in human parieto-frontal networks. J Neurosci. 2011b;31:9599–9610. doi: 10.1523/JNEUROSCI.0080-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallivan JP, McLean DA, Flanagan JR, Culham JC. Where one hand meets the other: limb-specific and action-dependent movement plans decoded from preparatory signals in single human frontoparietal brain areas. J Neurosci. 2013;33:1991–2008. doi: 10.1523/JNEUROSCI.0541-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia Guijo C, Rubio Esteban G, Pascual Martin-Gamero ML, Escamilla Crespo C. Balint’s syndrome secondary to cerebral toxoplasmosis in a female patient with AIDS. Rev Clin Esp. 1990 Nov;187(7):375–6. [PubMed] [Google Scholar]
- Garcin R, Rondot P, Recondo J. Ataxie optique localisee aux deux hemichamps visuels homonymes gauches. Rev Neurol. 1967;116:707–714. [PubMed] [Google Scholar]
- Gamberini M, Passarelli L, Fattori P, Zucchelli M, Bakola S, Luppino G, Galletti C. Cortical connections of the visuomotor parietooccipital area V6Ad of the macaque monkey. J Comp Neurol. 2009;513:622–642. doi: 10.1002/cne.21980. [DOI] [PubMed] [Google Scholar]
- Gaveau V, Pelisson D, Blangero A, Urquirzar C, Prablanc C, Vighetto A, Pisella A. Saccade control and eye-hand coordination in optic ataxia. Neuropsychologia. 2008;46:475–486. doi: 10.1016/j.neuropsychologia.2007.08.028. [DOI] [PubMed] [Google Scholar]
- Glover S. Separate visual representations in the planning and control of action. Behav Brain Sci. 2004;27:3–78. doi: 10.1017/s0140525x04000020. [DOI] [PubMed] [Google Scholar]
- Gnadt JW, Andersen RA. Memory related motor planning activity in posterior parietal cortex of macaque. Exp Brain Res. 1988;70:216–220. doi: 10.1007/BF00271862. [DOI] [PubMed] [Google Scholar]
- Goodale MA, Milner DA. Separate visual pathways for perception and action. Trends Neurosci. 1992;15(1):20–25. doi: 10.1016/0166-2236(92)90344-8. [DOI] [PubMed] [Google Scholar]
- Gradinaru V, Mogri M, Thompson KR, Henderson JM, Deisseroth K. Optical deconstruction of Parkinsonian neural circuitry. Science. 2009;324:354–359. doi: 10.1126/science.1167093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grefkes C, Ritzl A, Zilles K, Fink GR. Human medial intraparietal cortex subserves visuomotor coordinate transformation. Neuroimage. 2004;23(4):1494–1506. doi: 10.1016/j.neuroimage.2004.08.031. [DOI] [PubMed] [Google Scholar]
- Gurjinder PS, Dhand UK, Chopra JS. Balint’s syndrome following eclampsia. Clin Neurol Neurosurg. 1989;91(2):161–5. doi: 10.1016/s0303-8467(89)80039-3. [DOI] [PubMed] [Google Scholar]
- Han X, Qian X, Bernstein JG, Zhou H-h, Franzesi GT, Stern P, Bronson RT, Graybiel AM, Desimone R, Boyden ES. Millisecond-timescale optical control of neural dynamics in the nonhuman primate brain. Neuron. 2009;62:191–198. doi: 10.1016/j.neuron.2009.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauschild M, Mulliken GH, Fineman I, Loeb GE, Andersen RA. Cognitive signals for brain-machine interfaces in posterior parietal cortex include continuous 3D trajectory commands. Proc Natl Acad Sci USA. 2012;109(42):17075–17080. doi: 10.1073/pnas.1215092109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heed T, Beurze SM, Toni I, Roder B, Medendorp WP. Functional rather than effector-specific organization of human posterior parietal cortex. J Neurosci. 2011;31:3066–3076. doi: 10.1523/JNEUROSCI.4370-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Himmelbach M, Karnath HO, Perenin MT, Franz VH, Stockmeier K. A general deficit of the ‘automatic pilot’ with posterior parietal cortex lesions. Neuropsychologia. 2006;44:2749–2756. doi: 10.1016/j.neuropsychologia.2006.04.030. [DOI] [PubMed] [Google Scholar]
- Hinkley LBN, Krubitzer LA, Padberg J, Disbrow EA. Visual-manual exploration and posterior parietal cortex in humans. J Neurophysiol. 2009;102:3433–3446. doi: 10.1152/jn.90785.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hof PR, Bouras C, Constantinidis J, Morrison JH. Selective disconnection of specific visual association pathways in cases of Alzheimer’s disease presenting with Balint’s syndrome. J Neuropathol Exp Neurology. 1990;49:168–184. doi: 10.1097/00005072-199003000-00008. [DOI] [PubMed] [Google Scholar]
- Holmes G. Disturbances of visual orientation. Br J Ophthalmol. 1918 Sep;2(9):449–516. doi: 10.1136/bjo.2.9.449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes G, Horrax G. Disturbances of spatial orientation and visual attention, with loss of stereoscopic vision. Arch Neurol Psychiatry. 1919;1:385–407. [Google Scholar]
- Hwang EJ, Hauschild M, Wilke M, Andersen RA. Inactivation of the parietal reach region causes optic ataxia, impairing reaches but not saccades. Neuron. 2012;76:1021–1029. doi: 10.1016/j.neuron.2012.10.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inouchi M, Matsumoto R, Taki J, Kikuchi T, Mitsueda-Ono T, Mikuni N, Wheaton L, Hallett M, Fukuyama J, Shibasaki H, et al. Role of posterior parietal cortex in reaching movements in humans: clinical implication for “optic ataxia”. Clincal Neruophysiology. 2013;124:2230–2241. doi: 10.1016/j.clinph.2013.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson SR, Newport R, Mort D, Husain M. Where the eye looks, the hand follows, limb-dependent magnetic misreaching in optic ataxia. Curr Biol. 2005;15:42–46. doi: 10.1016/j.cub.2004.12.063. [DOI] [PubMed] [Google Scholar]
- Jackson SR, Newport R, Husain M, Fowlie JE, O-Donoghue M, Bajaj N. There may be more to reaching than meets the eye: Re-thinking optic ataxia. Neuropsychologia. 2009;24:1397–1408. doi: 10.1016/j.neuropsychologia.2009.01.035. [DOI] [PubMed] [Google Scholar]
- Jastorff J, Begliomini C, Fabbri-Destro M, Rizzolatti G, Orban GA. Coding observed motor acts: different organizational principles in the parietal and premotor cortex of humans. J Neurophysiol. 2010;104:128–140. doi: 10.1152/jn.00254.2010. [DOI] [PubMed] [Google Scholar]
- Jax SA, Buxbaum LJ, Lie E, Coslett HB. More than (where the target) meets the eyes: Disrupted visuomotor transformations in optic ataxia. Neuropsychologia. 2009;47:230–238. doi: 10.1016/j.neuropsychologia.2008.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jennings JH, Sparta DR, Stamatakis AM, Ung RL, Pleil KE, Kash TL, Stuber GD. Distinct extended amygdala circuits for divergent motivational states. Nature. 2013;496:224–228. doi: 10.1038/nature12041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeannerod M, Arbib MA, Rizzolatti G, Sakata H. Grasping objects: the cortical mechanisms of visuomotor transformation. TINS. 1995;18:314–320. [PubMed] [Google Scholar]
- Kagan I, Iyer A, Lindner A, Andersen RA. Space representation for eye movements is more contralateral in monkeys than in humans. PNAS. 2010;107:7933–7938. doi: 10.1073/pnas.1002825107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karnath HO, Perenin MT. Cortical control of visually guided reaching, evidence from patients with Optic Ataxia. Cereb Cortex. 2005;15:1561–1569. doi: 10.1093/cercor/bhi034. [DOI] [PubMed] [Google Scholar]
- Kas A, de Souza C, et al. Neural correlates of cognitive impairment in posterior cortical atrophy. Brain. 2011;134:1464–1478. doi: 10.1093/brain/awr055. [DOI] [PubMed] [Google Scholar]
- Khan AZ, Pisella L, Vighetto A, Cotton F, Luaute J, Boisson D, Salemme R, Crawford JD, Rosetti Y. Optic ataxia errors depend on remapped, not viewed, target location. Nature Neurosci. 2005;8:418–420. doi: 10.1038/nn1425. [DOI] [PubMed] [Google Scholar]
- Khan AZ, Pisella L, Blohm G. Causal evidence for posterior parietal cortex involvement in visual-to-motor transformations of reach targets. Cortex. 2013;49:2439–2448. doi: 10.1016/j.cortex.2012.12.004. [DOI] [PubMed] [Google Scholar]
- Klaes C, Westendorff S, Chakrabarti S, Gail A. Choosing goals, not rules: deciding among rule-based action plans. Neuron. 2011;70:536–548. doi: 10.1016/j.neuron.2011.02.053. [DOI] [PubMed] [Google Scholar]
- Kirshner HS, Lavin PJ. Posterior cortical atrophy: a brief review. Curr Neurol Neurosci Rep. 2006 Nov;6(6):477–80. doi: 10.1007/s11910-006-0049-0. [DOI] [PubMed] [Google Scholar]
- Konen CS, Kastner S. Representation of eye movements and stimulus motion in topographically organized areas of human posterior parietal cortex. J Neurosci. 2008;13;28(33):8361–75. doi: 10.1523/JNEUROSCI.1930-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konen CS, Mruczek RE, Montoya JL, Kastner S. Functional organization of human posterior parietal cortex: grasping- and reaching-related activations relative to topographically organized cortex. J Neurophysiol. 2013;109(12):2897–908. doi: 10.1152/jn.00657.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S, Abhayambika A, Sundaram AN, Sharpe JA. Posterior reversible encephalopathy syndrome presenting as Balint syndrome. J Neuroophthalmol. 2011 Sep;31(3):224–7. doi: 10.1097/WNO.0b013e31821b5f92. [DOI] [PubMed] [Google Scholar]
- Lammel S, Lim BK, Ran C, Huang KW, Betley MJ, Tye KM, Deisseroth K, Malenka RC. Input-specific control of reward and aversion in the ventral tegmental area. Nature. 2012;491:212–217. doi: 10.1038/nature11527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehmann SJ, Scherberger H. Reach and gaze representations in Macaque parietal and premotor grasp Areas. J Neurosci. 2013;33(16):7038–7049. doi: 10.1523/JNEUROSCI.5568-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levy I, Schluppeck D, Heeger DJ, Glimcher PW. Specificity of human cortical areas for reaches and saccades. J Neurosci. 2007;27:4687–4696. doi: 10.1523/JNEUROSCI.0459-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis JW, Van Essen DC. Mapping of architectonic subdivisions in the macaque monkey, with emphasis on parieto-occipital cortex. J Com Neurol. 2000;428:79–111. doi: 10.1002/1096-9861(20001204)428:1<79::aid-cne7>3.0.co;2-q. [DOI] [PubMed] [Google Scholar]
- Luppino G, Murata A, Govoni P, Matelli M. Largely segregated parietofrontal connections linking rostral intraparietal cortex (areas AIP and VIP) and the ventral premotor cortex (areas F5 and F4) Exp brain Res. 1999;128:181–187. doi: 10.1007/s002210050833. [DOI] [PubMed] [Google Scholar]
- Li CS, Mazzoni P, Andersen RA. The effect of reversible inactivation of macaque lateral intraparietal area on visual and memory saccades. J Neurophysiol. 1999;81:1827–1838. doi: 10.1152/jn.1999.81.4.1827. [DOI] [PubMed] [Google Scholar]
- Liu Y, Yttri EA, Snyder LH. Intention and attention: different functional roles for LIPd and LIPv. Nat Neruosci. 2010;13:495–502. doi: 10.1038/nn.2496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lynch JC, Mountcastle VB, Talbot WH, Yin TCT. Parietal lobe mechanisms for directed visual-attention. J Neurophysiol. 1977;40:362–389. doi: 10.1152/jn.1977.40.2.362. [DOI] [PubMed] [Google Scholar]
- Lynch JC, Graybiel AM, Lobeck LJ. The differential projection of two cytoarchitectonic subregions of the inferior parietal lobule of macaque upon the deep layers of the superior colliculus. J Comp Neurol. 1985;235:241–254. doi: 10.1002/cne.902350207. [DOI] [PubMed] [Google Scholar]
- McIntosh RD, Mulroue A, Blangero A, Pisella L, Rosetti Y. Correlated deficits of perception and action in optic ataxia. Neuropsychologia. 2011;49:131–137. doi: 10.1016/j.neuropsychologia.2010.11.017. [DOI] [PubMed] [Google Scholar]
- McMonagle P, Deering F, Berliner Y, Kertesz A. The cognitive profile of posterior cortical atrophy. Neurology. 2006;66(3):331–8. doi: 10.1212/01.wnl.0000196477.78548.db. [DOI] [PubMed] [Google Scholar]
- Medendorp WP, Goltz HC, Villis T, Crawford JD. Gaze-centered updating of visual space in human parietal cortex. Journal of Neuroscience. 2003;23(15):6209–6214. doi: 10.1523/JNEUROSCI.23-15-06209.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meek BP, Shelton P, Marotta JJ. Posterior cortical atrophy: visuomotor deficits in reaching and grasping. Front Hum Neurosci. 2013 Jun 21;7:294. doi: 10.3389/fnhum.2013.00294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mejia N, Park S, Ning M, Buonanno F. Pearls and Oysters: Reversible iatrogenic Balint syndrome. Neurology. 2008;70(24):e97–e98. doi: 10.1212/01.wnl.0000314730.64232.5a. [DOI] [PubMed] [Google Scholar]
- Mendez MF, Turner J, Gilmore G, Remler B, Tomask RL. Balint’s syndrome in Alzheimer’s disease, visuospatial functions. Int J Neurosci. 1990;54:339–346. doi: 10.3109/00207459008986653. [DOI] [PubMed] [Google Scholar]
- Merchut MP, Richie B. Transient visuospatial disorder from angiographic contrast. Arch Neurol. 2002 May;59(5):851–4. doi: 10.1001/archneur.59.5.851. [DOI] [PubMed] [Google Scholar]
- Montero J, Pena J, Genis D, Rubio F, Peres-Serra J, Barraquer-Bordas L. Balint’s syndrome. Report of four cases with watershed parieto-occipital lesions from vertebrobasilar ischemia or systemic hypotension. Acta Neurol Belg. 1982 Sep-Oct;82(5):270–80. [PubMed] [Google Scholar]
- Moreaud O. Balint Syndrome. Arch Neurol. 2003 Sep;60(9):1329–31. doi: 10.1001/archneur.60.9.1329. [DOI] [PubMed] [Google Scholar]
- Mountcastle VB, Lynch JC, Georgopoulos A, Sakata H, Acuna C. Posterior parietal association cortex of monkey-command functions for operations within extrapersonal space. J Neurophysiol. 1975;38:371–908. doi: 10.1152/jn.1975.38.4.871. [DOI] [PubMed] [Google Scholar]
- Mulliken G, Musallam S, Andersen RA. Forward estimation of movement state in posterior parietal cortex. Proc Natl Acad Sci USA. 2008;105(24):8170–8177. doi: 10.1073/pnas.0802602105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murata A, Gallese V, Luppino G, Kaseda M, Sakata H. Selectivity for the shape, size and orientation of objects for grasping in neurons of monkey parietal area AIP. J Neurophysiol. 2000;83:2580–2601. doi: 10.1152/jn.2000.83.5.2580. [DOI] [PubMed] [Google Scholar]
- Musallam S, Corneil BD, Greger B, Scherberger H, Andersen RA. Cognitive control signals for neural prosthetics. Science. 2004;305(5681):258–262. doi: 10.1126/science.1097938. [DOI] [PubMed] [Google Scholar]
- Passarelli L, Rosa MG, Gamberini M, Bakola S, Burman KJ, Fattori P, Galletti C. Cortical connections of area V6Av in the macaque: a visual-input node to the eye/hand coordination system. J Neurosci. 2011;31:1790–1801. doi: 10.1523/JNEUROSCI.4784-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perenin MT, Vighetto A. Optic ataxia, a specific disruption in visuomotor mechanisms. Brain. 1988;111:643–674. doi: 10.1093/brain/111.3.643. [DOI] [PubMed] [Google Scholar]
- Petit L, Haxby JV. Functional anatomy of pursuit eye movements in humans as revealed by fMRI. Journal of Neurophysiology. 1999;82(1):463–471. doi: 10.1152/jn.1999.82.1.463. [DOI] [PubMed] [Google Scholar]
- Pisella L, Grea H, Tilikete C, Vighetto A, Desmurget M, Rode G, Boisson D, Rosetti Y. An ‘automatic pilot’ for the hand in human posterior parietal cortex, toward reinterpreting optic ataxia. Nat Neurosci. 2000;3:729–736. doi: 10.1038/76694. [DOI] [PubMed] [Google Scholar]
- Pisella L, Sergio L, Blangero A, Torchin H, Vighetto A, Rossetti Y. Optic ataxia and the function of the dorsal steam: contributions to perception and action. Neuropsychologia. 2009;47:2033–3044. doi: 10.1016/j.neuropsychologia.2009.06.020. [DOI] [PubMed] [Google Scholar]
- Pitzalis S, Sereno MI, Committeri G, Fattori P, Galati G, Tonsoni A, Galletti C. The human homologue of macaque area V6A. Neuroimage. 2013;82:517–530. doi: 10.1016/j.neuroimage.2013.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prado J, Clavinger S, Otzenberger H, Schieber C, Kennedy H, Perenin MT. Two cortical systems for reaching in central and peripheral vision. Neuron. 2005;48:849–858. doi: 10.1016/j.neuron.2005.10.010. [DOI] [PubMed] [Google Scholar]
- Rafal R. Balint’s Syndrome. Handbook of Neuropsychology. (2) 2001;V [Google Scholar]
- Ristic AJ, Marjanovic I, Brajkovic L, Wolgamuth BR, Odalovic S, Lavrnic S, Vojvodic N, Jankovic S, Bascarevic V, Sokic D. Balint-like syndrome as an unusual representation of non-convulsive status epilepticus. Epileptic Disord. 2012 Mar;14(1):80–4. doi: 10.1684/epd.2012.0492. [DOI] [PubMed] [Google Scholar]
- Robinson DL, Goldberg ME, Stanton GB. Parietal association cortex in the primate: sensory mechanisms and behavioral modulations. J Neurophysiol. 1978;41:910–32. doi: 10.1152/jn.1978.41.4.910. [DOI] [PubMed] [Google Scholar]
- Rondot P, Recondo J, Dumas JLR. Visuomotor ataxia. Brain. 1977;100:355–376. doi: 10.1093/brain/100.2.355. [DOI] [PubMed] [Google Scholar]
- Salinas E, Thier P. Gain modulation: a major computational principle of the central nervous system. Neuron. 2000;27:15–21. doi: 10.1016/s0896-6273(00)00004-0. [DOI] [PubMed] [Google Scholar]
- Schindler I, Rice NJ, McIntosh R, Rossetti Y, Vighetto A, Milner DA. Automatic avoidance of obstacles is a dorsal stream function: evidence from optic ataxia. Nat Neurosci. 2004;7:7. doi: 10.1038/nn1273. [DOI] [PubMed] [Google Scholar]
- Sereno MI, Pitzalis S, Martinez A. Mapping of contralateral space in retinotopic coordinates by a parietal cortical area in humans. Science. 2001;294(5545):1350–1354. doi: 10.1126/science.1063695. [DOI] [PubMed] [Google Scholar]
- Shadlen MN, Newsome WT. Motion perception: seeing and deciding. Proc Natl Acad Sci USA. 1996;93:628–633. doi: 10.1073/pnas.93.2.628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snyder LH, Batista AP, Andersen RA. Coding of intention in the posterior parietal cortex. Nature. 1997;386:167–170. doi: 10.1038/386167a0. [DOI] [PubMed] [Google Scholar]
- Snyder LH, Batista A, Andersen RA. Change in motor plan, without a change in the spatial locus of attention, modulates activity in posterior parietal cortex. J Neurophysiol. 1998;79:2814–2819. doi: 10.1152/jn.1998.79.5.2814. [DOI] [PubMed] [Google Scholar]
- Snyder LH, Batista AP, Andersen RA. Saccade-related activity in the parietal reach region. J Neurophysiol. 2000;83:1099–1102. doi: 10.1152/jn.2000.83.2.1099. [DOI] [PubMed] [Google Scholar]
- Tanne-Gariepy J, Rouiller EM, Boussaoud D. Parietal inputs to dorsal versus ventral premotor areas in macaque monkey: evidence for largely segregated visuomotor pathways. Exp Brain Res. 2002;145:91–103. doi: 10.1007/s00221-002-1078-9. [DOI] [PubMed] [Google Scholar]
- Thier P, Andersen RA. Electrical microstimulation distinguishes distinct saccade-related areas in the posterior parietal cortex. J Neurophysiol. 1998;80(5):1713–1735. doi: 10.1152/jn.1998.80.4.1713. [DOI] [PubMed] [Google Scholar]
- Trillenberg P, Sprenger A, Petersen D, Kompf D, Heide W, Helmchen C. Functional dissociation of saccade and hand reaching control with bilateral lesions of the medial wall of the intraparietal sulcus, Implications for optic ataxia. Neuroimage. 2007;36:T69–T76. doi: 10.1016/j.neuroimage.2007.03.038. [DOI] [PubMed] [Google Scholar]
- Tsutsui K-I, Jiang M, Yara K, Sakata H, Taira M. Integration of perspective and disparity cues in surface-orientation-selective neurons of area CIP. J Neurophysiol. 2001;86:2856–2867. doi: 10.1152/jn.2001.86.6.2856. [DOI] [PubMed] [Google Scholar]
- Tye KM, Prakash R, Kim SY, Fenno LE, Grosenick L, Zarabi H, Thompson KR, Gradinaru V, Ramakrishnan C, Deisseroth K. Amygdala circuitry mediating reversible and bidirectional control of anxiety. Nature. 2011;471:358–362. doi: 10.1038/nature09820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Werf J, Jensen O, Fries P, Medendorp WP. Neuronal synchronization in human posterior parietal cortex during reach planning. J Neruosci. 2010;30:1402–1412. doi: 10.1523/JNEUROSCI.3448-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vesia M, Prime SL, Yan X, Sergio L, Crawford JD. Specificity of human parietal saccade and reach regions during transcranial magnetic stimulation. J Neurosci. 2010;30:13053–13065. doi: 10.1523/JNEUROSCI.1644-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vesia M, Crawford JD. Specialization of reach function in human posterior parietal cortex. Exp Brain Res. 2012;221:1–18. doi: 10.1007/s00221-012-3158-9. [DOI] [PubMed] [Google Scholar]
- Vesia M, Bolton DA, Mochizuki G, Staines WR. Human parietal and primary motor cortical interactions are selectively modulated during the transport and grip formation of goal-directed hand actions. Neuropsychologia. 2013;15:410–417. doi: 10.1016/j.neuropsychologia.2012.11.022. [DOI] [PubMed] [Google Scholar]
- Wardak C, Olivier E, Duhamel JR. Saccadic target selection deficits after lateral intraparietal area inactivation in monkeys. J Neurosci. 2002;22(22):9877–9884. doi: 10.1523/JNEUROSCI.22-22-09877.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilke M, Kagan I, Andersen RA. Functional imaging reveals rapid reorganization of cortical activity after parietal inactivation in monkeys. Proc Natl Acad Sci USA. 2012;109:8274–8279. doi: 10.1073/pnas.1204789109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilke M, Kagan I, Andersen RA. Effects of pulvinar inactivation on spatial decision-making between equal and asymmetric reward options. J Cogn Neurosci. 2013;25(8):1270–1283. doi: 10.1162/jocn_a_00399. [DOI] [PubMed] [Google Scholar]
- Xia M, Wang J, He Y. BrainNet Viewer:a network visualization tool for human brain connectomics. PLoS One. 2013 Jul 4;8(7):e68910. doi: 10.1371/journal.pone.0068910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yttri EA, Liu Y, Snyder LH. Lesions of cortical area LIP affect reach onset only when the reach is accompanied by a saccade, revealing an active eye-hand coordination circuit. Proc Natl Acad Sci USA. 2013;110:2371–2376. doi: 10.1073/pnas.1220508110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yttri EA, Wang C, Liu Y, Snyder LH. The parietal reach region is limb specific and not involved in eye-hand coordination. J Neurophysiol. 2013 doi: 10.1152/jn.00058.2013. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zipser D, Andersen RA. A back propagation programmed network that simulates response properties of a subset of posterior parietal neurons. Nature. 1988;331:679–684. doi: 10.1038/331679a0. [DOI] [PubMed] [Google Scholar]