

Abstract
Recent studies have shown significant associations of aberrant DNA methylation in spermatozoa with idiopathic male infertility, increased frequency of spontaneous abortions and imprinting disorders. Thus, the analysis of DNA methylation of specific genes in spermatozoa has the potential to become a new valuable diagnostic marker in clinical andrology. This perspective article discusses the current state and value of DNA methylation analysis in the diagnostic setup of infertile men and outlines challenges and perspectives. It highlights the potential of DNA methylation in andrological diagnostics and its putative benefit in the examination of hitherto idiopathic infertile patients is described.
Keywords: DNA methylation, male infertility, spermatogenesis
INTRODUCTION
Worldwide 10%–15% of couples are affected by infertility which can be attributed to various factors. Approximately 50% of reported cases are accounted for by male factor infertility, resulting in a prevalence of about 7% of all men.1,2,3 These numbers highlight the need for reliable diagnostic tools and further investigations of potential treatments. The classical andrological diagnostic procedure starts with the clinical and biochemical/endocrinological examination of the patient and is followed by semen analyses. These procedures have remained relatively unchanged for decades and consist of the inspection of macroscopic and microscopic appearance of the semen and the determination of sperm number, motility, vitality and morphology. Furthermore leukocyte peroxidase activity is measured and a mixed antiglobulin reaction test is recommended.4 As in 4% of infertile patients and 20%–30% of azoospermic patients established genetic causes might underlie their infertility, patients are also routinely screened for chromosomal aberrations, microdeletions of the azoospermia factor-loci and mutations of the CFTR gene.3,5 In addition, further genetic causes for infertility are suspected like copy number variants (CNVs).6,7,8 In this context, Tüttelmann and colleagues investigated the role of CNVs in male infertility by analyzing normozoospermic controls and patients with idiopathic severe oligozoospermia or with Sertoli-cell-only syndrome. Although the mean number of CNVs were comparable between all groups, they found recurring CNVs which were present only in patients with severe oligozoospermia (10 CNVs), only in Sertoli-cell-only syndrome (3 CNVs) or in both groups with spermatogenic failure (1 CNV), but not in controls.6 In addition, the analysis of X-linked CNVs in men with different sperm count demonstrated 73 CNVs being related to spermatogenesis. Interestingly, the patients had a higher excessive rate of deletions/person and a higher mean sequence loss/person resulting in a higher global burden of deletions in patients compared to controls.7 Furthermore, Lopes et al.8 analyzed recently CNVs in the genome of idiopathic infertile men and normozoospermic controls and found that a man's risk of idiopathic spermatogenic failure is changed by rare autosomal deletions (by 10%), rare X-linked CNVs (by 29%) and rare Y-linked duplications (by 88%). In summary, all described CNVs mark candidate genes which possibly cause spermatogenic failure or enhance the risk for it and thus represent novel targets for future investigations.
Apart from CNVs several single nucleotide polymorphisms and gene polymorphisms have also been described to be associated with poor male reproductive parameters. Such variants were detected in numerous genes including PRM1 (protamine 1),9 DAZL (deleted in azoospermia-like),10 MTHFR (methylenetetrahydrofolate reductase)11,12 and the GST (Glutathione S-transferases) genes.13
However, 30%–40% of infertile patients are still characterized as idiopathic as the underlying (molecular) reason for their infertility is not known.2,3 Due to the high percentage of idiopathic male infertility new insights and approaches concerning the molecular and genetic nature of impaired spermatogenesis are urgently needed to improve diagnosis and treatment. Currently, the development of novel technologies such as whole-genome sequencing and single-cell sequencing14,15,16 lead to exciting new findings in this research field and inspires the search for novel biomarkers (e.g. genes, proteins and metabolites) which could be used in the diagnosis of male infertility.17,18
Among them recent descriptions of a strong association between aberrant DNA methylation in spermatozoa and idiopathic male infertility19,20,21,22,23 indicate that epigenetics might have a strong impact on the quantitative and qualitative aspects of spermatogenesis. This assumption is further strengthened by studies outlining a possible predictive power of spermatozoal DNA methylation in pregnancy outcome.24,25,26,27 Thus, the analysis of DNA methylation of specific genes in spermatozoa could serve as a new valuable and noninvasive diagnostic marker in clinical andrology. We therefore believe that it is timely to consider the value of epigenetics in the diagnostic setup of infertile men and to critically evaluate the current challenges and perspectives of the approach.
DNA METHYLATION AND SPERMATOGENESIS
Epigenetics describes mitotically and/or meiotically heritable changes in gene function beyond the DNA sequence itself such as DNA methylation, histone modification, miRNA and non-coding RNAs.28,29 Spermatozoa possess specific DNA methylation patterns which are obtained during early stages of spermatogenesis.30,31 In this context, some genes become differentially ‘imprinted’ by DNA methylation being important for fertilization and subsequent embryogenesis.32,33 Several recent studies have shown a strong association of aberrant DNA methylation (epimutations) of some imprinted genes in spermatozoa with idiopathic infertility.19,20,21,22,23,34,35,36 Interestingly, men with moderate or severe oligospermia often displayed abnormalities in both maternal and paternal imprints.19 Furthermore, Marques et al.20 described that nearly 50% of patients of an oligozoospermic cohort had defective DNA methylation of the paternally imprinted gene H19 and/or the maternally imprinted gene MEST. Our group demonstrated that low sperm counts are associated with aberrant DNA methylation of H19 and more so of MEST. Additionally, aberrant DNA methylation of MEST was found to be associated with decreased progressive sperm motility and poor sperm morphology.22 Two other studies showed that men with severe or moderate oligozoospermia display more aberrant imprints than normozoospermic men.23,34 Furthermore, Camprubí and colleagues demonstrated that infertile patients had more aberrant imprints than fertile men.36 Studies by Hammoud et al.21 and Minor et al.35 supported these findings by demonstrating that DNA methylation patterns in sperm of infertile patients were considerably altered at H19 and various additional imprinted loci (LIT1, MEST, SNRPN, PLAGL1 and PEG3). As several studies described associations of male infertility with spermatozoal DNA methylation of H19 and/or MEST, we performed meta-analyses of these studies (if suitable data for a meta-analysis were available) demonstrating that it is more likely for infertile patients to have aberrant MEST (odds ratio: 3.4, 95% confidence interval: 1.98–5.84, P < 0.0001) or H19 (odds ratio: 14.62, 95% confidence interval: 7.34–29.12, P < 0.0001) DNA methylation in spermatozoa compared to fertile men (Figure 1).
Figure 1.

Meta-analyses of studies which analyzed normal and abnormal DNA methylation of H19 and MEST in spermatozoa of infertile men in comparison to these values of fertile men. For each study the odds ratio with 95% confidence interval is shown. Odds ratio >1 means that it is more likely for infertile men to have aberrant imprints than for fertile men, odds ratio <1 signifies that it is less likely for infertile men to have aberrant imprints compared to fertile men.
Apart from imprinted genes, other studies described associations of aberrant DNA methylation of non-imprinted genes and CpG loci with oligozoospermia, abnormal sperm morphology and reduced sperm motility.37,38,39,40,41
Epigenetic aberrations of spermatozoa have been described to be possibly inheritable to the offspring42 and aberrant DNA methylation patterns are known to be responsible for imprinting disorders (e.g. Beckwith-Wiedeman syndrome and Angelman syndrome). Interestingly, these aberrant imprints and imprinting disorders seem to occur more frequently after assisted reproductive techniques (ARTs) than after natural conception-although the prevalence of these disorders is still extremely low.43,44,45,46,47,48,49 In general, the epigenome of the spermatozoa seems to have an impact on embryo development50,51 and epimutations of the spermatozoa could be the cause for miscarriages as associations of spontaneous abortions and aberrant spermatozoal DNA methylation have been described.26,27
These findings have led to the concept that spermatozoal DNA methylation has some potential predictive power for pregnancy outcome.24,25,26,27 In this context, a study by El Hajj et al.25 demonstrated significantly higher (P < 0.001) ALU DNA methylation in sperm samples which led to pregnancy and live birth; whereas, significantly lower (P = 0.027) ALU DNA methylation levels were found in spontaneous abortions. However, Benchaib et al.24 demonstrated by immunostaining of 5-methylcytosine that global DNA methylation of human spermatozoa was associated to pregnancy rate but not to fertilization rate or rate of good quality embryos. Furthermore, DNA methylation aberrations of H19 and the MTHFR promoter in paternal spermatozoa were described in cases of spontaneous abortions, suggesting an impact of spermatozoal DNA methylation on pregnancy outcome.26,27 In Supplementary Table 1 (6.9MB, tif) the studies which analyzed potential associations of spermatozoal DNA methylation with poor semen parameters or pregnancy outcome are summarized.
List of studies which analyzed potential associations of aberrant spermatozoal DNA methylation with male infertitlity, pregnancy rate and outcome
In addition, retained histones could play an essential role in the association of aberrant DNA methylation with male infertility. During spermatogenesis histones are replaced by protamines; however, this process is not completed and up to 15% of the histones remain in spermatozoal DNA.52,53 These retained histones mark genes and loci that play a role in development (e.g. imprinted genes) and can be epigenetically modified.54,55 Failures in histone retention and modification or aberrations in protamine distribution might not only have an impact on male fertility as already described in several studies,56,57,58,59 but also could affect spermatozoal DNA methylation patterns. Indeed, aberrant spermatozoal DNA methylation has been already shown in men with abnormal protamine replacement.21,40,60
Recently, DNA hydroxymethylation was described as an additional type of DNA methylation, but it is still obscure if 5-hydroxymethylcytosine is only an intermediate in the removal of 5-methylcytosine or a separate epigenetic mark.61 Until now, most of the conventional methods for DNA methylation analysis can distinguish only between methylated and unmethylated cytosine, but not between different kinds of methylation. Thus, methods have been and will be developed to investigate 5-hydroxymethylation.62,63 With these new techniques, future studies will be able to elucidate the role of hydroxymethylation in andrology as this DNA modification could also play an essential role in spermatogenesis and embryo development.
CHALLENGES FOR THE IMPLEMENTATION OF DNA METHYLATION ANALYSIS IN ANDROLOGICAL DIAGNOSTICS
Although as of today numerous studies support the potential utility of DNA methylation in andrological workup, none of these studies have evaluated whether epigenetic analysis is suitable or for that matter feasible in a routine andrological lab.
Essential requirements for the application of DNA methylation analysis in the daily clinical routine are still missing. For this purpose and as one of the first steps, time- and cost-effective technologies which provide dependable and reliable results have to be established. Several different technologies (which are to greater or lesser extent time- and cost-effective) for DNA methylation analysis such whole-genome bisulfite sequencing, genome-wide DNA methylation arrays and locus-specific DNA methylation analysis such as pyrosequencing or methylation-specific PCR-based analyses are currently available and summarized in Table 1. Alternatively, already available commercial tests for DNA methylation analysis in routine diagnostics could be used. These tests are often based on bisulfite conversion of DNA and subsequent methylation specific real-time PCRs of the relevant gene(s).64
Table 1.
List of frequently used methods for DNA methylation analysis

Some technologies are limited in their sensitivity of DNA methylation detection (immunostaining) or in the amount of investigated CpGs (e.g. pyrosequencing or methylation-specific PCR-based analysis); however, these methods can be performed at affordable costs. In contrast, technologies such as whole-genome bisulfite sequencing or high resolution methylation arrays are able to measure multitudinous sequences across the genome, but are requiring sophisticated bioinformatic settings and are run at relatively high costs. If considering a clinical/andrological setting, a cost-effective, easy to perform and still informative technology seems preferable. Thus, a locus-specific DNA methylation analysis might provide sufficient information and is more rapid and less expensive than whole-genome analyses.
Another important aspect which has to be considered when analyzing DNA methylation in spermatozoa is the purification of semen samples. Independent of which methods are being applied, it is of crucial importance that the sample consists of spermatozoa only and is not contaminated by somatic cells or bacteria. One commonly used method, also recommended by the World Health Organization, is to perform swim-up purification before semen analysis which provides a relatively pure fraction of motile sperm.4
Further specific issues that need to be addressed are whether spermatozoal DNA methylation patterns represent a constant parameter over time and to which extent intra- and/or interindividual variations of gene-specific DNA methylation patterns exist. For the use as a diagnostic parameter, spermatozoal DNA methylation should ideally be temporally stable within individuals and should only display a small variability between healthy men.
Until now only a few studies have analyzed the temporal stability as well as the inter- and intraindividual variability of DNA methylation in spermatozoa. While one study described that DNA methylation in spermatozoa is stable over a short period of time (up to 1 year),65 others found locus-, cell- and age-dependent differences.66,67 Concerning inter- and intraindividual variability, Krausz et al.68 recently analyzed multitudinous cytosine positions across the human genome by using high resolution Infinium 450K methylation arrays and demonstrated that DNA methylation in spermatozoa is highly conserved between normozoospermic men and stable in different subpopulations of the same individual.
In addition, quantitative relevant reference values of DNA methylation in spermatozoa need to be established69,70 to classify patients’ spermatozoal DNA methylation into ‘normal’ and ‘abnormal’. Such a reference should be calculated by studying a large, healthy control group from which the normal range is defined.71 Recently, our group established a first reference range for normal spermatozoal MEST DNA methylation (0%–15%) based on the 95th percentile of MEST DNA methylation of 31 highly selected men with normal andrological parameters.72 In the future, multicenter studies in several countries on different continents need to be performed in order to establish reference values based on different ethnicities and laboratory techniques.
Nevertheless, prior to introduction into clinical diagnostics the clinical benefit of each novel biomarker has to be evaluated. Although the number of potential biomarkers is increasing due to new technologies, only few of them are appropriate for the use in clinics since associations with clinical parameters do not always result in clinical importance. In addition, some markers are not valid for all patients, but only for a specific subgroup which could present a limitation of a clinical marker.18
As diagnostic protocols and performance can be quite different in several laboratories and diagnoses can vary widely in both accuracy and precision, standardizations of protocols, diagnostic procedures, the interpretation of results and patients reports are necessary. Furthermore, internal and external quality control procedures should be set up and performed. One example for this standardization and quality control assessment is the diagnostic performance for Y-chromosomal microdeletions (azoospermia factor). For this the European Academy of Andrology and the European Molecular Genetics Quality Network have teamed up together and supported the publication of Laboratory guidelines73 (http://www.emqn.org/bpguidelines.php). In this context, societies such as Asian Society of Andrology or European Society of Human Reproduction and Embryology could also have a leading role in the establishment of such a quality assessment scheme for DNA methylation analysis in spermatozoa.
PERSPECTIVES
Several studies have already shown that aberrant DNA methylation in spermatozoa is associated with oligozoospermia, abnormal sperm morphology and decreased progressive motility (Figure 2); however, the underlying mechanisms and exact nature of associations are still unknown. For example it is unknown whether DNA methylation in spermatozoa is influenced by the origin of infertility which could be due to either central or testicular causes. By the same token it is currently unclear to which extent epimutations affect qualitative and quantitative aspects of spermatogenesis and vice versa.
Figure 2.
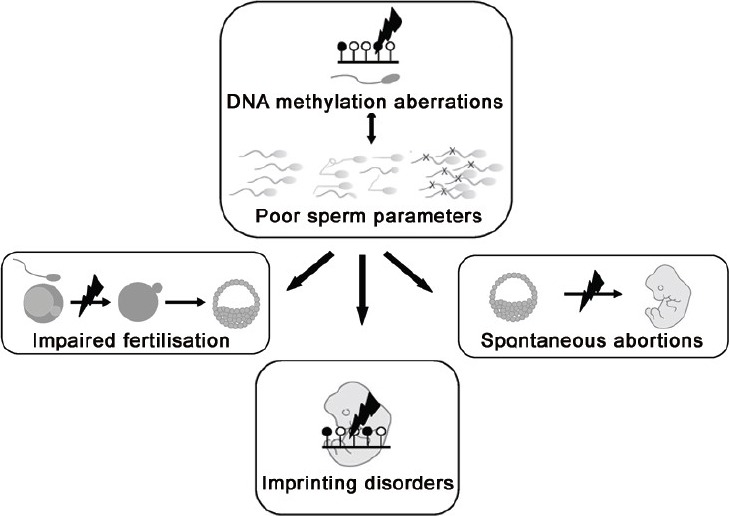
Aberrant imprints and abnormal semen parameters are associated with impaired fertilization, imprinting disorders and spontaneous abortions.
In addition, the knowledge of spermatozoal DNA methylation status could provide important information to subsequent fertilization, pregnancy success and possible outcome. Thus far ‘classical’ semen parameters only provide little information on the fertility capacity and cannot predict pregnancy rates or outcome, making DNA methylation a potentially invaluable tool in andrological examinations which in turn might yield a novel diagnostic parameter and prognostic factor for the outcome of any ART treatment.
The epigenetic status of spermatozoa could also affect the offspring's health by potential transmission of epimutations. Besides imprinting disorders (as already described above) epigenetic aberrations can have severe consequences on genomic stability and gene expression and thereby could cause serious diseases such as lupus, asthma, malignancies and several neurological disorders. For some of these diseases, commercial DNA methylation tests are already available now.64,74 However, it is still unknown if epimutations of spermatozoa can be transmitted to the offspring and affect its health as there are two main events in which epigenetic patterns are erased and renewed: during gametogenesis and in the early zygote after fertilization. These two epigenetic reprogramming events avoid in principle the transmission of parental DNA methylation aberrations to the offspring; but the reprogramming in the early zygote does not include imprinted genes making the transmission of spermatozoal epimutations to the first offspring (F1-generation) possible.75
It is also very clear that more studies are needed to determine to what extent DNA methylation analysis will be useful as diagnostic factor for male (in)fertility and as a prognostic factor for pregnancy. If information on the DNA methylation status could provide further hitherto unknown insights into the fertility capacity of the patient it would significantly improve the diagnostics for couples with longstanding history of male idiopathic infertility.
In the decade of whole genome sequencing, technologies in the field of epigenetics are rapidly developing. It is to be expected that methods for DNA methylation will probably be improved within the upcoming years and a cost-effective analysis for diagnostic procedures will be available soon. On the horizon one might even think about DNA methylation analysis of one single living sperm using techniques such as Raman microscopy,76,77 which could be used for ART. This would revolutionize the examination and treatment of infertile couples and increase the success of ART as sperm could be systematically selected for its potential to induce pregnancy.
Taken together, the current findings point to the potential of the approach; however, we encourage further investigations of the role of spermatozoal DNA methylation and subsequently fertilization, embryo development and pregnancy outcome to establish a novel prognostic parameter in the andrological workup of infertile men. Novel methods which enable the time- and cost-effective sequencing of the spermatozoal epigenome could make the epigenetic analysis of spermatozoa a standard measurement in andrology. If andrologists, andrological societies and the genetic quality assessment groups would combine their forces and activities we are convinced that this would lead to a new level in the diagnostic setup of male infertility.
AUTHOR CONTRIBUTIONS
RK helped in designing the concept of this article, performed literature research and wrote the manuscript. JG planned this article, designed the concept of this article, helped in literature research and writing the manuscript and supervised this project. All authors read and approved the final manuscript.
COMPETING INTERESTS
All authors declare no competing interests.
ACKNOWLEDGMENTS
Our work was supported by Graduate Program Cell Dynamics and Disease (CEDAD) and the International Max Planck Research School-Molecular Biomedicine (IMPRS-MBM) and by German Research Foundation (Research Unit ‘Germ cell potential’ (FOR 1041)).
Supplementary information is linked to the online version of the paper on the Asian Journal of Andrology website.
REFERENCES
- 1.Anderson JE, Farr SL, Jamieson DJ, Warner L, Macaluso M. Infertility services reported by men in the United States: national survey data. Fertil Steril. 2009;91:2466–70. doi: 10.1016/j.fertnstert.2008.03.022. [DOI] [PubMed] [Google Scholar]
- 2.Tüttelmann F, Werny F, Cooper TG, Kliesch S, Simoni M, et al. Clinical experience with azoospermia: aetiology and chances for spermatozoa detection upon biopsy. Int J Androl. 2011;34:291–8. doi: 10.1111/j.1365-2605.2010.01087.x. [DOI] [PubMed] [Google Scholar]
- 3.Hamada AJ, Esteves SC, Agarwal A. A comprehensive review of genetics and genetic testing in azoospermia. Clinics (Sao Paulo) 2013;68:39–60. doi: 10.6061/clinics/2013(Sup01)06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.World Health Organization. 5th ed. Geneva: world Health Organization; 2010. WHO Laboratory Manual for the Examination and Processing of Human Semen. [Google Scholar]
- 5.McLachlan RI, O’Bryan MK. Clinical Review#: State of the art for genetic testing of infertile men. J Clin Endocrinol Metab. 2010;95:1013–24. doi: 10.1210/jc.2009-1925. [DOI] [PubMed] [Google Scholar]
- 6.Tüttelmann F, Simoni M, Kliesch S, Ledig S, Dworniczak B, et al. Copy number variants in patients with severe oligozoospermia and sertoli-cell-only syndrome. PLoS ONE. 2011;6:e19426. doi: 10.1371/journal.pone.0019426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Krausz C, Giachini C, Lo Giacco D, Daguin F, Chianese C, et al. High resolution X chromosome-specific array-CGH detects new CNVs in infertile males. PLoS ONE. 2012;7:e44887. doi: 10.1371/journal.pone.0044887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lopes AM, Aston KI, Thompson E, Carvalho F, Goncalves J, et al. Human spermatogenic failure purges deleterious mutation load from the autosomes and both sex chromosomes, including the gene DMRT1. PLoS Genet. 2013;9:e1003349. doi: 10.1371/journal.pgen.1003349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.He XJ, Ruan J, Du WD, Chen G, Zhou Y, et al. PRM1 variant rs35576928 (Arg>Ser) is associated with defective spermatogenesis in the Chinese Han population. Reprod Biomed Online. 2012;25:627–34. doi: 10.1016/j.rbmo.2012.09.005. [DOI] [PubMed] [Google Scholar]
- 10.Teng YN, Chang YP, Tseng JT, Kuo PH, Lee IW, et al. A single-nucleotide polymorphism of the DAZL gene promoter confers susceptibility to spermatogenic failure in the Taiwanese Han. Hum Reprod. 2012;27:2857–65. doi: 10.1093/humrep/des227. [DOI] [PubMed] [Google Scholar]
- 11.Singh K, Singh SK, Sah R, Singh I, Raman R. Mutation C677T in the methylenetetrahydrofolate reductase gene is associated with male infertility in an Indian population. Int J Androl. 2005;28:115–9. doi: 10.1111/j.1365-2605.2004.00513.x. [DOI] [PubMed] [Google Scholar]
- 12.Gupta N, Gupta S, Dama M, David A, Khanna G, et al. Strong association of 677 C>T substitution in the MTHFR gene with male infertility--a study on an Indian population and a meta-analysis. PLoS ONE. 2011;6:e22277. doi: 10.1371/journal.pone.0022277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wu W, Lu J, Tang Q, Zhang S, Yuan B, et al. GSTM1 and GSTT1 null polymorphisms and male infertility risk: an updated meta-analysis encompassing 6934 subjects. Sci Rep. 2013;3:2258. doi: 10.1038/srep02258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Laird PW. Principles and challenges of genomewide DNA methylation analysis. Nat Rev Genet. 2010;11:191–203. doi: 10.1038/nrg2732. [DOI] [PubMed] [Google Scholar]
- 15.Li N, Ye M, Li Y, Yan Z, Butcher LM, et al. Whole genome DNA methylation analysis based on high throughput sequencing technology. Methods. 2010;52:203–12. doi: 10.1016/j.ymeth.2010.04.009. [DOI] [PubMed] [Google Scholar]
- 16.El Hajj N, Trapphoff T, Linke M, May A, Hansmann T, et al. Limiting dilution bisulfite (pyro)sequencing reveals parent-specific methylation patterns in single early mouse embryos and bovine oocytes. Epigenetics. 2011;6:1176–88. doi: 10.4161/epi.6.10.17202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kovac JR, Pastuszak AW, Lamb DJ. The use of genomics, proteomics, and metabolomics in identifying biomarkers of male infertility. Fertil Steril. 2013;99:998–1007. doi: 10.1016/j.fertnstert.2013.01.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Palmer SS, Barnhart KT. Biomarkers in reproductive medicine: the promise, and can it be fulfilled? Fertil Steril. 2013;99:954–62. doi: 10.1016/j.fertnstert.2012.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kobayashi H, Sato A, Otsu E, Hiura H, Tomatsu C, et al. Aberrant DNA methylation of imprinted loci in sperm from oligospermic patients. Hum Mol Genet. 2007;16:2542–51. doi: 10.1093/hmg/ddm187. [DOI] [PubMed] [Google Scholar]
- 20.Marques CJ, Costa P, Vaz B, Carvalho F, Fernandes S, et al. Abnormal methylation of imprinted genes in human sperm is associated with oligozoospermia. Mol Hum Reprod. 2008;14:67–74. doi: 10.1093/molehr/gam093. [DOI] [PubMed] [Google Scholar]
- 21.Hammoud SS, Purwar J, Pflueger C, Cairns BR, Carrell DT. Alterations in sperm DNA methylation patterns at imprinted loci in two classes of infertility. Fertil Steril. 2010;94:1728–33. doi: 10.1016/j.fertnstert.2009.09.010. [DOI] [PubMed] [Google Scholar]
- 22.Poplinski A, Tüttelmann F, Kanber D, Horsthemke B, Gromoll J. Idiopathic male infertility is strongly associated with aberrant methylation of MEST and IGF2/H19 ICR1. Int J Androl. 2010;33:642–9. doi: 10.1111/j.1365-2605.2009.01000.x. [DOI] [PubMed] [Google Scholar]
- 23.Sato A, Hiura H, Okae H, Miyauchi N, Abe Y, et al. Assessing loss of imprint methylation in sperm from subfertile men using novel methylation polymerase chain reaction Luminex analysis. Fertil Steril. 2011;95:129–34. doi: 10.1016/j.fertnstert.2010.06.076. [DOI] [PubMed] [Google Scholar]
- 24.Benchaib M, Braun V, Ressnikof D, Lornage J, Durand P, et al. Influence of global sperm DNA methylation on IVF results. Hum Reprod. 2005;20:768–73. doi: 10.1093/humrep/deh684. [DOI] [PubMed] [Google Scholar]
- 25.El Hajj N, Zechner U, Schneider E, Tresch A, Gromoll J, et al. Methylation status of imprinted genes and repetitive elements in sperm DNA from infertile males. Sex Dev. 2011;5:60–9. doi: 10.1159/000323806. [DOI] [PubMed] [Google Scholar]
- 26.Rotondo JC, Bosi S, Bazzan E, Di Domenico M, De Mattei M, et al. Methylenetetrahydrofolate reductase gene promoter hypermethylation in semen samples of infertile couples correlates with recurrent spontaneous abortion. Hum Reprod. 2012;27:3632–8. doi: 10.1093/humrep/des319. [DOI] [PubMed] [Google Scholar]
- 27.Ankolkar M, Patil A, Warke H, Salvi V, Kedia Mokashi N, et al. Methylation analysis of idiopathic recurrent spontaneous miscarriage cases reveals aberrant imprinting at H19 ICR in normozoospermic individuals. Fertil Steril. 2012;98:11861–92. doi: 10.1016/j.fertnstert.2012.07.1143. [DOI] [PubMed] [Google Scholar]
- 28.Qiu J. Epigenetics: unfinished symphony. Nature. 2006;441:143–5. doi: 10.1038/441143a. [DOI] [PubMed] [Google Scholar]
- 29.Carrell DT. Epigenetics of the male gamete. Fertil Steril. 2012;97:267–74. doi: 10.1016/j.fertnstert.2011.12.036. [DOI] [PubMed] [Google Scholar]
- 30.Hajkova P, Erhardt S, Lane N, Haaf T, El-Maarri O, et al. Epigenetic reprogramming in mouse primordial germ cells. Mech Dev. 2002;117:15–23. doi: 10.1016/s0925-4773(02)00181-8. [DOI] [PubMed] [Google Scholar]
- 31.Santos F, Dean W. Epigenetic reprogramming during early development in mammals. Reproduction. 2004;127:643–51. doi: 10.1530/rep.1.00221. [DOI] [PubMed] [Google Scholar]
- 32.Trasler JM. Epigenetics in spermatogenesis. Mol Cell Endocrinol. 2009;306:33–6. doi: 10.1016/j.mce.2008.12.018. [DOI] [PubMed] [Google Scholar]
- 33.Bartolomei MS, Ferguson-Smith AC. Mammalian genomic imprinting. Cold Spring Harb Perspect Biol. 2011;3 doi: 10.1101/cshperspect.a002592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Boissonnas CC, Abdalaoui HE, Haelewyn V, Fauque P, Dupont JM, et al. Specific epigenetic alterations of IGF2-H19 locus in spermatozoa from infertile men. Eur J Hum Genet. 2010;18:73–80. doi: 10.1038/ejhg.2009.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Minor A, Chow V, Ma S. Aberrant DNA methylation at imprinted genes in testicular sperm retrieved from men with obstructive azoospermia and undergoing vasectomy reversal. Reproduction. 2011;141:749–57. doi: 10.1530/REP-11-0008. [DOI] [PubMed] [Google Scholar]
- 36.Camprubí C, Pladevall M, Grossmann M, Garrido N, Pons MC, et al. Semen samples showing an increased rate of spermatozoa with imprinting errors have a negligible effect in the outcome of assisted reproduction techniques. Epigenetics. 2012;7:1115–24. doi: 10.4161/epi.21743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Houshdaran S, Cortessis VK, Siegmund K, Yang A, Laird PW, et al. Widespread epigenetic abnormalities suggest a broad DNA methylation erasure defect in abnormal human sperm. PLoS ONE. 2007;2:e1289. doi: 10.1371/journal.pone.0001289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Navarro-Costa P, Nogueira P, Carvalho M, Leal F, Cordeiro I, et al. Incorrect DNA methylation of the DAZL promoter CpG island associates with defective human sperm. Hum Reprod. 2010;25:2647–54. doi: 10.1093/humrep/deq200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wu W, Shen O, Qin Y, Niu X, Lu C, et al. Idiopathic male infertility is strongly associated with aberrant promoter methylation of methylenetetrahydrofolate reductase (MTHFR) PLoS ONE. 2010;5:e13884. doi: 10.1371/journal.pone.0013884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nanassy L, Carrell DT. Abnormal methylation of the promoter of CREM is broadly associated with male factor infertility and poor sperm quality but is improved in sperm selected by density gradient centrifugation. Fertil Steril. 2011;95:2310–4. doi: 10.1016/j.fertnstert.2011.03.096. [DOI] [PubMed] [Google Scholar]
- 41.Pacheco SE, Houseman EA, Christensen BC, Marsit CJ, Kelsey KT, et al. Integrative DNA methylation and gene expression analyses identify DNA packaging and epigenetic regulatory genes associated with low motility sperm. PLoS ONE. 2011;6:e20280. doi: 10.1371/journal.pone.0020280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kobayashi H, Hiura H, John RM, Sato A, Otsu E, et al. DNA methylation errors at imprinted loci after assisted conception originate in the parental sperm. Eur J Hum Genet. 2009;17:1582–91. doi: 10.1038/ejhg.2009.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.DeBaun MR, Niemitz EL, Feinberg AP. Association of in vitro fertilization with beckwith-wiedemann syndrome and epigenetic alterations of LIT1 and H19. Am J Hum Genet. 2003;72:156–60. doi: 10.1086/346031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gicquel C, Gaston V, Mandelbaum J, Siffroi JP, Flahault A, et al. In vitro fertilization may increase the risk of beckwith-wiedemann syndrome related to the abnormal imprinting of the KCN1OT gene. Am J Hum Genet. 2003;72:1338–41. doi: 10.1086/374824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Maher ER, Brueton LA, Bowdin SC, Luharia A, Cooper W, et al. Beckwith-Wiedemann syndrome and assisted reproduction technology (ART) J Med Genet. 2003;40:62–4. doi: 10.1136/jmg.40.1.62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sutcliffe AG, Peters CJ, Bowdin S, Temple K, Reardon W, et al. Assisted reproductive therapies and imprinting disorders–a preliminary british survey. Hum Reprod. 2006;21:1009–11. doi: 10.1093/humrep/dei405. [DOI] [PubMed] [Google Scholar]
- 47.Amor DJ, Halliday J. A review of known imprinting syndromes and their association with assisted reproduction technologies. Hum Reprod. 2008;23:2826–34. doi: 10.1093/humrep/den310. [DOI] [PubMed] [Google Scholar]
- 48.Manipalviratn S, DeCherney A, Segars J. Imprinting disorders and assisted reproductive technology. Fertil Steril. 2009;91:305–15. doi: 10.1016/j.fertnstert.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.van Montfoort AP, Hanssen LL, de Sutter P, Viville S, Geraedts JP, et al. Assisted reproduction treatment and epigenetic inheritance. Hum Reprod Update. 2012;18:171–97. doi: 10.1093/humupd/dmr047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jenkins TG, Carrell DT. The paternal epigenome and embryogenesis: poising mechanisms for development. Asian J Androl. 2011;13:76–80. doi: 10.1038/aja.2010.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Schagdarsurengin U, Paradowska A, Steger K. Analysing the sperm epigenome: roles in early embryogenesis and assisted reproduction. Nat Rev Urol. 2012;9:609–19. doi: 10.1038/nrurol.2012.183. [DOI] [PubMed] [Google Scholar]
- 52.Tanphaichitr N, Sobhon P, Taluppeth N, Chalermisarachai P. Basic nuclear proteins in testicular cells and ejaculated spermatozoa in man. Exp Cell Res. 1978;117:347–56. doi: 10.1016/0014-4827(78)90148-9. [DOI] [PubMed] [Google Scholar]
- 53.Oliva R. Protamines and male infertility. Hum Reprod Update. 2006;12:417–35. doi: 10.1093/humupd/dml009. [DOI] [PubMed] [Google Scholar]
- 54.Hammoud SS, Nix DA, Zhang H, Purwar J, Carrell DT, et al. Distinctive chromatin in human sperm packages genes for embryo development. Nature. 2009;460:473–8. doi: 10.1038/nature08162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Brykczynska U, Hisano M, Erkek S, Ramos L, Oakeley EJ, et al. Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nat Struct Mol Biol. 2010;17:679–87. doi: 10.1038/nsmb.1821. [DOI] [PubMed] [Google Scholar]
- 56.Carrell DT, Liu L. Altered protamine 2 expression is uncommon in donors of known fertility, but common among men with poor fertilizing capacity, and may reflect other abnormalities of spermiogenesis. J Androl. 2001;22:604–10. [PubMed] [Google Scholar]
- 57.Aoki VW, Liu L, Jones KP, Hatasaka HH, Gibson M, et al. Sperm protamine 1/protamine 2 ratios are related to in vitro fertilization pregnancy rates and predictive of fertilization ability. Fertil Steril. 2006;86:1408–15. doi: 10.1016/j.fertnstert.2006.04.024. [DOI] [PubMed] [Google Scholar]
- 58.Hammoud SS, Nix DA, Hammoud AO, Gibson M, Cairns BR, et al. Genome-wide analysis identifies changes in histone retention and epigenetic modifications at developmental and imprinted gene loci in the sperm of infertile men. Human Reprod. 2011;26:2558–69. doi: 10.1093/humrep/der192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Simon L, Castillo J, Oliva R, Lewis SE. Relationships between human sperm protamines, DNA damage and assisted reproduction outcomes. Reprod Biomed Online. 2011;23:724–34. doi: 10.1016/j.rbmo.2011.08.010. [DOI] [PubMed] [Google Scholar]
- 60.Aston KI, Punj V, Liu L, Carrell DT. Genome-wide sperm deoxyribonucleic acid methylation is altered in some men with abnormal chromatin packaging or poor in vitro fertilization embryogenesis. Fertil Steril. 2012;97:285–92. doi: 10.1016/j.fertnstert.2011.11.008. [DOI] [PubMed] [Google Scholar]
- 61.Branco MR, Ficz G, Reik W. Uncovering the role of 5-hydroxymethylcytosine in the epigenome. Nat Rev Genet. 2011;13:7–13. doi: 10.1038/nrg3080. [DOI] [PubMed] [Google Scholar]
- 62.Huang Y, Pastor WA, Shen Y, Tahiliani M, Liu DR, et al. The behaviour of 5-hydroxymethylcytosine in bisulfite sequencing. PLoS ONE. 2010;5:e8888. doi: 10.1371/journal.pone.0008888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kriukiene E, Liutkeviciute Z, Klimasauskas S. 5-hydroxymethylcytosine–the elusive epigenetic mark in mammalian DNA. Chem Soc Rev. 2012;41:6916–30. doi: 10.1039/c2cs35104h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Heyn H, Esteller M. DNA methylation profiling in the clinic: applications and challenges. Nat Rev Genet. 2012;13:679–92. doi: 10.1038/nrg3270. [DOI] [PubMed] [Google Scholar]
- 65.Cortessis VK, Siegmund K, Houshdaran S, Laird PW, Sokol RZ. Repeated assessment by high-throughput assay demonstrates that sperm DNA methylation levels are highly reproducible. Fertil Steril. 2011;96:1325–30. doi: 10.1016/j.fertnstert.2011.09.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Flanagan JM, Popendikyte V, Pozdniakovaite N, Sobolev M, Assadzadeh A, et al. Intra- and interindividual epigenetic variation in human germ cells. Am J Hum Genet. 2006;79:67–84. doi: 10.1086/504729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Jenkins TG, Aston KI, Cairns BR, Carrell DT. Paternal aging and associated intraindividual alterations of global sperm 5-methylcytosine and 5-hydroxymethylcytosine levels. Fertil Steril. 2013;100:945–51. doi: 10.1016/j.fertnstert.2013.05.039. [DOI] [PubMed] [Google Scholar]
- 68.Krausz C, Sandoval J, Sayols S, Chianese C, Giachini C, et al. Novel insights into DNA methylation features in spermatozoa: stability and peculiarities. PLoS ONE. 2012;7:e44479. doi: 10.1371/journal.pone.0044479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Talens RP, Boomsma DI, Tobi EW, Kremer D, Jukema JW, et al. Variation, patterns, and temporal stability of DNA methylation: considerations for epigenetic epidemiology. FASEB J. 2010;24:3135–44. doi: 10.1096/fj.09-150490. [DOI] [PubMed] [Google Scholar]
- 70.Schneider E, Pliushch G, El Hajj N, Galetzka D, Puhl A, et al. Spatial, temporal and interindividual epigenetic variation of functionally important DNA methylation patterns. Nucleic Acids Res. 2010;38:3880–90. doi: 10.1093/nar/gkq126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Marshall WJ, Bangert SK. 2nd ed. Edinburgh: Churchill Livingstone; 2008. Clinical Biochemistry: metabolic and Clinical Aspects; pp. 17–20. [Google Scholar]
- 72.Kläver R, Tüttelmann F, Bleiziffer A, Haaf T, Kliesch S, et al. DNA methylation in spermatozoa as a prospective marker in andrology. Andrology. 2013;1:731–40. doi: 10.1111/j.2047-2927.2013.00118.x. [DOI] [PubMed] [Google Scholar]
- 73.Simoni M, Bakker E, Krausz C. EAA/EMQN best practice guidelines for molecular diagnosis of y-chromosomal microdeletions. State of the art 2004. Int J Androl. 2004;27:240–9. doi: 10.1111/j.1365-2605.2004.00495.x. [DOI] [PubMed] [Google Scholar]
- 74.Kelly TK, De Carvalho DD, Jones PA. Epigenetic modifications as therapeutic targets. Nat Biotechnol. 2010;28:1069–1078. doi: 10.1038/nbt.1678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Reik W, Dean W, Walter J. Epigenetic reprogramming in mammalian development. Science. 2001;293:1089–93. doi: 10.1126/science.1063443. [DOI] [PubMed] [Google Scholar]
- 76.Kelly JG, Najand GM, Martin FL. Characterisation of DNA methylation status using spectroscopy (mid-IR versus Raman) with multivariate analysis. J Biophotonics. 2011;4:345–54. doi: 10.1002/jbio.201000085. [DOI] [PubMed] [Google Scholar]
- 77.Mallidis C, Wistuba J, Bleisteiner B, Damm OS, Gross P, et al. In situ visualization of damaged DNA in human sperm by raman microspectroscopy. Hum Reprod. 2011;26:1641–9. doi: 10.1093/humrep/der122. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
List of studies which analyzed potential associations of aberrant spermatozoal DNA methylation with male infertitlity, pregnancy rate and outcome