

SUMMARY
Recent advances in our understanding of tissue regeneration and the development of efficient approaches to induce and differentiate pluripotent stem cells for cell replacement therapies promise exciting avenues for treating degenerative age-related diseases. However, clinical studies and insights from model organisms have identified major roadblocks that normal aging processes impose on tissue regeneration. These new insights suggest that specific targeting of environmental niche components, including growth factors, ECM and immune cells, and intrinsic stem cell properties that are affected by aging will be critical for development of new strategies to improve stem cell function and optimize tissue repair processes.
INTRODUCTION
A central goal of regenerative medicine is the ability to restore or rejuvenate tissues using endogenous stem cells or exogenous replacement cells derived from stem or progenitor cells. This approach relies on harnessing tissue repair processes that have evolved to heal damaged tissues and to maintain tissue homeostasis. Improving and restoring such processes is likely to be a promising strategy for tissue rejuvenation and to allay age-related diseases.
Stem cells (SCs) are characterized by their multi-potency and their capacity to self-renew, giving rise to both progeny that differentiates to repair tissues and to progeny that retains SC properties to ensure the long-term maintenance of the SC pool (Fuchs and Chen, 2013). Somatic SC populations differ according to the regenerative requirement of their host tissue. In high-turnover tissues like the intestine or the hematopoietic system, the majority of the SC or progenitor population is active throughout life, while in tissues with low rates of homeostatic renewal like the muscle, SCs remain quiescent and are only activated by injury (reviewed in Barker et al., 2010; Cheung and Rando, 2013; Sanchez Alvarado and Yamanaka, 2014). In organs that lack SCs, induced pluripotent stem cell (iPSC)-derived replacement cells constitute a promising therapeutic avenue for functional restoration (reviewed in Fox et al., 2014). iPSCs regain the same developmental potential of embryonic SCs, and can thus, in principle, be differentiated into any tissue of interest (Tapia and Scholer, 2016).
Regardless of whether endogenous somatic SCs or iPSCs are used, the ability to regenerate somatic tissues is impacted by the systemic milieu and the tissue microenvironment, and it has become clear in recent years that modulating the systemic parameters and the microenvironment in aged or diseased conditions can significantly impact regenerative success (Forbes and Rosenthal, 2014; Lane et al., 2014; Wagers, 2012). Extracellular signals can influence all aspects of SC function: quiescence, proliferation, multipotency and differentiation. These signals are derived either from the systemic environment, reaching the SC via the vasculature, or the local environment – the niche. The niche can be considered any cell, tissue or extracellular matrix that resides in direct contact or close proximity to the SC population and that influences SC biology. Examples of niche components include other somatic and stromal cells, immune cells, extracellular matrix, innervating neuronal fibers, and the vasculature. Nevertheless, niche structure varies among the different somatic SC types, reflecting the different compositions of the host tissues and physiological demands of their somatic cells and resident SCs (Bjornsson et al., 2015; Scadden, 2014).
Signals derived from the blood that influence SC function include soluble molecules secreted by any tissue in the body, which can be hormones, growth factors and any other signaling molecules or immune-derived signals secreted by infiltrating immune cells. These signals may influence SC function directly or through modulation of the local environment. Indirect action includes changes to the makeup of the resident immune cell population, modulation of stromal or somatic cell function, and perturbation of the secretory phenotype of these cells (Jones and Wagers, 2008; Li et al., 2016a; Morgner et al., 2015; Rezza et al., 2016; Scadden, 2014; Wagers, 2012).
Both the niche and the systemic environment, but also SCs themselves (Fig. 1) change with age, and these changes limit the success of regenerative processes. Strategies targeted at improving the quality of these interactions in coordination with specific SC–directed interventions are likely to significantly enhance the success of regenerative therapies (Table 1). This may include identifying and using immune cell-derived factors that stimulate a specific aspect of the regenerative process or targeting the immune cells themselves with instructive signals to modulate regeneration. Such approaches are particularly relevant in conditions in which the immune environment is compromised, namely in degenerative diseases that are associated with inflammation, and in old patients, which often present with chronic low-level inflammation.
Fig. 1. Intrinsic regulators of SC function and their dysregulation with aging.
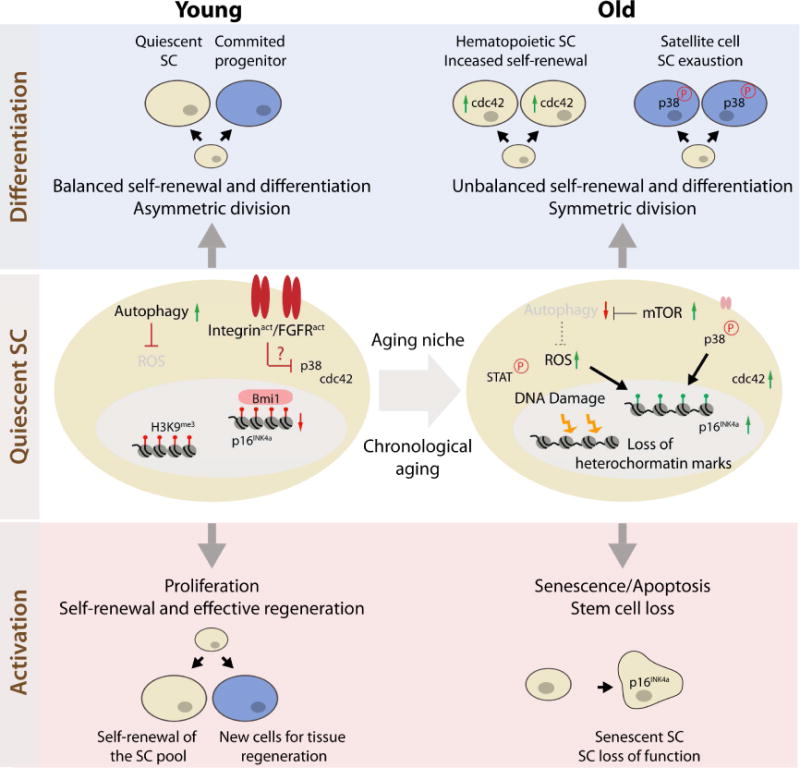
The figure summarizes molecular interactions identified independently in multiple systems, and these interactions are depicted in a generic cell type when not specified. Quiescent SC (middle panel): p16 is upregulated in old satellite cells and HSCs. In satellite cells the epigenetic de-repression of the INK4A locus (loss of Bmi1) is a consequence of accumulated ROS due to mTOR-dependent inhibition of the autophagic flux. An increase in p38 signaling further contributes to p16 induction in old satellite cells. In HSCs, p16 induction is a consequence of DNA damage and telomere shortening. Both satellite cells and HSCs display changes in epigenetic marks with aging. Moreover, Mesenchymal stem cells display a generalized reduction in the heterochromatin-associated H3K9me3 with age.
Differentiation (top panel): In old HSCs, elevated Cdc42 leads to cell polarity defects that favor self-renewal expansion at the expense of differentiation. Upon regenerative pressure, old satellite cells show unbalanced self-renewal and differentiation due to increased symmetric divisions. Symmetric segregation of p38 signaling in old satellite cells, due to p38 hyperactivation or altered β1-integrin activity and insensitivity to Fgf2, prevents asymmetric division and leads to stem cell exhaustion by differentiation.
Activation (bottom panel): Upon proliferative pressure, these changes result in satellite cell senescence as well as HSC senescence and apoptosis.
Table 1.
Therapeutic molecules with translational potential in regenerative medicine previously tested in model organisms. i.p., intraperitoneal; s.c., subcutaneous; i.m., intramuscular; i.vit., intravitreal.
| Therapeutic Molecule | Delivery | Cellular Target | Molecular Target/Effect | Reference |
|---|---|---|---|---|
| IL-22 (F-652) | Systemic (i.p./s.c. injection) | ISCs | Activation of Stat3 | Lindemans et al., 2015 |
| DOT1L inhibitor (epz-4777/epz-5676) | Systemic (i.p. injection) | unknown | Inhibition of DOT1L, NF-κB effector | Soria-Valles et al., 2015 |
| DOT1L inhibitor (epz-4777/epz-5676) | In vitro treatment | iPSCs | Inhibition of DOT1L, NF-κB effector Improved reprograming efficiency | Soria-Valles et al., 2015 |
| Fibronectin | Local (i.m.) | Satellite cells | Activation of integrin signaling - Protein tyrosine kinase 2 (PTK2, FAK) | Lukjanenko et al., 2016 |
| *β1-integrin-activating antibody TS2/16 | Local (i.m.) | Satellite cells | Activation of β1-integrin/FGFR | Rozo et al., 2016 |
| pyridone-6 | Local (Topical on skin) | HFSCs | Inhibition of JAK/STAT | Doles et al., 2012 |
| Tyr AG 490 | Local (i.m.) | Satellite cells | Inhibition of Jak2 | Price et al., 2014 |
| *5,15 diphenylporphrine | Local (i.m.) | Satellite cells | Inhibition of Stat3 | Price et al., 2014 |
| rIL-33 | Local (i.m.) or Systemic (i.p.) | Muscle Tregs | Activation of IL1RL1 (ST2) | Kuswanto et al., 2016 |
| GnRH | Systemic (s.c. injection) | unknown | unknown | Zhang et al., 2013 |
| hrMANF | Local (i.vit.) | Eye Macrophages | unknown | Neves et al., 2016 |
| ABT263 | Systemic (Gavage) | Senescent cells | Inhibition of BCL-2/BCL-xL | Chang et al., 2016 |
| SB203580 | Ex-vivo, prior to transplant | Satellite cells | Inhibition of p38 MAPK activity | Bernet et al., 2014 |
| SB202190 | Ex-vivo, prior to transplant | Satellite cells | Inhibition of p38 MAPK activity | Cosgrove et al., 2014 |
| Trolox | Systemic (i.p. injection) | Satellite cells | Anti-oxidant | Garcia-Prat et al., 2016 |
| CASIN | Ex-vivo, prior to transplant | HSCs | Inhibition of Cdc42 activity | Florian et al., 2012 |
| Oxytocin | Systemic (s.c. injection) | Satellite cells | Activation of MAPK/ERK signaling pathway | Elabd et al., 2014 |
| GDF11 | Systemic (i.p. injection) | Vasculature (in the brain) Unknown in other tissues |
Activation TGFβ signaling in endothelial cells of the brain ERK1/2 activation in renal epithelia |
Loffredo et al., 2013 Katsimpardi et al., 2014 Sinha et al., 2014 Zhang et al., 2016 |
Beyond inflammation, supplementation of growth factors, hormones or other signaling molecules which decline with aging are possible approaches to improve regeneration. Furthermore, successful regeneration can benefit from the elimination of detrimental local or systemic signals that accumulate with age, or of their cellular sources, such as senescent cells. In this review, we discuss potential regenerative medicine strategies to allay age-related changes to SCs and their environment and promise to enhance the repair capacity of old and diseased tissues. We specifically highlight new studies that have identified a growing number of promising rejuvenating interventions (Table 1) and propose a combined approach to target SC intrinsic limitations and niche deregulations simultaneously.
AGING ROADBLOCKS AND TARGETS FOR IMPROVEMENT
Aging is associated with a loss of regenerative capacity in many tissues. Age-related changes in the systemic environment, the niche, as well as the SCs themselves contribute to this loss. The contribution of the aging systemic environment has been elegantly demonstrated by studies using heterochronic parabiosis. By joining the circulatory system of young and old animals it was shown that the regenerative capacity of the skeletal muscle, liver, brain, bone and pancreas of young parabionts is worsened by exposure to an old systemic environment, whereas young blood can improve regeneration in old animals, reduce cardiac hypertrophy, and improve cognitive function (Baht et al., 2015; Conboy et al., 2005; Loffredo et al., 2013; Ruckh et al., 2012; Salpeter et al., 2013; Villeda et al., 2011; Villeda et al., 2014). A recent study suggests that similar effects can be obtained through a single heterochronic blood exchange (Rebo et al., 2016).
Intrinsic impairments to SC function, however, appear not to be reversible by systemic or niche rejuvenation, and intrinsic and extrinsic determinants of SC aging have been described as independent contributors to the loss of regenerative capacity (Liu and Rando, 2011; Oh et al., 2014). Interestingly, the observation that extrinsic impairments precede the development of intrinsic roadblocks suggests that dysregulated environmental cues may be drivers of intrinsic defects in old SCs (reviewed in Sacco and Puri, 2015). Identifying the nodal points where extrinsic and intrinsic age–related changes converge to drive the loss of SC function is thus critical. Such insight is beginning to emerge in recent years.
Intrinsic Limitations of Aged Stem Cells
In many tissues, active SCs persist into old age, but are often compromised in their ability to perform their functions (Schultz and Sinclair, 2016). Overcoming intrinsic alterations to SC function (Fig. 1, middle panel) is essential for interventions designed to recruit endogenous SCs for regenerative purposes. In the muscle, recent studies have shed light on the transition of normally quiescent SCs that can be activated during muscle regeneration towards a fully senescent state that impairs their activation in geriatric animals. SC quiescence is a tightly regulated process that ensures that the progenitor population retains long-term potential to be readily activated in response to injury (reviewed in Cheung and Rando, 2013). At geriatric ages (28 months and more in the mouse), quiescent muscle SCs are found in a pre-senescent state that is associated with the loss of epigenetic silencing of the INK4a locus and the consequent upregulation of the cell cycle inhibitor p16INK4a. These cells are unable to perform regenerative functions and will undergo full senescence upon regenerative pressure (Sousa-Victor et al., 2014).
Interestingly, the balance between quiescence and senescence depends on mechanisms that avert the accumulation of intracellular damage. Quiescent muscle SCs display a continuous high basal autophagy level, which is necessary for the clearance of damaged organelles and proteins. A failure to maintain autophagy flux in old satellite cells leads to increased ROS levels, INK4a de-repression and ultimately entry into senescence (Garcia-Prat et al., 2016). A similar quiescence-to-senescence transition, associated with increased levels of p16INK4a, is observed in aged hematopoietic stem cells (HSCs) as a consequence of telomere shortening and accumulation of DNA damage (Wang et al., 2014) (Fig. 1, middle and bottom panel).
Quiescence in HSCs is associated with an attenuation of DNA repair pathways (Beerman et al., 2014), while cycling old HSCs have increased levels of replication stress associated with cell cycle defects and chromosome gaps or breaks (Flach et al., 2014). The resulting accumulation of DNA damage and genome-wide mutations in aged SCs can promote changes in chromatin that lead to a broad deregulation in gene expression. Transcriptome analysis of aged hair follicle SCs revealed that the sustained DNA damage response in cycling cells during aging leads to a loss of the SC population from the skin through terminal epidermal differentiation (Matsumura et al., 2016).
Age-dependent alterations in the epigenome also affect chromatin structure and transcriptional regulation, consequently perturbing SC function (Rando and Chang, 2012). Mesenchymal SCs from aged individuals display a generalized reduction in the heterochromatin-associated H3K9me3 mark and a down-regulation of proteins involved in heterochromatin maintenance (Zhang et al., 2015). Similar alterations can be found in an embryonic SC model of Werner progeria syndrome, supporting the idea that heterochromatin disorganization may cause cellular dysfunction with age (Zhang et al., 2015). Epigenomic profiling of aged HSCs revealed that the activating H3K4me3 mark is increased in genes involved in maintaining HSC identity, while differentiation-promoting genes are increasingly repressed with age (Sun et al., 2014). These epigenetic changes are consistent with the reported increased numbers and reduced differentiation capacity of aged HSCs (Florian et al., 2012; Janzen et al., 2006; Rossi et al., 2005). Similar studies in quiescent satellite cells revealed that the young muscle SC pool is characterized by a permissive chromatin state, while aged satellite cells display a global increase in the repressive H3K27me3 mark (Liu et al., 2013). Although these findings suggest that changes in the epigenetic landscape are a general trait of aged SCs, further studies are required to determine the extent to which these alterations have a causal or correlative relation with SC aging. A recent study showed that a site-specific chromatin alteration in aged activated satellite cells leads to the induction of Hox9 and consequent activation of several pathways detrimental for proper satellite cell function. The inhibition of chromatin activation or deletion of Hoxa9 was sufficient to improve satellite cell function and muscle regeneration in old mice, suggesting a causal relation between aberrant epigenetic stress response and functionality of SCs of aged mice (Schworer et al., 2016). The same in depth analysis into causality will have to be applied to the effect of DNA damage in SC aging, considering studies in which defects in DNA repair lead to the accumulation of DNA damage with no effect on satellite cell function (Cousin et al., 2013).
If endogenous SCs are to be used to repair tissues in old patients, strategies to reverse some of the described age-related intrinsic alterations that impair regenerative capacity have to be devised (Table 1). Intervening at the level of gene regulation, through the silencing of p16INK4a expression, was shown to be effective in restoring quiescence and muscle SC function of geriatric mice (Sousa-Victor et al., 2014). Because p16INK4a upregulation is a common hallmark of SC aging in many tissues (Martin et al., 2014), this strategy may have a broader application in the regeneration of aged tissues. It should be noted, however, that reducing p16INK4a levels can also result in increased cancer incidence (Kim and Sharpless, 2006). Alternatively, it is also possible to target p16INK4a-expressing cells for elimination using senolytic agents (Chang et al., 2016). While this approach may have broader beneficial implications in the aging niche (see below), it can also lead to the elimination of senescent SCs independently, thus ensuring that a healthier SC pool is maintained through old age.
Another possible approach is to intervene at the level of dysregulated cell signaling pathways that cause alterations in gene regulation, including the increase in p16INK4a expression. Pharmacological inhibition of p38 MAPK signaling decreases the expression of cell cycle inhibitors and enhances the regenerative capacity of aged satellite cells in muscle transplantation experiments (Bernet et al., 2014; Cosgrove et al., 2014). Similarly, inhibition of JAK-STAT signaling improves satellite cell function in older mice (Price et al., 2014). Pharmacological interventions that restore basal autophagic flux in geriatric satellite cells are also effective in preventing irreversible cell cycle exit and improving muscle SC function (Garcia-Prat et al., 2016). Other examples of the use of compounds to target dysregulated pathways include the partial rejuvenation of aged HSCs through mTOR (Chen et al., 2009) and Cdc42 (Florian et al., 2012) inhibition. Increased mTOR activation represents a conserved mechanism for organismal aging (Johnson et al., 2013), and elevated activity of the small RhoGTPase Cdc42 has been reported in several aged mouse tissues (Wang et al., 2007). Genetic models of increased Cdc42 display phenotypic and functional changes in HSCs that are consistent with accelerated aging, including reduced reconstitution capacity and cell polarity defects. Treating aged HSCs with CASIN, a specific inhibitor of Cdc42, is sufficient to restore the reconstitution capacity and polarity of the old HSC population, opening the possibility that other aging phenotypes could also be reversed with a similar treatment (Florian et al., 2012; Florian et al., 2013). Defects in cell polarity are also observed in aged muscle SCs, in this case due to excessive p38 signaling. In fact, part of the mechanism through which p38 inhibition restores the function of old satellite cells is by restoring asymmetric SC division, a process that is necessary to sustain self-renewal (Bernet et al., 2014; Rozo et al., 2016) (Fig. 1, top panel).
The effects of aging in the generation of replacement cells
In many organs, endogenous repair mechanisms are inefficient, even at young age, due to the absence or limited functionality of a resident stem/progenitor cell pool. In these situations, tissue repair therapies have to rely on the transplantation of replacement cells generated in vitro. The success of such therapies depends on the efficient functional engraftment of replacement cells into the host tissue and on the quality of the replacement cells generated in vitro (Chen et al., 2012; Fox et al., 2014; Lamba et al., 2009; Lu et al., 2015; MacLaren et al., 2006).
Replacement cells can be generated through differentiation of induced pluripotent stem cells (iPSCs) derived from patient-specific or donor somatic cells, mesenchymal stem cells or embryonic stem cells (ESCs). Alternatively, they can also be generated through direct trans-differentiation of somatic cells. iPCSs are generated by reprogramming fully differentiated cells into a pluripotent state (Takahashi et al., 2007) which can then be differentiated into any cell type, while direct trans-differentiation involves the direct conversion of a somatic cell type into another, without intermediate induction of a pluripotent state (Huang et al., 2011; Ieda et al., 2010; Sekiya and Suzuki, 2011; Vierbuchen et al., 2010). Notably, whereas reprogramming into iPSCs erases the transcriptomic signatures of aging present in old fibroblasts, direct trans-differentiation of old fibroblasts into neurons preserves the aging-related profile (Mertens et al., 2015). Accordingly, neurons generated from iPSCs preserve youthful characteristics when compared to the ones generated by direct trans-differentiation (Mertens et al., 2015; Miller et al., 2013; Yang et al., 2015). Thus, while trans-differentiation may be an effective way to model diseases of aging in a dish in a patient-dependent manner, in the context of cell-replacement therapies, generation of replacement cells by sequential reprogramming and differentiation may be advantageous. The rejuvenation potential of cellular reprograming can also be used to directly target cells in vivo, as long as only partial reprograming is induced to avoid teratoma formation. This strategy was recently shown to have a general beneficial effect on organismal aging (Ocampo et al., 2016). Interestingly, aging itself favors the in vivo reprogramming process. The effect seems to be mediated by neighboring senescent cells that secrete IL-6 (Chiche et al., 2016; Mosteiro et al., 2016).
However, aging can also be a limiting factor for reprogramming (Li et al., 2009; Soria-Valles et al., 2015), limiting the success in generating replacement cells from old donors via iPSCs. Shorter telomere length, activation of the INK4a locus, p53-mediated DNA damage response, replication stress and NF-κB activation are characteristics of aging cells which impose a barrier to reprograming. Moreover, the reversal of these characteristics during the reprograming process is required for efficient iPSC generation (Banito et al., 2009; Li et al., 2009; Marion et al., 2009a; Marion et al., 2009b; Ruiz et al., 2015; Soria-Valles et al., 2015). Inhibition of several of these pathways has been efficiently applied to improve the reprograming efficiency from old donors (Table 1). Inhibition of the INK4 locus improves reprogramming efficiency of old mouse skin fibroblasts (Li et al., 2009), while inhibition of the chromatin modifying factor DOT1L, an effector of NF-κB activity, improves reprogramming efficiency of old (but not young) human fibroblasts (Soria-Valles et al., 2015).
It has further been noted that even in young donors, senescence and genomic instability arise naturally during reprogramming and impose a barrier to efficient iPSC generation (Banito et al., 2009; Ruiz et al., 2015). Consistently, silencing of senescence effectors (Banito et al., 2009) or reduction in replication stress by increasing CHK1 levels or nucleoside supplementation (Ruiz et al., 2015) facilitate reprogramming. Reducing replication stress further reduces genomic instability arising from reprogramming (Ruiz et al., 2015), and p53 inhibition enhances reprogramming efficiency under exogenously inflicted DNA damage. This perturbation, however, compromises genomic stability in the generated iPSCs, suggesting that p53 activation is a protective mechanism to ensure genomic integrity in the newly generated SCs (Marion et al., 2009a). Thus, while being a limiting factor to reprograming efficiency, p53 activation may be important to ensure the quality of the replacement cells generated by reprogramming from old donors with compromised genomic integrity. Mutations may also arise in the mtDNA of donor cells and be clonally expanded during iPSC generation. Because the frequency of mtDNA defects in iPSCs increases with the age of the donor cell, it is important to monitor mtDNA mutations in iPSCs generated from older patients (Kang et al., 2016).
If patient-derived iPSCs are to be used, aging limits the success of cell replacement-based regeneration in two ways: by reducing the ability to generate iPS cells and by preventing efficient implantation of iPSC-derived progenitor cells. The studies discussed above provide clues for strategies that can be applied to increase the efficiency of reprograming from old donors while ensuring genomic stability of the replacement cells. Studies discussed in the following, in turn, have provided interesting new insights into interventions that may improve engraftment and tissue repair through modulating niche and systemic conditions.
Changes in Biophysical Properties of Aged and Damaged Tissues
Niche cues that modulate SC behavior and orchestrate efficient tissue regeneration include a wide range of biophysical and biomechanical signals. The ability of a SC to efficiently respond to stimuli depends on interactions with the surrounding extracellular matrix (ECM) and is modulated by physical cues such as substrate elasticity or stiffness, surface topography and mechanical stresses (Conway and Schaffer, 2012; Nowell et al., 2016).
Age-related changes in the composition and structure of the ECM have a strong impact on the regenerative process (Phillip et al., 2015; Watt and Huck, 2013). Tissue aging is associated with loss of junctional integrity and gaps between epithelial cells, likely due to a decrease in the levels of cell adhesion molecules such as cadherins (Akintola et al., 2008), and associated with increased tissue stiffening of several organs (Kwak, 2013; Wood et al., 2014). The accumulation of senescent cells in old tissues and the consequent presence of senecence-associated secretory phenotype (SASP) factors (see below), that include ECM components and proteases, can also contribute to the altered biophysical properties of the aging niche (Coppe et al., 2010; Frantz et al., 2010).
A common feature of age-associated degenerative diseases and repeated tissue injuries is the excessive deposition of connective tissue associated with fibrosis, a process that significantly impacts the composition of the ECM (Brack et al., 2007; Burkauskiene et al., 2006; Calabresi et al., 2007; Gagliano et al., 2000). In the regenerating skeletal muscle, dysregulation of the pathways responsible for limiting matrix production results in the interstitial accumulation of collagen and impairment of regeneration (Lemos et al., 2015; Pessina et al., 2015). Attenuation of fibrosis also facilitates hepatic regeneration (Nakamura et al., 2000). In general, scar tissue creates an unfavorable environment for stem-cell based therapies, thus inhibiting fibrogenesis may be a potential point of intervention for regenerative therapies.
A more complete understanding of how cell-matrix interactions influence SC behavior enables advances in niche reconstruction that incorporate factors beyond biochemical signals (Dalby et al., 2007; Gilbert et al., 2010; McBeath et al., 2004; Quarta et al., 2016; Yim et al., 2007). Similarly, current methods for SC delivery in regenerative therapies strive to create local environments where survival and engraftment of SCs is enhanced by biochemical, biophysical, and biomechanical cues (Choi et al., 2014). A study that cultured satellite cells on hydrogel substrates with properties matching the rigidity of muscle tissue, demonstrates that biophysical solutions can be combined with biochemical treatments, such as p38 inhibitors, to rejuvenate muscle SCs for transplantation (Cosgrove et al., 2014). Another approach to enhance SC delivery and integration is to instruct cells to produce their own ECM while in culture, generating hybrid scaffolds that allow attachment and migration of SCs and can subsequently be transplanted to damaged tissues (Matsuura et al., 2014; Schenke-Layland et al., 2009). Indeed, in vivo studies show that activated satellite cells interact with interstitial fibrogenic cells, regulating collagen biosynthesis to ensure proper ECM deposition (Fry et al., 2017). Future advances in the use of biomaterials will likely help overcome some of the biophysical limitations to the application of stem-cell based therapies to aging tissues. Nevertheless, these types of interventions will require further development of safe routes for endogenous SC transplantation.
Dysregulated growth factor signaling
Age-related changes in local niche factors have been found to perturb regenerative capacity of somatic SC populations in several tissues. In the aging skeletal muscle, increases in local FGF and TGFβ signaling are coupled with decreased expression of the Notch ligand Dl in the myofiber and decreases in deposition of the extracellular matrix protein fibronectin. These changes synergistically contribute to the loss of regenerative capacity, mostly by affecting satellite cell function (Lukjanenko et al., 2016; Sousa-Victor et al., 2015). The increase in FGF signaling causes a loss of quiescence of old satellite cells, and leads to satellite cell loss (Chakkalakal et al., 2012). The remaining satellite cells become unresponsive to FGF2 in regenerative conditions and fail to expand or self-renew, thus impairing regenerative capacity of the muscle (Bernet et al., 2014). Moreover, age-related alterations in SC adhesion and ECM interactions impair SC signal responsiveness. In healthy tissue, fibronectin is induced during muscle regeneration and promotes integrin signaling in satellite cells. However, fibronectin is not efficiently induced in old mice, consequently satellite cells undergo defective integrin signaling, resulting in a decreased response to FGF signaling (Lukjanenko et al., 2016; Rozo et al., 2016). Treatment with activators of integrin signaling can restore FGF responsiveness and rescue proliferative and self-renewal defects of old satellite cells. Consequently, integrin signaling activators or fibronectin supplementation can rescue regenerative capacity of old muscles (Lukjanenko et al., 2016; Rozo et al., 2016). These niche-specific signaling dysregulations synergize with a systemic increase in Wnt signaling, which directly affects satellite cell function, promoting aberrant fibrogenic commitment that compromises regenerative capacity and promotes fibrosis (Brack et al., 2007). Thus, Wnt inhibitors are an effective way to restore satellite cell function in aging (Brack et al., 2007). Similarly, increased Wnt signaling derived from old serum promotes mesenchymal SC aging that can be attenuated by Wnt inhibitors (Zhang et al., 2011) or ROS inhibition (Zhang et al., 2013a).
A recent study identified the lateral ventricle choroid plexus (LVCP), a primary producer of cerebrospinal fluid (CSF), as an important niche component for neural stem cells (NSCs) affected by aging (Silva-Vargas et al., 2016). Studying age-related changes in the secretome of the LVCP, the authors identified molecules with reduced secretion in old age, such as BMP5 and IGF1, which can be supplemented to improve the function of old NSCs (Silva-Vargas et al., 2016). An age-dependent increase in TGF-β signaling is also observed in the neurogenic niche, with negative impact on neurogenesis (Pineda et al., 2013), while reduced secretion of wnt3 from astrocytes impairs neurogenesis in old mice (Okamoto et al., 2011). Furthermore, in the bone marrow, the vasculature is an important component of the niche supporting hematopoietic stem cells (HSCs). During aging, niche-forming vessels are reduced, however the vessels can be restored by activation of endothelial Notch signaling (Kusumbe et al., 2016). Together, these studies demonstrate that interventions targeted at restoring niche signaling capacity may promote regenerative success (Table 1).
Heterochronic parabiosis studies also led to the identification of rejuvenating factors found in the young systemic milieu that can improve regeneration. Plasma levels of the hormone oxytocin decrease with age, and its systemic administration improves regenerative capacity in the skeletal muscle of old mice through direct action on satellite cells (Elabd et al., 2014). GDF11, a member of the TGF-β family, was also recently described as a circulatory factor declining in aging mice, and systemic supplementation of recombinant GDF11 enhances regenerative capacity of skeletal muscle (Sinha et al., 2014), neurogenesis in the brain (Katsimpardi et al., 2014), and tubular regeneration after acute kidney injury (Zhang et al., 2016) as well as prevents hypertrophy in the heart of old mice (Loffredo et al., 2013). Interestingly, the effects in the brain are mediated by vascular remolding that promotes increased blood flow (Katsimpardi et al., 2014), yet the mechanism of action in other tissues is still unclear. The effects of GDF11 on skeletal muscle and its systemic changes with aging have recently been questioned (Egerman et al., 2015; Hinken et al., 2016; Poggioli et al., 2016; Schafer et al., 2016), calling for further studies to fully determine if this promising multi-target factor will be useful in therapies aimed at enhancing regenerative capacity.
Cellular senescence
Cellular senescence is a state of irreversible cell cycle arrest that can be induced by a series of stimuli that are prominent in aging organisms, including telomere dysfunction, genomic instability, oxidative stress or oncogene activation (Campisi and d’Adda di Fagagna, 2007). Senescent cells accumulate with aging in several tissues of primates and rodents and are a common feature at sites of age-related pathologies (Herbig et al., 2006; Jeyapalan et al., 2007; van Deursen, 2014; Wang et al., 2009). Although the state of senescence can impair the regenerative capacity of a tissue by limiting SC function in a cell-autonomous way (see above), many of the detrimental effects of cellular senescence are thought to be a consequence of the secretion of chemokines (Il-8, GROα and GROβ,and several members of the CCL and CXCL family of chemokines) pro-inflammatory cytokines (Il-6, Il-1α and Il-1β, GM-CSF and M-CSF), IGF binding proteins and extracellular matrix remodeling proteases (matrix metalloproteinases-MMPs, and serine proteases), commonly referred to as the senescence-associated secretory phenotype (SASP) (Coppe et al., 2010; van Deursen, 2014). Senescence activation is also associated with a wide range of essential biological activities that include embryonic development (Munoz-Espin et al., 2013; Storer et al., 2013), wound healing (Demaria et al., 2014), tissue repair (Krizhanovsky et al., 2008), and induction of epithelial-to-mesenchymal transitions (Laberge et al., 2012). Senescence is thus simultaneously an important tumor suppressor mechanism with key biological functions, but also a likely contributor to the general state of low-grade chronic inflammation in older organisms (Franceschi and Campisi, 2014). This dual nature of senescence and the SASP as key factors in tissue repair and as drivers of degenerative age-related pathologies significantly impacts regeneration and stem-cell based therapies (Neves et al., 2015). The SASP also influences the immune response at multiple levels. One of the key functions of secreted factors originating in senescent cells is to attract immune cells, which in turn are responsible for the elimination of senescent cells (Kang et al., 2011; Xue et al., 2007). In the context of acute liver injury in mice, the mechanism of immune clearance of senescent activated hepatic stellate cells involves the cytotoxic action of recruited natural killer cells and is essential to limit the fibrogenic response to tissue damage and to facilitate repair (Krizhanovsky et al., 2008). Although the transient presence of SASP factors following injury is essential for efficient tissue repair, the age-associated accumulation of senescent cells, perhaps due to the decline of adequate clearance by the aging immune system, can result in the formation of dysregulated immune microenvironments. In the mouse skin, stromal-derived SASP factors can recruit suppressive myeloid cells, with a consequent inhibition of anti-tumor T-cell responses, establishing a tumor-permissive and chronic inflammatory microenvironment (Ruhland et al., 2016). These characteristics of the aging niche will likely influence the efficacy of stem-cell based therapies, requiring strategies that take into account SASP-mediated immune alterations.
The SASP can further contribute to age-related tissue dysfunction through the paracrine induction of secondary senescence in healthy neighboring cells (Acosta et al., 2013; Nelson et al., 2012). Selective clearance of senescent cells and SASP suppression in HSCs and bone stromal cells, improves the function of the remaining healthy population of HSCs, allowing long-term and multi-lineage engraftment after bone marrow transplantation (Chang et al., 2016). The process of paracrine senescence can thus both affect the intrinsic regenerative capacity of functional SCs and the quality of the regenerating niche.
Efficient stem-cell based therapies will likely have to consider strategies for attenuating the deleterious effects of senescence on the regenerative process (Table 1). One possible approach is to eliminate the already present senescent cells. This concept has been successfully tested in progeroid and physiologically aged mouse models where the genetically induced clearance of senescent cells was shown to attenuate age-related deterioration of several organs without apparent side effects (Baker et al., 2016; Baker et al., 2011). Pharmacological interventions that can selectively kill senescent cells, are thus potential avenues to rejuvenate tissue microenvironments and recover regenerative potential in humans. One such senolytic agent, ABT263 (a specific inhibitor of the anti-apoptotic proteins BCL-2 and BCL-xL) has been shown to effectively deplete senescent cells when orally administrated to aged mice or animals subjected to a strong genotoxic stress. This reduction in senescent cells was sufficient to improve the function of HSCs and satellite cells (Chang et al., 2016). Although ABT263 has some toxic side effects that might limit its broad application as an anti-aging drug, the discovery of other senolytic compounds will likely be of great interest for regenerative therapies. Alternatively, an approach aimed at preventing or attenuating the development of the SASP, could achieve similar effects while avoiding cytotoxicity. High-throughput screens for compounds that selectively reduce the secretion of proteins by senescent cells allowed for the identification of two glucocorticoids, corticosterone and cortisol, as compounds capable of suppressing the secretion of factors that comprise the SASP (Laberge et al., 2012). Rapamycin, an inhibitor of the mammalian TORC1 complex, also decreases the SASP by suppressing IL1A translation and inhibiting NF-κB activity (Laberge et al., 2015). Inhibition of the JAK/STAT pathway, responsible for regulating the expression of a number of pro-inflammatory cytokines, was also shown to be a viable strategy to suppress the SASP in multiple senescent cell types (Xu et al., 2015). A better understanding of the signaling pathways that regulate the development of secretory phenotypes by senescent cells should provide the basis for selecting other drug candidates to interfere with the SASP. Such drugs would be of interest as co-adjuvants in SC-based regenerative therapies.
Age-related inflammation
Inflammaging, a chronic, low-grade inflammation observed during aging, has been proposed to be associated with the pathology of most age-related diseases (Franceschi and Campisi, 2014). Supporting this view, healthy aging in semi-supercentenarians is correlated with low levels of inflammatory markers (Arai et al., 2015). Evidence that inflammaging contributes to the loss of regenerative capacity of many tissues comes from mouse and fly models, as well as from humans: In murine models of chronic inflammation driven by over-activation of the NF-κB pathway, anti-inflammatory treatments can rescue regenerative capacity in the gut and liver (Jurk et al., 2014). NF-κB activation and secretion of high levels of pro-inflammatory cytokines is also observed in mouse models of progeria, and inhibition of NF-κB signaling prevents age-associated dysfunction in these animals (Osorio et al., 2012; Soria-Valles et al., 2015). Preventing chronic activation of the pro-inflammatory NFkB/Rel and JAK/STAT signaling pathways in the fly intestine promotes regenerative homeostasis and extends lifespan (Guo et al., 2014; Li et al., 2016b). In the mammalian skeletal muscle, an age specific increase in niche-derived NF‐κB signaling impairs satellite cells function, which can be attenuated by systemic administration of sodium salicylate, an FDA‐approved NF‐κB inhibitor (Oh et al., 2016). Observations in humans, in turn, suggest a central role for NF-κB activation and TNFα signaling in skin aging (Haustead et al., 2016). Evidence from heterochronic parabiosis also suggests that several serum-derived factors contributing to the age-related loss of regenerative capacity are molecules involved in the regulation of inflammation. Examples include the pro-inflammatory cytokine CCl11 and β2-microglobulin (a component of the major histocompatibility complex), which are increased in old serum and CSF and negatively impact neurogenesis (Smith et al., 2015; Villeda et al., 2011). These studies suggest that inhibiting the activity of these molecules may be an effective way to restore SC function during aging, at least in the brain.
Another interesting example for inflammatory signaling influencing regenerative capacity is the circulating ligand responsible for Wnt signaling-dependent loss of regenerative capacity in the old skeletal muscle (Brack et al., 2007). Recent findings indicate that this factor is the unconventional Wnt ligand C1qa, which belongs to the complement system (Naito et al., 2012). Complement-mediated activation is a hallmark of inflammatory conditions and age–related diseases. It is associated with activation of the Nlrp3 inflammasome in macrophages and leads to the production of pro-inflammatory cytokines (Bajic et al., 2015; Celkova et al., 2015). While the effects of C1qa on satellite cells were due to activation of Wnt signaling, it is tempting to speculate that the age-related elevation of C1qa levels in the serum may contribute to a systemic inflammatory condition. Anti-complement drugs, which are already used for therapy in many inflammatory disease conditions (Morgan and Harris, 2015), may thus also improve regeneration in old patients.
Chronic activation of inflammatory cytokine production in aging also impacts SC function directly, at least in the skin, and inhibition of JAK/STAT signaling in epidermal SCs can restore functionality (Doles et al., 2012). Hyperactivation of JAK/STAT with aging was also reported in the skeletal muscle, contributing to satellite cell loss of function by promoting myogenic commitment at the expenses of symmetric expansion. Notably, SC specific inhibition of JAK/STAT signaling improved satellite cell function and regenerative capacity in the old skeletal muscle (Price et al., 2014). In Drosophila, similar chronic activation of JAK/STAT signaling has been reported to impact gastric SC function, promoting metaplasia of the gastric epithelium. Limiting JAK/STAT activation in the gastric region promotes intestinal homeostasis and extends lifespan (Li et al., 2016b).
Another example of pro-inflammatory cytokines affecting SC function directly has recently been reported: Chronic exposure of HSCs to the pro-inflammatory cytokine IL-1 results in precocious myeloid differentiation at the expense of self-renewal (Pietras et al., 2016). In old mice, HSC differentiation is also skewed towards the myeloid lineage (Rossi et al., 2005) suggesting that chronic activation of inflammatory signaling in old animals may be part of the mechanism involved in HSC aging. It should be noted, however, that acute inflammatory signaling is essential for rapid myeloid recovery following injury (Mirantes et al., 2014). The idea that acute inflammatory singling is important for tissue repair is also supported by a recent study showing that at the skin wound-edge, aged keratinocytes fail to activate STAT3, perturbing the re-epithelization process following skin wounding (Keyes et al., 2016). While chronic activation of STAT signaling is detrimental for epidermal SCs (Doles et al., 2012), in this context Il-6 supplementation improves wound repair in aged skin (Keyes et al., 2016).
Together, these studies suggest that to improve the efficacy of regenerative processes and stem-cell based therapies, the development of effective strategies that limit ‘inflammaging’ may be critical (Table 1). Such approaches will likely involve attenuating the negative effects of systemic and local pro-inflammatory molecules that build up in aging organisms. Because the SASP is thought to be a major source of ‘inflammaging’, it is likely that the strategies described above to eliminate senescent cells or modulate the SASP would have similar beneficial effects.
Innate immune cell dysregulation and immune modulation
The activation of SCs in response to tissue damage is coordinated with the activation of immune cells, which in turn play a central role in the regulation of the regenerative process (reviewed in Aurora and Olson, 2014)). Because immune cells regulate and are regulated by inflammatory signals, targeting immune cells directly is likely to be another effective way to allay age-related inflammation and improve regeneration.
A common theme emerging from work in different tissue systems is that macrophages are required for the regenerative response and that a biphasic activation of the immune system is necessary for efficient regeneration in multiple organs (Arnold et al., 2007; Deonarine et al., 2007; Lucas et al., 2010; Miron et al., 2013; Perdiguero et al., 2011; Pull et al., 2005; Saclier et al., 2013; Varga et al., 2016). The early phase of the regenerative response is dominated by pro-inflammatory macrophages, derived from circulating monocytes, which participate in debris clearance and SC activation. Hemocytes (immune cells in invertebrates) also regulate SC activation in flies (Ayyaz et al., 2015), suggesting that the role of innate immune cells in the regulation of regeneration is evolutionarily conserved. Following the pro-inflammatory stimulus, macrophages shift into anti-inflammatory phenotypes that coordinate cell differentiation and tissue reconstruction. The shift is intrinsically coordinated in macrophages (Mounier et al., 2013; Perdiguero et al., 2011) and is blocked if phagocytosis is inhibited (Arnold et al., 2007; Aurora and Olson, 2014).
There are also examples in which macrophages acquire anti-inflammatory phenotypes, independently of previous inflammatory signals, such as macrophages recruited via a non-vascular route (Wang and Kubes, 2016) and tissue-resident macrophages (Jenkins et al., 2011; Jenkins et al., 2013). The latter exist in several organs, and include lung alveolar macrophages, liver Kupffer cells, epidermal Langerhans cells, and brain microglia and persist independently of peripheral recruitment (Guilliams et al., 2013; Hashimoto et al., 2013; Schulz et al., 2012). The relative contribution of local versus monocyte-derived macrophages to regeneration is still under investigation. Nevertheless, the importance of these cells as modulators of regenerative capacity, and the relevance of the pleiotropy of phenotypes involved in this regulatory process, that extend beyond only inflammatory and anti-inflammatory macrophages, are increasingly recognized (Wynn and Vannella, 2016). Thus, immune modulation targeted at harnessing the pro-repair functions of macrophages may be an effective way to promote regeneration in old patients. In fact, one important observation in studies using heterochronic parabiosis is that the positive effects of the young systemic environment on neural repair depends on the recruitment of young circulatory monocytes (Ruckh et al., 2012), and correlates with increased densities of young anti-inflammatory macrophages at lesion sites (Miron et al., 2013). This is attributed to non-cell autonomous effects of young monocytes recruited into the lesion site that are able to rejuvenate the lesion environment and promote an anti-inflammatory phenotype that is usually deficient in lesions from old mice (Miron et al., 2013). Aging of the innate immune system itself may thus contribute to the age-associated loss of regenerative capacity, and the immune modulatory potential of young macrophages may be sufficient to improve the endogenous regenerative potential of old tissues.
Aging of the innate immune system is complex. Old monocytes/macrophages are dysfunctional and less efficient in fighting infections, yet they also present a state of chronic activation associated with increased and persistent production of inflammatory cytokines (Hearps et al., 2012; Qian et al., 2012; Shaw et al., 2013). These cells are also deficient in phagocytic activity, a critical trigger for the anti-inflammatory switch in macrophage function during tissue repair. Impaired phagocytic activity of old macrophages may thus contribute to the deficiency in anti-inflammatory macrophages found at lesion sites. Moreover, tissue resident macrophages in old animals can also be dysfunctional and chronically activated (Damani et al., 2011), which can further contribute to compromise tissue regenerative capacity (Ekdahl et al., 2003).
Beyond macrophages, other classes of immune cells have also been implicated in the regenerative process. Dendritic epidermal T cells (DETCs) are essential for proliferation and tissue re-epithelialization after skin wounding (Jameson et al., 2002). Recent work showed that DETCs are regulated through an epithelial-immune crosstalk, where keratinocytes upregulate Skint genes required for activation and maintenance of DETCs (Keyes et al., 2016). This crosstalk is impaired in aged skin, where keratinocytes fail to activate DETCs, resulting in defective wound-healing and can be restored by re-activation of STAT-regulated Skint levels (Keyes et al., 2016). T regulatory cells (Tregs), have also received special attention recently due to their ability to dampen inflammatory responses and promote tissue repair in the skeletal muscle, heart, skin, kidney, and brain (Burzyn et al., 2013; Gandolfo et al., 2009; Nosbaum et al., 2016; Raposo et al., 2014; Villalta et al., 2014; Wan et al., 2015; Weirather et al., 2014). Tregs promote regeneration through several mechanisms, including the promotion of anti-inflammatory phenotypes of macrophages, reduction of effector T cells and associated pro-inflammatory signals, and activation of stem/progenitor cells (Lei et al., 2015). At least in the skeletal muscle, this mechanism of immune regulation is also affected by aging: Recruitment of regulatory T cells (Tregs) into skeletal muscle injury sites is impaired in old animals, contributing to age-related loss of regenerative capacity (Kuswanto et al., 2016). This deficiency is associated with impaired production of Il-33 locally and Il-33 supplementation can restore regenerative capacity of the skeletal muscle (Kuswanto et al., 2016). Fibro-Adipogenic Progenitor (FAP)-like cells are a major source of Il-33, and were found to be localized preferentially within nerve structures. These findings highlight the importance of stromal cells (FAPs) as a source of immune modulatory factors during regeneration, and suggest that neuro-immune interactions may also be important regulators of skeletal muscle regenerative capacity in aging.
Several other lymphoid cell types have been implicated in the regenerative process, particularly in barrier epithelia such as the intestine, where immune cells participate in the defense against pathogen infection in addition to their role in tissue repair (Karin and Clevers, 2016). In these systems, neuro-immune interactions are also important coordinators of regeneration and inflammatory responses (Veiga-Fernandes and Mucida, 2016): In the intestine, innate lymphoid cells sense damage through glia-derived neurotrophic signals. In response, these cells upregulate Il-22 production, a cytokine important in gut inflammation (Ibiza et al., 2016) and intestinal SC function (Lindemans et al., 2015). The effects of aging in neuro-immune interactions in barrier epithelia are yet to be determined. Nevertheless, de-regulation of neuro-immune interactions in the brain, has been reported to play an important role in aging. NF-κB activation in hypothalamic microglia (the resident immune cells of the central nervous system) promotes TNFα production, which stimulates NF-κB signaling in local neurons. In turn, increased NF-kappa B signaling in hypothalamic neurons, results in epigenetic repression of the GnRH gene, leading to reduced GnRH release which accelerates the development of several age-related phenotypes (Zhang et al., 2013b). Importantly, GnRH restoration can rescue several age-related physiological dysfunctions, including the loss of neural SC function, suggesting that immune inhibition in the hypothalamus may represent a potential strategy to improve age-associated impairments systemically, by regulating neuro-immune interactions locally.
The findings described above introduce immune modulation aimed towards harnessing the anti-inflammatory potential of immune cells to promote endogenous repair mechanisms as a promising strategy to improve regenerative capacity in aging tissues (Table 1). This strategy likely applies not only to tissues where regeneration is sustained by an endogenous adult SC population, but also in cell-replacement based regenerative therapies. A proof of principle for such an approach is a recent study in which mesencephalic astrocyte-derived neurotrophic factor (MANF) was used to enhance the success of cell-replacement therapies in the retina, improving cell integration and vision recovery following transplantation (Neves et al., 2016). In this study, we discovered that MANF, a previously described neurotrophic factor derived from neural glia, is also expressed in immune cells and has an autocrine immune modulatory function promoting anti-inflammatory activation. Importantly, the immune modulatory function of MANF is essential for its neuroprotective and tissue-repair promoting activities in the retina (Neves et al., 2016), highlighting the importance of immune modulation in regenerative success. IGF-1 is yet another neurotrophic factor also shown to improve the success of cell-replacement therapies in the retina, when delivered via viral vectors prior to transplantation (West et al., 2012). Notably, in other regenerative and inflammatory contexts, IGF-1 also displays an immune modulatory function that alleviates inflammation by promoting anti-inflammatory phenotypes of macrophages in an autocrine manner (Tonkin et al., 2015) or by promoting recruitment of Tregs with anti-inflammatory activity (Johannesson et al., 2014). These functions are likely contributors to the improved regenerative success observed in the retina. The beneficial effect of IGF-1 in regenerative capacity, however, extends beyond the regulation of inflammation, suggesting a broad effect on different cellular targets (Mourkioti and Rosenthal, 2005). The multitasking capacity of neurotrophic factors in the context of a regenerative response was also appreciated in the hematopoietic system. HSCs express the GDNF family of ligands (GFLs) receptor RET, and GFL signaling through RET in HSCs promotes survival, expansion and in vivo transplantation efficiency (Fonseca-Pereira et al., 2014). Although none of these neurotrophic factors has been specifically tested in the aging context as a means to improve regeneration, they represent good candidates to modulate regenerative capacity by simultaneously promoting neuroprotection, modulating the immune environment and enhancing SC function.
Another recent study has provided further evidence for the usefulness of immune modulation in the success of regenerative therapies: In this case, extracellular matrix-derived biomaterial scaffolds are used to promote regenerative success by modulating the immune microenvironment in traumatic muscle wounds (Sadtler et al., 2016). The effects are mediated by T helper 2 cells, which promote anti-inflammatory polarization of local macrophages in an Il-4 dependent manner, potentiating functional tissue regeneration (Sadtler et al., 2016). Notably, the local administration of the scaffolds influences T cell responses systemically. This suggests that local biomaterial based therapies may promote type 2 immunity systemically, which may be beneficial in situations where chronic inflammation is detrimental for regenerative capacity in several tissues, as it happens in aging.
CONCLUSIONS AND PERSPECTIVES
Advances in regenerative medicine and the advent of SC therapies, now progressing into clinical trials (Trounson and McDonald, 2015), will likely elucidate current limitations to the application of SCs to treat disease. Because older individuals are more likely to suffer from chronic degenerative conditions that would benefit from regenerative therapies, it is imperative to address the roadblocks imposed by aging to SC-based therapeutic interventions. Our growing understanding of the basic mechanisms of organismal aging has uncovered critical limitations in the repair capacity of old tissues and highlights the necessity to coordinate SC specific interventions with niche and systemic modulation to improve regenerative success.
In the work discussed here, we point to interventions with significant success in pre-clinical studies (Table 1). These include ablation of senescent cells, inhibition of inflammatory signaling, modulation of immune cell phenotypes, and inhibition of different intracellular signaling pathways chronically activated during aging. While so far promising, the translational success of these early studies will require dealing with two major questions: First, the pathways targeted by these interventions are often important physiological regulators of regeneration, and their acute activation is critical for regenerative success. Thus therapeutic strategies should aim to modulate pathway activity for averting chronic activation rather than complete suppression. Secondly, systemic and local delivery of effector molecules may have effects in multiple cell types simultaneously, sometimes with opposing outcomes. Thus, identifying the cellular targets of each intervention is an important next step to ensure that the clinical translation of these therapies takes into account possible side effects. We anticipate that further studies in defined model systems will facilitate addressing these questions.
Acknowledgments
H.J. is supported by AG047497 and AG052989. J.N. and P.S.V. are supported by the Glenn Foundation for Medical Research.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the productionprocess errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Acosta JC, Banito A, Wuestefeld T, Georgilis A, Janich P, Morton JP, Athineos D, Kang TW, Lasitschka F, Andrulis M, et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nature cell biology. 2013;15:978–990. doi: 10.1038/ncb2784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akintola AD, Crislip ZL, Catania JM, Chen G, Zimmer WE, Burghardt RC, Parrish AR. Promoter methylation is associated with the age-dependent loss of N-cadherin in the rat kidney. American journal of physiology Renal physiology. 2008;294:F170–176. doi: 10.1152/ajprenal.00285.2007. [DOI] [PubMed] [Google Scholar]
- Arai Y, Martin-Ruiz CM, Takayama M, Abe Y, Takebayashi T, Koyasu S, Suematsu M, Hirose N, von Zglinicki T. Inflammation, But Not Telomere Length, Predicts Successful Ageing at Extreme Old Age: A Longitudinal Study of Semi-supercentenarians. EBioMedicine. 2015;2:1549–1558. doi: 10.1016/j.ebiom.2015.07.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold L, Henry A, Poron F, Baba-Amer Y, van Rooijen N, Plonquet A, Gherardi RK, Chazaud B. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. The Journal of experimental medicine. 2007;204:1057–1069. doi: 10.1084/jem.20070075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aurora AB, Olson EN. Immune modulation of stem cells and regeneration. Cell stem cell. 2014;15:14–25. doi: 10.1016/j.stem.2014.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayyaz A, Li H, Jasper H. Haemocytes control stem cell activity in the Drosophila intestine. Nature cell biology. 2015;17:736–748. doi: 10.1038/ncb3174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baht GS, Silkstone D, Vi L, Nadesan P, Amani Y, Whetstone H, Wei Q, Alman BA. Exposure to a youthful circulaton rejuvenates bone repair through modulation of beta-catenin. Nature communications. 2015;6:7131. doi: 10.1038/ncomms8131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajic G, Degn SE, Thiel S, Andersen GR. Complement activation, regulation, and molecular basis for complement-related diseases. The EMBO journal. 2015;34:2735–2757. doi: 10.15252/embj.201591881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker DJ, Childs BG, Durik M, Wijers ME, Sieben CJ, Zhong J, Saltness RA, Jeganathan KB, Verzosa GC, Pezeshki A, et al. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature. 2016;530:184–189. doi: 10.1038/nature16932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, Kirkland JL, van Deursen JM. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011;479:232–236. doi: 10.1038/nature10600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banito A, Rashid ST, Acosta JC, Li S, Pereira CF, Geti I, Pinho S, Silva JC, Azuara V, Walsh M, et al. Senescence impairs successful reprogramming to pluripotent stem cells. Genes & development. 2009;23:2134–2139. doi: 10.1101/gad.1811609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barker N, Bartfeld S, Clevers H. Tissue-resident adult stem cell populations of rapidly self-renewing organs. Cell stem cell. 2010;7:656–670. doi: 10.1016/j.stem.2010.11.016. [DOI] [PubMed] [Google Scholar]
- Beerman I, Seita J, Inlay MA, Weissman IL, Rossi DJ. Quiescent hematopoietic stem cells accumulate DNA damage during aging that is repaired upon entry into cell cycle. Cell stem cell. 2014;15:37–50. doi: 10.1016/j.stem.2014.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernet JD, Doles JD, Hall JK, Kelly Tanaka K, Carter TA, Olwin BB. p38 MAPK signaling underlies a cell-autonomous loss of stem cell self-renewal in skeletal muscle of aged mice. Nature medicine. 2014;20:265–271. doi: 10.1038/nm.3465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjornsson CS, Apostolopoulou M, Tian Y, Temple S. It takes a village: constructing the neurogenic niche. Developmental cell. 2015;32:435–446. doi: 10.1016/j.devcel.2015.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brack AS, Conboy MJ, Roy S, Lee M, Kuo CJ, Keller C, Rando TA. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science. 2007;317:807–810. doi: 10.1126/science.1144090. [DOI] [PubMed] [Google Scholar]
- Burkauskiene A, Mackiewicz Z, Virtanen I, Konttinen YT. Age-related changes in myocardial nerve and collagen networks of the auricle of the right atrium. Acta cardiologica. 2006;61:513–518. doi: 10.2143/AC.61.5.2017765. [DOI] [PubMed] [Google Scholar]
- Burzyn D, Kuswanto W, Kolodin D, Shadrach JL, Cerletti M, Jang Y, Sefik E, Tan TG, Wagers AJ, Benoist C, et al. A special population of regulatory T cells potentiates muscle repair. Cell. 2013;155:1282–1295. doi: 10.1016/j.cell.2013.10.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calabresi C, Arosio B, Galimberti L, Scanziani E, Bergottini R, Annoni G, Vergani C. Natural aging, expression of fibrosis-related genes and collagen deposition in rat lung. Experimental gerontology. 2007;42:1003–1011. doi: 10.1016/j.exger.2007.06.016. [DOI] [PubMed] [Google Scholar]
- Campisi J, d’Adda di Fagagna F. Cellular senescence: when bad things happen to good cells. Nature reviews Molecular cell biology. 2007;8:729–740. doi: 10.1038/nrm2233. [DOI] [PubMed] [Google Scholar]
- Celkova L, Doyle SL, Campbell M. NLRP3 Inflammasome and Pathobiology in AMD. Journal of clinical medicine. 2015;4:172–192. doi: 10.3390/jcm4010172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakkalakal JV, Jones KM, Basson MA, Brack AS. The aged niche disrupts muscle stem cell quiescence. Nature. 2012;490:355–360. doi: 10.1038/nature11438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang J, Wang Y, Shao L, Laberge RM, Demaria M, Campisi J, Janakiraman K, Sharpless NE, Ding S, Feng W, et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nature medicine. 2016;22:78–83. doi: 10.1038/nm.4010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C, Liu Y, Zheng P. mTOR regulation and therapeutic rejuvenation of aging hematopoietic stem cells. Science signaling. 2009;2:ra75. doi: 10.1126/scisignal.2000559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen W, Jongkamonwiwat N, Abbas L, Eshtan SJ, Johnson SL, Kuhn S, Milo M, Thurlow JK, Andrews PW, Marcotti W, et al. Restoration of auditory evoked responses by human ES-cell-derived otic progenitors. Nature. 2012;490:278–282. doi: 10.1038/nature11415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheung TH, Rando TA. Molecular regulation of stem cell quiescence. Nature reviews Molecular cell biology. 2013;14:329–340. doi: 10.1038/nrm3591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiche A, Le Roux I, von Joest M, Sakai H, Aguin SB, Cazin C, Salam R, Fiette L, Alegria O, Flamant P, et al. Injury-Induced Senescence Enables In Vivo Reprogramming in Skeletal Muscle. Cell stem cell. 2016 doi: 10.1016/j.stem.2016.11.020. [DOI] [PubMed] [Google Scholar]
- Choi YC, Choi JS, Woo CH, Cho YW. Stem cell delivery systems inspired by tissue-specific niches. Journal of controlled release : official journal of the Controlled Release Society. 2014;193:42–50. doi: 10.1016/j.jconrel.2014.06.032. [DOI] [PubMed] [Google Scholar]
- Conboy IM, Conboy MJ, Wagers AJ, Girma ER, Weissman IL, Rando TA. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature. 2005;433:760–764. doi: 10.1038/nature03260. [DOI] [PubMed] [Google Scholar]
- Conway A, Schaffer DV. Biophysical regulation of stem cell behavior within the niche. Stem cell research & therapy. 2012;3:50. doi: 10.1186/scrt141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coppe JP, Desprez PY, Krtolica A, Campisi J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annual review of pathology. 2010;5:99–118. doi: 10.1146/annurev-pathol-121808-102144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cosgrove BD, Gilbert PM, Porpiglia E, Mourkioti F, Lee SP, Corbel SY, Llewellyn ME, Delp SL, Blau HM. Rejuvenation of the muscle stem cell population restores strength to injured aged muscles. Nature medicine. 2014;20:255–264. doi: 10.1038/nm.3464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cousin W, Ho ML, Desai R, Tham A, Chen RY, Kung S, Elabd C, Conboy IM. Regenerative capacity of old muscle stem cells declines without significant accumulation of DNA damage. PloS one. 2013;8:e63528. doi: 10.1371/journal.pone.0063528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalby MJ, Gadegaard N, Curtis AS, Oreffo RO. Nanotopographical control of human osteoprogenitor differentiation. Current stem cell research & therapy. 2007;2:129–138. doi: 10.2174/157488807780599220. [DOI] [PubMed] [Google Scholar]
- Damani MR, Zhao L, Fontainhas AM, Amaral J, Fariss RN, Wong WT. Age-related alterations in the dynamic behavior of microglia. Aging cell. 2011;10:263–276. doi: 10.1111/j.1474-9726.2010.00660.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demaria M, Ohtani N, Youssef SA, Rodier F, Toussaint W, Mitchell JR, Laberge RM, Vijg J, Van Steeg H, Dolle ME, et al. An essential role for senescent cells in optimal wound healing through secretion of PDGF-AA. Developmental cell. 2014;31:722–733. doi: 10.1016/j.devcel.2014.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deonarine K, Panelli MC, Stashower ME, Jin P, Smith K, Slade HB, Norwood C, Wang E, Marincola FM, Stroncek DF. Gene expression profiling of cutaneous wound healing. Journal of translational medicine. 2007;5:11. doi: 10.1186/1479-5876-5-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doles J, Storer M, Cozzuto L, Roma G, Keyes WM. Age-associated inflammation inhibits epidermal stem cell function. Genes & development. 2012;26:2144–2153. doi: 10.1101/gad.192294.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egerman MA, Cadena SM, Gilbert JA, Meyer A, Nelson HN, Swalley SE, Mallozzi C, Jacobi C, Jennings LL, Clay I, et al. GDF11 Increases with Age and Inhibits Skeletal Muscle Regeneration. Cell metabolism. 2015;22:164–174. doi: 10.1016/j.cmet.2015.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekdahl CT, Claasen JH, Bonde S, Kokaia Z, Lindvall O. Inflammation is detrimental for neurogenesis in adult brain. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:13632–13637. doi: 10.1073/pnas.2234031100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elabd C, Cousin W, Upadhyayula P, Chen RY, Chooljian MS, Li J, Kung S, Jiang KP, Conboy IM. Oxytocin is an age-specific circulating hormone that is necessary for muscle maintenance and regeneration. Nature communications. 2014;5:4082. doi: 10.1038/ncomms5082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flach J, Bakker ST, Mohrin M, Conroy PC, Pietras EM, Reynaud D, Alvarez S, Diolaiti ME, Ugarte F, Forsberg EC, et al. Replication stress is a potent driver of functional decline in ageing haematopoietic stem cells. Nature. 2014;512:198–202. doi: 10.1038/nature13619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Florian MC, Dorr K, Niebel A, Daria D, Schrezenmeier H, Rojewski M, Filippi MD, Hasenberg A, Gunzer M, Scharffetter-Kochanek K, et al. Cdc42 activity regulates hematopoietic stem cell aging and rejuvenation. Cell stem cell. 2012;10:520–530. doi: 10.1016/j.stem.2012.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Florian MC, Nattamai KJ, Dorr K, Marka G, Uberle B, Vas V, Eckl C, Andra I, Schiemann M, Oostendorp RA, et al. A canonical to non-canonical Wnt signalling switch in haematopoietic stem-cell ageing. Nature. 2013;503:392–396. doi: 10.1038/nature12631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonseca-Pereira D, Arroz-Madeira S, Rodrigues-Campos M, Barbosa IA, Domingues RG, Bento T, Almeida AR, Ribeiro H, Potocnik AJ, Enomoto H, et al. The neurotrophic factor receptor RET drives haematopoietic stem cell survival and function. Nature. 2014;514:98–101. doi: 10.1038/nature13498. [DOI] [PubMed] [Google Scholar]
- Forbes SJ, Rosenthal N. Preparing the ground for tissue regeneration: from mechanism to therapy. Nature medicine. 2014;20:857–869. doi: 10.1038/nm.3653. [DOI] [PubMed] [Google Scholar]
- Fox IJ, Daley GQ, Goldman SA, Huard J, Kamp TJ, Trucco M. Stem cell therapy. Use of differentiated pluripotent stem cells as replacement therapy for treating disease. Science. 2014;345:1247391. doi: 10.1126/science.1247391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. The journals of gerontology Series A, Biological sciences and medical sciences. 2014;69(Suppl 1):S4–9. doi: 10.1093/gerona/glu057. [DOI] [PubMed] [Google Scholar]
- Frantz C, Stewart KM, Weaver VM. The extracellular matrix at a glance. Journal of cell science. 2010;123:4195–4200. doi: 10.1242/jcs.023820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fry CS, Kirby TJ, Kosmac K, McCarthy JJ, Peterson CA. Myogenic Progenitor Cells Control Extracellular Matrix Production by Fibroblasts during Skeletal Muscle Hypertrophy. Cell stem cell. 2017;20:56–69. doi: 10.1016/j.stem.2016.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuchs E, Chen T. A matter of life and death: self-renewal in stem cells. EMBO reports. 2013;14:39–48. doi: 10.1038/embor.2012.197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gagliano N, Arosio B, Santambrogio D, Balestrieri MR, Padoani G, Tagliabue J, Masson S, Vergani C, Annoni G. Age-dependent expression of fibrosis-related genes and collagen deposition in rat kidney cortex. The journals of gerontology Series A, Biological sciences and medical sciences. 2000;55:B365–372. doi: 10.1093/gerona/55.8.b365. [DOI] [PubMed] [Google Scholar]
- Gandolfo MT, Jang HR, Bagnasco SM, Ko GJ, Agreda P, Satpute SR, Crow MT, King LS, Rabb H. Foxp3+ regulatory T cells participate in repair of ischemic acute kidney injury. Kidney international. 2009;76:717–729. doi: 10.1038/ki.2009.259. [DOI] [PubMed] [Google Scholar]
- Garcia-Prat L, Martinez-Vicente M, Perdiguero E, Ortet L, Rodriguez-Ubreva J, Rebollo E, Ruiz-Bonilla V, Gutarra S, Ballestar E, Serrano AL, et al. Autophagy maintains stemness by preventing senescence. Nature. 2016;529:37–42. doi: 10.1038/nature16187. [DOI] [PubMed] [Google Scholar]
- Gilbert PM, Havenstrite KL, Magnusson KE, Sacco A, Leonardi NA, Kraft P, Nguyen NK, Thrun S, Lutolf MP, Blau HM. Substrate elasticity regulates skeletal muscle stem cell self-renewal in culture. Science. 2010;329:1078–1081. doi: 10.1126/science.1191035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo L, Karpac J, Tran SL, Jasper H. PGRP-SC2 promotes gut immune homeostasis to limit commensal dysbiosis and extend lifespan. Cell. 2014;156:109–122. doi: 10.1016/j.cell.2013.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haustead DJ, Stevenson A, Saxena V, Marriage F, Firth M, Silla R, Martin L, Adcroft KF, Rea S, Day PJ, et al. Transcriptome analysis of human ageing in male skin shows mid-life period of variability and central role of NF-kappaB. Scientific reports. 2016;6:26846. doi: 10.1038/srep26846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hearps AC, Martin GE, Angelovich TA, Cheng WJ, Maisa A, Landay AL, Jaworowski A, Crowe SM. Aging is associated with chronic innate immune activation and dysregulation of monocyte phenotype and function. Aging cell. 2012;11:867–875. doi: 10.1111/j.1474-9726.2012.00851.x. [DOI] [PubMed] [Google Scholar]
- Herbig U, Ferreira M, Condel L, Carey D, Sedivy JM. Cellular senescence in aging primates. Science. 2006;311:1257. doi: 10.1126/science.1122446. [DOI] [PubMed] [Google Scholar]
- Hinken AC, Powers JM, Luo G, Holt JA, Billin AN, Russell AJ. Lack of evidence for GDF11 as a rejuvenator of aged skeletal muscle satellite cells. Aging cell. 2016;15:582–584. doi: 10.1111/acel.12475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang P, He Z, Ji S, Sun H, Xiang D, Liu C, Hu Y, Wang X, Hui L. Induction of functional hepatocyte-like cells from mouse fibroblasts by defined factors. Nature. 2011;475:386–389. doi: 10.1038/nature10116. [DOI] [PubMed] [Google Scholar]
- Ibiza S, Garcia-Cassani B, Ribeiro H, Carvalho T, Almeida L, Marques R, Misic AM, Bartow-McKenney C, Larson DM, Pavan WJ, et al. Glial-cell-derived neuroregulators control type 3 innate lymphoid cells and gut defence. Nature. 2016;535:440–443. doi: 10.1038/nature18644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ieda M, Fu JD, Delgado-Olguin P, Vedantham V, Hayashi Y, Bruneau BG, Srivastava D. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell. 2010;142:375–386. doi: 10.1016/j.cell.2010.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jameson J, Ugarte K, Chen N, Yachi P, Fuchs E, Boismenu R, Havran WL. A role for skin gammadelta T cells in wound repair. Science. 2002;296:747–749. doi: 10.1126/science.1069639. [DOI] [PubMed] [Google Scholar]
- Janzen V, Forkert R, Fleming HE, Saito Y, Waring MT, Dombkowski DM, Cheng T, DePinho RA, Sharpless NE, Scadden DT. Stem-cell ageing modified by the cyclin-dependent kinase inhibitor p16INK4a. Nature. 2006;443:421–426. doi: 10.1038/nature05159. [DOI] [PubMed] [Google Scholar]
- Jeyapalan JC, Ferreira M, Sedivy JM, Herbig U. Accumulation of senescent cells in mitotic tissue of aging primates. Mechanisms of ageing and development. 2007;128:36–44. doi: 10.1016/j.mad.2006.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johannesson B, Sattler S, Semenova E, Pastore S, Kennedy-Lydon TM, Sampson RD, Schneider MD, Rosenthal N, Bilbao D. Insulin-like growth factor-1 induces regulatory T cell-mediated suppression of allergic contact dermatitis in mice. Disease models & mechanisms. 2014;7:977–985. doi: 10.1242/dmm.015362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson SC, Rabinovitch PS, Kaeberlein M. mTOR is a key modulator of ageing and age-related disease. Nature. 2013;493:338–345. doi: 10.1038/nature11861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones DL, Wagers AJ. No place like home: anatomy and function of the stem cell niche. Nature reviews Molecular cell biology. 2008;9:11–21. doi: 10.1038/nrm2319. [DOI] [PubMed] [Google Scholar]
- Jurk D, Wilson C, Passos JF, Oakley F, Correia-Melo C, Greaves L, Saretzki G, Fox C, Lawless C, Anderson R, et al. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nature communications. 2014;2:4172. doi: 10.1038/ncomms5172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang E, Wang X, Tippner-Hedges R, Ma H, Folmes CD, Gutierrez NM, Lee Y, Van Dyken C, Ahmed R, Li Y, et al. Age-Related Accumulation of Somatic Mitochondrial DNA Mutations in Adult-Derived Human iPSCs. Cell stem cell. 2016;18:625–636. doi: 10.1016/j.stem.2016.02.005. [DOI] [PubMed] [Google Scholar]
- Kang TW, Yevsa T, Woller N, Hoenicke L, Wuestefeld T, Dauch D, Hohmeyer A, Gereke M, Rudalska R, Potapova A, et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature. 2011;479:547–551. doi: 10.1038/nature10599. [DOI] [PubMed] [Google Scholar]
- Karin M, Clevers H. Reparative inflammation takes charge of tissue regeneration. Nature. 2016;529:307–315. doi: 10.1038/nature17039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katsimpardi L, Litterman NK, Schein PA, Miller CM, Loffredo FS, Wojtkiewicz GR, Chen JW, Lee RT, Wagers AJ, Rubin LL. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science. 2014;344:630–634. doi: 10.1126/science.1251141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keyes BE, Liu S, Asare A, Naik S, Levorse J, Polak L, Lu CP, Nikolova M, Pasolli HA, Fuchs E. Impaired Epidermal to Dendritic T Cell Signaling Slows Wound Repair in Aged Skin. Cell. 2016;167:1323–1338 e1314. doi: 10.1016/j.cell.2016.10.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim WY, Sharpless NE. The regulation of INK4/ARF in cancer and aging. Cell. 2006;127:265–275. doi: 10.1016/j.cell.2006.10.003. [DOI] [PubMed] [Google Scholar]
- Krizhanovsky V, Yon M, Dickins RA, Hearn S, Simon J, Miething C, Yee H, Zender L, Lowe SW. Senescence of activated stellate cells limits liver fibrosis. Cell. 2008;134:657–667. doi: 10.1016/j.cell.2008.06.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kusumbe AP, Ramasamy SK, Itkin T, Mae MA, Langen UH, Betsholtz C, Lapidot T, Adams RH. Age-dependent modulation of vascular niches for haematopoietic stem cells. Nature. 2016;532:380–384. doi: 10.1038/nature17638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuswanto W, Burzyn D, Panduro M, Wang KK, Jang YC, Wagers AJ, Benoist C, Mathis D. Poor Repair of Skeletal Muscle in Aging Mice Reflects a Defect in Local, Interleukin-33-Dependent Accumulation of Regulatory T Cells. Immunity. 2016;44:355–367. doi: 10.1016/j.immuni.2016.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwak HB. Aging, exercise, and extracellular matrix in the heart. Journal of exercise rehabilitation. 2013;9:338–347. doi: 10.12965/jer.130049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laberge RM, Sun Y, Orjalo AV, Patil CK, Freund A, Zhou L, Curran SC, Davalos AR, Wilson-Edell KA, Liu S, et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nature cell biology. 2015;17:1049–1061. doi: 10.1038/ncb3195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laberge RM, Zhou L, Sarantos MR, Rodier F, Freund A, de Keizer PL, Liu S, Demaria M, Cong YS, Kapahi P, et al. Glucocorticoids suppress selected components of the senescence-associated secretory phenotype. Aging cell. 2012;11:569–578. doi: 10.1111/j.1474-9726.2012.00818.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamba DA, Gust J, Reh TA. Transplantation of human embryonic stem cell-derived photoreceptors restores some visual function in Crx-deficient mice. Cell stem cell. 2009;4:73–79. doi: 10.1016/j.stem.2008.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lane SW, Williams DA, Watt FM. Modulating the stem cell niche for tissue regeneration. Nature biotechnology. 2014;32:795–803. doi: 10.1038/nbt.2978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lei H, Schmidt-Bleek K, Dienelt A, Reinke P, Volk HD. Regulatory T cell-mediated anti-inflammatory effects promote successful tissue repair in both indirect and direct manners. Frontiers in pharmacology. 2015;6:184. doi: 10.3389/fphar.2015.00184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemos DR, Babaeijandaghi F, Low M, Chang CK, Lee ST, Fiore D, Zhang RH, Natarajan A, Nedospasov SA, Rossi FM. Nilotinib reduces muscle fibrosis in chronic muscle injury by promoting TNF-mediated apoptosis of fibro/adipogenic progenitors. Nature medicine. 2015;21:786–794. doi: 10.1038/nm.3869. [DOI] [PubMed] [Google Scholar]
- Li C, Zhen G, Chai Y, Xie L, Crane JL, Farber E, Farber CR, Luo X, Gao P, Cao X, et al. RhoA determines lineage fate of mesenchymal stem cells by modulating CTGF-VEGF complex in extracellular matrix. Nature communications. 2016a;7:11455. doi: 10.1038/ncomms11455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H, Collado M, Villasante A, Strati K, Ortega S, Canamero M, Blasco MA, Serrano M. The Ink4/Arf locus is a barrier for iPS cell reprogramming. Nature. 2009;460:1136–1139. doi: 10.1038/nature08290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H, Qi Y, Jasper H. Preventing Age-Related Decline of Gut Compartmentalization Limits Microbiota Dysbiosis and Extends Lifespan. Cell host & microbe. 2016b;19:240–253. doi: 10.1016/j.chom.2016.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindemans CA, Calafiore M, Mertelsmann AM, O’Connor MH, Dudakov JA, Jenq RR, Velardi E, Young LF, Smith OM, Lawrence G, et al. Interleukin-22 promotes intestinal-stem-cell-mediated epithelial regeneration. Nature. 2015;528:560–564. doi: 10.1038/nature16460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu L, Cheung TH, Charville GW, Hurgo BM, Leavitt T, Shih J, Brunet A, Rando TA. Chromatin modifications as determinants of muscle stem cell quiescence and chronological aging. Cell reports. 2013;4:189–204. doi: 10.1016/j.celrep.2013.05.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu L, Rando TA. Manifestations and mechanisms of stem cell aging. The Journal of cell biology. 2011;193:257–266. doi: 10.1083/jcb.201010131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loffredo FS, Steinhauser ML, Jay SM, Gannon J, Pancoast JR, Yalamanchi P, Sinha M, Dall’Osso C, Khong D, Shadrach JL, et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell. 2013;153:828–839. doi: 10.1016/j.cell.2013.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu WY, Bird TG, Boulter L, Tsuchiya A, Cole AM, Hay T, Guest RV, Wojtacha D, Man TY, Mackinnon A, et al. Hepatic progenitor cells of biliary origin with liver repopulation capacity. Nature cell biology. 2015;17:971–983. doi: 10.1038/ncb3203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucas T, Waisman A, Ranjan R, Roes J, Krieg T, Muller W, Roers A, Eming SA. Differential roles of macrophages in diverse phases of skin repair. J Immunol. 2010;184:3964–3977. doi: 10.4049/jimmunol.0903356. [DOI] [PubMed] [Google Scholar]
- Lukjanenko L, Jung MJ, Hegde N, Perruisseau-Carrier C, Migliavacca E, Rozo M, Karaz S, Jacot G, Schmidt M, Li L, et al. Loss of fibronectin from the aged stem cell niche affects the regenerative capacity of skeletal muscle in mice. Nature medicine. 2016 doi: 10.1038/nm.4126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacLaren RE, Pearson RA, MacNeil A, Douglas RH, Salt TE, Akimoto M, Swaroop A, Sowden JC, Ali RR. Retinal repair by transplantation of photoreceptor precursors. Nature. 2006;444:203–207. doi: 10.1038/nature05161. [DOI] [PubMed] [Google Scholar]
- Marion RM, Strati K, Li H, Murga M, Blanco R, Ortega S, Fernandez-Capetillo O, Serrano M, Blasco MA. A p53-mediated DNA damage response limits reprogramming to ensure iPS cell genomic integrity. Nature. 2009a;460:1149–1153. doi: 10.1038/nature08287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marion RM, Strati K, Li H, Tejera A, Schoeftner S, Ortega S, Serrano M, Blasco MA. Telomeres acquire embryonic stem cell characteristics in induced pluripotent stem cells. Cell stem cell. 2009b;4:141–154. doi: 10.1016/j.stem.2008.12.010. [DOI] [PubMed] [Google Scholar]
- Martin N, Beach D, Gil J. Ageing as developmental decay: insights from p16(INK4a) Trends in molecular medicine. 2014;20:667–674. doi: 10.1016/j.molmed.2014.09.008. [DOI] [PubMed] [Google Scholar]
- Matsumura H, Mohri Y, Binh NT, Morinaga H, Fukuda M, Ito M, Kurata S, Hoeijmakers J, Nishimura EK. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science. 2016;351:aad4395. doi: 10.1126/science.aad4395. [DOI] [PubMed] [Google Scholar]
- Matsuura K, Utoh R, Nagase K, Okano T. Cell sheet approach for tissue engineering and regenerative medicine. Journal of controlled release : official journal of the Controlled Release Society. 2014;190:228–239. doi: 10.1016/j.jconrel.2014.05.024. [DOI] [PubMed] [Google Scholar]
- McBeath R, Pirone DM, Nelson CM, Bhadriraju K, Chen CS. Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Developmental cell. 2004;6:483–495. doi: 10.1016/s1534-5807(04)00075-9. [DOI] [PubMed] [Google Scholar]
- Mertens J, Paquola AC, Ku M, Hatch E, Bohnke L, Ladjevardi S, McGrath S, Campbell B, Lee H, Herdy JR, et al. Directly Reprogrammed Human Neurons Retain Aging-Associated Transcriptomic Signatures and Reveal Age-Related Nucleocytoplasmic Defects. Cell stem cell. 2015;17:705–718. doi: 10.1016/j.stem.2015.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller JD, Ganat YM, Kishinevsky S, Bowman RL, Liu B, Tu EY, Mandal PK, Vera E, Shim JW, Kriks S, et al. Human iPSC-based modeling of late-onset disease via progerin-induced aging. Cell stem cell. 2013;13:691–705. doi: 10.1016/j.stem.2013.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirantes C, Passegue E, Pietras EM. Pro-inflammatory cytokines: emerging players regulating HSC function in normal and diseased hematopoiesis. Experimental cell research. 2014;329:248–254. doi: 10.1016/j.yexcr.2014.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miron VE, Boyd A, Zhao JW, Yuen TJ, Ruckh JM, Shadrach JL, van Wijngaarden P, Wagers AJ, Williams A, Franklin RJ, et al. M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nature neuroscience. 2013;16:1211–1218. doi: 10.1038/nn.3469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan BP, Harris CL. Complement, a target for therapy in inflammatory and degenerative diseases. Nature reviews Drug discovery. 2015;14:857–877. doi: 10.1038/nrd4657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgner J, Ghatak S, Jakobi T, Dieterich C, Aumailley M, Wickstrom SA. Integrin-linked kinase regulates the niche of quiescent epidermal stem cells. Nature communications. 2015;6:8198. doi: 10.1038/ncomms9198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mosteiro L, Pantoja C, Alcazar N, Marion RM, Chondronasiou D, Rovira M, Fernandez-Marcos PJ, Munoz-Martin M, Blanco-Aparicio C, Pastor J, et al. Tissue damage and senescence provide critical signals for cellular reprogramming in vivo. Science. 2016;354 doi: 10.1126/science.aaf4445. [DOI] [PubMed] [Google Scholar]
- Mounier R, Theret M, Arnold L, Cuvellier S, Bultot L, Goransson O, Sanz N, Ferry A, Sakamoto K, Foretz M, et al. AMPKalpha1 regulates macrophage skewing at the time of resolution of inflammation during skeletal muscle regeneration. Cell metabolism. 2013;18:251–264. doi: 10.1016/j.cmet.2013.06.017. [DOI] [PubMed] [Google Scholar]
- Mourkioti F, Rosenthal N. IGF-1, inflammation and stem cells: interactions during muscle regeneration. Trends in immunology. 2005;26:535–542. doi: 10.1016/j.it.2005.08.002. [DOI] [PubMed] [Google Scholar]
- Munoz-Espin D, Canamero M, Maraver A, Gomez-Lopez G, Contreras J, Murillo-Cuesta S, Rodriguez-Baeza A, Varela-Nieto I, Ruberte J, Collado M, et al. Programmed cell senescence during mammalian embryonic development. Cell. 2013;155:1104–1118. doi: 10.1016/j.cell.2013.10.019. [DOI] [PubMed] [Google Scholar]
- Naito AT, Sumida T, Nomura S, Liu ML, Higo T, Nakagawa A, Okada K, Sakai T, Hashimoto A, Hara Y, et al. Complement C1q activates canonical Wnt signaling and promotes aging-related phenotypes. Cell. 2012;149:1298–1313. doi: 10.1016/j.cell.2012.03.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura T, Sakata R, Ueno T, Sata M, Ueno H. Inhibition of transforming growth factor beta prevents progression of liver fibrosis and enhances hepatocyte regeneration in dimethylnitrosamine-treated rats. Hepatology. 2000;32:247–255. doi: 10.1053/jhep.2000.9109. [DOI] [PubMed] [Google Scholar]
- Nelson G, Wordsworth J, Wang C, Jurk D, Lawless C, Martin-Ruiz C, von Zglinicki T. A senescent cell bystander effect: senescence-induced senescence. Aging cell. 2012;11:345–349. doi: 10.1111/j.1474-9726.2012.00795.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neves J, Demaria M, Campisi J, Jasper H. Of flies, mice, and men: evolutionarily conserved tissue damage responses and aging. Developmental cell. 2015;32:9–18. doi: 10.1016/j.devcel.2014.11.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neves J, Zhu J, Sousa-Victor P, Konjikusic M, Riley R, Chew S, Qi Y, Jasper H, Lamba DA. Immune modulation by MANF promotes tissue repair and regenerative success in the retina. Science. 2016;353:aaf3646. doi: 10.1126/science.aaf3646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nosbaum A, Prevel N, Truong HA, Mehta P, Ettinger M, Scharschmidt TC, Ali NH, Pauli ML, Abbas AK, Rosenblum MD. Cutting Edge: Regulatory T Cells Facilitate Cutaneous Wound Healing. J Immunol. 2016;196:2010–2014. doi: 10.4049/jimmunol.1502139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowell CS, Odermatt PD, Azzolin L, Hohnel S, Wagner EF, Fantner GE, Lutolf MP, Barrandon Y, Piccolo S, Radtke F. Chronic inflammation imposes aberrant cell fate in regenerating epithelia through mechanotransduction. Nature cell biology. 2016;18:168–180. doi: 10.1038/ncb3290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ocampo A, Reddy P, Martinez-Redondo P, Platero-Luengo A, Hatanaka F, Hishida T, Li M, Lam D, Kurita M, Beyret E, et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell. 2016;167:1719–1733 e1712. doi: 10.1016/j.cell.2016.11.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh J, Lee YD, Wagers AJ. Stem cell aging: mechanisms, regulators and therapeutic opportunities. Nature medicine. 2014;20:870–880. doi: 10.1038/nm.3651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh J, Sinha I, Tan KY, Rosner B, Dreyfuss JM, Gjata O, Tran P, Shoelson SE, Wagers AJ. Age-associated NF-kappaB signaling in myofibers alters the satellite cell niche and re-strains muscle stem cell function. Aging. 2016;8:2871–2896. doi: 10.18632/aging.101098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okamoto M, Inoue K, Iwamura H, Terashima K, Soya H, Asashima M, Kuwabara T. Reduction in paracrine Wnt3 factors during aging causes impaired adult neurogenesis. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2011;25:3570–3582. doi: 10.1096/fj.11-184697. [DOI] [PubMed] [Google Scholar]
- Osorio FG, Barcena C, Soria-Valles C, Ramsay AJ, de Carlos F, Cobo J, Fueyo A, Freije JM, Lopez-Otin C. Nuclear lamina defects cause ATM-dependent NF-kappaB activation and link accelerated aging to a systemic inflammatory response. Genes & development. 2012;26:2311–2324. doi: 10.1101/gad.197954.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perdiguero E, Sousa-Victor P, Ruiz-Bonilla V, Jardi M, Caelles C, Serrano AL, Munoz-Canoves P. p38/MKP-1-regulated AKT coordinates macrophage transitions and resolution of inflammation during tissue repair. The Journal of cell biology. 2011;195:307–322. doi: 10.1083/jcb.201104053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pessina P, Kharraz Y, Jardi M, Fukada S, Serrano AL, Perdiguero E, Munoz-Canoves P. Fibrogenic Cell Plasticity Blunts Tissue Regeneration and Aggravates Muscular Dystrophy. Stem cell reports. 2015;4:1046–1060. doi: 10.1016/j.stemcr.2015.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillip JM, Aifuwa I, Walston J, Wirtz D. The Mechanobiology of Aging. Annual review of biomedical engineering. 2015;17:113–141. doi: 10.1146/annurev-bioeng-071114-040829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pietras EM, Mirantes-Barbeito C, Fong S, Loeffler D, Kovtonyuk LV, Zhang S, Lakshminarasimhan R, Chin CP, Techner JM, Will B, et al. Chronic interleukin-1 exposure drives haematopoietic stem cells towards precocious myeloid differentiation at the expense of self-renewal. Nature cell biology. 2016;18:607–618. doi: 10.1038/ncb3346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pineda JR, Daynac M, Chicheportiche A, Cebrian-Silla A, Sii Felice K, Garcia-Verdugo JM, Boussin FD, Mouthon MA. Vascular-derived TGF-beta increases in the stem cell niche and perturbs neurogenesis during aging and following irradiation in the adult mouse brain. EMBO molecular medicine. 2013;5:548–562. doi: 10.1002/emmm.201202197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poggioli T, Vujic A, Yang P, Macias-Trevino C, Uygur A, Loffredo FS, Pancoast JR, Cho M, Goldstein J, Tandias RM, et al. Circulating Growth Differentiation Factor 11/8 Levels Decline With Age. Circulation research. 2016;118:29–37. doi: 10.1161/CIRCRESAHA.115.307521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price FD, von Maltzahn J, Bentzinger CF, Dumont NA, Yin H, Chang NC, Wilson DH, Frenette J, Rudnicki MA. Inhibition of JAK-STAT signaling stimulates adult satellite cell function. Nature medicine. 2014;20:1174–1181. doi: 10.1038/nm.3655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pull SL, Doherty JM, Mills JC, Gordon JI, Stappenbeck TS. Activated macrophages are an adaptive element of the colonic epithelial progenitor niche necessary for regenerative responses to injury. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:99–104. doi: 10.1073/pnas.0405979102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qian F, Wang X, Zhang L, Chen S, Piecychna M, Allore H, Bockenstedt L, Malawista S, Bucala R, Shaw AC, et al. Age-associated elevation in TLR5 leads to increased inflammatory responses in the elderly. Aging cell. 2012;11:104–110. doi: 10.1111/j.1474-9726.2011.00759.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quarta M, Brett JO, DiMarco R, De Morree A, Boutet SC, Chacon R, Gibbons MC, Garcia VA, Su J, Shrager JB, et al. An artificial niche preserves the quiescence of muscle stem cells and enhances their therapeutic efficacy. Nature biotechnology. 2016 doi: 10.1038/nbt.3576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rando TA, Chang HY. Aging, rejuvenation, and epigenetic reprogramming: resetting the aging clock. Cell. 2012;148:46–57. doi: 10.1016/j.cell.2012.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raposo C, Graubardt N, Cohen M, Eitan C, London A, Berkutzki T, Schwartz M. CNS repair requires both effector and regulatory T cells with distinct temporal and spatial profiles. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2014;34:10141–10155. doi: 10.1523/JNEUROSCI.0076-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rebo J, Mehdipour M, Gathwala R, Causey K, Liu Y, Conboy MJ, Conboy IM. A single heterochronic blood exchange reveals rapid inhibition of multiple tissues by old blood. Nature communications. 2016;7:13363. doi: 10.1038/ncomms13363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rezza A, Wang Z, Sennett R, Qiao W, Wang D, Heitman N, Mok KW, Clavel C, Yi R, Zandstra P, et al. Signaling Networks among Stem Cell Precursors, Transit-Amplifying Progenitors, and their Niche in Developing Hair Follicles. Cell reports. 2016;14:3001–3018. doi: 10.1016/j.celrep.2016.02.078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi DJ, Bryder D, Zahn JM, Ahlenius H, Sonu R, Wagers AJ, Weissman IL. Cell intrinsic alterations underlie hematopoietic stem cell aging. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:9194–9199. doi: 10.1073/pnas.0503280102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rozo M, Li L, Fan CM. Targeting beta1-integrin signaling enhances regeneration in aged and dystrophic muscle in mice. Nature medicine. 2016 doi: 10.1038/nm.4116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruckh JM, Zhao JW, Shadrach JL, van Wijngaarden P, Rao TN, Wagers AJ, Franklin RJ. Rejuvenation of regeneration in the aging central nervous system. Cell stem cell. 2012;10:96–103. doi: 10.1016/j.stem.2011.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruhland MK, Loza AJ, Capietto AH, Luo X, Knolhoff BL, Flanagan KC, Belt BA, Alspach E, Leahy K, Luo J, et al. Stromal senescence establishes an immunosuppressive microenvironment that drives tumorigenesis. Nature communications. 2016;7:11762. doi: 10.1038/ncomms11762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruiz S, Lopez-Contreras AJ, Gabut M, Marion RM, Gutierrez-Martinez P, Bua S, Ramirez O, Olalde I, Rodrigo-Perez S, Li H, et al. Limiting replication stress during somatic cell reprogramming reduces genomic instability in induced pluripotent stem cells. Nature communications. 2015;6:8036. doi: 10.1038/ncomms9036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sacco A, Puri PL. Regulation of Muscle Satellite Cell Function in Tissue Homeostasis and Aging. Cell stem cell. 2015;16:585–587. doi: 10.1016/j.stem.2015.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saclier M, Yacoub-Youssef H, Mackey AL, Arnold L, Ardjoune H, Magnan M, Sailhan F, Chelly J, Pavlath GK, Mounier R, et al. Differentially activated macrophages orchestrate myogenic precursor cell fate during human skeletal muscle regeneration. Stem Cells. 2013;31:384–396. doi: 10.1002/stem.1288. [DOI] [PubMed] [Google Scholar]
- Sadtler K, Estrellas K, Allen BW, Wolf MT, Fan H, Tam AJ, Patel CH, Luber BS, Wang H, Wagner KR, et al. Developing a pro-regenerative biomaterial scaffold microenvironment requires T helper 2 cells. Science. 2016;352:366–370. doi: 10.1126/science.aad9272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salpeter SJ, Khalaileh A, Weinberg-Corem N, Ziv O, Glaser B, Dor Y. Systemic regulation of the age-related decline of pancreatic beta-cell replication. Diabetes. 2013;62:2843–2848. doi: 10.2337/db13-0160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez Alvarado A, Yamanaka S. Rethinking differentiation: stem cells, regeneration, and plasticity. Cell. 2014;157:110–119. doi: 10.1016/j.cell.2014.02.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scadden DT. Nice neighborhood: emerging concepts of the stem cell niche. Cell. 2014;157:41–50. doi: 10.1016/j.cell.2014.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schafer MJ, Atkinson EJ, Vanderboom PM, Kotajarvi B, White TA, Moore MM, Bruce CJ, Greason KL, Suri RM, Khosla S, et al. Quantification of GDF11 and Myostatin in Human Aging and Cardiovascular Disease. Cell metabolism. 2016;23:1207–1215. doi: 10.1016/j.cmet.2016.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schenke-Layland K, Rofail F, Heydarkhan S, Gluck JM, Ingle NP, Angelis E, Choi CH, MacLellan WR, Beygui RE, Shemin RJ, et al. The use of three-dimensional nanostructures to instruct cells to produce extracellular matrix for regenerative medicine strategies. Biomaterials. 2009;30:4665–4675. doi: 10.1016/j.biomaterials.2009.05.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schultz MB, Sinclair DA. When stem cells grow old: phenotypes and mechanisms of stem cell aging. Development. 2016;143:3–14. doi: 10.1242/dev.130633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schworer S, Becker F, Feller C, Baig AH, Kober U, Henze H, Kraus JM, Xin B, Lechel A, Lipka DB, et al. Epigenetic stress responses induce muscle stem-cell ageing by Hoxa9 developmental signals. Nature. 2016;540:428–432. doi: 10.1038/nature20603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekiya S, Suzuki A. Direct conversion of mouse fibroblasts to hepatocyte-like cells by defined factors. Nature. 2011;475:390–393. doi: 10.1038/nature10263. [DOI] [PubMed] [Google Scholar]
- Shaw AC, Goldstein DR, Montgomery RR. Age-dependent dysregulation of innate immunity. Nature reviews Immunology. 2013;13:875–887. doi: 10.1038/nri3547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva-Vargas V, Maldonado-Soto AR, Mizrak D, Codega P, Doetsch F. Age-Dependent Niche Signals from the Choroid Plexus Regulate Adult Neural Stem Cells. Cell stem cell. 2016 doi: 10.1016/j.stem.2016.06.013. [DOI] [PubMed] [Google Scholar]
- Sinha M, Jang YC, Oh J, Khong D, Wu EY, Manohar R, Miller C, Regalado SG, Loffredo FS, Pancoast JR, et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science. 2014;344:649–652. doi: 10.1126/science.1251152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith LK, He Y, Park JS, Bieri G, Snethlage CE, Lin K, Gontier G, Wabl R, Plambeck KE, Udeochu J, et al. beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis. Nature medicine. 2015;21:932–937. doi: 10.1038/nm.3898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soria-Valles C, Osorio FG, Gutierrez-Fernandez A, De Los Angeles A, Bueno C, Menendez P, Martin-Subero JI, Daley GQ, Freije JM, Lopez-Otin C. NF-kappaB activation impairs somatic cell reprogramming in ageing. Nature cell biology. 2015;17:1004–1013. doi: 10.1038/ncb3207. [DOI] [PubMed] [Google Scholar]
- Sousa-Victor P, Garcia-Prat L, Serrano AL, Perdiguero E, Munoz-Canoves P. Muscle stem cell aging: regulation and rejuvenation. Trends in endocrinology and metabolism: TEM. 2015;26:287–296. doi: 10.1016/j.tem.2015.03.006. [DOI] [PubMed] [Google Scholar]
- Sousa-Victor P, Gutarra S, Garcia-Prat L, Rodriguez-Ubreva J, Ortet L, Ruiz-Bonilla V, Jardi M, Ballestar E, Gonzalez S, Serrano AL, et al. Geriatric muscle stem cells switch reversible quiescence into senescence. Nature. 2014;506:316–321. doi: 10.1038/nature13013. [DOI] [PubMed] [Google Scholar]
- Storer M, Mas A, Robert-Moreno A, Pecoraro M, Ortells MC, Di Giacomo V, Yosef R, Pilpel N, Krizhanovsky V, Sharpe J, et al. Senescence is a developmental mechanism that contributes to embryonic growth and patterning. Cell. 2013;155:1119–1130. doi: 10.1016/j.cell.2013.10.041. [DOI] [PubMed] [Google Scholar]
- Sun D, Luo M, Jeong M, Rodriguez B, Xia Z, Hannah R, Wang H, Le T, Faull KF, Chen R, et al. Epigenomic profiling of young and aged HSCs reveals concerted changes during aging that reinforce self-renewal. Cell stem cell. 2014;14:673–688. doi: 10.1016/j.stem.2014.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–872. doi: 10.1016/j.cell.2007.11.019. [DOI] [PubMed] [Google Scholar]
- Tapia N, Scholer HR. Molecular Obstacles to Clinical Translation of iPSCs. Cell stem cell. 2016;19:298–309. doi: 10.1016/j.stem.2016.06.017. [DOI] [PubMed] [Google Scholar]
- Tonkin J, Temmerman L, Sampson RD, Gallego-Colon E, Barberi L, Bilbao D, Schneider MD, Musaro A, Rosenthal N. Monocyte/Macrophage-derived IGF-1 Orchestrates Murine Skeletal Muscle Regeneration and Modulates Autocrine Polarization. Molecular therapy : the journal of the American Society of Gene Therapy. 2015;23:1189–1200. doi: 10.1038/mt.2015.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trounson A, McDonald C. Stem Cell Therapies in Clinical Trials: Progress and Challenges. Cell stem cell. 2015;17:11–22. doi: 10.1016/j.stem.2015.06.007. [DOI] [PubMed] [Google Scholar]
- van Deursen JM. The role of senescent cells in ageing. Nature. 2014;509:439–446. doi: 10.1038/nature13193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varga T, Mounier R, Horvath A, Cuvellier S, Dumont F, Poliska S, Ardjoune H, Juban G, Nagy L, Chazaud B. Highly Dynamic Transcriptional Signature of Distinct Macrophage Subsets during Sterile Inflammation, Resolution, and Tissue Repair. J Immunol. 2016;196:4771–4782. doi: 10.4049/jimmunol.1502490. [DOI] [PubMed] [Google Scholar]
- Veiga-Fernandes H, Mucida D. Neuro-Immune Interactions at Barrier Surfaces. Cell. 2016;165:801–811. doi: 10.1016/j.cell.2016.04.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vierbuchen T, Ostermeier A, Pang ZP, Kokubu Y, Sudhof TC, Wernig M. Direct conversion of fibroblasts to functional neurons by defined factors. Nature. 2010;463:1035–1041. doi: 10.1038/nature08797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villalta SA, Rosenthal W, Martinez L, Kaur A, Sparwasser T, Tidball JG, Margeta M, Spencer MJ, Bluestone JA. Regulatory T cells suppress muscle inflammation and injury in muscular dystrophy. Science translational medicine. 2014;6:258ra142. doi: 10.1126/scitranslmed.3009925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villeda SA, Luo J, Mosher KI, Zou B, Britschgi M, Bieri G, Stan TM, Fainberg N, Ding Z, Eggel A, et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature. 2011;477:90–94. doi: 10.1038/nature10357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villeda SA, Plambeck KE, Middeldorp J, Castellano JM, Mosher KI, Luo J, Smith LK, Bieri G, Lin K, Berdnik D, et al. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nature medicine. 2014;20:659–663. doi: 10.1038/nm.3569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagers AJ. The stem cell niche in regenerative medicine. Cell stem cell. 2012;10:362–369. doi: 10.1016/j.stem.2012.02.018. [DOI] [PubMed] [Google Scholar]
- Wan X, Hou LJ, Zhang LY, Huang WJ, Liu L, Zhang Q, Hu B, Chen W, Chen X, Cao CC. IKKalpha is involved in kidney recovery and regeneration of acute ischemia/reperfusion injury in mice through IL10-producing regulatory T cells. Disease models & mechanisms. 2015;8:733–742. doi: 10.1242/dmm.018200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang C, Jurk D, Maddick M, Nelson G, Martin-Ruiz C, von Zglinicki T. DNA damage response and cellular senescence in tissues of aging mice. Aging cell. 2009;8:311–323. doi: 10.1111/j.1474-9726.2009.00481.x. [DOI] [PubMed] [Google Scholar]
- Wang J, Lu X, Sakk V, Klein CA, Rudolph KL. Senescence and apoptosis block hematopoietic activation of quiescent hematopoietic stem cells with short telomeres. Blood. 2014;124:3237–3240. doi: 10.1182/blood-2014-04-568055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L, Yang L, Debidda M, Witte D, Zheng Y. Cdc42 GTPase-activating protein deficiency promotes genomic instability and premature aging-like phenotypes. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:1248–1253. doi: 10.1073/pnas.0609149104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watt FM, Huck WT. Role of the extracellular matrix in regulating stem cell fate. Nature reviews Molecular cell biology. 2013;14:467–473. doi: 10.1038/nrm3620. [DOI] [PubMed] [Google Scholar]
- Weirather J, Hofmann UD, Beyersdorf N, Ramos GC, Vogel B, Frey A, Ertl G, Kerkau T, Frantz S. Foxp3+ CD4+ T cells improve healing after myocardial infarction by modulating monocyte/macrophage differentiation. Circulation research. 2014;115:55–67. doi: 10.1161/CIRCRESAHA.115.303895. [DOI] [PubMed] [Google Scholar]
- West EL, Pearson RA, Duran Y, Gonzalez-Cordero A, MacLaren RE, Smith AJ, Sowden JC, Ali RR. Manipulation of the recipient retinal environment by ectopic expression of neurotrophic growth factors can improve transplanted photoreceptor integration and survival. Cell transplantation. 2012;21:871–887. doi: 10.3727/096368911X623871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood LK, Kayupov E, Gumucio JP, Mendias CL, Claflin DR, Brooks SV. Intrinsic stiffness of extracellular matrix increases with age in skeletal muscles of mice. J Appl Physiol. 2014;117:363–369. doi: 10.1152/japplphysiol.00256.2014. 1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu M, Tchkonia T, Ding H, Ogrodnik M, Lubbers ER, Pirtskhalava T, White TA, Johnson KO, Stout MB, Mezera V, et al. JAK inhibition alleviates the cellular senescence-associated secretory phenotype and frailty in old age. Proceedings of the National Academy of Sciences of the United States of America. 2015;112:E6301–6310. doi: 10.1073/pnas.1515386112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue W, Zender L, Miething C, Dickins RA, Hernando E, Krizhanovsky V, Cordon-Cardo C, Lowe SW. Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas. Nature. 2007;445:656–660. doi: 10.1038/nature05529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y, Jiao J, Gao R, Le R, Kou X, Zhao Y, Wang H, Gao S, Wang Y. Enhanced Rejuvenation in Induced Pluripotent Stem Cell-Derived Neurons Compared with Directly Converted Neurons from an Aged Mouse. Stem cells and development. 2015;24:2767–2777. doi: 10.1089/scd.2015.0137. [DOI] [PubMed] [Google Scholar]
- Yim EK, Pang SW, Leong KW. Synthetic nanostructures inducing differentiation of human mesenchymal stem cells into neuronal lineage. Experimental cell research. 2007;313:1820–1829. doi: 10.1016/j.yexcr.2007.02.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang DY, Pan Y, Zhang C, Yan BX, Yu SS, Wu DL, Shi MM, Shi K, Cai XX, Zhou SS, et al. Wnt/beta-catenin signaling induces the aging of mesenchymal stem cells through promoting the ROS production. Molecular and cellular biochemistry. 2013a;374:13–20. doi: 10.1007/s11010-012-1498-1. [DOI] [PubMed] [Google Scholar]
- Zhang DY, Wang HJ, Tan YZ. Wnt/beta-catenin signaling induces the aging of mesenchymal stem cells through the DNA damage response and the p53/p21 pathway. PloS one. 2011;6:e21397. doi: 10.1371/journal.pone.0021397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang G, Li J, Purkayastha S, Tang Y, Zhang H, Yin Y, Li B, Liu G, Cai D. Hypothalamic programming of systemic ageing involving IKK-beta, NF-kappaB and GnRH. Nature. 2013b;497:211–216. doi: 10.1038/nature12143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W, Li J, Suzuki K, Qu J, Wang P, Zhou J, Liu X, Ren R, Xu X, Ocampo A, et al. Aging stem cells. A Werner syndrome stem cell model unveils heterochromatin alterations as a driver of human aging. Science. 2015;348:1160–1163. doi: 10.1126/science.aaa1356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Li Q, Liu D, Huang Q, Cai G, Cui S, Sun X, Chen X. GDF11 improves tubular regeneration after acute kidney injury in elderly mice. Scientific reports. 2016;6:34624. doi: 10.1038/srep34624. [DOI] [PMC free article] [PubMed] [Google Scholar]