

Abstract
Autophagy is a conserved process in which cytoplasmic components are sequestered for degradation in the vacuole/lysosomes in eukaryotic cells. Autophagy is induced under a variety of starvation conditions, such as the depletion of nitrogen, carbon, phosphorus, zinc, and others. However, apart from nitrogen starvation, it remains unclear how these stimuli induce autophagy. In yeast, for example, it remains contentious whether autophagy is induced under carbon starvation conditions, with reports variously suggesting both induction and lack of induction upon depletion of carbon. We therefore undertook an analysis to account for these inconsistencies, concluding that autophagy is induced in response to abrupt carbon starvation when cells are grown with glycerol but not glucose as the carbon source. We found that autophagy under these conditions is mediated by nonselective degradation that is highly dependent on the autophagosome-associated scaffold proteins Atg11 and Atg17. We also found that the extent of carbon starvation–induced autophagy is positively correlated with cells' oxygen consumption rate, drawing a link between autophagy induction and respiratory metabolism. Further biochemical analyses indicated that maintenance of intracellular ATP levels is also required for carbon starvation–induced autophagy and that autophagy plays an important role in cell viability during prolonged carbon starvation. Our findings suggest that carbon starvation–induced autophagy is negatively regulated by carbon catabolite repression.
Keywords: ATP, autophagy, energy metabolism, glucose, yeast
Introduction
Autophagy is a conserved process in which cytoplasmic components are sequestered for degradation in the vacuole/lysosomes in eukaryotes. During autophagy, a portion of the cytoplasm is sequestered into a double-membrane structure known as the autophagosome, which is subsequently delivered to the vacuole and degraded by a variety of vacuolar hydrolases. The resulting metabolites are recycled for use as inputs in cellular metabolism, including energy generation and the biosynthesis of proteins and membranes (1). Thus, defects in autophagy are known to cause aberrations in differentiation and development in yeast, nematodes, flies, and mice and have been implicated in a range of human diseases (2–4). In the yeast Saccharomyces cerevisiae, autophagy is particularly critical for the maintenance of cell viability during starvation, as well as the sporulation of cells (2, 5, 6). These observations underscore the fundamental yet incompletely characterized role of autophagy in cells.
Previous studies in yeast have identified ∼40 autophagy-related (Atg) proteins. Of these, 18 proteins are required for the formation of autophagosomes (Atg1–10, -12–14, -16–18, -29, and -31). A subset of “core” Atg proteins localizes to the preautophagosomal structure (PAS),2 from which the autophagosome is generated (7, 8). The concerted function of these core Atg proteins is essential for the progression of the two broad classes of autophagy that have been described: bulk and receptor-mediated autophagy. Bulk autophagy involves the sequestration and degradation of cytoplasmic components in response to starvation (9). In contrast, receptor-mediated autophagy refers to the marking by receptor proteins of specific targets, including aggregated proteins and damaged or over-abundant organelles, which are subsequently sequestered and degraded by the core autophagy machinery (10). The degradation of specific targets, such as mitochondria and peroxisomes, is collectively known as selective autophagy.
This diversity in function of the core autophagy machinery is afforded by scaffold proteins, which are employed by the cell to facilitate appropriate PAS formation and autophagy progression in response to cellular requirements. When bulk autophagy is induced, the Atg17–Atg29–Atg31 complex acts as an essential scaffold during autophagosome formation (11, 12). In contrast, cells require Atg11 as a scaffold protein to form the autophagosome during receptor-mediated autophagy (13, 14). In the latter case, Atg11 acts as an adapter protein, bringing cellular material marked for degradation by receptor proteins (including Atg19, Atg34, Atg32, Atg33, Atg36, Atg39, and Atg40) (15–19). The Atg17–Atg29–Atg31 complex and Atg11 thereby allow autophagy to act as a concerted degradation program within the cell. Thus, the deletion of Atg17 blocks bulk autophagy but not selective autophagy in cells (16). Meanwhile, the deletion of Atg11 results in the suppression of receptor-mediated autophagy but has only a marginal effect on the degradation of cytoplasmic components via bulk autophagy (20).
Carbon, which is essential for all living organisms, is the primary element of all classes of organic macromolecules, and its metabolism is central to the generation of energy (in the form of ATP). As a heterotroph, yeast assimilates a broad range of carbon sources. Of these, the monosaccharide glucose is the most preferred carbon source for S. cerevisiae and is metabolized by glycolysis without entering the tricarboxylic acid cycle, instead being diverted to ethanol production (alcoholic fermentation). The presence of a sufficient concentration of glucose results in the stringent repression of gene expression required for utilization of alternative carbon sources and respiration, a phenomenon known as catabolite repression. The presence of sufficient glucose therefore results in the generation of ATP almost exclusively from glycolysis rather than mitochondrial respiration, even when alternative carbon sources are available (21). In contrast, the absence of glucose results in the “derepression” of gene expression required for the catabolism of alternative sources such as glycerol, ethanol, acetate, and lactate and the acquisition of mitochondrial respiration (22).
The regulation of autophagy is closely linked to the metabolic state of the cell. The induction of autophagy has been described in response to a variety of nutrient starvation conditions, such as the depletion of nitrogen, carbon, sulfur, phosphorus, and zinc (9, 23, 24). Although the regulation and signaling responsible for nitrogen starvation–induced autophagy are relatively well-characterized, the mechanistic bases of autophagy induction in response to other starvation conditions are not well understood. A range of studies suggest that carbon starvation is able to induce autophagy (9, 25–28). However, a recent study contends that autophagy induction is blocked in cells subjected to carbon starvation and instead nutrients are replenished via endocytosis as a substitute for autophagy under these conditions (29). In addition, it has been shown that autophagy may be dispensable for cell survival during carbon starvation (29, 30). Thus, it remains contentious whether autophagy is induced under carbon starvation conditions. In this work, we investigate the effect of various growth conditions on carbon starvation–induced autophagy.
Results
Autophagy is induced in cells grown in glycerol medium but not in glucose medium in response to carbon starvation
In this study we employed an assay in which cells were initially grown on synthetic media, comprising a source of carbon, casamino acids, and ammonium sulfate, before being subjected to carbon starvation, whereby cells were transferred to the same medium lacking a source of carbon.
Using this approach, we examined cleavage of GFP-Atg8 and Ape1 maturation (Fig. 1A). Atg8 is recruited at the PAS and isolation membrane via lipidation during autophagosome formation. Following its incorporation into the autophagosomes, Atg8 is transported to the vacuole and degraded by vacuolar hydrolysis. Therefore, autophagy induction can be analyzed by detecting free GFP accumulating following cleavage of GFP-Atg8 (31). Ape1 is a cargo protein for the cytoplasm to vacuole targeting pathway and autophagy. Thus, Ape1 is transported to the vacuole via autophagy during starvation conditions, where it is converted from the proform (pApe1) to the mature form (mApe1) by vacuolar hydrolases (15).
Figure 1.
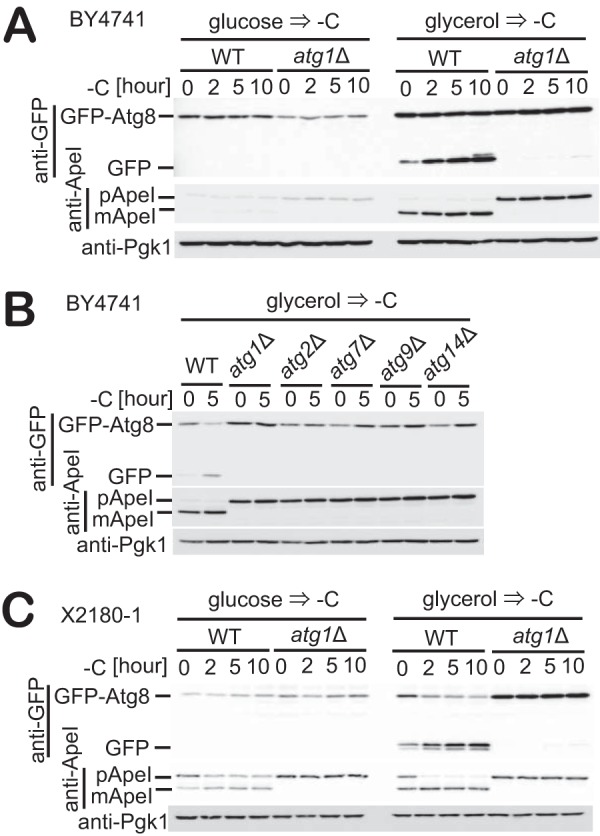
Autophagy is induced in glycerol-grown cells in response to carbon starvation. A, WT and atg1Δ cells expressing GFP-Atg8 grown in glucose or glycerol medium were subjected to carbon starvation. The cleavage of GFP-Atg8 and Ape1 maturation at the indicated time was assessed by immunoblotting with antibodies against GFP, Ape1, and Pgk1. Pgk1 was used as a loading control. B, WT, atg1Δ, atg2Δ, atg7Δ, atg9Δ, or atg14Δ cells expressing GFP-Atg8 grown in glycerol medium were subjected to carbon starvation. Immunoblotting was performed as described in A. C, WT or atg1Δ cells expressing GFP-Atg8 grown in glucose or glycerol medium were exposed to carbon starvation. Immunoblotting was performed as described in A.
In wild-type cells (BY4741) grown in glucose medium, we observed neither induction of autophagy nor any increase in Ape1 maturation when cells were subjected to carbon starvation (Fig. 1A, left). This result is consistent with previously reported data (29). We also observed that in cells subjected to carbon starvation, the expression of both GFP-Atg8 (under the control of its endogenous promotor) and Ape1 did not increase (Fig. 1A, left). However, autophagy was induced when cells grown in glycerol medium were subjected to carbon starvation (Fig. 1A, right). This autophagy was completely dependent on Atg1, suggesting the involvement of the canonical autophagy mechanism (Fig. 1A). We also observed that carbon starvation–induced autophagy requires the individual functions of Atg1, Atg2, Atg7, Atg9, and Atg14, which are representative for each functional unit of the core Atg machinery (Fig. 1B). In addition, cells grown in glycerol medium showed consistently high expression levels of both GFP-Atg8 and Ape1 compared with the expression levels of these proteins in glucose-grown cells (Fig. 1A, right).
We further tested whether this phenotype is reproduced in an alternative strain background. We also observed autophagy induction upon carbon starvation when prototrophic X2180-1 cells were grown in glycerol medium but not glucose medium (Fig. 1C), reproducing our observations in BY4741 cells (Fig. 1A). These results indicate that carbon starvation–induced autophagy following growth in the presence of glycerol but not glucose is not unique to BY4741 and is unrelated to the auxotrophy of the cells.
Behavior of Atg proteins under carbon starvation
Next, we examined the localization of Atg proteins by fluorescence microscopy when cells grown in glucose or glycerol medium were subjected to carbon starvation conditions. In glucose-grown cells, GFP-Atg8 displayed a diffuse cytosolic pattern under growing conditions (Fig. 2A). When subjected to carbon starvation, GFP fluorescence was not observed in the vacuole, rather accumulating as multiple puncta that are morphologically distinct from a typical PAS, even following 5 h of starvation (Fig. 2, A and B). Similarly, most other Atg proteins also accumulated to multiple punctate structures in the cytoplasm (Fig. 2B and supplemental Fig. S1). In contrast, cells grown in glycerol medium formed a single GFP-Atg8 dot structure per cell adjacent to the vacuole when subjected to carbon starvation, strongly suggesting PAS formation (Fig. 2A). The number of cells with this GFP-Atg8 dot structure increased in response to carbon starvation (Fig. 2A). Furthermore, this dot structure colocalized with Atg2-mCherry, Atg17-mCherry, and Atg11-mCherry (Fig. 2C). Together, these data indicate that cells grown in glycerol medium form a PAS that facilitates autophagy induction during carbon starvation, whereas PAS formation is defective in glucose-grown cells, resulting in a failure of autophagy induction during carbon starvation.
Figure 2.

Behavior of Atg proteins under carbon starvation. A, cells expressing GFP-Atg8 grown in glucose or glycerol medium were exposed to carbon starvation for 1 h before fluorescence images were acquired. The number of cells with the PAS (GFP-Atg8) was counted, and their percentages relative to the total numbers of cells are shown. B, cells expressing Atg2-GFP, GFP-Atg8, Atg9-GFP, Atg11-GFP, Atg14-GFP, or Atg17-GFP grown in glucose medium were exposed to carbon starvation for 5 h, and fluorescence images were then acquired. C, cells expressing GFP-Atg8 and Atg2-mCherry, GFP-Atg8 and Atg17-mCherry, Atg11-GFP and Atg17-mCherry, or Atg11-GFP and Atg2-mCherry were grown in glycerol medium and exposed to carbon starvation for 1 h before fluorescence microscopy. D, WT and atg1Δ cells expressing GFP-Atg8 grown in glycerol medium were exposed to carbon starvation. Then, fluorescence images were acquired every 20 s at 0.-μm step increments. The Z-slices were stacked. Scale bar, 5 μm.
We also observed the intracellular dynamics of GFP-Atg8 to examine the process from PAS formation to fusion of autophagosomes with vacuoles. Changes in GFP-Atg8 dot structure were observed periodically, with the dots becoming brighter, and the appearance of diffuse fluorescence within the vacuole was observed over the course of about 10 min (Fig. 2D; supplemental movie 1). In contrast, GFP-Atg8 dots observed in a strain lacking Atg1 moved in a random manner around the periphery of vacuole (Fig. 2D; supplemental movie 2). These results are consistent with descriptions of normal autophagy-associated GFP-Atg8 dynamics, as reported previously (7). Therefore, carbon starvation–induced autophagy appears to be facilitated by the canonical autophagy mechanism, as observed under nitrogen starvation conditions in glucose-grown cells.
Atg11 is required for autophagy induction in response to carbon starvation
Previous studies have reported the induction of mitophagy in cells grown in glycerol medium (16, 17). We therefore investigated whether mitophagy or any other selective autophagy is induced when cells grown in glycerol medium are subjected to carbon starvation. We first examined vacuolar cleavage of GFP-fusion proteins for substrates of several selective autophagic pathways (mitophagy, ERphagy, and pexophagy). The vacuolar cleavage of these GFP-fusion proteins was not observed in cells exposed to carbon starvation (supplemental Fig. S2A). In addition, the vacuolar degradation of lipid droplets was not observed (supplemental Fig. S2B). We also examined the cleavage of GFP-Atg8 in cells lacking each receptor protein required for various forms of selective autophagy. Autophagy progressed normally in cells lacking Atg19, Atg34, Atg32, Atg33, Atg36, Atg39, and Atg40 that were subjected to carbon starvation (supplemental Fig. S2C). Taken together, these results indicate that carbon starvation does not induce the established forms of selective autophagy.
In S. cerevisiae, the two major modes of autophagy, receptor-mediated and starvation–induced bulk autophagy, require the scaffold proteins Atg11 or Atg17, respectively. We next examined which of these scaffold proteins is required for carbon starvation–induced autophagy (Fig. 3A). As expected, under carbon starvation, both autophagy induction and Ape1 maturation were blocked in cells lacking Atg17 and the core Atg protein Atg1. Autophagy was not observed in cells deleted for both Atg11 and Atg17, which are completely defective in PAS formation. However, we also found that carbon starvation–induced cleavage of GFP-Atg8 is highly dependent on Atg11 (Fig. 3A), suggesting the involvement of this scaffold protein in addition to Atg17 during carbon starvation.
Figure 3.
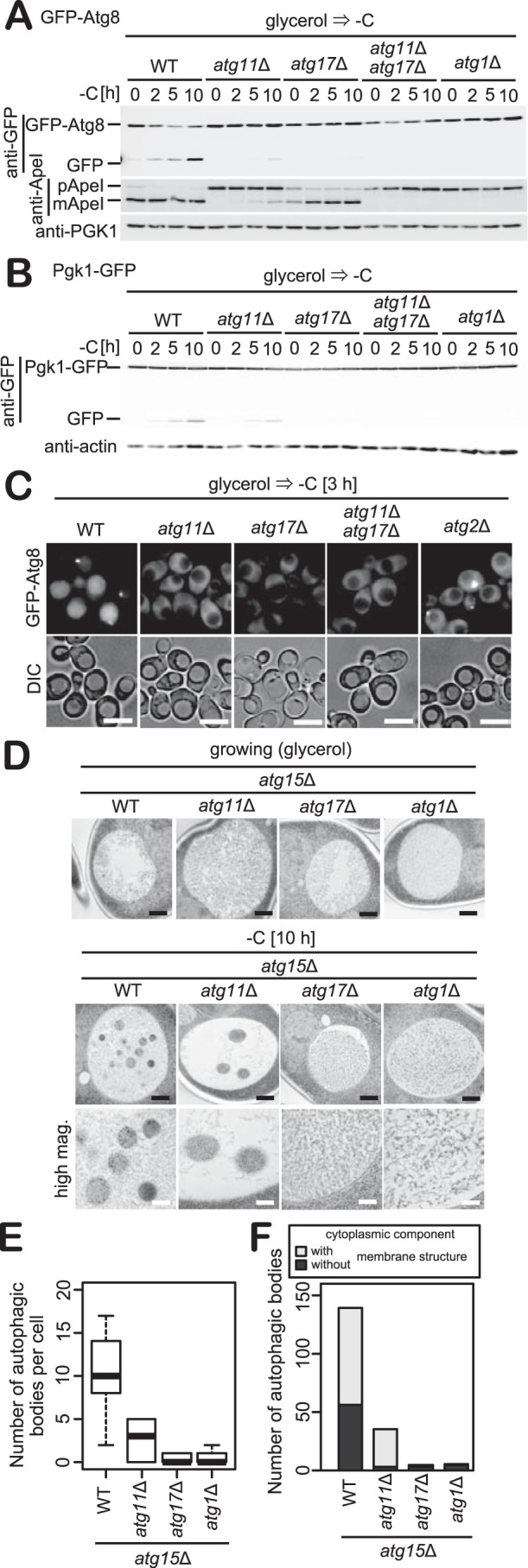
Carbon starvation–induced autophagy is highly dependent on Atg11. A and B, WT, atg11Δ, atg17Δ, atg11Δatg17Δ, or atg2Δcells expressing GFP-Atg8 or Pgk1-GFP grown in glycerol medium were exposed to carbon starvation. The cleavage of GFP-Atg8 or Pgk1-GFP and the Ape1 maturation were analyzed by immunoblotting with antibodies against GFP, Ape1, and Pgk1. Pgk1 or actin was used as a loading control. C, WT, atg11Δ, atg17Δ, atg11Δatg17Δ, or atg2Δ cells expressing GFP-Atg8 grown in glycerol medium were exposed to carbon starvation for 3 h before fluorescence microscopy. Scale bars, 5 μm. D, atg15Δ, atg15Δatg11Δ, and atg15Δatg1Δcells grown in glycerol medium were exposed to carbon starvation for 10 h. Images were acquired by transmission electron microscopy. Black bars, 500 nm; white bars, 200 nm. E, box plot of the number of autophagic bodies per cell in images obtained under the same conditions as shown in D. Whiskers indicate highest and lowest values, the top and bottom of the box represent upper and lower quartiles, respectively, and the central line indicates the median of data. n = 13 (WT, atg11Δ) or 12 (atg17Δ, atg1Δ). F, bars indicate total number of autophagic bodies containing cytoplasmic components with or without membrane structures in D.
We further examined the induction of bulk autophagy using Pgk1-GFP, an established reporter on the delivery of cytosolic components to the vacuole (32). We observed that the cleavage of Pgk1-GFP is highly dependent on Atg11, consistent with data demonstrating GFP-Atg8 cleavage (Fig. 3C). Microscopic examination of GFP-Atg8 further revealed that under carbon starvation conditions, the vacuolar fluorescence of GFP-Atg8 was abolished in strains deleted for Atg11, Atg17, Atg11 and Atg17, or Atg2 during the first 3 h of carbon starvation (Fig. 3C). Deletion of Atg11 resulted in a striking inhibition of PAS formation under these conditions, as for atg17Δ cells (Fig. 3C). From these analyses, we conclude that carbon starvation induces bulk autophagy that is highly dependent on Atg11.
Next, we observed autophagic bodies using transmission electron microscopy (TEM) (Fig. 3D). We used cells lacking Atg15, a lipase required for the degradation of autophagic bodies. Under growing conditions, spherical bodies were not observed in the vacuole of cells grown in glycerol medium. However, when subjected to carbon starvation, atg15Δ cells displayed a large number of spherical bodies in the vacuole. In atg15Δatg11Δ cells, the number of spherical bodies was dramatically reduced (Fig. 3E). In atg15Δatg17Δ and atg15Δatg2Δ cells, spherical bodies were not observed in the vacuole (Fig. 3E), consistent with our findings that Pgk1-GFP or GFP-Atg8 do not undergo cleavage in these strains, as shown in Fig. 3, A and B. TEM data further indicate that the morphological changes associated with microautophagy were rarely observed during carbon starvation (Fig. 3D). Taken together, these results indicate that carbon starvation–induced autophagy is facilitated by macroautophagy rather than microautophagy or endocytosis (supplemental Fig. S3).
Further examination revealed that the size of autophagic bodies in atg15Δ cells subjected to carbon starvation was about 250 nm in diameter (Fig. 3D). This size is similar to that reported for the mitophagosome (200–300 nm) rather than the diameter (300–900 nm) of autophagic bodies observed in glucose-grown cells under conditions of nitrogen starvation (9, 16). Examination of the content of autophagic bodies in carbon-starved atg15Δ cells revealed that autophagic bodies contain cytoplasmic components and membrane structures of undetermined origin (Fig. 3F). In contrast, in atg15Δatg11Δ cells, membrane structures were not observed in autophagic bodies (Fig. 3F), although the number of autophagic bodies was drastically reduced (Fig. 3E). These results suggest that cells may use membrane structures as an initiation site from which the autophagosome originates during carbon starvation–induced autophagy.
De-repression of catabolite repression is required for autophagy induction in response to carbon starvation
We examined autophagy induction in cells grown in a range of preculture conditions comprising various fermentable (glucose, raffinose, and galactose) and non-fermentable (ethanol, acetate, lactate, and glycerol) carbon sources. As shown in Fig. 1, autophagy was not induced in glucose-grown cells during carbon starvation (Fig. 4A). However, autophagy was observed when cells grown in raffinose or galactose were subjected to carbon starvation (Fig. 4A). Growth of cells in any of the non-fermentable carbon sources resulted in the induction of autophagy upon the onset of carbon starvation (Fig. 4A).
Figure 4.

Extent of autophagy in response to carbon starvation is highly correlated with oxygen consumption rate. A, WT and atg2Δ cells expressing GFP-Atg8 grown in the presence of indicated carbon sources were exposed to carbon starvation for 5 h. The cleavages of GFP-Atg8 and Ape1 maturation were analyzed by immunoblotting with antibodies against GFP, Ape1, and Pgk1. Pgk1 was used as a loading control. B, oxygen consumption rate in cells grown in indicated carbon sources was measured. A correlation between oxygen consumption and autophagy activity (upper) or Atg8 protein level (lower) is shown. Extent of autophagy was shown as the ratios of free GFP band intensities and total GFP ones (GFP-Atg8 and free GFP) acquired from A. Error bars denote S.D. C, WT, hxk1Δhxk2Δ. or atg2Δ cells expressing GFP-Atg8 grown in glucose were exposed to carbon starvation for 5 h. Immunoblotting was performed as described in A. D, oxygen consumption rate in WT or hxk1Δhxk2Δ cells grown in glucose medium was measured. Error bars denote S.D. **, p < 0.05 by Student's t test. E, WT and atg2Δ cells expressing GFP-Atg8 grown in glucose at the indicated cell densities were exposed to carbon starvation for 5 h. Immunoblotting was performed as described in A. F, WT, Snf1-AID, or atg2Δ cells expressing GFP-Atg8 grown in glycerol medium were treated with IAA for 30 min before being subjected to carbon starvation for 5 h. Immunoblotting was performed as described in A. G, oxygen consumption rate in WT or reg1Δ cells grown in glucose medium was measured. Error bars denote S.D. **, p < 0.05 by Student's t test. H, WT, reg1Δ, snf1Δ, or atg1Δ cells expressing GFP-Atg8 grown in glucose medium were exposed to carbon starvation for 5 h. Immunoblotting was performed as described in A.
When cells are grown in the presence of glucose, the expression of a broad array of genes involved in the utilization of carbon sources other than glucose and respiration is strictly repressed, a phenomenon referred to as catabolite repression (21). Thus, we monitored oxygen consumption rates in cells grown in medium containing different carbon sources. We observed that the extent of autophagy upon carbon starvation was positively correlated with the cellular oxygen consumption rate (Fig. 4B, upper panel). It was further observed that the expression of Atg8 also correlated with oxygen consumption (Fig. 4B, lower panel). These observations demonstrate a correlation between the degree of de-repression of catabolite repression and the cell's capacity to induce autophagy in response to carbon starvation.
To further investigate this probable link between catabolite repression and autophagy induction, we examined GFP-Atg8 cleavage in hxt1Δhxt2Δ cells, in which catabolite repression does not occur. In hxt1Δhxt2Δ cells, carbon starvation–induced autophagy was observed even in cells grown in glucose medium (Fig. 4C). The elevated oxygen consumption rate was also detected when hxt1Δhxt2Δ cells were grown on glucose medium (Fig. 4D). In yeast, the exhaustion of glucose leads to a metabolic reconfiguration known as the diauxic shift, whereby cells switch from the fermentation of glucose by glycolysis to a respiratory mode of growth utilizing ethanol (33). To determine whether this metabolic change is also associated with autophagy induction upon carbon starvation, we examined GFP-Atg8 cleavage in cells at a variety of growth phases. During the log phase of growth, when glucose is still available in media, autophagy was not induced following the onset of carbon starvation (Fig. 4E). This inhibition of autophagy persisted until just before entering diauxic shift (Fig. 4E, OD600 = 5.7). In contrast, our assessment of cells undergoing the diauxic shift revealed an elevation in oxygen consumption rate (supplemental Fig. S4A) and the induction of autophagy upon carbon starvation (Fig. 4E, OD600 = 6.5).
The expression of Atg8 was correlated with the oxygen consumption rate (Fig. 4B, lower panel). It is known that the expression level of Atg8 controls the size of autophagosomes and extent of autophagy under nitrogen starvation conditions (34). Thus, we examined the relationship between the level of GFP-Atg8 expression and autophagy induction. However, cells that show high expression levels of Atg8 lacking Ume6, Pho23, Rpd3, and Sin3 were not able to induce autophagy when subjected to carbon starvation (supplemental Fig. S4, B–D) (35, 36). In contrast, carbon starvation resulted in autophagy induction in cells in which an increase in both Atg8 expression and oxygen consumption rates are observed (supplemental Fig. S4, E and F) (37), indicating that high expression of Atg8 is not sufficient for autophagy induction in response to carbon starvation.
Snf1 activity is required for autophagy induction in response to carbon starvation
In the absence of catabolite repression, autophagy is induced in cells that have been subjected to carbon starvation, as shown in Fig. 4C. A central player in catabolite repression is Snf1, a kinase that is activated when the concentration of glucose drops below a certain threshold (21). We next examined whether Snf1 is required for autophagy induction when cells are subjected to carbon starvation. As Snf1 is essential for respiratory growth, we used the auxin-inducible degron (AID) system to conditionally knock down Snf1 expression. We first confirmed that levels of Snf1 in cells with AID-tagged Snf1 (Snf1-AID) were knocked down effectively when cells were treated with indole-3-acetic acid (IAA) (Fig. 4F). When subjected to carbon starvation, autophagy induction was blocked in cells expressing Snf1-AID when IAA was added to culture medium (Fig. 4F). Next, we examined autophagy induction in cells lacking Reg1. Because the Reg1–Glc7 complex negatively regulates Snf1 activity (21), reg1Δ cells are able to respire even in the presence of glucose (Fig. 4G). As for hxt1Δhxt2Δ cells, in reg1Δ cells, carbon starvation–induced autophagy even in cells grown in glucose medium (Fig. 4H), indicating that Snf1 activity is required for autophagy induction upon carbon starvation.
Activation of PKA and Torc1 suppresses autophagy induction in response to carbon starvation
cAMP-dependent protein kinase (PKA) and Torc1 negatively regulate autophagy induction (38–40). We therefore set out to determine whether these pathways are involved in the regulation of carbon starvation–induced autophagy. First, we monitored PKA activity (Fig. 5A) by assessing the phosphorylation of Cki1, a reporter of PKA activation. We used the first 200 amino acids of Cki1 containing mutations at two known protein kinase C sites (S25A and S30A) making its phosphorylation exclusively PKA-dependent (Ser-185) (41). In cells growing in the presence of glucose, the phosphorylation level of Cki1 was high (Fig. 5A, glucose). When these cells were subjected to carbon starvation, the phosphorylation dropped immediately (Fig. 5A, glucose). In contrast, in cells grown in glycerol medium, the degree of Cki1 phosphorylation was lower than that of cells grown in glucose medium (Fig. 5A, glycerol). This phosphorylation was unchanged in response to carbon starvation (Fig. 5A, glycerol). We next examined whether activation of PKA suppresses carbon starvation–induced autophagy. PKA activity is up-regulated by deletion of Bcy1, a protein that encodes the regulatory subunit of PKA (42). We used the AID system to conditionally knock down the expression of Bcy1 as Bcy1 is essential for growth on nonfermentable carbon sources. Cells expressing AID-tagged Bcy1 (AID-Bcy1) showed efficient knockdown of Bcy1 when treated with IAA (Fig. 5B). The level of AID-Bcy1 decreased upon carbon starvation (Fig. 5B). In cells grown in glycerol medium, the cleavage of GFP was reduced in cells expressing AID-Bcy1 that were subjected to carbon starvation, even in the absence of IAA. This indicated that AID tagging itself affects the function of Bcy1 to some degree. In addition, in cells grown in glucose, carbon starvation–induced autophagy observed in reg1Δ cells was blocked by knockdown of Bcy1 (Fig. 5C). Thus, these results indicate that activation of PKA suppresses carbon starvation–induced autophagy.
Figure 5.
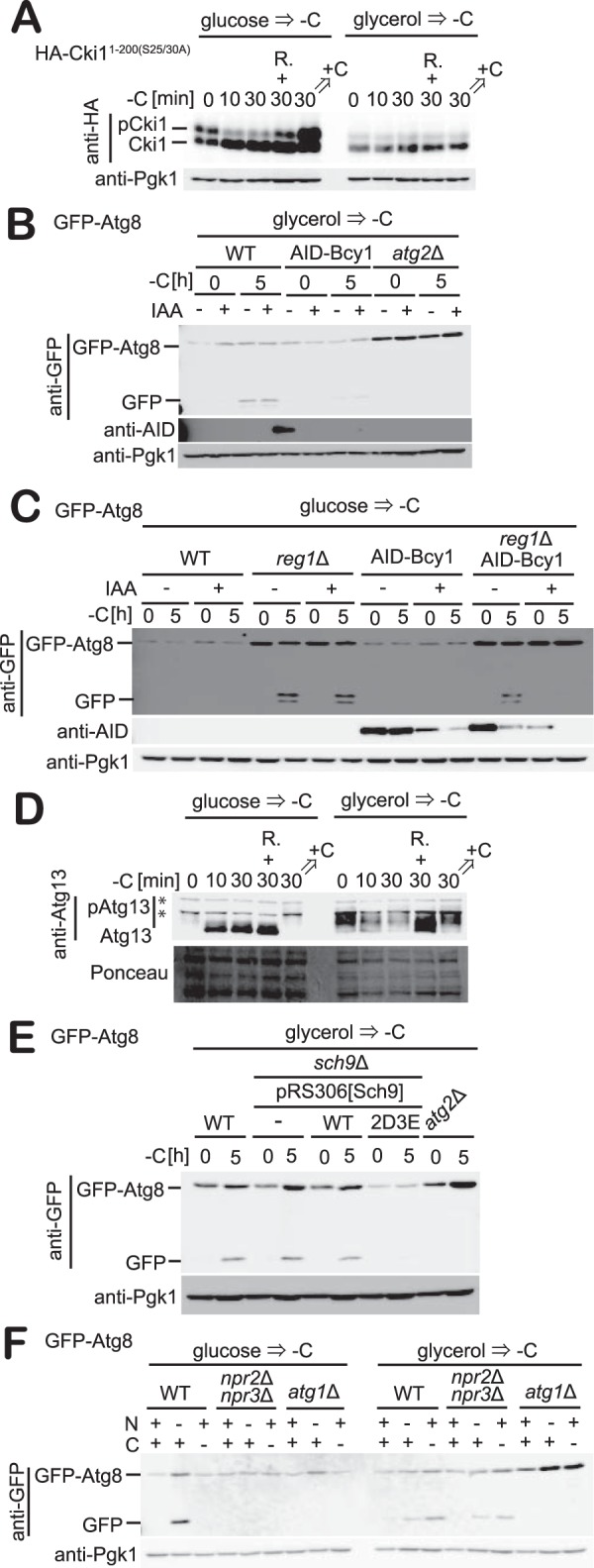
Activation of PKA or Torc1 suppresses carbon starvation–induced autophagy. A, cells expressing HA-Cki11–200(S25A/S30A) grown in glucose or glycerol medium were subjected to carbon starvation. Phosphorylation was analyzed by immunoblotting with antibodies against HA. R denotes addition of rapamycin. +C denotes addition of carbon source. B, WT, AID-Bcy1, or atg2Δ cells expressing GFP-Atg8 grown in glycerol medium were treated with IAA for 1 h and exposed to carbon starvation for 5 h. The cleavages of GFP-Atg8 at the indicated time were analyzed by immunoblotting with antibodies against GFP, Ape1, AID, and Pgk1. Pgk1 was used as a loading control. C, WT, reg1Δ, AID-Bcy1, or reg1ΔAID-Bcy1 cells expressing GFP-Atg8 grown in glucose medium were treated with IAA for 1 h and exposed to carbon starvation for 5 h. Immunoblotting was performed as described in B. D, cells grown in glucose or glycerol medium were exposed to carbon starvation. The phosphorylation status of Atg13 was analyzed by immunoblotting with antibodies against Atg13. R denotes addition of rapamycin. +C denotes addition of carbon source. E, WT, sch9Δ, atg2Δ, sch9Δ harboring plasmids for Sch9(WT) or Sch9(2D3E) cells expressing GFP-Atg8 grown in glycerol medium were exposed to carbon starvation for 5 h. Immunoblotting was performed as described in B. F, WT, npr2Δnpr3Δ, or atg1Δ cells expressing GFP-Atg8 grown in glucose or glycerol medium were exposed to carbon or nitrogen starvation for 5 h. Immunoblotting was performed as described in A.
Next, we monitored Torc1 activity (Fig. 5D). To monitor Torc1 activity, we examined the phosphorylation level of Atg13 (Fig. 5D), a substrate of Torc1 that is hyperphosphorylated under nutrient-rich conditions. When subjected to starvation conditions such as nitrogen starvation or rapamycin treatment, Atg13 is dephosphorylated and initiates the assembly of the PAS and consequently autophagosome formation (43, 44). In cells growing in the presence of glucose, Atg13 was extensively phosphorylated (Fig. 5D). This phosphorylation was drastically reduced when cells were subjected to carbon starvation, similarly to that of Cki1 (Fig. 5D). In contrast, Atg13 phosphorylation was limited in glycerol-grown cells maintained under nutrient-replete conditions, a phenotype that was unchanged when cells were subjected to carbon starvation (Fig. 5D).
We next examined whether activation of Torc1 suppresses carbon starvation–induced autophagy (Fig. 5E). Sch9 is a substrate of Torc1, and hyperactive mutants of Sch9 are able to suppress the induction of autophagy by rapamycin treatment (45, 46). We used a hyperactive mutant of Sch9 (Sch92D3E), in which five phosphorylation sites are substituted with acidic residues (T723D, S263D, T737E, S758E, and S765E). In cells grown in glycerol medium, expression of this hyperactive Sch9 mutant resulted in impaired expression of GFP-Atg8 and blocked autophagy induction when subjected to carbon starvation (Fig. 5E). These results indicate that activation of Torc1 suppresses autophagy induction upon carbon starvation. Furthermore, we also examined the effect of deletion of Npr2 and Npr3 on carbon starvation–induced autophagy (Fig. 5F). Npr2/3 are components of the SEA (Seh1-associated) complex, which transmits nitrogen or amino acids starvation signals to Torc1. Therefore, autophagy induction is suppressed in cells lacking Npr2/3 when cells grown in glucose medium are subjected to nitrogen or amino acid starvation (47, 48). However, the induction of carbon starvation–induced autophagy was unperturbed in npr2Δnpr3Δ cells grown in glycerol medium (Fig. 5F), indicating that the SEA complex is not required for carbon starvation–induced autophagy.
Intracellular ATP is required for the induction of autophagy during carbon starvation
As shown in Figs. 1 and 5, autophagy was not observed in glucose-grown cells subjected to carbon starvation, although the substrates of PKA and Torc1 were rapidly dephosphorylated under these conditions. We therefore focused on the metabolic status of cells upon carbon starvation. We monitored intracellular ATP levels in cells grown in glucose or glycerol medium when subjected to carbon starvation (Fig. 6A). In cells grown in glucose, the intracellular ATP level dropped immediately after carbon starvation (Fig. 6A). This decline persisted for at least 3 h following the onset of carbon starvation (Fig. 6A). In contrast, the ATP levels in cells grown in glycerol medium were maintained during carbon starvation (Fig. 6A). Oxygen consumption in cells grown in glycerol medium following carbon starvation was observed, but oxygen consumption was not apparent in glucose-grown cells (Fig. 6B). The intracellular ATP level and oxygen consumption rate phenotypes were also reproduced in reg1Δ cells (Fig. 6, C and D).
Figure 6.

Intracellular ATP level is correlated with autophagy induction in response to carbon starvation. A and B, WT cells grown in glucose or glycerol medium were exposed to carbon starvation. Intracellular ATP or oxygen consumption rates were measured. Error bars denote S.D. **, p < 0.05 by Student's t test (n = 3). C and D, WT or reg1Δ cells were grown in glucose medium and exposed to carbon starvation. Intracellular ATP or oxygen consumption rates were measured. Error bars denote S.D. **, p < 0.05; *, p < 0.5 by Student's t test (n = 3). E, WT and atg2Δ cells expressing GFP-Atg8 grown in glycerol medium were exposed to carbon starvation in the presence of antimycin A (5 μm) for 5 h. The cleavage of GFP-Atg8 was analyzed by immunoblotting with antibodies against GFP and Pgk1. Pgk1 was used as a loading control. F, cells grown in glycerol medium were exposed to carbon starvation with antimycin A (5 μm) for 3 h. Intracellular ATP level was measured. Error bars denote S.D. **, p < 0.05 by Student's t test (n = 3). G, WT and atg2Δ cells expressing GFP-Atg8 grown in glucose medium were shifted to medium containing indicated concentrations of glucose. The GFP-Atg8 and Ape1 maturation were analyzed by immunoblotting with antibodies against GFP, Ape1, and Pgk1. Pgk1 was used as a loading control. H, cells grown in glucose medium were shifted to medium containing 0.05% glucose. Intracellular ATP level was measured. Error bars denote S.D. **, p < 0.05 by Student's t test (n = 3).
We also examined the effect of antimycin A, a cytochrome c reductase inhibitor, on carbon starvation–induced autophagy (Fig. 6, E and F). Treatment with antimycin A resulted in a decrease in intracellular ATP levels in cells grown in glycerol medium following carbon starvation (Fig. 6F) and inhibited progression of autophagy under carbon starvation conditions (Fig. 6E). These results indicate that mitochondrial activity is required for the maintenance of intracellular ATP levels and autophagy induction during carbon starvation conditions. Finally, we examined the effect of a shift from media containing 2% glucose to media containing trace amounts of glucose on autophagy induction. When cells were shifted to medium containing 0.05% glucose, autophagy induction was observed from 1 h after this shift (Fig. 6G), whereas a shift to medium containing 0.2% glucose resulted in a delay in autophagy induction (from 3 h, Fig. 6G), suggesting that autophagy is induced upon the exhaustion of glucose in the medium. We also observed that such a shift to medium containing small amounts of glucose resulted in the maintenance of intracellular ATP levels (Fig. 6H). Taken together, these results indicate that the maintenance of intracellular ATP levels following carbon starvation is essential for autophagy induction.
Autophagy is required for the maintenance of cell viability during carbon starvation
We next examined the physiological relevance of autophagy in response to carbon starvation. In response to nitrogen starvation, cells enter a state in which the cell cycle slows and is ultimately arrested at G1/G0 phase (49, 50). To determine whether this phenomenon is relevant to carbon starvation–induced autophagy, we monitored cell growth during nitrogen or carbon starvation in cells grown in glucose or glycerol medium (Fig. 7A). When cells are grown in glucose medium, the onset of nitrogen starvation allows limited cell proliferation, whereas growth immediately ceases upon carbon starvation (Fig. 7A). In the case of glycerol-grown cells, nitrogen starvation supported only modest cell growth, whereas carbon starvation immediately prevented any further cell growth (Fig. 7A), indicating that autophagy is not at all able to support cell growth during carbon starvation.
Figure 7.
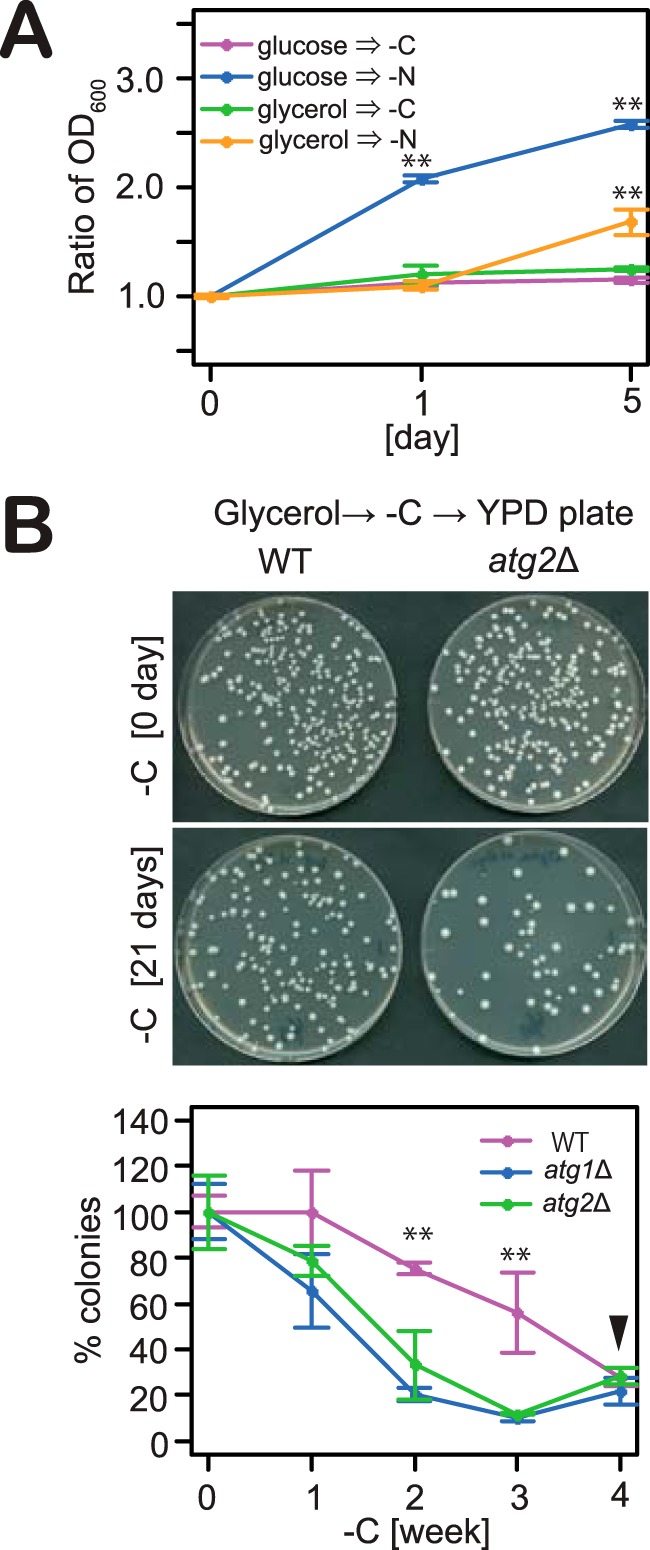
Autophagy induction is required for the maintenance of homeostasis during carbon starvation. A, cells grown in glucose or glycerol medium were exposed to nitrogen or carbon starvation, and A600 values were measured. Error bars denote S.D. **, p < 0.05 by Student's t test (n = 3). B, WT, atg1Δ, and atg2Δ cells grown in glycerol medium were exposed to carbon starvation. At indicated time cultures were diluted 1 × 105-fold and plated onto YPD agar. The plates were incubated at 30 °C for 4 days. The number of colonies on plates was counted. Error bars denote S.D. **, p < 0.05 by Student's t test (n = 3). Arrowhead indicates culture re-growth (“gasping”) (68, 69).
Studies have shown that glucose-grown cells defective in autophagy show a dramatic decrease in viability when subjected to nitrogen starvation due to the inadequate supply of autophagic flux-derived amino acids (2, 6, 51). In terms of carbon starvation, it has been reported that no change in viability is observed when glucose-grown cells defective in autophagy are subjected to carbon starvation (29). We therefore assessed the viability of cells grown in glycerol medium during carbon starvation (Fig. 7B), finding that the viability of wild-type cells reduced gradually over 4 weeks (Fig. 7B). In contrast, cells defective in autophagy (atg1Δ or atg2Δ cells) displayed significantly decreased viability after 3 weeks of carbon starvation. These results therefore indicate that autophagy plays an important role in the maintenance of cell viability during carbon starvation.
Discussion
In this study, we investigated how growth conditions affect carbon starvation–induced autophagy in S. cerevisiae. Our data indicate that autophagy is induced by carbon starvation when catabolite repression does not occur. Autophagy is not induced when cells grown to the log phase of growth in glucose are subjected to carbon starvation (Fig. 1A). In contrast, autophagy was observed upon carbon starvation of cells grown in non-fermentable carbon sources or in glucose-grown cells undergoing diauxic shift (Figs. 1 and 4E). These observations support previous reports that carbon starvation–induced autophagy is inhibited in cells grown to log phase on glucose and requires the Hsp31 minifamily of proteins, which is essential for the transition to diauxic shift (26, 29). Together, these findings suggest that autophagy induction in response to carbon starvation occurs only in the absence of catabolite repression. This finding complements our observation that the level of autophagy induction in response to carbon starvation is highly correlated with the cellular oxygen consumption rate, which is suppressed as a consequence of catabolite repression (Fig. 4, A and B). In addition, we found that autophagy induction in response to carbon starvation required the activation of Snf1, which is a key regulator of catabolite repression (Fig. 4, F–H). These results provide strong evidence that carbon starvation–induced autophagy requires derepression of catabolite repression.
As shown in Figs. 1 and 5, autophagy was not induced in cells grown in glucose medium upon carbon starvation, although the signaling requirements for autophagy induction appear to have been met. Thus, this inhibition of autophagy induction upon carbon starvation is likely due to differences in the metabolism of cells grown in fermentable and non-fermentable carbon sources. In particular, when cells are grown in glucose, intracellular ATP is exhausted rapidly when subjected to carbon starvation (Fig. 6A) (52–54). During autophagosome formation, cells require ATP for the conjugation of Atg8 and Atg12–Atg5, in addition to autophagy-related kinase activities (Atg1 and PI3K) (55). However, microscopic analysis revealed that in cells grown in glucose medium, the accumulation of scaffold proteins to the PAS was not observed upon carbon starvation (Fig. 2). This result suggests that ATP is required before PAS formation. Meanwhile, for cells in which catabolite repression does not occur, the intracellular ATP level is maintained following the onset of carbon starvation (Fig. 6, A and C) (53, 54). We therefore interpret these data as suggesting that cells require mitochondrial activity to maintain intracellular ATP levels during carbon starvation.
It has previously been reported that rapamycin treatment after carbon starvation leads to autophagy induction in cells grown in glucose medium (29, 56). However, in our experiments, rapamycin treatment after carbon starvation did not result in the induction of autophagy (data not shown). As shown in Fig. 5D, rapamycin did not enhance the dephosphorylation of Atg13 by carbon starvation. These findings indicate that treatment with rapamycin does not show any additional effect in carbon starvation–induced autophagy in cells grown in glucose medium.
A range of studies have demonstrated the induction of autophagy in glucose-grown cells at the log phase of growth in response to carbon starvation (23, 25, 27). Although we were unable to observe the induction of autophagy under these conditions (Fig. 1A), autophagy induction was apparent in cells shifted to media containing concentrations of glucose below the threshold required for catabolite repression (Fig. 6G) (57). We also observed that a shift to media containing glucose at these low concentrations resulted in the maintenance of intracellular ATP levels following the onset of carbon starvation (Fig. 6H). These results suggest that trace amounts of glucose within media, which results in a switch from fermentative to respiratory metabolism, allows the maintenance of intracellular ATP levels, thereby facilitating autophagy induction. We therefore suspect that the autophagy induction observed in the other studies noted above may be caused by residual glucose not completely eliminated from culture media, for example by extensive washing of cells. This explanation accounts for the reported discrepancies in autophagy induction under carbon starvation conditions.
Mammalian cells exhibit high respiratory activity and generate ATP via oxidative phosphorylation, even in the presence of glucose (58). The results presented in this study can therefore be reconciled with previous reports that autophagy is induced during carbon starvation conditions in mammalian cells (59). This autophagy induction upon carbon starvation was prevented by hypoxia but not normoxia (60). This finding tallies with our finding that treatment with antimycin A, which inhibits mitochondrial electron transport, prevents carbon starvation–induced autophagy (Fig. 6E). Thus, these data suggest that mammalian cells may require mitochondrial activity to induce autophagy in response to carbon starvation.
In S. cerevisiae, autophagy induction triggered by nitrogen starvation or treatment with rapamycin requires Atg17, which acts as the primary scaffold protein of the PAS but not Atg11, which is implicated in receptor-mediated autophagy. Here, we showed that autophagy in response to carbon starvation requires Atg11 as a scaffold protein for the PAS (Fig. 3). In yeast strains other than S. cerevisiae, Atg11 is required for bulk autophagy. For example, the thermotolerant yeast Kluyveromyces marxianus requires Atg11 for autophagy induced by rapamycin treatment (61), and the fission yeast Schizosaccharomyces pombe employs Atg11 as an essential component of autophagy induction upon nitrogen starvation (62). When S. cerevisiae is grown in the presence of glucose, cells convert glucose to ethanol even under aerobic conditions. During fermentation, cells acquire energy from glycolysis and fermentation but not oxidative respiration. This regulatory system is referred to as the Crabtree effect. In comparison with S. cerevisiae, the Crabtree effect is much less pronounced in K. marxianus and S. pombe (63). In our experiments, autophagy induction in response to carbon starvation was observed in respiring cells but not fermenting cells (Fig. 4). These findings therefore may suggest that Atg11 is involved in bulk autophagy induction when cells acquire energy by respiratory metabolism.
In this work, we showed that autophagy induction under carbon starvation is negatively correlated with catabolite repression. However, additional experiments are needed to further understand the relationship between autophagy induction and changes in the metabolic status of the cell in response to carbon starvation. Future studies in yeast will address these important questions in further detail.
Experimental procedures
Yeast strains and growth conditions
Yeast strains used in this study are listed in Table 1. Gene deletions and epitope tagging of genes at endogenous loci were constructed by PCR-based methods and were validated by PCR (64, 65). The strains for the AID-mediated deletion of proteins were constructed by adding the AID tag sequence to the C termini of Snf1 or the N termini of Bcy1 (66, 67). Cells were inoculated from plates into YPD medium and grown overnight. Cultures were then diluted in synthetic medium (0.17% yeast nitrogen base without amino acids and ammonium sulfate, 0.5% ammonium sulfate, 0.5% casamino acid, 0.002% tryptophan, 0.002% adenine, and 0.002% uracil) containing 2% glucose (glucose medium) or 2% glycerol and 0.1% glucose (glycerol medium) and grown until log phase. Following this, cultures were again diluted in glucose or glycerol medium and cultured overnight. Cells at A600 = 0.6 were then harvested and washed twice with distilled water before being transferred to carbon starvation medium (0.17% yeast nitrogen base without amino acids and ammonium sulfate, 0.5% ammonium sulfate, 0.5% casamino acids, 0.002% tryptophan, 0.002% adenine, and 0.002% uracil). 0.1% glucose was supplemented to glycerol media to accelerate the onset of growth. After overnight culture on glycerol medium, glucose was completely consumed, and cell growth is on glycerol. The same procedure was employed to preculture cells on other carbon sources, including galactose, raffinose, acetate, ethanol, and lactate. For rapamycin treatment, rapamycin (LC Laboratories) was added to a final concentration of 200 ng/ml, and the same volume of DMSO was added to control samples. For auxin treatment, IAA (Nacalai Tesque) was added to a final concentration of 0.5 mm, and the same volume of DMSO was added to control samples (66).
Table 1.
Yeast strains used in this study
| Strain name | Genotype |
|---|---|
| BY4741 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 |
| X2180-1 | MATα SUC2 mal mel gal2 CUP1 |
| ScAA824 | BY4741 GFP-ATG8-His3 |
| ScAA951 | BY4741 his3Δ1::GFP-ATG8-His3 atg2Δ::kanMX4 |
| ScAA1123 | BY4741 his3Δ1::GFP-ATG8-His3 atg1Δ::kanMX4 |
| ScAA991 | BY4741 his3Δ1::GFP-ATG8-His3 atg7Δ::kanMX4 |
| ScAA992 | BY4741 his3Δ1::GFP-ATG8-His3 atg9Δ::kanMX4 |
| ScAA993 | BY4741 his3Δ1:: GFP-ATG8-His3 atg14Δ::kanMX4 |
| ScAA890 | BY4741 his3Δ1::PGK1-GFP-His3 |
| ScAA987 | BY4741 his3Δ1::PGK1-GFP-His3 atg2Δ::kanMX4 |
| ScAA931 | X2180-1 atg8::GFP-ATG8-hphNT1 |
| ScAA956 | X2180-1 atg8::GFP-ATG8-hphNT1 atg1Δ::natNT2 |
| ScAA886 | BY4741 his3Δ1::GFP-ATG8-His3 atg2::ATG2-2mCherry-hphNT1 |
| ScAA1029 | BY4741 his3Δ1::GFP-ATG8-His3 atg17::ATG17-2mCherry-hphNT1 |
| ScAA925 | BY4741 atg11::ATG11-GFP-kanMX4 atg17::ATG17-2mCherry-hphNT1 |
| ScAA928 | BY4741 atg11::ATG11-GFP-kanMX4 atg2::ATG2-2mCherry-hphNT1 |
| ScAA891 | BY4741 his3Δ1::PGK1-GFP-His3 atg11Δ::kanMX4 |
| ScAA990 | BY4741 his3Δ1::PGK1-GFP-His3 atg17Δ::kanMX4 |
| ScAA947 | BY4741 his3Δ1::GFP-ATG8-His3 atg11Δ::kanMX4 |
| ScAA949 | BY4741 his3Δ1::GFP-ATG8-His3 atg17Δ::kanMX4 |
| ScAA979 | BY4741 his3Δ1::GFP-ATG8-His3 atg17Δ::kanMX4 atg11Δ::hphNT1 |
| ScAA915 | BY4741 his3Δ1::GFP-ATG8-His3 atg15Δ::natNT2 |
| ScAA916 | BY4741 his3Δ1::GFP-ATG8-His3 atg1Δ::hphNT1 atg15Δ::natNT2 |
| ScAA916 | BY4741 his3Δ1::GFP-ATG8-His3 atg11Δ::kanMX4 atg15Δ::natNT2 |
| ScAA1113 | BY4741 his3Δ1::GFP-ATG8-His3 atg15Δ::natNT2 atg17Δ::hphNT1 |
| ScAA1186 | BY4741 his3Δ1::GFP-ATG8-His3 hxk1Δ::natNT2 hxk2Δ::hphNT1 |
| ScAA1022 | BY4741 snf1::SNF1-3HA-hphNT1 |
| ScAA572 | BY4741 atg8:: GFP-ATG8::hphNT1 |
| ScAA1083 | BY4741 ura3Δ0::ADHpromoter-OsTIR-9myc-URA3 atg8:: GFP-ATG8::hphNT1 snf1::SNF1-AID-natNT2 |
| ScAA577 | BY4741 atg8:: GFP-ATG8::hphNT1 atg2Δ::kanMX4 |
| ScAA966 | BY4741 his3Δ1::GFP-ATG8-His3 reg1Δ::natNT2 |
| ScAA1023 | BY4741 his3Δ1::GFP-ATG8-His3 snf1Δ::kanMX4 |
| ScAA1050 | BY4741 CYCpromoter-3HA-Cki11–200 (S25/30A) |
| ScAA1030 | BY4741 ura3Δ0::ADHpromoter-OsTIR-9myc-URA3 atg8:: GFP-ATG8::hphNT1 bcy1::natNT2-ADHpromoter-AID-BCY1 |
| ScAA1106 | BY4741 atg8:: GFP-ATG8::hphNT1 reg1Δ::kanMX4 |
| ScAA1107 | BY4741 ura3Δ0::ADHpromoter-OsTIR-9myc-URA3 atg8:: GFP-ATG8::hphNT1 bcy1::natNT2-ADHpromoter-AID-BCY1 reg1Δ::kanMX4 |
| ScAA932 | BY4741 sch9::SCH9-3HA-hphNT1 |
| ScAA995 | BY4741 gln3::GLN3-3HA-hphNT1 |
| ScAA1038 | BY4741 his3Δ1::GFP-ATG8-His3 npr2Δ::kanMX4 npr3Δ::hphNT1 |
| ScAA1221 | BY4741 his3Δ1::GFP-ATG8-His3 sch9Δ::kanMX4 |
| ScAA958 | BY4741 his3Δ1::GFP-ATG8-His3 bcy1Δ::natNT2 |
| ScAA1001 | BY4741 atg1Δ::kanMX4 |
| ScAA816 | BY4741 atg2Δ::kanMX4 |
| ScAA1371 | BY4741 atg1::ATG1-GFP-kanMX4 |
| ScAA1372 | BY4741 atg2::ATG2-GFP-kanMX4 |
| ScAA1373 | BY4741 atg5::ATG5-GFP-kanMX4 |
| ScAA1374 | BY4741 atg6::ATG6-GFP-kanMX4 |
| ScAA1375 | BY4741 atg9::ATG9-GFP-kanMX4 |
| ScAA1376 | BY4741 atg11::ATG11-GFP-kanMX4 |
| ScAA1377 | BY4741 atg17::ATG17-GFP-kanMX4 |
| ScAA1084 | BY4741 his3Δ1::GFP-ATG8-His3 atg19Δ::kanMX4 |
| ScAA1085 | BY4741 his3Δ1::GFP-ATG8-His3 atg32Δ::kanMX4 |
| ScAA1086 | BY4741 his3Δ1::GFP-ATG8-His3 atg33Δ::kanMX4 |
| ScAA1087 | BY4741 his3Δ1::GFP-ATG8-His3 atg34Δ::kanMX4 |
| ScAA1088 | BY4741 his3Δ1::GFP-ATG8-His3 atg36Δ::kanMX4 |
| ScAA1090 | BY4741 his3Δ1::GFP-ATG8-His3 atg39Δ::kanMX4 |
| ScAA1091 | BY4741 his3Δ1::GFP-ATG8-His3 atg40Δ::kanMX4 |
| ScAA844 | BY4741 om45::OM45-GFP-hphNT1 |
| ScAA1044 | BY4741 om45::OM45-GFP-hphNT1 atg1Δ::kanMX4 |
| YKM841 | BY4741 ATG8 wt-T-P-kanMX PEX11-EGFP-natNT2 |
| YKM842 | BY4741 ATG8 wt-T-P-kanMX PEX11-EGFP-natNT2 atg1Δ::zeoNT3 |
| YKM936 | BY4741 sec63::SEC63-GFP-kanMX4 |
| YKM939 | BY4741 sec63::SEC63-GFP-kanMX4 atg1Δ::natNT2 |
| YKM206 | BY4741 ATG8 wt-T-P-kanMX ERG6-EGFP-hphNT1 |
| YKM226 | BY4741 ATG8 wt-T-P-kanMX ERG6-EGFP-hphNT1 atg1Δ::natNT2 |
| ScAA1125 | BY4741 his3Δ1::GFP-ATG8-His3 ume6Δ::natNT2 |
| ScAA850 | BY4741 vht1::VHT1-GFP-kanMX4 |
| ScAA883 | BY4741 vht1::VHT1-GFP-kanMX4 atg1Δ::natNT2 |
| ScAA885 | BY4741 vht1::VHT1-GFP-kanMX4 vps4Δ::natNT2 |
| ScAA862 | BY4741 his3Δ1::GFP-ATG8-His3 vps4Δ::kanMX4 |
| ScAA873 | BY4741 his3Δ1::GFP-ATG8-His3 vtc4Δ::kanMX4 |
Plasmid construction
All plasmids used in this study are listed in Table 2.
Table 2.
Plasmids used in this study
| Name | Description |
|---|---|
| pAA1 | pRS303-GFP-ATG8 |
| pKE47 | pRS303-PGK1-GFP |
| pAA2 | pRS306-SCH9 |
| pAA3 | pRS306-SCH92D3E (T723D, S263D, T737E, S758E, S765E) |
Immunoblotting
Yeast cell pellets were suspended in 100 μl of 10% trichloroacetic acid, kept on ice for 5 min, and centrifuged at 15,000 × g for 5 min. The pellets were washed with 1 ml of cold acetone, dried at room temperature, and suspended in SDS sample buffer followed by cell disruption at room temperature using FastPrep-24 (MP Biomedicals) and 0.5-mm YZB zirconia beads (Yasui Kikai). These samples were centrifuged at 15,000 × g for 5 min, incubated with agitation at 65 °C for 10 min, and centrifuged at 15,000 × g for 5 min. The supernatants were used for immunoblotting analysis. Antibodies against GFP (11814460001; Roche Applied Science), AID (BioROIS), Pgk1 (Invitrogen), HA (3F10; Roche Applied Science), Ape1 (laboratory stock), and Atg13 (43) were used for the detection of relevant proteins. Horseradish peroxidase-conjugated antibodies (Jackson ImmunoResearch) were used as secondary antibodies. Chemiluminescence signals produced by substrate for HRP enzyme (FEMTOGLOWPlus; Michigan Diagnostics) were detected using a CCD camera system (LAS4000; GE Healthcare).
Microscopic image acquisition
Fluorescence microscopy was performed at room temperature using an inverted fluorescence microscope (IX81; Olympus) equipped with an electron-multiplying CCD camera (ImagEM C9100-13) and a 150× objective lens (UAPON 150× OTIRF, NA/1.45; Olympus). GFP and mCherry were excited using a 488-nm blue laser (50 milliwatts; Coherent) and a 561-nm yellow laser (50 milliwatts; Coherent), respectively. Fluorescence was filtered with a Di01-R488/561-25 dichroic mirror (Semrock) and an Em01-R488/568-25 bandpass filter (Semrock) and separated into two channels using a U-SIP splitter (Olympus) equipped with a DM565HQ dichroic mirror (Olympus). The fluorescence was further filtered using an FF02-525/50-25 bandpass filter (Semrock) for the GFP channel and an FF01-624/40-25 bandpass filter (Semrock) for the mCherry channel. Images were acquired and processed using the MetaMorph software (Molecular Devices).
Oxygen consumption of cells
Respiration of yeast cells was measured noninvasively at room temperature using a Fibox3 oxygen meter (PreSence) with continuous stirring. Cells growing at the indicated growth phase were collected before dilution at A600 = 2.0 in phosphate buffer (0.1 m, pH 5.8) and monitoring of oxygen consumption.
Determination of cell viability
Cells initially grown in glycerol were exposed to carbon-starvation medium. At the indicated time points, cultures were diluted 1.0 × 105-fold and plated onto YPD agar plates. Plates were incubated at 30 °C for 4 days and colonies numbers counted.
ATP measurement
A600 = 1.0 of cells was washed, collected by centrifugation, and frozen using liquid nitrogen. The pellets were resuspended in 2.5% trichloroacetic acid and then diluted 1:50 with 25 mm Tris-HCl, pH 8.8. 20 μl of the samples was assayed using the ENLITEN® ATP assay kit (Promega). Luminescence signals were detected using a multimode microplate reader (Varioskan Lux; Thermo Fisher Scientific).
Author contributions
A. A. and Y. O. designed experiments and wrote the manuscript; A. A. and M. K. performed the experiments.
Supplementary Material
Acknowledgment
We thank Dr. Alexander May for proofreading.
This work was supported in part by Grants-in-aid for Scientific Research 23000015 (to Y. O.) and 16H06375 (to Y. O.) from the Ministry of Education, Culture, Sports, Science and Technology of Japan. The authors declare that they have no conflicts of interest with the contents of this article.
This article contains supplemental Figs. S1–S4 and Movies S1 and S2.
- PAS
- preautophagosomal structure
- AID
- auxin-inducible degron
- IAA
- indole-3-acetic acid
- TEM
- transmission electron microscopy.
References
- 1. Rabinowitz J. D., and White E. (2010) Autophagy and metabolism. Science 330, 1344–1348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Tsukada M., and Ohsumi Y. (1993) Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett. 333, 169–174 [DOI] [PubMed] [Google Scholar]
- 3. Mizushima N. (2005) The pleiotropic role of autophagy: from protein metabolism to bactericide. Cell Death Differ. 12, 1535–1541 [DOI] [PubMed] [Google Scholar]
- 4. Mizushima N., Levine B., Cuervo A. M., and Klionsky D. J. (2008) Autophagy fights disease through cellular self-digestion. Nature 451, 1069–1075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Onodera J., and Ohsumi Y. (2004) Ald6p is a preferred target for autophagy in yeast, Saccharomyces cerevisiae. J. Biol. Chem. 279, 16071–16076 [DOI] [PubMed] [Google Scholar]
- 6. Suzuki S. W., Onodera J., and Ohsumi Y. (2011) Starvation induced cell death in autophagy-defective yeast mutants is caused by mitochondria dysfunction. PLoS ONE 6, e17412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Suzuki K., Kirisako T., Kamada Y., Mizushima N., Noda T., and Ohsumi Y. (2001) The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation. EMBO J. 20, 5971–5981 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Suzuki K., Kubota Y., Sekito T., and Ohsumi Y. (2007) Hierarchy of Atg proteins in pre-autophagosomal structure organization. Genes Cells 12, 209–218 [DOI] [PubMed] [Google Scholar]
- 9. Takeshige K., Baba M., Tsuboi S., Noda T., and Ohsumi Y. (1992) Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. J. Cell Biol. 119, 301–311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Zaffagnini G., and Martens S. (2016) Mechanisms of selective autophagy. J. Mol. Biol. 428, 1714–1724 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Kawamata T., Kamada Y., Suzuki K., Kuboshima N., Akimatsu H., Ota S., Ohsumi M., and Ohsumi Y. (2005) Characterization of a novel autophagy-specific gene, ATG29. Biochem. Biophys. Res. Commun. 338, 1884–1889 [DOI] [PubMed] [Google Scholar]
- 12. Cao Y., and Klionsky D. J. (2007) Physiological functions of Atg6/Beclin 1: a unique autophagy-related protein. Cell Res. 17, 839–849 [DOI] [PubMed] [Google Scholar]
- 13. Kim J., Huang W. P., and Klionsky D. J. (2001) Membrane recruitment of Aut7p in the autophagy and cytoplasm to vacuole targeting pathways requires Aut1p, Aut2p, and the autophagy conjugation complex. J. Cell Biol. 152, 51–64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Shintani T., and Klionsky D. J. (2004) Cargo proteins facilitate the formation of transport vesicles in the cytoplasm to vacuole targeting pathway. J. Biol. Chem. 279, 29889–29894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Scott S. V., Guan J., Hutchins M. U., Kim J., and Klionsky D. J. (2001) Cvt19 is a receptor for the cytoplasm-to-vacuole targeting pathway. Mol. Cell 7, 1131–1141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Okamoto K., Kondo-Okamoto N., and Ohsumi Y. (2009) Mitochondria-anchored receptor Atg32 mediates degradation of mitochondria via selective autophagy. Dev. Cell 17, 87–97 [DOI] [PubMed] [Google Scholar]
- 17. Kanki T., Wang K., Cao Y., Baba M., and Klionsky D. J. (2009) Atg32 is a mitochondrial protein that confers selectivity during mitophagy. Dev. Cell 17, 98–109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Motley A. M., Nuttall J. M., and Hettema E. H. (2012) Pex3-anchored Atg36 tags peroxisomes for degradation in Saccharomyces cerevisiae. EMBO J. 31, 2852–2868 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Mochida K., Oikawa Y., Kimura Y., Kirisako H., Hirano H., Ohsumi Y., and Nakatogawa H. (2015) Receptor-mediated selective autophagy degrades the endoplasmic reticulum and the nucleus Nature. 522, 359–362 [DOI] [PubMed] [Google Scholar]
- 20. Mao K., Chew L. H., Inoue-Aono Y., Cheong H., Nair U., Popelka H., Yip C. K., and Klionsky D. J. (2013) Atg29 phosphorylation regulates coordination of the Atg17-Atg31-Atg29 complex with the Atg11 scaffold during autophagy initiation. Proc. Natl. Acad. Sci. U.S.A. 110, E2875–84 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Kayikci Ö., and Nielsen J. (2015) Glucose repression in Saccharomyces cerevisiae. FEMS Yeast Res. 15, fov068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Fendt S.-M., and Sauer U. (2010) Transcriptional regulation of respiration in yeast metabolizing differently repressive carbon substrates. BMC Syst. Biol. 4, 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Yokota H., Gomi K., and Shintani T. (2017) Induction of autophagy by phosphate starvation in an Atg11-dependent manner in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 483, 522–527 [DOI] [PubMed] [Google Scholar]
- 24. Kawamata T., Horie T., Matsunami M., Sasaki M., and Ohsumi Y. (2017) Zinc starvation induces autophagy in yeast. J. Biol. Chem. 292, 8520–8530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Noda T., Matsuura A., Wada Y., and Ohsumi Y. (1995) Novel system for monitoring autophagy in the yeast Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 210, 126–132 [DOI] [PubMed] [Google Scholar]
- 26. Miller-Fleming L., Antas P., Pais T. F., Smalley J. L., Giorgini F., and Outeiro T. F. (2014) Yeast DJ-1 superfamily members are required for diauxic-shift reprogramming and cell survival in stationary phase. Proc. Natl. Acad. Sci. U.S.A. 111, 7012–7017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Yi C., Tong J., Lu P., Wang Y., Zhang J., Sun C., Yuan K., Xue R., Zou B., Li N., Xiao S., Dai C., Huang Y., Xu L., Li L., Chen S., Miao D., Deng H., Li H., and Yu L. (2017) Formation of a Snf1-Mec1-Atg1 module on mitochondria governs energy deprivation-induced autophagy by regulating mitochondrial respiration. Dev. Cell 41, 59–71 [DOI] [PubMed] [Google Scholar]
- 28. Seo A. Y., Lau P.-W., Feliciano D., Sengupta P., Gros M. A., Cinquin B., Larabell C. A., and Lippincott-Schwartz J. (2017) AMPK and vacuole-associated Atg14p orchestrate μ-lipophagy for energy production and long-term survival under glucose starvation. Elife 6, e21690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Lang M. J., Martinez-Marquez J. Y., Prosser D. C., Ganser L. R., Buelto D., Wendland B., and Duncan M. C. (2014) Glucose starvation inhibits autophagy via vacuolar hydrolysis and induces plasma membrane internalization by down-regulating recycling. J. Biol. Chem. 289, 16736–16747 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Klosinska M. M., Crutchfield C. A., Bradley P. H., Rabinowitz J. D., and Broach J. R. (2011) Yeast cells can access distinct quiescent states. Genes Dev. 25, 336–349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Cheong H., and Klionsky D. J. (2008) Biochemical methods to monitor autophagy-related processes in yeast. Methods Enzymol. 451, 1–26 [DOI] [PubMed] [Google Scholar]
- 32. Welter E., Thumm M., and Krick R. (2010) Quantification of nonselective bulk autophagy in S. cerevisiae using Pgk1-GFP. Autophagy 6, 794–797 [DOI] [PubMed] [Google Scholar]
- 33. Turcotte B., Liang X. B., Robert F., and Soontorngun N. (2010) Transcriptional regulation of nonfermentable carbon utilization in budding yeast. FEMS Yeast Res. 10, 2–13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Xie Z., Nair U., and Klionsky D. J. (2008) Atg8 controls phagophore expansion during autophagosome formation. Mol. Biol. Cell. 19, 3290–3298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Bartholomew C. R., Suzuki T., Du Z., Backues S. K., Jin M., Lynch-Day M. A., Umekawa M., Kamath A., Zhao M., Xie Z., Inoki K., and Klionsky D. J. (2012) Ume6 transcription factor is part of a signaling cascade that regulates autophagy. Proc. Natl. Acad. Sci. U.S.A. 109, 11206–11210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Jin M., He D., Backues S. K., Freeberg M. A., Liu X., Kim J. K., and Klionsky D. J. (2014) Transcriptional regulation by Pho23 modulates the frequency of autophagosome formation. Curr. Biol. 24, 1314–1322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Bernard A., Jin M., González-Rodríguez P., Füllgrabe J., Delorme-Axford E., Backues S. K., Joseph B., and Klionsky D. J. (2015) Rph1/KDM4 mediates nutrient-limitation signaling that leads to the transcriptional induction of autophagy. Curr. Biol. 25, 546–555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Noda T., and Ohsumi Y. (1998) Tor, a phosphatidylinositol kinase homologue, controls autophagy in yeast. J. Biol. Chem. 273, 3963–3966 [DOI] [PubMed] [Google Scholar]
- 39. Budovskaya Y. V., Stephan J. S., Reggiori F., Klionsky D. J., and Herman P. K. (2004) The Ras/cAMP-dependent protein kinase signaling pathway regulates an early step of the autophagy process in Saccharomyces cerevisiae. J. Biol. Chem. 279, 20663–20671 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Stephan J. S., Yeh Y. Y., Ramachandran V., Deminoff S. J., and Herman P. K. (2010) The Tor and cAMP-dependent protein kinase signaling pathways coordinately control autophagy in Saccharomyces cerevisiae. Autophagy 6, 294–295 [DOI] [PubMed] [Google Scholar]
- 41. Deminoff S. J., Howard S. C., Hester A., Warner S., and Herman P. K. (2006) Using substrate-binding variants of the cAMP-dependent protein kinase to identify novel targets and a kinase domain important for substrate interactions in Saccharomyces cerevisiae. Genetics 173, 1909–1917 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Toda T., Cameron S., Sass P., Zoller M., Scott J. D., McMullen B., Hurwitz M., Krebs E. G., and Wigler M. (1987) Cloning and characterization of BCY1, a locus encoding a regulatory subunit of the cyclic AMP-dependent protein kinase in Saccharomyces cerevisiae. Mol. Cell. Biol. 7, 1371–1377 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Kamada Y., Funakoshi T., Shintani T., Nagano K., Ohsumi M., and Ohsumi Y. (2000) Tor-mediated induction of autophagy via an Apg1 protein kinase complex. J. Cell Biol. 150, 1507–1513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Fujioka Y., Suzuki S. W., Yamamoto H., Kondo-Kakuta C., Kimura Y., Hirano H., Akada R., Inagaki F., Ohsumi Y., and Noda N. N. (2014) Structural basis of starvation-induced assembly of the autophagy initiation complex. Nat. Struct. Mol. Biol. 21, 513–521 [DOI] [PubMed] [Google Scholar]
- 45. Urban J., Soulard A., Huber A., Lippman S., Mukhopadhyay D., Deloche O., Wanke V., Anrather D., Ammerer G., Riezman H., Broach J. R., De Virgilio C., Hall M. N., and Loewith R. (2007) Sch9 is a major target of TORC1 in Saccharomyces cerevisiae. Mol. Cell 26, 663–674 [DOI] [PubMed] [Google Scholar]
- 46. Yorimitsu T., Zaman S., Broach J. R., and Klionsky D. J. (2007) Protein kinase A and Sch9 cooperatively regulate induction of autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell. 18, 4180–4189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Graef M., and Nunnari J. (2011) Mitochondria regulate autophagy by conserved signalling pathways. EMBO J. 30, 2101–2114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Kira S., Tabata K., Shirahama-Noda K., Nozoe A., Yoshimori T., and Noda T. (2014) Reciprocal conversion of Gtr1 and Gtr2 nucleotide-binding states by Npr2-Npr3 inactivates TORC1 and induces autophagy. Autophagy 10, 1565–1578 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Matsui A., Kamada Y., and Matsuura A. (2013) The role of autophagy in genome stability through suppression of abnormal mitosis under starvation. PLoS Genet. 9, e1003245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. An Z., Tassa A., Thomas C., Zhong R., Xiao G., Fotedar R., Tu B. P., Klionsky D. J., and Levine B. (2014) Autophagy is required for G1/G0 quiescence in response to nitrogen starvation in Saccharomyces cerevisiae. Autophagy 10, 1702–1711 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Onodera J., and Ohsumi Y. (2005) Autophagy is required for maintenance of amino acid levels and protein synthesis under nitrogen starvation. J. Biol. Chem. 280, 31582–31586 [DOI] [PubMed] [Google Scholar]
- 52. Wilson W. A., Hawley S. A., and Hardie D. G. (1996) Glucose repression/derepression in budding yeast: SNF1 protein kinase is activated by phosphorylation under derepressing conditions, and this correlates with a high AMP:ATP ratio. Curr. Biol. 6, 1426–1434 [DOI] [PubMed] [Google Scholar]
- 53. Ashe M. P., De Long S. K., and Sachs A. B. (2000) Glucose depletion rapidly inhibits translation initiation in yeast. Mol. Biol. Cell. 11, 833–848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Aoh Q. L., Graves L. M., and Duncan M. C. (2011) Glucose regulates clathrin adaptors at the trans-Golgi network and endosomes. Mol. Biol. Cell. 22, 3671–3683 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Nakatogawa H., Suzuki K., Kamada Y., and Ohsumi Y. (2009) Dynamics and diversity in autophagy mechanisms: lessons from yeast. Nat. Rev. Mol. Cell Biol. 10, 458–467 [DOI] [PubMed] [Google Scholar]
- 56. Dechant R., Saad S., Ibáñez A. J., and Peter M. (2014) Cytosolic pH regulates cell growth through distinct GTPases, Arf1 and Gtr1, to promote ras/PKA and TORC1 activity. Mol. Cell 55, 409–421 [DOI] [PubMed] [Google Scholar]
- 57. Meijer M. M., Boonstra J., Verkleij A. J., and Verrips C. T. (1998) Glucose repression in Saccharomyces cerevisiae is related to the glucose concentration rather than the glucose flux. J. Biol. Chem. 273, 24102–24107 [DOI] [PubMed] [Google Scholar]
- 58. Pilatus U., Aboagye E., Artemov D., Mori N., Ackerstaff E., and Bhujwalla Z. M. (2001) Real-time measurements of cellular oxygen consumption, pH, and energy metabolism using nuclear magnetic resonance spectroscopy. Magn. Reson. Med. 45, 749–755 [DOI] [PubMed] [Google Scholar]
- 59. Kim J., Kim Y. C., Fang C., Russell R. C., Kim J. H., Fan W., Liu R., Zhong Q., and Guan K. L. (2013) Differential regulation of distinct Vps34 complexes by AMPK in nutrient stress and autophagy. Cell 152, 290–303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Sutter B. M., Wu X., Laxman S., and Tu B. P. (2013) Methionine inhibits autophagy and promotes growth by inducing the SAM-responsive methylation of PP2A. Cell 154, 403–415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Yamamoto H., Shima T., Yamaguchi M., Mochizuki Y., Hoshida H., Kakuta S., Kondo-Kakuta C., Noda N. N., Inagaki F., Itoh T., Akada R., and Ohsumi Y. (2015) The thermotolerant yeast Kluyveromyces marxianus is a useful organism for structural and biochemical studies of autophagy. J. Biol. Chem. 290, 29506–29518 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Sun L. L., Li M., Suo F., Liu X. M., Shen E. Z., Yang B., Dong M. Q., He W. Z., and Du L. L. (2013) Global analysis of fission yeast mating genes reveals new autophagy factors. PLoS Genet. 9, e1003715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Hagman A., Säll T., Compagno C., and Piskur J. (2013) Yeast “Make-Accumulate-Consume” life strategy evolved as a multi-step process that predates the whole genome duplication. PLoS ONE 8, e68734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Knop M., Siegers K., Pereira G., Zachariae W., Winsor B., Nasmyth K., and Schiebel E. (1999) Epitope tagging of yeast genes using a PCR-based strategy: more tags and improved practical routines. Yeast 15, 963–972 [DOI] [PubMed] [Google Scholar]
- 65. Janke C., Magiera M. M., Rathfelder N., Taxis C., Reber S., Maekawa H., Moreno-Borchart A., Doenges G., Schwob E., Schiebel E., and Knop M. (2004) A versatile toolbox for PCR-based tagging of yeast genes: new fluorescent proteins, more markers and promoter substitution cassettes. Yeast 21, 947–962 [DOI] [PubMed] [Google Scholar]
- 66. Nishimura K., Fukagawa T., Takisawa H., Kakimoto T., and Kanemaki M. (2009) An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nat. Methods 6, 917–922 [DOI] [PubMed] [Google Scholar]
- 67. Tanaka C., Tan L. J., Mochida K., Kirisako H., Koizumi M., Asai E., Sakoh-Nakatogawa M., Ohsumi Y., and Nakatogawa H. (2014) Hrr25 triggers selective autophagy-related pathways by phosphorylating receptor proteins. J. Cell Biol. 207, 91–105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Fabrizio P., Battistella L., Vardavas R., Gattazzo C., Liou L. L., Diaspro A., Dossen J. W., Gralla E. B., and Longo V. D. (2004) Superoxide is a mediator of an altruistic aging program in Saccharomyces cerevisiae. J. Cell Biol. 166, 1055–1067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Burtner C. R., Murakami C. J., Kennedy B. K., and Kaeberlein M. (2009) A molecular mechanism of chronological aging in yeast. Cell Cycle 8, 1256–1270 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.