

Abstract
The field of cognitive neuroscience was transformed by the discovery that exercise induces neurogenesis in the adult brain, with the potential to improve brain health and stave off the effects of neurodegenerative disease. However, the basic mechanisms underlying exercise-brain connections are not well understood. Here, we use an evolutionary-neuroscience approach to develop the Adaptive Capacity Model (ACM), detailing how and why physical activity improves brain function based on an energy-minimizing strategy. Building on studies showing a combined benefit of exercise and cognitive challenge to enhance neuroplasticity, our ACM addresses two fundamental questions: 1) what are the proximate and ultimate mechanisms underlying age-related brain atrophy? and 2) how do lifestyle changes influence the trajectory of healthy and pathological aging?
Over the last decade, numerous studies have shown a beneficial effect of aerobic exercise on cognition across the lifespan, with particularly important implications for the aging brain [1–4]. These positive exercise-induced brain changes have been associated with cognitive benefits supporting exercise as a promising lifestyle intervention, offering potential therapeutic possibilities for both healthy aging and for individuals suffering from neurodegenerative diseases, such as Alzheimer’s disease [5–7]. However, despite growing interest in the use of physical activity as a therapeutic intervention, the basic mechanisms underlying exercise-brain connections are not well understood and studies reveal a high degree of variation in the effects of exercise on the brain. One key limiting factor is that we lack a theoretical explanatory model describing the interactions between exercise, cognition, and the brain, leaving researchers without clearly defined predictions or explanations for the variation across studies (see Box 1). Understanding the source of the observed individual differences in response to exercise is particularly important for generating interventions that provide more consistent and universally effective outcomes across the lifespan.
BOX 1. Existing models of lifestyle and brain aging: Reserve, Scaffolding, and Maintenance.
Several models have emerged over the past two decades suggesting that differences in activities across the lifespan, including exercise participation, can help to explain individual differences in cognitive outcomes observed during aging and in the context of brain injury and neurodegenerative disease [104–109]. Reserve, neurocognitive scaffolding, and brain maintenance models generally suggest that a range of early and lifelong activities such as level of educational attainment, social engagement, and exercise can help build or maintain brain tissue or network flexibility that can either delay or compensate for structural brain changes that occur during aging or in response to developing disease pathology. While these models have proven extremely popular and valuable in addressing how individuals might differ in trajectories of cognitive decline, they do have some limitations in their application. For example, existing models do not explain why a specific type of behavior should lead to a lifelong build up of brain or neurocognitive reserve against developing pathology, reducing our ability to design interventions that specifically leverage their mechanistic origins to amplify or more consistently generate benefits. Additionally, existing models do not explicitly and consistently predict how particular behaviors will impact specific regional brain structures or networks.
Thus, despite their utility in explaining some of the variance in cognitive outcomes during aging, the limitations of existing models, with partially complementary and also overlapping generalized constructs, leaves us without a clear and specific direction for designing new studies and making precise public health recommendations. The Adaptive Capacity Model (ACM) is built on an evolutionary neuroscience approach to brain health that establishes a theoretical context for why one type of behavior, i.e., exercise, is especially beneficial to brain aging and cognition, and further generates explicit predictions for how this type of behavior impacts the brain across the lifespan, providing a clear direction for intervention research based on behaviorally-linked neuroplasticity. While we focus here on exercise, we expect that applying such an evolutionary neuroscience framework to other types of behaviors (e.g., educational attainment, social activity) that have been associated with altering the course of cognitive and brain aging has the potential to advance our understanding of the underlying proximate and ultimate mechanisms associated with other sources of individual differences observed during aging.
Here, we use an evolutionary neuroscience perspective to propose a new model detailing how and why physical activity influences human brain function. We suggest that exercise is linked to cognition because our evolutionary history included a shift to cognitively demanding foraging patterns that required combining elements of motor control, memory, spatial navigation, and executive functions with high levels of aerobic physical activity. Integrating concepts in evolutionary medicine with those from clinical and cognitive neuroscience, we propose a new Adaptive Capacity Model (ACM) to explain how and why exercise protects the brain across the lifespan, leading to enhanced cognitive functions and the potential for successful cognitive aging. The ACM suggests that our evolutionary history as cognitively engaged, “endurance athletes” during foraging led to enhanced neural responses to exercise. In addition, when faced with chronic inactivity over the lifespan common to modern industrialized societies, our brains adaptively reduce capacity as part of an energy-saving strategy, leading to age-related brain atrophy. By detailing specific cognitive domains that are linked to physically active search and decision-making (e.g., those required during foraging in natural settings), the ACM provides a blueprint for a novel research pathway that capitalizes on our evolved neurophysiology to enhance brain function across the lifespan. In addition, the ACM may help us understand why previous exercise, as well as, cognitive training interventions have highly variable results, since differences among studies in controlling for cognitive demands, types of cognitive tasks, and intensity and timing of concurrent physical activity could influence observed outcomes. Having a theoretical model, like the ACM, provides an important framework to support the development and evaluation of targeted brain-based interventions that use exercise combined with cognitive stimulation to generate more consistently beneficial outcomes in the context of healthy and pathological aging.
The Adaptive Capacity Model
The foundation for the ACM is the wealth of data that have shown that aerobic exercise can induce neuroplasticity in both non-human animal models and humans. In general, results from rodent studies have focused on the neuroplastic response to aerobic activity in the hippocampus. These highly controlled experiments are consistent, reliable, and show that aerobic exercise leads to the formation of new neurons (neurogenesis) within this brain structure, which has been associated with improved performance on cognitive tasks [1,8–12]. Exercise appears to trigger hippocampal neurogenesis primarily through the upregulation of neurotrophins and growth factors, with brain-derived neurotrophic factor (BDNF) playing a key role [2,10–15]. In addition, angiogenesis and enhanced cerebral perfusion may play an important role in exercise-induced structural brain benefits [10]. Studies in humans support both the effects of exercise on the brain and the hypothesized perfusion and BDNF-based molecular mechanisms, however there is more variation across human studies. For example, while cross-sectional and intervention studies suggest that aerobic fitness and exercise participation lead to improved memory and executive function, and enhanced brain structure [16–22], a recent meta-analysis highlights a high degree of variation across studies [23]. We suggest that an evolutionary neuroscience approach can help us understand why results differ among human studies and can lead to more consistent outcomes from future work.
An evolutionary neuroscience approach to brain health
Recently, the emerging field of evolutionary medicine has pioneered the use of evolutionary theory to explore the effects of lifestyle on health and well-being [24–27]. Researchers have suggested that human evolutionary history may be responsible for both the physiological benefits of a highly active lifestyle and the dangers of more sedentary living [24–26,28–31]. Beginning around two million years ago, human ancestors transitioned from a relatively sedentary ape-like existence to a hunting and gathering lifestyle which required an increased amount of physical activity compared to earlier hominins [25,26,28,30,32–39]. Human physiology evolved within the context of high levels of daily aerobic activity, and it is likely that our reliance on exercise for health maintenance is a legacy of our evolutionary past.
The links between health and physical activity rely on the ability of our physiological systems to adapt to stimuli. Much work has shown that physiological systems evolved to meet changes in demands with changes in capacity, and these alterations in capacity are constrained by energy costs [25,26,40]. In this case, capacity refers to the ability of physiological systems to perform required tasks, which can determine the health of specific organ systems and the ability to function in everyday activities [41]. Physiological capacity reductions in response to lack of stimuli are adaptive since they reduce metabolic requirements (e.g., basal metabolism), freeing up energy for other aspects of biology that improve reproductive success [26]. This interaction between energy availability and reproduction is well documented in humans and severe energy stress (e.g., caloric restriction or extremely high workloads), can lead to the disruption of reproduction, which is returned when energy availability increases [42].
Human physiology seems specifically adapted to respond to the aerobic exercise-induced stresses required of a hunting and gathering lifestyle, and when faced with a less active lifestyle common to many of today’s industrialized societies, organ systems undergo a reduction in capacity, often predisposing us to chronic disease [26]. Based on analyses of modern human hunting and gathering populations [39,43], it is likely that foraging occurs at moderate aerobic intensities (~40–85% of maximum aerobic capacity) over long time periods, and thus, we might expect that physiological adaptations are tuned to this range of intensities. Since humans never had to adapt to long periods of inactivity, including very low amounts of moderate intensity activity, our physiological response to long-term inactivity can lead to chronic disease [26], and this may be especially relevant in the context of cognitive and brain aging.
For example, in the face of chronically low levels of physical activity, the human cardiovascular system reduces peripheral vasculature and vascular elasticity, leading to heightened cardiovascular disease risk [26]. Because the primary function of the cardiovascular system (i.e., moving oxygen-containing blood to working muscles and organs) is taxed during exercise, chronic inactivity reduces this stimulus and elicits a detrimental response [26,34]. Capacity reductions are most clearly seen in analyses of individuals who undergo exercise training, followed by a period of detraining (inactivity) [44]. Periods of detraining lead to increased endothelial dysfunction [45], reductions in cardiac stroke volume, reductions in heart mass [46], and reductions in peripheral capillary density [47]. Other examples of harmful inactivity-induced capacity reductions include loss of muscle mass and strength (sarcopenia) [48] and loss of bone mineral density (osteoporosis) [26,34].
There is strong evidence that adaptive physiological capacity is a product of our evolutionary history as aerobic athletes. For example, a key requirement for the evolution of adaptive capacity is that the physiological response to exercise must be heritable. In humans, studies have found that the physiological response to exercise is, in fact, heritable, and that there is heterogeneity in individual response to exercise programs, including a small percentage of non-responders [49]. This heterogeneity in exercise-response is expected in traits that vary within populations and can therefore be acted upon by natural selection. Further support for the evolution of adaptive physiological capacity comes from artificial selection experiments in rodent models that act, in some ways, as analogs for the evolution of human endurance capabilities. In one experiment, mice were selectively bred for high amounts of voluntary wheel running [50], and in another, rats were selectively bred for high and low aerobic capacity [51]. In both mice and rats, organ plasticity in response to exercise is enhanced in selected lines compared with control lines that were bred randomly (liver and kidney mass in mice [50] and heart and lung mass in rats [51]). These experiments provide a novel window into the underlying evolutionary mechanisms behind adaptive physiological capacity, and, when viewed through the lens of human evolution, suggest that our physiology was strongly shaped by selection for aerobic activity.
Based on an energy-minimizing strategy inherent in the ACM, we would expect the human brain to be particularly susceptible to capacity reductions. The human brain has the highest mass-specific metabolic rate of any organ [52], and it’s total energy costs account for approximately 20% of human basal metabolism [53,54]. However, it is surprising that a lack of exercise might lead to capacity reductions in this organ system since physical activity is not generally considered a cognitively challenging behavior [55]. Here, we draw on data from animal model and human intervention studies to show that the stimulus for exercise-induced brain capacity maintenance or enhancement is likely related to the cognitive demands of aerobically active foraging. That is, links between exercise and the brain likely evolved to improve foraging success in our ancestors, and a lack of either exercise in general or cognitive demands during exercise may lead to capacity reductions or sub-optimal capacity maintenance in the brain similar to those seen in other organ systems.
Mechanisms underlying the Adaptive Capacity Model (ACM)
The ACM differs from prior models of lifestyle-brain interactions by starting with the premise that our behavioral set-point is lifelong physical activity (Figure 1). Rather than building-up capacity from a sedentary life, we suggest that the adult human body consistently responds to exercise-induced stimulation by maintaining brain capacity, and that a lack of stimulation is treated as an opportunity for energy savings through capacity reduction. Such reductions in capacity in the context of neurologically healthy aging are expressed as age-related brain atrophy. This concept also suggests that genetic risk factors for neurodegenerative diseases, such as Alzheimer’s disease, that were maintained throughout our evolutionary history (e.g., the Apolipoprotein E [APOE] ε4 allele [56]) may only lead to enhanced risk of cognitive impairment or vulnerability to dementia when met with a lack of exercise-induced stimulation (Figure 1).
Figure 1.
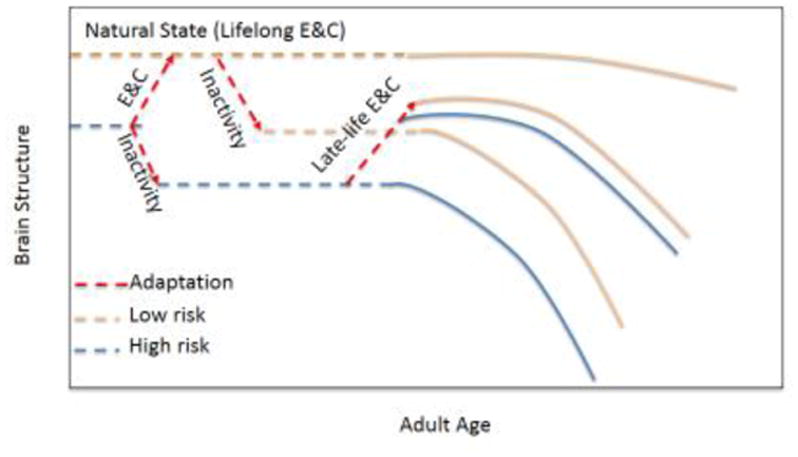
The Adaptive Capacity Model (ACM). Dotted lines indicate adult lifespan prior to detectable cognitive decline due to aging or neurodegenerative disease. Solid lines indicate periods of potentially observable cognitive decline. Peach lines represent individuals with low risk for late-life neurodegenerative disease (e.g., Alzheimer’s disease or cerebrovascular disease), while blue lines represent those with high risk for neurodegenerative disease. It is expected that individual differences in early life development, such as with early access to healthcare, in utero exposures, nutrition, genetics (e.g., apolipoprotein E ε4 alleles), and other early lifestyle and health factors can alter the start point for the adult trajectory of risk for developing age-related cognitive decline or late-life neurodegenerative disease. The ACM predicts that individuals who engage in cognitively challenging aerobic exercise (E&C) will maintain brain structure across much of their lifespan, as this represents the brain’s natural state, consistent with our evolutionary history. When met with periods of inactivity during adult aging, the brain responds adaptively by diminishing capacity to reduce energy costs, leading to decreased structure and associated function, as reflected by age-related regional brain atrophy. Individuals with reduced brain structure due to chronic inactivity experience greater vulnerability to either age-related cognitive decline (low risk) or developing neurodegenerative disease (high risk). Importantly, the ACM suggests that engaging in late-life E&C after extended periods of inactivity can adaptively increase capacity, altering the trajectory of brain changes to reduce the impacts of cognitive aging and the risk for dementia during the adult lifespan. It is expected that cognitive decline becomes evident when the combination of reduced capacity and level of risk for neurodegenerative disease is sufficient to overcome an individual’s constellation of lifelong developed capabilities, leading to observable effects of cognitive aging or neurodegenerative disease. Thus, for a young adult, diminished regional brain morphology due to inactivity can be offset by driving existing task-related networks to capacity, engaging alternative brain networks, or using a combination of both to maintain levels of cognitive performance. For an older adult with adaptively reduced brain capacity due to chronic inactivity, as reflected by regional brain atrophy, and especially for those with higher risk for developing late-life neurodegenerative disease pathology, the use of such existing task-related and/or alternative brain networks are no longer sufficient to hold off the expression of cognitive decline related to aging or neurodegenerative disease. In turn, the ACM predicts that the course of cognitive decline and the associated lifelong trajectory can be altered by engagement in E&C during the adult lifespan.
A key element of the ACM is the identification of stimuli for maintaining brain capacity. We suggest that the combination of exercise and cognitive engagement acts as an acute stimulus for preservation or enhancement of neural capacity (see Figure 2A and Box 2), and that the effects of long-term physically active cognitive challenges protect the brain from energy-saving atrophy across the lifespan. At its core, movement and physical activity require activation of motor and sensory domains (Figure 2A). However, movement through complex environments also engages several components of the executive function domain including planning, inhibition, monitoring, and attentional shifting/multi-tasking [57]. As locomotor speed increases (e.g., during high speed walking or running), processing demands on the brain consequently increase as well [58]. Finally, during the search for food (e.g., foraging), demands are placed on several aspects of memory and spatial navigation [59,60]. Thus, exercise itself can be cognitively challenging (see also [61]), and added demands of movement and search in complex environments increase cognitive requirements during exercise.
Figure 2.

Mechanisms underlying the ACM. A. Cognitively challenging physical activity, traditionally experienced during foraging, represents the evolutionary origins of the ACM. During foraging bouts, individuals must combine aerobic activity with control of motor systems, spatial navigation and memory, executive functions including decision making and planning, and control of sensory and attentional systems. This combination makes foraging a cognitively complex behavior, requiring an adaptive neural response to processing demands that can be further amplified when moving through novel environments and at increasing speeds. B. Acute effects of aerobic exercise. At the acute level, cognitively challenging aerobic activity (e.g., during foraging) begins with a neurogenic trigger (physical movement). Movement initiates the upregulation of neurotrophins and increased brain perfusion. In turn, the effects may induce hippocampal neurogenesis, as well as synaptogenesis and myelin remodeling with associated white matter (WM) and cortical brain effects. These non-specific effects are referred to as on-demand potential that may be realized when combined with cognitive challenges during or immediately following the aerobic activity. If these challenges occur, the acute effects of cognitively challenging exercise lead to neuron survival in the hippocampus and strengthened connections and myelination in key white matter tracts and associated cortical brain regions. C. Lifetime effects of cognitively challenging exercise. Combining aerobic exercise and cognitive challenges across the lifespan leads to the maintenance of brain structure and associated function during aging. Inactivity over the lifespan leads to an adaptive brain response that reduces structure (i.e., brain atrophy), potentially leading to the expression of cognitive decline or neurodegenerative disease. However, increasing cognitively challenging physical activity (Ex + Cog) after a lifetime of inactivity can alter the trajectory of cognitive aging and potentially reduce risks for the development of neurodegenerative disease.
BOX 2. Is exercise-induced neuroplasticity enhanced by human evolution?
While we believe the Adaptive Capacity Model (ACM) has applications for both humans and non-human animal models, there is reason to suspect that humans are uniquely sensitive to combined exercise and cognitive training. In fact, human evolutionary history suggests that the links between cognitive stimuli and exercise may be especially beneficial to humans compared with our closest living relatives. If true, these links may be a key element of our evolutionary success and suggest that use of animal models may require a careful consideration of species-specific evolutionary histories, as well as the inherent differences in brain organization and corresponding cognitive processes.
As described in the main text, human evolution was marked by a transition from relatively low levels of aerobic physical activity to higher levels of activity beginning around 2 million years ago [25,110,111]. This shift was spurred by a changing climate that forced our ancestors from more forested environments with high food density to more open habitats and an associated need to hunt and forage for heterogeneously distributed foods over long distances, potentially requiring greater spatial navigation, memory, and attention skills [59,60,110]. Our early bipedal ancestors fed on mainly fruits and vegetation in more forested areas, a lifestyle in some ways more similar to that of living chimpanzees, where foraging may not have required the same set of cognitive challenges (see [112]). Daily mobility in these species was likely low, with shorter daily travel needed for foraging bouts [113]. The shift to hunting and gathering not only altered the types of foods our ancestors accessed, but also increased aerobic physical activity and daily travel distances [25,39].
These differences in foraging strategy may have led to differences in neural susceptibility to plasticity. In cross-sectional analyses, humans have a greater degree of variation in brain morphological asymmetry compared with chimpanzees [114], suggesting potentially higher levels of plasticity across the lifespan. In a follow-up study of brain structure from imaging scans of related individuals (twin humans and closely related chimpanzees), Gomez-Robles et al. [115] showed that while total brain size was highly heritable in both species, the size of specific brain structures was less heritable in humans than chimpanzees. These results suggest that the human brain may be uniquely adapted to widespread neuroplasticity compared with the chimpanzee brain [115]. It is possible that the human brain remains flexible throughout life as an adaptation to the varied environments and cognitive demands humans experience. A more homogeneous environment in which to forage (e.g., tropical forests of chimpanzees) may have led selection to canalize brain structure.
The ACM builds on the work of Kempermann et al. [62] who first suggested that exercise in animal models stresses cognitive domains linked to foraging, triggering a neurogenic response in the hippocampus. The link between cognitive demands of foraging and exercise may have been a strong selection pressure for exercise-induced neurogenesis, as well as for modifying cortical brain connectivity through changes in synaptogenesis and myelination. Kempermann et al. [62] suggest the brain uses movement as a trigger for neurogenesis through the production of BDNF, signaling the possible cognitive needs that might benefit from the generation of new neurons. Survival of these neurons may be enhanced by cognitive challenges or novelty experienced during or after physical activity [62,63]. This is not to suggest that neurogenesis only leads to long-term benefits when combined with cognitive challenge, but that there is an increased effect on neuron survival when physical and cognitive challenges are combined. In addition, previous experiences may predict neurogenic needs, and thus, the size of the activity-generated pool is related to the prior levels of novelty and need experienced by an individual during exercise earlier in life [62]. In addition, Kempermann (2008) suggests that novelty and exploration may be key elements of these combined physical and cognitive challenges. Recent work provides a clue as to how movement triggers production of neurotrophins critical for neuroplasticity [64]. In rodent models, exercising muscles produce myokines, including FNDC5 and cathepsin B, which regulate metabolic processes and may improve muscle function while also inducing increased levels of BDNF [64–66].
On an acute level, a single bout of exercise would therefore trigger biochemical mechanisms that begin at the level of the muscle and can be utilized by the brain if faced with cognitive demands (“on-demand potential”; Figure 2B). When multiple cognitive domains are engaged during and shortly after movement, this on-demand potential would generate beneficial effects on the brain that lead to improved foraging success. It is important to note that exercise itself can be cognitively demanding. Thus, it is likely that, for example, neuronal survival can and does occur without specific additional higher-order cognitive challenges due to the inherent cognitive demands of exercise [61], however this hypothesis suggests that adding higher-level cognitive demands can enhance the exercise-induced benefits.
Over a lifetime, stresses on this system would act as a physiological stimulus that reduces the potential for energy-saving atrophy (Figure 2C). By contrast, a lack of overall exercise will reduce the triggered biochemical mechanisms available to convert cognitive effort into brain benefits, and signal energy saving reductions of brain volume in areas not cognitively engaged. Exercise without cognitive demands may lead to sub-optimal brain benefits, since on-demand potential is not fully realized. Thus, the well-established morphological effects of brain aging [67–70] may, in part, reflect the relatively inactive lifestyles of adults living in modern industrialized societies.
Regionally specific effects and origins of healthy aging
In addition to these more general predictions regarding overall brain structure, the ACM allows us to develop predictions for brain regions that may preferentially show exercise-induced neuro-plastic effects when specific cognitive demands are linked to physical activity (Figure 1B). A growing body of evidence suggests that neuroplasticity in the adult brain depends, at least to a great extent, on a combination of neurogenesis, synaptogenesis, and the capacity for myelin remodeling to alter the brain’s response to changes in environmental experience and developing disease [71,72]. Since neurogenesis may be more energetically expensive to maintain and activate [73], there may be evolutionary reasons for the differential reliance on neuro-plastic regional brain responses through adaptive changes in synaptic plasticity and myelin remodeling and/or myelination. We predict capacity maintenance through neurogenesis only when necessary to meet specific cognitive demands. For example, in the hippocampus, exercise-induced neurogenesis is a key aspect of spatial pattern separation [74,75], likely important for successful foraging [76]. However, maintenance and strengthening of white matter tracts may provide an energetically efficient and consistent benefit to motor control or executive functions [77,78], rather than depending on neurogenesis in broader regions of the brain that are less functionally localized or regionally accessible to the formation of new neurons during adulthood [79]. Maintenance or enhancement of brain structure and function in cortical brain regions may be related to synaptogenesis and white matter tract integrity as part of an energy saving mechanism. In turn, loss of synapses and white matter integrity in response to decreased activity would lead to a measurable reduction in gray matter, expressed together as regional brain atrophy. In this case, reductions in those cognitive activities preferentially engaged during physically active foraging behavior (i.e., executive functions, memory, spatial navigation, and processing speed) may lead selective brain regions and their associated neural systems to be more vulnerable to brain atrophy during healthy aging, as part of a neuroplastic response to reduced activity. As such, the observed selective regional vulnerability to atrophy of the frontal, temporal, and other brain regions often seen during healthy cognitive aging may, in part, reflect the mismatch between our human evolutionary history and the more sedentary lifestyles of today.
Not only does the ACM provide clear predictions and a foundation for novel interventions, this model may also help to explain the origins of healthy brain aging, why there is variation across human studies of exercise-induced neuroplasticity, and how we can design interventions to produce more consistently beneficial outcomes. Although atrophy can occur in response to brain injury or the pathology of developing neurodegenerative disease, in the context of neurologically healthy aging, the ACM suggests that age-related brain atrophy is part of a physiologically adaptive, energy saving response to reduced lifestyle demands for cognitively engaged physical activity. Variance in outcomes may be linked to use of a wide-range of exercise modes in prior interventions, including stationary equipment and home-based walking programs [80,81]. It is possible that human exercise in relatively cognitively impoverished environments (e.g., gym-based stationary exercise or lab-based exercise interventions) does not provide the optimal cognitive stimulus to maximize the effects of exercise on the brain. In addition, aerobic exercise through largely known environments (e.g., participants’ own neighborhoods) with expected environmental stimuli may not maximize the potential benefits of exercise for the brain. Finally, cross-sectional studies often pool subjects based on a wide variety of exercise experiences and levels of activity with no way of teasing apart the cognitive challenges of specific exercise regimes. Without controlling for individual differences in activity-linked cognitive demands, the ACM predicts a high degree of variation across the brain in responding to physical activity, along with corresponding differences in effect sizes.
Support for the Adaptive Capacity Model
Combined exercise and cognitive training in animal models
Work in animal models provides support for the basic mechanisms underlying the ACM and has generally focused on exercise-induced hippocampal neurogenesis in response to combinations of exercise and environmental enrichment. These studies support the predictions of the ACM by showing that new neurons generated through exercise are more likely to survive when exercise bouts are followed by some kind of cognitive challenge. For example, Fabel et al. [82] sequentially combined physical activity and cage enrichment in mice. Their study showed that cage enrichment alone, and wheel running alone both led to enhanced hippocampal neurogenesis. However, when researchers followed a period of wheel-running with a period of cage enrichment, an additive effect occurred and led to significantly increased neurogenesis. In another study, Motta-Teixeira et al. [83] exposed groups of rats that had been exercised for one week to a cognitive task that stressed spatial working memory (a Morris water maze where platform location changed regularly). Compared to exercised subjects who performed a task with a visible platform, a greater number of new neurons survived in the hippocampus in the group that had the cognitive challenge following exercise. In addition, the number of new neurons that survived was inversely correlated with the time delay in presenting the cognitive task, supporting the idea that close temporal proximity of physical and cognitive activity may be especially beneficial [83]. It is important to note that studies have also shown that wheel running alone can increase neurogenesis without the addition of cognitive enrichment [12,84,85]. These studies suggest that there is clearly a benefit to isolated exercise, that exercise may include cognitive demands, and that the additive effects of cognitive challenge may be dependent on the specific tasks or enrichment associated with the exercise bout.
Exercise itself seems to be a key trigger for BDNF upregulation (molecular mechanism for neurogenesis) in these combined activity and cognitive challenge experiments. For example, mice housed with running wheels show significantly higher levels of BDNF in the hippocampus compared with mice living in similarly enriched environments without access to running wheels [84]. Bechara and Kelly [86] showed that, while enrichment may have an effect on object recognition memory, only enrichment combined with exercise leads to upregulation of BDNF and associated cell proliferation in the hippocampus.
Finally, exercise intensity seems to play a role in the mechanisms underlying the neural benefits of aerobic activity. For example, Inoue et al. [87] showed that spatial memory and hippocampal neurogenesis are both enhanced by moderate intensity exercise, but that higher exercise intensities do not provide a similar benefit. These results support the notion that adaptive capacity changes are tuned to the moderate aerobic intensities used during foraging (e.g., [61]). Thus, animal models provide strong evidence that combinations of aerobic exercise and cognitive challenge stimulate specific beneficial effects in the brain.
Combined exercise and cognitive training in humans
A recent rise in interest in combining exercise and cognitive training interventions provides an opportunity to explore potential support for the ACM in humans. In general, researchers have used one of two experimental intervention designs [88]. In “separate” designs, exercise sessions are performed independently from cognitive training sessions, with delays of hours or even days. In “simultaneous” designs, exercise and cognitive training sessions occur at the same time. A meta-analysis that included 20 studies suggests that generally combining physical activity and cognitive training leads to greater cognitive improvements compared to control or physical activity only groups, but not compared with groups engaged only in cognitive training [88]. However, the comparison with cognitive training alone was based solely on “separate” study designs, leaving the potential benefit of simultaneous training relative to cognitive training alone unaddressed [88].
When examined carefully, there is more support for a major cognitive effect when exercise and cognitive training are performed close in time, in line with the proposed ACM. For example, the studies that show no significant cognitive improvements in combined training groups generally have individuals engaging in cognitive and aerobic training on separate days [89–92]. When training sessions are performed sequentially on the same day, the effects of combined training on cognition are stronger, and generally improve when the delay between sessions is minimized [93]. For example, a single bout of exercise performed either immediately before or after learning a motor skill led to improvements in skill acquisition in healthy adults, while exercise performed 1 or 2 hours following task learning led to no improvements in skill acquisition [94,95].
While close timing seems to be key for improvements, a recent study suggests that certain aspects of memory may actually benefit from a time delay [96]. For example, long-term memory assessed using a declarative memory task was improved following a four-hour delay between learning and exercise, compared with no delay [96]. While this result seems to contradict the predictions of the ACM regarding proximity of exercise and cognitive challenge, it is important to note that cognitively demanding foraging typically takes place over several hours. It is possible that memory consolidation in a declarative task benefits from a time delay, while procedural memory tasks and executive functions benefit from training performed in closer proximity to the physical activity. In these cases, neural-based memory consolidation may be linked with cognitive demands faced earlier in a foraging bout.
Fewer studies have examined simultaneous exercise and cognitive training, but the results of such studies are highly promising, especially when the cognitive task is thematically linked to the exercise session. Anderson-Hanley et al. [97] showed, in a randomized controlled trial, that three months of exercise on a stationary bicycle where subjects controlled a bike in a virtual tour (e.g., cognitively engaged in the movement on a screen) significantly improved executive functions compared to individuals who exercised on the bike without the cognitive stimulation. BDNF levels increased following the three-month trial in both groups, but the increase was significantly higher in the virtual bike tour group, suggesting a BDNF enhancement during the cognitive challenge [97]. In an extension of this work, Barcelos et al. [98] recently showed that a similar trial that included a group playing a video game while bicycling showed an even greater improvement in executive functions relative to exercise with the virtual tour alone. These study designs, where the physical and cognitive challenge are associated with a common goal-directed activity, are consistent with our evolutionary-neuroscience hypothesis and provide strong support for the ACM.
Other simultaneous designs have also shown encouraging results. For example, a cognitive and physical training program using a virtual dance game and treadmill walking combined with a memory task led to significant cognitive improvements in executive function and episodic memory that were maintained in a 1-year follow-up [99]. In another study, recall of foreign language vocabulary was greater when subjects learned vocabulary while exercising on a stationary cycle compared with subjects who learned vocabulary while sedentary [100].
In addition to studies that directly examined the effects of simultaneous aerobic exercise and cognitive training, there is evidence that low intensity movement itself is a key element of successful spatial navigation in humans [101]. In a series of studies, Lövden and colleagues examined changes in cognitive performance, markers of neuroplasticity, and changes in brain structure in individuals that completed a four-month training regime of virtual spatial navigation tasks performed while walking at a low intensity on a treadmill compared with four months of walk training without navigation. After the intervention, the combined training group performed better on navigation tasks than the group that only walked [101,102]. Structural neuroimaging analyses of the hippocampus indicated that combined training over four months reduced age-related declines in hippocampal volume compared to four months of treadmill walking alone [101]. Lövden and colleagues [102] also showed that, in individuals with a polymorphism that enhances activity-dependent BDNF production, four months of combined training increased imaging markers of neural integrity (hippocampal N-acetylaspartate) compared with four months of treadmill walking without the navigation task, suggesting at least one genetic mechanism and associated molecular pathway for differences in response to the combination of exercise and cognitive training.
Overall, these studies strongly support the hypothesis that combining physical activity and cognitively demanding tasks can have a greater effect on the human brain than aerobic exercise alone. Evidence suggests that the timing of exercise in relation to the cognitive challenge is important, and that long delays (e.g., performing tasks on separate days) do not have as strong a positive impact on cognition. For certain cognitive domains, simultaneous exercise and cognitive training may lead to the greatest benefit, providing a firm foundation for the ACM.
Conclusions
Although there are still many questions left unanswered, there is strong evidence that physical activity is beneficial for brain health and growing evidence that combining exercise with cognitive tasks may provide neural protection over the adult lifespan. The ACM differs from other models of human brain health and aging by providing a blueprint, based on an evolutionary neuroscience approach, for generating testable predictions linking exercise to cognitive health late in life, and by suggesting that novel interventions designed to stimulate key cognitive domains during exercise may be especially beneficial to brain health across the lifespan. Importantly, by extending the energy-minimizing principle to the evolution of the human brain, the ACM suggests that brain atrophy observed in neurologically healthy aging is part of a neuroplastic adaptive response to lower energy demands due to suboptimal levels of physical-cognitive activity over the lifespan, providing a basis for the effects of brain and cognitive aging that is distinct from neurodegenerative disease and brain injury.
By starting with an evolutionary neuroscience foundation, the ACM provides a clear direction for improving on areas of previous work. We can generate interventions that challenge the range of cognitive domains likely key to foraging success, leveraging our evolutionary history, and therefore, focusing on those cognitive processes that may be most responsive in the context of mobility. For example, combining exercise with spatial navigation and memory tasks provides a clear link to the likely cognitive demands of foraging. Similarly, dual tasking (e.g., performing cognitive tasks while walking) is a fundamental element of foraging and executive planning and motor control functions are challenged while moving on the landscape. These kinds of tasks can be generated through software applications for use during exercise.
The ACM also predicts that human physiology is well adapted to moderate aerobic intensities, suggesting that cognitively demanding exercise at very high or very low intensities may not provide the greatest benefits. While researchers have not explored the effects of changing exercise intensities in these combined challenges on cognitive performance, there is some evidence that, as in rodent models, moderate intensity exercise provides the greatest cognitive benefits in humans [103], in line with predictions of the ACM, and future work in humans should alter exercise intensities in combined challenges. While these moderate intensities may be linked to our evolutionary history, there is little evidence to date the mode of physical activity (e.g., cycling vs. walking or running) has a large impact on the cognitive benefits of activity. This is likely because the triggers for exercise-induced plasticity are engagement of the aerobic system, rather than a specific pattern of movements per se, provided that the level of inherent cognitive challenge during the exercise activity is comparable. Finally, presenting both novel environments and novel tasks may be key drivers of exercise-induced neuroplasticity. Such environments can be experienced in real-life situations, or through virtual reality or 2D software applications, however more detailed studies of the effects of novel experiences is needed to support this hypothesis.
While the mechanisms underlying the beneficial effects of simultaneous exercise and cognitively demanding tasks have yet to be fully delineated, we believe there is ample evidence to support the further investigation of these methods in studies across the lifespan. The ACM illustrates that taking a cross-disciplinary, evolutionary neuroscience approach and using cross-cutting, clinical-translational methods to study the brain-body link may help us gain a better understanding of how and why differences in lifestyle influence the course of healthy and pathological aging, potentially leading to better treatment options and prevention therapies. Furthermore, extending this approach to the realms of affective and social neuroscience may lead to a comprehensive framework for understanding how engaging in multiple lifestyle behaviors influence brain structure and function over the lifespan. Future studies that integrate knowledge of our unique evolutionary history with clinical and cognitive neuroscience research may help us to better understand, treat, and prevent the effects of human brain aging and disease in the present day.
Outstanding Questions Box.
A key aspect of the ACM is that exercise combined with cognitive demands can lead to long-term maintenance of brain structure and function if experienced consistently over time. However, there is a great need for more research that details the links between cognitively demanding exercise and brain health proposed by the ACM, especially at key time-points across the lifespan that help tease apart the best ways to use physical activity to improve brain aging outcomes. For example, while studies have demonstrated beneficial impacts of self-reported early-adult exercise on brain aging[7,87–93], no studies to date have assessed the impact of lifelong simultaneous exercise and cognitive challenge on the brain. We suggest that several unresolved questions could form the basis of novel intervention studies:
How do the effects of cognitively demanding exercise differ across the human lifespan?
Are there ways to engage the beneficial trigger mechanism of cognitively demanding exercise without stressing the aerobic system?
What intensity levels and durations work best for initiating the cognitive improvements associated with exercise?
Is there a genetic predisposition for a beneficial response to cognitively demanding aerobic activity?
Are there particular types of exercise (e.g., running, walking, cycling, others) that maximize the beneficial response to cognitively demanding aerobic activity?
Can cognitively demanding exercise prevent, delay, or diminish the effects of developing neurodegenerative disease or brain injury during aging?
Trends Box.
Recent work has shown that exercise can significantly improve brain structure and function in adults, especially during aging
We currently lack a comprehensive theoretical model to explain why exercise can lead to improved brain function
Taking an evolutionary neuroscience approach suggests that physiological systems, including the brain, respond to activity-related stress by expanding capacity, and that reductions in capacity represent an energy-minimizing strategy in response to inactivity
From an evolutionary neuroscience perspective, physical activity stresses brain function due to the cognitively demanding foraging context in which our ancestors engaged in aerobic physical activity
The proposed Adapative Capacity Model (ACM) links evolutionary theory with cognitive neuroscience to show that cognitively demanding exercise is beneficial to brain structure and function, and that we can take advantage of this adaptation to help prevent declines due to aging and developing neurological disease.
Acknowledgments
The authors would like to acknowledge support from the National Institute on Aging (AG019610, AG049464), the National Science Foundation (1440867), the state of Arizona and ADHS, the Arizona Advanced Research Institute for Biomedical Imaging, Tech Launch Arizona, Ken and Linda Robin, and the McKnight Brain Research Foundation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Colcombe SJ, Kramer AF. Fitness effects on the cognitive function of older adults: a meta-analytic study. Psychol Sci. 2003;14:125–130. doi: 10.1111/1467-9280.t01-1-01430. [DOI] [PubMed] [Google Scholar]
- 2.Cotman CW, Berchtold NC. Exercise: a behavioral intervention to enhance brain health and plasticity. Trends Neurosci. 2002;25:295–301. doi: 10.1016/s0166-2236(02)02143-4. [DOI] [PubMed] [Google Scholar]
- 3.Hillman CH, et al. Be smart, exercise your heart: exercise effects on brain and cognition. Nat Rev Neuro. 2008;9:58–65. doi: 10.1038/nrn2298. [DOI] [PubMed] [Google Scholar]
- 4.Kramer AF, et al. Exercise, cognition, and the aging brain. J Appl Physiol. 2006;101:1237–1242. doi: 10.1152/japplphysiol.00500.2006. [DOI] [PubMed] [Google Scholar]
- 5.Buchman A, et al. Total daily physical activity and the risk of AD and cognitive decline in older adults. Neurology. 2012;78:1323–1329. doi: 10.1212/WNL.0b013e3182535d35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Intlekofer KA, Cotman CW. Exercise counteracts declining hippocampal function in aging and Alzheimer’s disease. Neurobiol Dis. 2013;57:47–55. doi: 10.1016/j.nbd.2012.06.011. [DOI] [PubMed] [Google Scholar]
- 7.Rovio S, et al. Leisure-time physical activity at midlife and the risk of dementia and Alzheimer’s disease. Lancet Neurol. 2005;4:705–711. doi: 10.1016/S1474-4422(05)70198-8. [DOI] [PubMed] [Google Scholar]
- 8.Colcombe SJ, et al. Aerobic exercise training increases brain volume in aging adults. J Gerontol. 2006;61A:1166–1170. doi: 10.1093/gerona/61.11.1166. [DOI] [PubMed] [Google Scholar]
- 9.Ehninger D, Kempermann G. Neurogenesis in the adult hippocampus. Cell Tissue Res. 2008;331:243–250. doi: 10.1007/s00441-007-0478-3. [DOI] [PubMed] [Google Scholar]
- 10.Fabel K, et al. VEGF is necessary for exercise-induced adult hippocampal neurogenesis. Eur J Neurosci. 2003;18:2803–2812. doi: 10.1111/j.1460-9568.2003.03041.x. [DOI] [PubMed] [Google Scholar]
- 11.Van Praag H. Neurogenesis and exercise: past and future directions. Neuromolecular Med. 2008;10:128–140. doi: 10.1007/s12017-008-8028-z. [DOI] [PubMed] [Google Scholar]
- 12.Van Praag H, et al. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat Neurosci. 1999;2:266–270. doi: 10.1038/6368. [DOI] [PubMed] [Google Scholar]
- 13.Allen SJ, Dawbarn D. Clinical relevance of the neurotrophins and their receptors. Clin Sci. 2006;110:175–191. doi: 10.1042/CS20050161. [DOI] [PubMed] [Google Scholar]
- 14.Cotman CW, et al. Exercise builds brain health: key roles of growth factor cascades and inflammation. Trends Neurosci. 2007;30:464–472. doi: 10.1016/j.tins.2007.06.011. [DOI] [PubMed] [Google Scholar]
- 15.Vaynman S, et al. Hippocampal BDNF mediates the efficacy of exercise on synaptic plasticity and cognition. Eur J Neurosci. 2004;20:2580–2590. doi: 10.1111/j.1460-9568.2004.03720.x. [DOI] [PubMed] [Google Scholar]
- 16.Angevaren M, et al. Physical activity and enhanced fitness to improve cognitive function in older people without known cognitive impairment. Cochrane Db Syst Rev. 2008:3. doi: 10.1002/14651858.CD005381.pub3. [DOI] [PubMed] [Google Scholar]
- 17.Etnier JL, et al. The influence of physical fitness and exercise upon cognitive functioning: A meta-analysis. J Sport Exerc Psychol. 1997;19:249–277. [Google Scholar]
- 18.Heyn P, et al. The effects of exercise training on elderly persons with cognitive impairment and dementia: A meta-analysis. Arch Phys Med Rehabil. 2004;85:1694–1704. doi: 10.1016/j.apmr.2004.03.019. [DOI] [PubMed] [Google Scholar]
- 19.Kramer AF. Ageing, fitness and neurocognitive function. Nature. 1999;402:750–750. doi: 10.1038/22682. [DOI] [PubMed] [Google Scholar]
- 20.Smith PJ, et al. Aerobic exercise and neurocognitive performance: a meta-analytic review of randomized controlled trials. Psychosom Med. 2010;72:239–252. doi: 10.1097/PSY.0b013e3181d14633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.van Uffelen JG, et al. The effects of exercise on cognition in older adults with and without cognitive decline: a systematic review. Clin J Sport Med. 2008;18:486–500. doi: 10.1097/JSM.0b013e3181845f0b. [DOI] [PubMed] [Google Scholar]
- 22.Verstynen TD, et al. Caudate nucleus volume mediates the link between cardiorespiratory fitness and cognitive flexibility in older adults. J Aging Res. 2012;2012 doi: 10.1155/2012/939285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Roig M, et al. The effects of cardiovascular exercise on human memory: a review with meta-analysis. Neurosci Biobehav Rev. 2013;37:1645–1666. doi: 10.1016/j.neubiorev.2013.06.012. [DOI] [PubMed] [Google Scholar]
- 24.Eaton SB, Eaton SB. An evolutionary perspective on human physical activity: implications for health. Comp Biochem Physiol A Mol Integr Physiol. 2003;136:153–159. doi: 10.1016/s1095-6433(03)00208-3. [DOI] [PubMed] [Google Scholar]
- 25.Lieberman D. The Story of the Human Body: Evolution, Health and Disease. Penguin UK: 2013. [PubMed] [Google Scholar]
- 26.Lieberman DE. Is Exercise Really Medicine? An Evolutionary Perspective. Curr Sports Med Rep. 2015;14:313–319. doi: 10.1249/JSR.0000000000000168. [DOI] [PubMed] [Google Scholar]
- 27.Stearns SC, Koella JC. Evolution in health and disease. Oxford University Press; 2008. [Google Scholar]
- 28.Cordain L, et al. Physical activity, energy expenditure and fitness: an evolutionary perspective. Int J Sports Med. 1998;19:328–335. doi: 10.1055/s-2007-971926. [DOI] [PubMed] [Google Scholar]
- 29.Eaton SB, et al. Stone agers in the fast lane: chronic degenerative diseases in evolutionary perspective. Am J Med. 1988;84:739–749. doi: 10.1016/0002-9343(88)90113-1. [DOI] [PubMed] [Google Scholar]
- 30.Malina RM, Little BB. Feature Article Physical Activity: The Present in the Context of the Past. Am J Hum Biol. 2008;20:373–391. doi: 10.1002/ajhb.20772. [DOI] [PubMed] [Google Scholar]
- 31.Noakes T, Spedding M. Olympics: Run for your life. Nature. 2012;487:295–296. doi: 10.1038/487295a. [DOI] [PubMed] [Google Scholar]
- 32.Aiello LC, Wells JC. Energetics and the evolution of the genus Homo. Annu Rev Anthropol. 2002:323–338. [Google Scholar]
- 33.Leonard WR, Robertson ML. Comparative primate energetics and hominid evolution. Am J Phys Anthropol. 1997;102:265–281. doi: 10.1002/(SICI)1096-8644(199702)102:2<265::AID-AJPA8>3.0.CO;2-X. [DOI] [PubMed] [Google Scholar]
- 34.Lieberman DE, et al. The First Humans–Origin and Early Evolution of the Genus Homo. Springer; 2009. Brains, brawn, and the evolution of human endurance running capabilities; pp. 77–92. [Google Scholar]
- 35.Pontzer H, et al. Energy expenditure and activity among Hadza hunter-gatherers. Am J Hum Biol. 2015;27:628–637. doi: 10.1002/ajhb.22711. [DOI] [PubMed] [Google Scholar]
- 36.Pontzer H, et al. Hunter-gatherer energetics and human obesity. PLoS One. 2012;7:e40503. doi: 10.1371/journal.pone.0040503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Raichlen DA, et al. Calcaneus length determines running economy: implications for endurance running performance in modern humans and Neandertals. J Hum Evol. 2011;60:299–308. doi: 10.1016/j.jhevol.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 38.Raichlen DA, Polk JD. Linking brains and brawn: exercise and the evolution of human neurobiology. Proc R Soc Lond B Biol Sci. 2013;280:20122250. doi: 10.1098/rspb.2012.2250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Raichlen DA, et al. Physical activity patterns and biomarkers of cardiovascular disease risk in hunter-gatherers. Am J Hum Biol. 2016 doi: 10.1002/ajhb.22919. [DOI] [PubMed] [Google Scholar]
- 40.Weibel ER, et al. The concept of symmorphosis: a testable hypothesis of structure-function relationship. Proc Natl Acad Sci USA. 1991;88:10357–10361. doi: 10.1073/pnas.88.22.10357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tanaka H, Seals DR. Invited review: dynamic exercise performance in masters athletes: insight into the effects of primary human aging on physiological functional capacity. J Appl Physiol. 2003;95:2152–2162. doi: 10.1152/japplphysiol.00320.2003. [DOI] [PubMed] [Google Scholar]
- 42.Ellison PT. Energetics and reproductive effort. Am J Hum Biol. 2003;15:342–351. doi: 10.1002/ajhb.10152. [DOI] [PubMed] [Google Scholar]
- 43.Gurven M, et al. Physical activity and modernization among Bolivian Amerindians. PLoS One. 2013;8:e55679. doi: 10.1371/journal.pone.0055679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mujika I, Padilla S. Detraining: loss of training-induced physiological and performance adaptations. Part II. Sports Med. 2000;30:145–154. doi: 10.2165/00007256-200030030-00001. [DOI] [PubMed] [Google Scholar]
- 45.Vona M, et al. Effects of different types of exercise training followed by detraining on endothelium-dependent dilation in patients with recent myocardial infarction. Circulation. 2009;119:1601–1608. doi: 10.1161/CIRCULATIONAHA.108.821736. [DOI] [PubMed] [Google Scholar]
- 46.Martin WH, et al. Effects of physical deconditioning after intense endurance training on left ventricular dimensions and stroke volume. J Am Coll Cardiol. 1986;7:982–989. doi: 10.1016/s0735-1097(86)80215-7. [DOI] [PubMed] [Google Scholar]
- 47.Bangsbo J, Mizuno M. Morphological and metabolic alterations in soccer players with detraining and retraining and their relation to performance 1988 [Google Scholar]
- 48.Montero-Fernandez N, Serra-Rexach J. Role of exercise on sarcopenia in the elderly. Eur J Phys Rehabil Med. 2013;49:131–143. [PubMed] [Google Scholar]
- 49.Bouchard C, et al. Familial aggregation of V̇o2 max response to exercise training: results from the HERITAGE Family Study. J Appl Physiol. 1999;87:1003–1008. doi: 10.1152/jappl.1999.87.3.1003. [DOI] [PubMed] [Google Scholar]
- 50.Swallow JG, et al. Phenotypic and evolutionary plasticity of organ masses in response to voluntary exercise in house mice1. Integr Comp Biol. 2005;45:426–437. doi: 10.1093/icb/45.3.426. [DOI] [PubMed] [Google Scholar]
- 51.Swallow JG, et al. Phenotypic and evolutionary plasticity of body composition in rats selectively bred for high endurance capacity. J Appl Physiol. 2010;109:778–785. doi: 10.1152/japplphysiol.01026.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Aiello LC, Wheeler P. The expensive-tissue hypothesis: the brain and the digestive system in human and primate evolution. Cur Anthropol. 1995;36:199–221. [Google Scholar]
- 53.Clarke DD, Sokoloff L. Regulation of cerebral metabolic rate 1999 [Google Scholar]
- 54.Sokoloff L. The metabolism of the central nervous system in vivo. Handbook of physiology, section I, Neurophysiology. 1960;3:1843–1864. [Google Scholar]
- 55.Liu-Ambrose T, et al. Emerging concept: ‘central benefit model’ of exercise in falls prevention. Br J Sports Med. 2012 doi: 10.1136/bjsports-2011-090725. bjsports-2011-090725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Raichlen DA, Alexander GE. Exercise, APOE genotype, and the evolution of the human lifespan. Trends Neurosci. 2014;37:247–255. doi: 10.1016/j.tins.2014.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Yogev-Seligmann G, et al. The role of executive function and attention in gait. Mov Disord. 2008;23:329–342. doi: 10.1002/mds.21720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Brick N, et al. Attentional focus in endurance activity: new paradigms and future directions. International Review of Sport and Exercise Psychology. 2014;7:106–134. [Google Scholar]
- 59.Cashdan E, Gaulin SJ. Why Go There? Evolution of Mobility and Spatial Cognition in Women and Men. Human Nature. 2016;27:1–15. doi: 10.1007/s12110-015-9253-4. [DOI] [PubMed] [Google Scholar]
- 60.Trumble BC, et al. No sex or age difference in dead-reckoning ability among Tsimane forager-horticulturalists. Human Nature. 2016;27:51–67. doi: 10.1007/s12110-015-9246-3. [DOI] [PubMed] [Google Scholar]
- 61.Raichlen DA, et al. Differences in Resting State Functional Connectivity between Young Adult Endurance Athletes and Healthy Controls. Front Hum Neurosci. 2016:10. doi: 10.3389/fnhum.2016.00610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kempermann G, et al. Why and how physical activity promotes experience-induced brain plasticity. Front Neurosci. 2010;4:189. doi: 10.3389/fnins.2010.00189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Curlik D, Shors T. Training your brain: do mental and physical (MAP) training enhance cognition through the process of neurogenesis in the hippocampus? Neuropharmacology. 2013;64:506–514. doi: 10.1016/j.neuropharm.2012.07.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Wrann CD. FNDC5/Irisin–their role in the nervous system and as a mediator for beneficial effects of exercise on the brain. Brain Plasticity. 2015;1:55–61. doi: 10.3233/BPL-150019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Moon HY, et al. Running-induced systemic cathepsin B secretion is associated with memory function. Cell Metab. 2016;24:332–340. doi: 10.1016/j.cmet.2016.05.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wrann CD, et al. Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway. Cell Metab. 2013;18:649–659. doi: 10.1016/j.cmet.2013.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Alexander GE, et al. Regional network of magnetic resonance imaging gray matter volume in healthy aging. Neuroreport. 2006;17:951–956. doi: 10.1097/01.wnr.0000220135.16844.b6. [DOI] [PubMed] [Google Scholar]
- 68.Bergfield KL, et al. Age-related networks of regional covariance in MRI gray matter: reproducible multivariate patterns in healthy aging. Neuroimage. 2010;49:1750–1759. doi: 10.1016/j.neuroimage.2009.09.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Raz N, et al. Neuroanatomical correlates of cognitive aging: evidence from structural magnetic resonance imaging. Neuropsychology. 1998;12:95–114. doi: 10.1037//0894-4105.12.1.95. [DOI] [PubMed] [Google Scholar]
- 70.Raz N, Rodrigue KM. Differential aging of the brain: patterns, cognitive correlates and modifiers. Neurosci Biobehav Rev. 2006;30:730–748. doi: 10.1016/j.neubiorev.2006.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Alexander GE. An emerging role for imaging white matter in the preclinical risk for Alzheimer Disease: linking β-Amyloid to myelin. JAMA Neurology. 2017;74:17–19. doi: 10.1001/jamaneurol.2016.4123. [DOI] [PubMed] [Google Scholar]
- 72.O’Rourke M, et al. Adult myelination: wrapping up neuronal plasticity. Neural Regener Res. 2014;9:1261–1264. doi: 10.4103/1673-5374.137571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Harris JJ, Attwell D. The energetics of CNS white matter. The Journal of Neuroscience. 2012;32:356–371. doi: 10.1523/JNEUROSCI.3430-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Clelland C, et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science. 2009;325:210–213. doi: 10.1126/science.1173215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Creer DJ, et al. Running enhances spatial pattern separation in mice. Proc Natl Acad Sci USA. 2010;107:2367–2372. doi: 10.1073/pnas.0911725107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Mattson MP. Lifelong brain health is a lifelong challenge: from evolutionary principles to empirical evidence. Ageing Res Rev. 2015;20:37–45. doi: 10.1016/j.arr.2014.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Scholz J, et al. Training induces changes in white-matter architecture. Nat Neurosci. 2009;12:1370–1371. doi: 10.1038/nn.2412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Voineskos AN, et al. Age-related decline in white matter tract integrity and cognitive performance: a DTI tractography and structural equation modeling study. Neurobiol Aging. 2012;33:21–34. doi: 10.1016/j.neurobiolaging.2010.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Paredes MF, et al. Brain size and limits to adult neurogenesis. J Comp Neurol. 2016;524:646–664. doi: 10.1002/cne.23896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Erickson KI, et al. Exercise training increases size of hippocampus and improves memory. Proc Natl Acad Sci USA. 2011;108:3017–3022. doi: 10.1073/pnas.1015950108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Topp R, Stevenson JS. The effects of attendance and effort on outcomes among older adults in a long-term exercise program. Res Nurs Health. 1994;17:15–24. doi: 10.1002/nur.4770170104. [DOI] [PubMed] [Google Scholar]
- 82.Fabel K, et al. Additive effects of physical exercise and environmental enrichment on adult hippocampal neurogenesis in mice. Front Neurosci. 2009;3:2. doi: 10.3389/neuro.22.002.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Motta-Teixeira LC, et al. Spatial learning and neurogenesis: Effects of cessation of wheel running and survival of novel neurons by engagement in cognitive tasks. Hippocampus. 2016;26:794–803. doi: 10.1002/hipo.22560. [DOI] [PubMed] [Google Scholar]
- 84.Kobilo T, et al. Running is the neurogenic and neurotrophic stimulus in environmental enrichment. Learn Mem. 2011;18:605–609. doi: 10.1101/lm.2283011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Mustroph ML, et al. Aerobic exercise is the critical variable in an enriched environment that increases hippocampal neurogenesis and water maze learning in male C57BL/6J mice. Neuroscience. 2012;219:62–71. doi: 10.1016/j.neuroscience.2012.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Bechara R, Kelly A. Exercise improves object recognition memory and induces BDNF expression and cell proliferation in cognitively enriched rats. Behav Brain Res. 2013;245:96–100. doi: 10.1016/j.bbr.2013.02.018. [DOI] [PubMed] [Google Scholar]
- 87.Inoue K, et al. Long-term mild, rather than intense, exercise enhances adult hippocampal neurogenesis and greatly changes the transcriptomic profile of the hippocampus. PLoS One. 2015;10:e0128720. doi: 10.1371/journal.pone.0128720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Zhu X, et al. The more the better? A meta-analysis on effects of combined cognitive and physical intervention on cognition in healthy older adults. Ageing Res Rev. 2016;31:67–79. doi: 10.1016/j.arr.2016.07.003. [DOI] [PubMed] [Google Scholar]
- 89.Barnes DE, et al. The Mental Activity and eXercise (MAX) trial: a randomized controlled trial to enhance cognitive function in older adults. JAMA internal medicine. 2013;173:797–804. doi: 10.1001/jamainternmed.2013.189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Legault C, et al. Designing clinical trials for assessing the effects of cognitive training and physical activity interventions on cognitive outcomes: the Seniors Health and Activity Research Program Pilot (SHARP-P) study, a randomized controlled trial. BMC Geriatr. 2011;11:1. doi: 10.1186/1471-2318-11-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Rahe J, et al. Cognitive training with and without additional physical activity in healthy older adults: cognitive effects, neurobiological mechanisms, and prediction of training success. Front Aging Neurosci. 2015:7. doi: 10.3389/fnagi.2015.00187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Shatil E. Does combined cognitive training and physical activity training enhance cognitive abilities more than either alone? A four-condition randomized controlled trial among healthy older adults. Front Aging Neurosci. 2013;5:8. doi: 10.3389/fnagi.2013.00008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Roig M, et al. Time-Dependent Effects of Cardiovascular Exercise on Memory. Exerc Sport Sci Rev. 2016;44:81–88. doi: 10.1249/JES.0000000000000078. [DOI] [PubMed] [Google Scholar]
- 94.Statton MA, et al. A single bout of moderate aerobic exercise improves motor skill acquisition. PLoS One. 2015;10:e0141393. doi: 10.1371/journal.pone.0141393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Thomas R, et al. Acute Exercise and Motor Memory Consolidation: The Role of Exercise Intensity. PLoS ONE. 2016;11:e0159589. doi: 10.1371/journal.pone.0159589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.van Dongen EV, et al. Physical Exercise Performed Four Hours after Learning Improves Memory Retention and Increases Hippocampal Pattern Similarity during Retrieval. Curr Biol. 2016;26:1722–1727. doi: 10.1016/j.cub.2016.04.071. [DOI] [PubMed] [Google Scholar]
- 97.Anderson-Hanley C, et al. Exergaming and older adult cognition: a cluster randomized clinical trial. Am J Prev Med. 2012;42:109–119. doi: 10.1016/j.amepre.2011.10.016. [DOI] [PubMed] [Google Scholar]
- 98.Barcelos N, et al. Aerobic and Cognitive Exercise (ACE) Pilot Study for Older Adults: Executive Function Improves with Cognitive Challenge While Exergaming. J Int Neuropsychol Soc. 2015;21:768–779. doi: 10.1017/S1355617715001083. [DOI] [PubMed] [Google Scholar]
- 99.Eggenberger P, et al. Does multicomponent physical exercise with simultaneous cognitive training boost cognitive performance in older adults? A 6-month randomized controlled trial with a 1-year follow-up. Clin Interv Aging. 2015;10:1335. doi: 10.2147/CIA.S87732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Schmidt-Kassow M, et al. Exercising during learning improves vocabulary acquisition: behavioral and ERP evidence. Neurosci Lett. 2010;482:40–44. doi: 10.1016/j.neulet.2010.06.089. [DOI] [PubMed] [Google Scholar]
- 101.Lövdén M, et al. Spatial navigation training protects the hippocampus against age-related changes during early and late adulthood. Neurobiol Aging. 2012;33:620.e629–620.e622. doi: 10.1016/j.neurobiolaging.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 102.Lövdén M, et al. Performance-related increases in hippocampal N-acetylaspartate (NAA) induced by spatial navigation training are restricted to BDNF Val homozygotes. Cereb Cortex. 2010:bhq230. doi: 10.1093/cercor/bhq230. [DOI] [PubMed] [Google Scholar]
- 103.McMorris T, Hale BJ. Differential effects of differing intensities of acute exercise on speed and accuracy of cognition: a meta-analytical investigation. Brain Cogn. 2012;80:338–351. doi: 10.1016/j.bandc.2012.09.001. [DOI] [PubMed] [Google Scholar]
- 104.Nyberg L, et al. Memory aging and brain maintenance. Trends Cogn Sci. 2012;16:292–305. doi: 10.1016/j.tics.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 105.Park DC, Reuter-Lorenz P. The adaptive brain: aging and neurocognitive scaffolding. Annu Rev Psychol. 2009;60:173. doi: 10.1146/annurev.psych.59.103006.093656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Satz P, et al. Brain and cognitive reserve: Mediator (s) and construct validity, a critique. J Clin Exp Neuropsychol. 2011;33:121–130. doi: 10.1080/13803395.2010.493151. [DOI] [PubMed] [Google Scholar]
- 107.Scarmeas N, et al. Influence of leisure activity on the incidence of Alzheimer’s disease. Neurology. 2001;57:2236–2242. doi: 10.1212/wnl.57.12.2236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Stern Y. What is cognitive reserve? Theory and research application of the reserve concept. J Int Neuropsychol Soc. 2002;8:448–460. [PubMed] [Google Scholar]
- 109.Stern Y, et al. Relationship between lifetime occupation and parietal flow Implications for a reserve against Alzheimer’s disease pathology. Neurology. 1995;45:55–60. doi: 10.1212/wnl.45.1.55. [DOI] [PubMed] [Google Scholar]
- 110.Bramble DM, Lieberman DE. Endurance running and the evolution of Homo. Nature. 2004;432:345–352. doi: 10.1038/nature03052. [DOI] [PubMed] [Google Scholar]
- 111.Carrier DR. The energetic paradox of human running and hominid evolution [and comments and reply] Cur Anthropol. 1984;25:483–495. [Google Scholar]
- 112.Kaplan H, et al. A theory of human life history evolution: diet, intelligence, and longevity. Evol Anthropol. 2000;9:156–185. [Google Scholar]
- 113.Pontzer H, Wrangham RW. Climbing and the daily energy cost of locomotion in wild chimpanzees: implications for hominoid locomotor evolution. J Hum Evol. 2004;46:315–333. doi: 10.1016/j.jhevol.2003.12.006. [DOI] [PubMed] [Google Scholar]
- 114.Gómez-Robles A, et al. Proc R Soc B. The Royal Society; 2013. Increased morphological asymmetry, evolvability and plasticity in human brain evolution; p. 20130575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Gómez-Robles A, et al. Relaxed genetic control of cortical organization in human brains compared with chimpanzees. Proceedings of the National Academy of Sciences. 2015;112:14799–14804. doi: 10.1073/pnas.1512646112. [DOI] [PMC free article] [PubMed] [Google Scholar]