

Abstract
Older adults are at a high risk of falls, and most falls occur during locomotor activities like walking. This study aimed to improve our understanding of changes in neuromuscular control associated with increased risk of falls in older adults in the presence of dynamic balance challenges during walking. Motor module (also known as muscle synergy) analyses identified changes in the neuromuscular recruitment of leg muscles during walking with and without perturbations designed to elicit the visual perception of lateral instability. During normal walking we found that a history of falls (but not age) was associated with reduced motor module complexity and that age (but not a history of falls) was associated with increased step-to-step variability of module recruitment timing. Furthermore, motor module complexity was unaltered in the presence of optical flow perturbations. The specific effects of a history of falls on leg muscle recruitment included an absence and/or inability to independently recruit motor modules normally recruited to perform biomechanical functions important for walking balance control. These results suggest that fallers do not recruit the appropriate motor modules necessary for well-coordinated walking balance control even in the presence of perturbations. The identified changes in the modular control of walking balance in older fallers may either represent a neural deficit that leads to poor balance control or a prior history of falls that results in a compensatory motor adaptation. In either case, our study provides initial evidence that a reduced motor repertoire in older adult fallers may be a constraint on their ability to appropriately respond to balance challenges during walking.
NEW & NOTEWORTHY This is the first study to demonstrate a reduced motor repertoire during walking in older adults with a history of falls but without any overt neurological deficits. Furthermore, using virtual reality during walking to elicit the visual perception of lateral instability, we provide initial evidence that a reduced motor repertoire in older adult fallers may be a constraint on their ability to appropriately respond to balance challenges during walking.
Keywords: gait, muscle synergy, stability, virtual reality, vision
INTRODUCTION
Older adults are at an exceptionally high risk of falls, and most falls occur during locomotor activities such as walking (Tinetti et al. 1988). Moreover, recent evidence suggests that the rate of injurious falls among older adults is accelerating and outpacing the growth in the older adult population (Cigolle et al. 2015). Thus there is a critical need to better understand the mechanisms governing falls risk in the elderly to empower our translational efforts to mitigate those risks. Numerous factors have been associated with falls risk in older adults, including age-associated declines in sensory, motor, and cognitive-motor acuity (Melzer et al. 2004; Menz et al. 2005; Springer et al. 2006; Thelen et al. 1996). Such declines may lead to altered neuromuscular control of walking that limits an individual’s ability to respond to environmental challenges that could precipitate a fall. This study aims to improve our understanding of changes in neuromuscular control associated with increased risk of falls in older adults, especially in the presence of dynamic balance challenges during walking.
Motor module (also known as muscle synergy) analysis has frequently been used to characterize altered neuromuscular control in mobility-impaired populations, but whether or how motor module recruitment is affected by normal aging nor how this is related to falls risk has received much less attention. Motor modules are defined as groups of coactive muscles that are flexibly recruited over time to transform movement goals into the required biomechanical output for a given motor behavior (Allen and Neptune 2012; Berniker et al. 2009; Chvatal et al. 2011; d’Avella and Bizzi 2005; Ting and Macpherson 2005). The number of motor modules recruited to perform a motor behavior is frequently used as a measure of neuromuscular complexity, where reduced neuromuscular complexity (i.e., fewer motor modules) has been associated with reduced walking performance in mobility impaired populations [e.g., stroke (Clark et al. 2010) Parkinson’s disease (Rodriguez et al. 2013), spinal cord injury (Fox et al. 2013; Hayes et al. 2014), and cerebral palsy (Steele et al. 2015; Tang et al. 2015)]. This reduced neuromuscular complexity further limits an individual’s ability to perform more complex locomotor tasks [e.g., changing speed, cadence, step length, and step height (Routson et al. 2014)], which require more precise balance control. Similarly, altered consistency and timing of motor module recruitment across the gait cycle is also related to reduced walking performance in mobility-impaired populations (Allen et al. 2017; Routson et al. 2013).
Perhaps due to subtler neuromuscular changes associated with normal aging, very few studies have sought to determine whether and how aging alters motor module recruitment associated with gait and balance performance. Evidence for altered motor module recruitment in old age comes from recordings of individual leg muscle activity patterns in which older adults typically coactivate their agonists and antagonists more than young adults (Franz and Kram 2013; Hortobágyi et al. 2009; Peterson and Martin 2010). Consistent with this implication, two recent analyses from the same cohort of subjects found small changes in motor module number, structure, and recruitment timing in older vs. young adults, most evident when subjects walked at faster than preferred cadences (Artoni et al. 2013; Monaco et al. 2010). The earlier of those studies concluded that the emerging muscle synergies may be more associated with the biomechanical demands of locomotion than with age-associated changes in neuromuscular control. However, this interpretation is inconsistent with findings that the modular structure and organization of walking balance control differs between groups with varying levels of balance proficiency and fall risk (Allen et al. 2017; Bowden et al. 2010; Clark et al. 2010; Sawers et al. 2015). Moreover, this cumulative literature suggests that the effects of a history of falls on motor module recruitment in older adults may be larger than the effects of age alone.
Growing evidence suggests that preclinical walking balance deficits, especially due to normal aging, may go undetected using measurements recorded during normal, unencumbered walking. Thus balance perturbations, a class of experimental paradigms used to provoke instability, have been increasingly used to study the neuromuscular mechanisms governing walking balance control. In the context of age-related balance impairment, older adults rely more on visual feedback than young adults to plan and orchestrate the motor adjustments needed to regulate balance during both standing and walking (Bugnariu and Fung 2007; Eikema et al. 2013; Jeka et al. 2010; O’Connor et al. 2008; Sundermier et al. 1996; Yeh et al. 2014). Accordingly, we have shown that optical flow perturbations during walking, or perturbations designed to elicit the visual perception of lateral instability, can reveal age-related differences in metrics of walking balance that are not otherwise apparent during normal, unperturbed walking (Francis et al. 2015; Franz et al. 2015; Qiao et al. 2018a). Moreover, in their seminar paper, Lord and Webster (1990) found that this reliance on visual feedback was much more pronounced in older subjects with a history of falls compared with nonfallers. Perhaps consequently, we recently discovered that optical flow perturbations of sufficient amplitude reduced dynamic stability during walking more in older adult fallers than in nonfallers (Qiao et al. 2018b). Together with evidence for disproportionate increases in antagonist coactivation in older adults with a history of falls (Marques et al. 2013), these findings allude to the potential for larger perturbation effects on motor module complexity due to a history of falls than to aging alone. Indeed, changes in motor module recruitment underlying the control of walking in the presence of such balance perturbations, while currently unknown, could inform the development of improved detection and falls prevention techniques.
Therefore, the purpose of this study was to investigate the effects of aging and a history of falls on the neuromuscular control of walking with and without optical flow balance perturbations. Specifically, we examined the complexity and consistency of motor module recruitment of leg muscles across varying levels of perturbation amplitudes. We hypothesized that 1) compared with young adults, older adults, and even more so, those with a history of falls, would exhibit decreased module complexity and consistency, 2) with larger effects in the presence of balance perturbations.
METHODS
Subjects
Twelve healthy young adults (6 female; means ± SD; age: 24.8 ± 4.6 yr; height: 1.70 ± 0.10 m; and mass: 65.4 ± 10.8 kg), eleven healthy older adults (6 female; age: 75.1 ± 5.8 yr; height: 1.75 ± 0.07; and mass: 70.3 ± 11.9 kg), and ten older adults with a history of falls (7 female age: 77.7 ± 7.7 yr; height: 1.7 ± 0.12 m; and mass: 71.3 ± 13.3 kg) participated in this study. A history of falls was determined through self-report of one or more falls in the past year as defined by the Kellogg International Work Group Definition. Subjects were excluded based on body mass index ≥30, sedentary lifestyle, orthopedic or neurological condition, or taking medication that causes dizziness. Visual acuity was also examined for all participants to ensure each had normal or corrected to normal vision (i.e., ~20/20). All subjects provided written informed consent before participating according to an experimental protocol approved by institutional review board of the University of North Carolina.
Data Collection and Processing
For all subjects, preferred overground walking speed was computed as the time taken to walk the middle 2 m of a 10-m walkway using two photocells (Brower Timing, Draper, UT). All further testing was performed while subjects walked on a dual-belt, force-measuring treadmill (Bertec, Columbus, OH). The treadmill was surrounded by a semicircular curved screen measuring 2.64-m high and 2.90-m wide (Fig. 1). Older adult subjects walked on the treadmill at their preferred walking speed (nonfallers: 1.17 ± 0.20 m/s and fallers: 1.04 ± 0.22 m/s, P = 0.082). Young adult data from a previously published study (Stokes et al. 2017) were reanalyzed for comparison to older adults. Young adult subjects walked on the treadmill at 1.25 m/s, which was comparable to their preferred overground walking speed (1.29 ± 0.18 m/s, P = 0.527). All subjects wore a harness secured via an overhead support system.
Fig. 1.

Subjects walked on a treadmill in an immersive virtual environment while watching a hallway with and without continuous mediolateral (ML) optical flow perturbations of 3 amplitudes. We recorded electromyographic activity from the following 7 leg muscles: tibialis anterior (TA), medial gastrocnemius (MG), soleus (SOL), peroneus longus (PL), vastus lateralis (VL), medial hamstrings (MH), and gluteus medius (GM). hs, Heel-strike.
After 5 min of acclimation to walking on the treadmill, all subjects completed four 2-min walking trials, in randomized order, while watching a speed-matched, virtual hallway rear projected onto the screen (Fig. 1) and controlled using Simulink Desktop Real-Time (MathWorks, Natick, MA), described in detail elsewhere (O’Connor 2012). Specifically, the real-time controller commanded the movement of the camera perspective in our rendered and projected virtual hallway (see Supplemental Video; Supplemental Material for this article is available online at the Journal website). The nominal position of the camera perspective was centered on the treadmill at a height prescribed to match that of each subject and the width of the virtual hallway replicated the width of projection screen. In some cases, walking trials consisted of continuous mediolateral oscillations that displaced the camera perspective relative to that nominal position on the treadmill, inducing optical flow perturbations with amplitudes of 20, 35, and 50 cm prescribed as follows. These perturbations were comprised of the sum of three sinusoids, such that the full amplitude was applied at 0.250 Hz and half that amplitude was applied at 0.125 and 0.442 Hz—designed to facilitate complex mediolateral oscillations that would be difficult to anticipate. Moreover, because the camera perspective translations were applied at the nominal position of the subject on the treadmill, the foreground moved at full amplitude while the far end of the hallway remained nearly stationary, thereby provoking balance corrections rather than heading corrections (see Supplemental Video). For all trials, subjects were simply instructed to “walk while looking down the hallway.”
Three-dimensional trajectories of 17 anatomical markers and 14 additional tracking markers affixed using rigid clusters placed on subjects’ pelvis and right and left legs were recorded (100 Hz) using a 14-camera motion capture system (Motion Analysis, Santa Rosa, CA). Marker trajectories were filtered using a fourth-order Butterworth filter with cut-off frequency of 8 Hz. We first scaled a 7-segment, 18-degree-of-freedom lower extremity model (Arnold et al. 2010) to subjects’ standing calibration trial, with functional hip joint centers identified from a leg circumduction task according to Piazza et al. (2001). From each subject’s scaled model, a global optimization inverse kinematics routine calculated time series of sagittal plane hip, knee, and ankle joint angles, and the coronal plane hip joint angle, by minimizing the weighted sum of squared differences between measured and modeled marker positions. We assembled stride-averaged joint angles for each condition using heel strikes identified from peaks in the fore-aft position of the heel markers relative to the sacral marker (Zeni et al. 2008). Finally, we extracted local maxima and minima from the subject’s stride-averaged joint angle profiles. A mixed, two-way factorial ANOVA tested for significant effects of group (between-subject factor: young, older nonfallers, and older fallers) and perturbation amplitude (within-subject factor: 0, 20, 35, and 50 cm) on each kinematic outcome measure using an α-level of 0.05. When a significant perturbation main effect was found, we focused our post hoc comparisons to elucidate differences between each amplitude and unperturbed walking.
Surface EMG data were recorded at 1,000 Hz using wireless electrodes (Trigno; Delsys, Boston, MA) from seven muscles on the right leg. Specifically, we recorded EMG from the tibialis anterior (TA), medial gastrocnemius (MG), soleus (SOL), peroneus longus (PL), vastus lateralis (VLAT), medial hamstrings (MH), and gluteus medius (GMED). All EMG data were high-pass filtered at 35 Hz (3rd-order Butterworth filter), demeaned, full-wave rectified, and then low-pass filtered at 10 Hz (3rd-order Butterworth filter). EMG data were normalized to the maximum observed in each muscle across all perturbation conditions and then resampled at each 0.5% of the gait cycle based on the heel-strikes identified from marker trajectories. Subject-specific EMG matrixes were then generated for each perturbation condition (four conditions: 0, 20, 35, and 50 cm) in which all gait cycles within a condition were concatenated end to end to form an m × t matrix, where m indicates the number of muscles (7) and t is the time base (no. of gait cycles × 201). The number of gait cycles varied across subjects and conditions, with a minimum size of 42 gait cycles.
Motor Module Extraction and Analysis
Motor modules for each subject were extracted separately from the EMG data matrixes for each perturbation condition using nonnegative matrix factorization (NMF; Lee and Seung 1999). NMF decomposes muscle activity into a reduced set of motor modules, such that EMG = W × C, where W is an m × n matrix with n motor modules and C is the n × t matrix of motor module activation coefficients. Each column of W represents the weights of each muscle in a module, and each row of C represents how that motor module is recruited across all gait cycles. To ensure equal weighting of each muscle during the extraction process, each row in the EMG data matrixes (i.e., muscle vector) was scaled to unit variance before motor module extraction with NMF and rescaled to original units afterwards (Torres-Oviedo and Ting 2007). We then examined several variables related to motor modules within and across groups as follows.
Motor module complexity and timing.
The number of motor modules, n, per condition was chosen as follows. From each data matrix, one to seven motor modules (W) were extracted. For each extraction, the goodness of fit between actual and reconstructed EMG (i.e., W × C) was evaluated with the variability accounted for (VAF), defined as 100 × squared uncentered Pearson’s correlation coefficient (Zar 1999). The 95% confidence interval (CI) on VAF was found by implementing a bootstrapping procedure in which the EMG data matrix was resampled 100 times with replacement. The VAF of the reconstructed EMG was recalculated for each resampling, and 95% CIs were constructed from these bootstrapped VAF values at each motor module number. The number of motor modules, n, was then chosen such that the lower bound of the 95% CI on VAF exceeded 90% (Fig. 2A; Cheung et al. 2009). As a complement to the number of motor modules, we also examined the VAF accounted for by one motor module (Fig. 2A; Shuman et al. 2016). To evaluate whether aging, a history of falls, or perturbation amplitude affected motor module complexity and timing (see below), we used a mixed factorial (group × perturbation) ANOVA and an α-level of 0.05. We found that neither measure of complexity changed with perturbation amplitude (see Fig. 4A). Therefore, all further analyses were performed using the motor modules identified during normal walking.
Fig. 2.

Motor modules identified from a representative young adult subject (n = 1). A: variability accounted for (VAF) plot from a representative subject illustrating the 2 measures of motor module complexity: number of motor modules and VAF by 1 motor module. B: the 4 motor modules identified in the representative subject (left) and their recruitment timing (right). Step-by-step variability was quantified by averaging the root mean square error (RMSE) in the timing curves across motor modules. C: the extracted motor modules well-reconstructed the original EMG (dashed line: original EMG, solid line: reconstructed EMG). TA, tibialis anterior; MG, medial gastrocnemius; SOL, soleus; PL, peroneus longus; VL, vastus lateralis; MG, medial hamstrings; GM, gluteus medius.
Fig. 4.
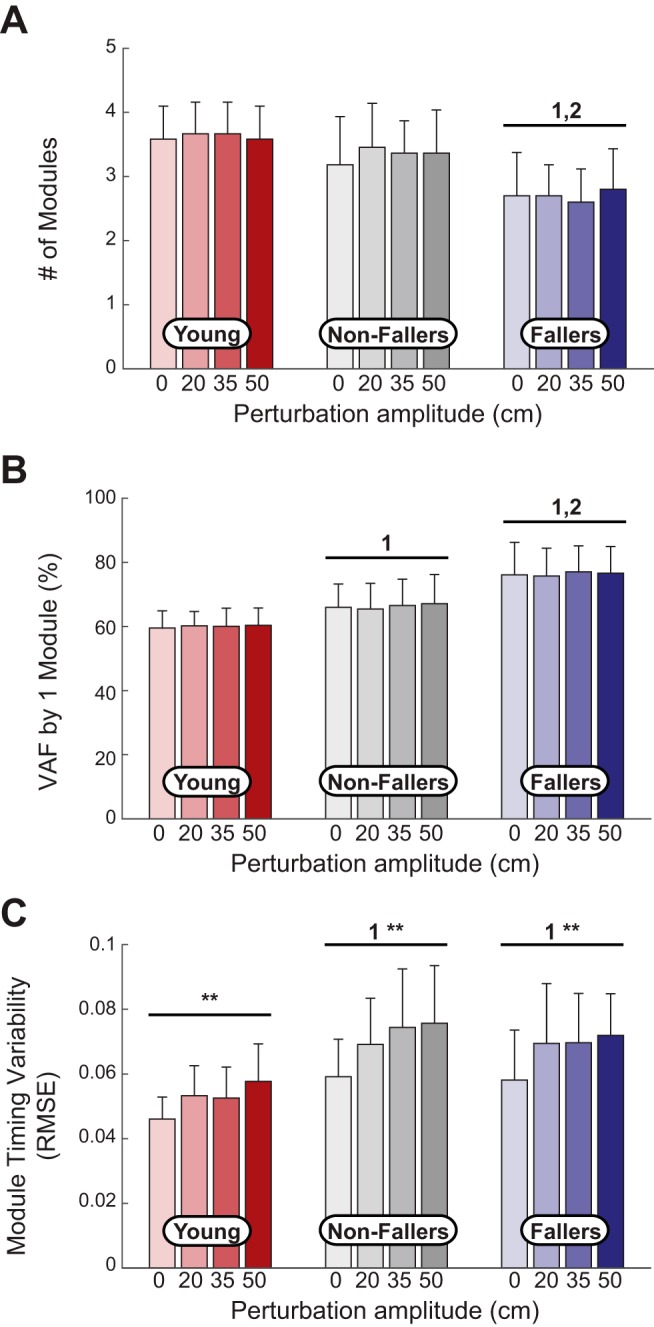
Motor module outcomes for each group across perturbation conditions. A: the number of motor modules were reduced in older fallers (n = 10) compared with young adults (n = 12) and older nonfallers (n = 11). There were no ANOVA main effects of perturbation magnitude in any group. B: the variability accounted for (VAF) explained by one motor module was increased in older fallers compared with both young adults and older nonfallers. There were no ANOVA main effects of perturbation magnitude in any group. C: older adults, regardless of a history of falls, exhibited larger step-by-step variability in motor module timing. All groups increased variability as perturbation amplitude was increased. RMSE, root mean square error. **Significant ANOVA main effect of perturbations. 1,2Significant difference from young adults and older nonfallers, respectively, via pairwise comparisons.
To examine the step-to-step variability of motor module recruitment timing, a new set of motor module recruitment coefficients, C, were calculated for the perturbation conditions using the Ws from the normal walking condition. Step-to-step variability was then quantified using the average root mean square error of the Cs for each perturbation condition (Fig. 2B; Sawers et al. 2015).
Motor module similarity across groups.
To examine whether the same motor modules were recruited across the young adult, older nonfallers, and older fallers groups, motor modules identified from each subject during normal walking within a group were pooled together and then sorted with hierarchical cluster analysis as in (Cheung et al. 2005). The Matlab statistics toolbox functions pdist (Minkowski, P = 3), linkage (ward), and cluster were applied to the pooled subjects from each group. The number of clusters for each group was determined as the minimum number of clusters such that no cluster contained more than one motor module from the same subject. We then compared the average motor modules in each of the young adult, older nonfallers, and older fallers clusters using Pearson’s correlation coefficient (r). A pair of motor modules was considered similar if r > 0.874, which corresponds to the critical value of r2 for 7 muscles at P = 0.01.
RESULTS
Differences in peak leg joint kinematics between groups were modest. Across all conditions, older fallers walked with comparable peak joint kinematics to older nonfallers but with less peak hip extension than young adults (P = 0.027; Fig. 3). Compared with normal walking, young adults walked only with less peak ankle extension in the presence of optical flow perturbations (P = 0.009; Fig. 3). The same amplitude perturbations elicited less peak hip extension and less peak hip adduction in older adults, regardless of a history of falls (P < 0.037). Those perturbations further elicited greater peak stance phase knee flexion and less peak ankle extension in older fallers compared with normal walking (P = 0.001).
Fig. 3.

Group mean hip, knee, and ankle joint angles in young (n = 12), older nonfallers (n = 11), and older fallers (n = 10) walking with and without optical flow perturbations plotted against an averaged gait cycle, from heel-strike to heel-strike. *Significant ANOVA main effect of perturbations. 1Significant difference compared with young adults via pairwise comparisons.
Modular Organization of Leg Muscle Activity During Normal Walking
We found that a history of falls (but not age) was associated with reduced motor module complexity. In normal walking, older fallers averaged fewer motor modules (2.7 ± 0.6) than both young adults (3.6 ± 0.5; P < 0.001) and older nonfallers (3.1 ± 0.8; P < 0.05; Fig. 4A). Similarly, the variance in leg muscle recruitment accounted for by one module was larger in older fallers compared with young adults (P < 0.01) and older nonfallers (P < 0.05; Fig. 4B).
Motor module composition and timing was very similar between young adults and older nonfallers (Fig. 5). A total of five motor modules were identified across the young adult participants, with module 5 only recruited in a single subject. The remaining four motor modules (modules 1–4) were recruited in the majority of both young adults and older nonfaller participants. Module 1 consisted primarily of the ankle plantarflexor muscles (MG, SOL and PL) and was active during late stance. Module 2 consisted primarily of the hamstrings (MH) and was active from late swing into early stance. Module 3 consisted of the ankle dorsiflexor (TA) and to some extent the knee extensor (VLAT) and was active primarily during leg swing. Finally, module 4 consisted mainly of extensor activity during early stance from hip extensors/abductors (GMED) and knee extensor (VLAT).
Fig. 5.

Motor modules recruited in each group during normal walking. In young adults (n = 12), a total of 5 motor modules were identified, with one module (module 5) only recruited in a single subject. The remaining 4 motor modules (modules 1–4) were recruited in at least 75% of young adult subjects, with qualitatively similar recruitment timing. These same four motor modules were also recruited in most of the older nonfallers (n = 11). In contrast, older fallers (n = 10) exhibited differences in motor module recruitment. Most older faller subjects recruited module 1 (plantarflexors during late stance). However, modules 2–4 were recruited in only half or fewer of older faller subjects, with module 3 [tibialis anterior (TA) during swing] recruited in only a single subject. Instead, many older faller subjects had a module with coactivation from all nonplantarflexor modules (module 6) that was active throughout much of stance. Text to the right of each module indicates the number of subjects within each group that recruited that motor module and its contribution to the overall variability accounted for (VAF) (means ± SD). MGAS, medial gastrocnemius; SOL, soleus; PERO, peroneus longus; VLAT, vastus lateralis; MH, medial hamstrings; GMED, gluteus medius.
In contrast, motor module composition differed in older adult fallers compared with nonfallers. The majority of older adult fallers (9 of 10) recruited module 1 (i.e., ankle plantarflexor module) with similar recruitment timing centered in late stance. However, only half of fallers recruited modules 2 and 4 (hamstring and proximal extensor modules, respectively), and only two fallers recruited module 3 (ankle dorsiflexor module). Instead, many fallers recruited a module resembling merging of modules 2, 3, and 4 with coactivation of TA, VLAT, MH, and GMED throughout much of the gait cycle (module 6).
We also found that age (but not a history of falls) was associated with increased step-to-step variability of module recruitment timing. Although the pattern of motor module recruitment timing across groups was qualitatively similar (Fig. 5, right), the step-to-step variability of module recruitment timing was on average 25% higher in older compared with younger adults regardless of a history of falls (Fig. 4C, P < 0.05). Indeed, we found no difference between older nonfallers and fallers for step-to-step variability of module recruitment timing.
Modular Organization of Leg Muscle Activity in the Presence of Perturbations
Neither measure of module complexity changed with perturbation amplitude (Fig. 4A), with the same motor modules that were recruited in normal walking (Fig. 5) also recruited in the presence of perturbations in all groups. Conversely, we found that step-to-step variability of recruitment timing was altered by optical perturbations during walking regardless of age or a history of falls. More specifically, although the average recruitment timing of motor modules was qualitatively similar across perturbation amplitudes (Fig. 6), step-to-step variability of module recruitment timing increased in the presence of perturbations in all groups (young adults: P = 0.02, older nonfallers: P = 0.04, and older fallers: P < 0.01).
Fig. 6.

Motor module recruitment timing across perturbation conditions. Recruitment timing was qualitatively similar across all perturbation magnitudes in each group (young: n = 12; older nonfallers: n = 11; and older fallers: n = 10).
DISCUSSION
This study investigated the complexity and consistency of motor module recruitment of leg muscles in subjects spanning a continuum of walking balance integrity (young, older nonfallers, and older fallers) during walking and changes thereof in response to balance challenges. Broadly speaking, we aimed to gain improved mechanistic insight into the neuromechanical mechanisms underlying age-associated falls risk. Although our findings generally supported our hypotheses, study outcomes were more complex than anticipated. Our first major finding was that a history of falls, but not aging per se, was associated with reduced motor module complexity. In addition, we found that age, independent of a history of falls, was associated with increased step-to-step variability of module recruitment timing. More surprisingly, module complexity was invariant in the presence of perturbations in all subject groups. Together, our findings suggest that a reduced motor repertoire in older adult fallers may be a constraint on their ability to appropriately respond to balance challenges during walking—novel evidence implicating altered neuromuscular control as a mechanism for age-associated falls risk.
Age and Fall History Effects on Motor Module Complexity During Unperturbed Walking
Our data suggest adults with a history of falls utilize a reduced motor repertoire to control leg muscle recruitment during normal, unperturbed walking. We acknowledge that our recorded muscles do not fully capture the control of all lower limb movements during walking. For example, compared with young adults, older adults often exhibit an increased reliance on hip flexor and extensor muscles (DeVita and Hortobagyi 2000), and we did not record from the gluteus maximus or iliopsoas in this study. Nevertheless, the four motor modules commonly recruited in our young and older adult nonfallers resembled those previously found during normal walking (Fig. 5) (Artoni et al. 2013; Cappellini et al. 2006; Clark et al. 2010; Davis and Vaughan 1993; Ivanenko et al. 2006; Monaco et al. 2010). In contrast, older adult fallers recruited a fewer number of motor modules than nonfallers, an outcome fully consistent with reduced modular complexity previously found in various populations known to be at a risk of falls [e.g., stroke (Clark et al. 2010), Parkinson’s disease (Rodriguez et al. 2013), spinal cord injury (Fox et al. 2013; Hayes et al. 2014), and cerebral palsy (Steele et al. 2015; Tang et al. 2015)]. However, this is the first study to demonstrate a reduced motor repertoire during walking in older adults with a history of falls but without any overt neurological deficits. Indeed, our subjects were otherwise seemingly well-matched, for example, for self-selected walking speed. Our results are also consistent with a recent paper investigating modular control in response to slip perturbations in which reduced complexity was identified in neurotypical adults who fell in response to an imposed slip (Sawers et al. 2017). Similarly, reduced modular complexity has been previously observed in neurotypical adults when their walking balance was challenged on a narrow beam compared with walking overground (Sawers et al. 2015). However, those paradigms were specifically designed to produce a “failure” or fall. In contrast, we find that altered leg muscle recruitment patterns in older adults with a history of falls are naturally emergent during normal walking. Given that only minor differences were observed in movement kinematics between groups, the altered modular patterns in older fallers cannot be attributed solely to differences in kinematics and thus likely represent a difference in neural control. Accordingly, these results provide evidence that reduced modular complexity (i.e., reduced motor repertoire) is highly prevalent in individuals with balance challenges even in the absence of overt neurological deficits.
The specific effects of a history of falls on leg muscle recruitment during walking included an absence and/or inability to independently recruit motor modules normally recruited to perform biomechanical functions important for walking balance control. The most striking result was that the independent TA module (Fig. 5, module 3) was absent in the majority of older adult fallers. This module is important for controlling foot and leg kinematics during the swing phase (Allen and Neptune 2012; Neptune et al. 2009), and its absence is consistent with prior work demonstrating that altered ankle dorsiflexor activity, strength, and range of motion in older adults are associated with a history of falls and predictor future falls (Kemoun et al. 2002; Whipple et al. 1987; Wolfson et al. 1995). Also absent in many subjects were modules 2 and 4, which are important for decelerating/controlling the swing leg from terminal swing into early stance and controlling vertical acceleration of the body during weight acceptance (i.e., early stance), respectively (Allen and Neptune 2012; Neptune et al. 2009). Instead, many subjects recruited a module that resembled a merging of modules 2, 3, and 4 with simultaneous activity of the TA, VLAT, MH, and GMED throughout much of stance and swing. Such modular merging is similar to that previously observed in individuals poststroke (Clark et al. 2010), which has been shown to result in an altered ability to control vertical support, mediolateral center of mass acceleration, and leg swing (Allen et al. 2013). In addition, GMED is important for regulating lateral foot placement and thus mediolateral balance control during walking (Rankin et al. 2014). Accordingly, our interpretation here in older adult fallers is that the inability to independently modulate these muscles during walking may contribute to an inability to effectively respond to balance perturbations in a task-specific manner.
There are at least two possible interpretations of our primary finding that older adult fallers have a reduced motor repertoire during normal, unperturbed walking; either the altered neural control strategy represents a neural deficit that leads to poor balance control or a prior history of falls results in a compensatory motor adaptation. Growing evidence suggests that even in older adults without any overt neurological deficits, there may be alterations in the peripheral and central neural control of walking that could precipitate a risk of falls. For example, compared with young adults, older adults often have reduced proprioceptive acuity and tactile sensation, functionally exacerbated by slowed nerve signal transmission (Dorfman and Bosley 1979; Goble et al. 2009). Indeed, those peripheral neural changes may explain the increased reliance on vision for balance control leveraged in our experimental paradigm (Sundermier et al. 1996). It is debated whether motor modules during locomotion reflect neural control mechanisms or emerge from biomechanical constraints. However, evidence from animal studies suggests that motor circuits related to motor modules exist at various levels of the nervous system, including the motor cortex (Overduin et al. 2012, 2015), brain stem (Chvatal et al. 2013; Roh et al. 2011), and spinal cord (Hart and Giszter 2010; Takei et al. 2017; Tresch et al. 1999). Thus, peripheral and/or central neural deficits due to aging could manifest functionally in walking as altered modular control of muscle activation. Future work will be required to tease out peripheral and central neural control contributions to altered modular control in older adult fallers. Nevertheless, the altered structure of the motor modules recruited by older fallers in this study is consistent with a muscle coordination mechanism that could impair balance control and/or lead to falls.
In contrast, the reduced motor repertoire observed in older falls may instead represent a goal-directed compensatory strategy after experiencing a fall, e.g., to increase stability in the presence of unreliable sensory feedback and/or to attenuate fear of falling. Although coactivation of muscles spanning the hip, knee, and ankle may have adverse effects on rapid, task-specific corrective responses to balance challenges, it may also increase mechanical impedance of the leg to abate instability. Antagonist coactivation, in particular, is commonly employed by young adults when walking on slippery surfaces (Whitmore et al. 2016) or walking down slopes (Hunter et al. 2010) and increases considerably in old age (Franz and Kram 2013; Hortobágyi et al. 2009; Peterson and Martin 2010). This feedforward control strategy is highly intuitive, as neural delays are often too long to allow feedback mechanisms to sufficiently respond to instabilities during movement. Although increased muscle coactivation is at least consistent with motor module merging, we only observed the latter in older adults with a prior history of falls. Accordingly, taking the alternative interpretation of our findings, older adult fallers may have purposefully and disproportionately coactivated their leg muscles, potentially due to a fear of falling. Indeed, heightened muscle coactivation is associated with fear of falling or perception of postural threat (Nagai et al. 2012).
Age and Fall History Effects on Motor Module Recruitment Timing During Unperturbed Walking
Despite no independent effect of age on motor module complexity, our results do suggest that age alone increases the variability of module recruitment timing during walking. Although older adult nonfallers recruited the same set of motor modules with similar recruitment timing as young adults, they did so with increased step-to-step variability. Some measures of variability, such as that of step width from one step to the next, have been implicated as a marker of falls risk in old age (Brach et al. 2008; Owings and Grabiner 2004). Increased modular recruitment variability is consistent with increased variability of gait patterns that often occur with aging (e.g., kinematics and spatiotemporal parameters), and some evidence suggests that this variability arises from increased sensory noise (Roos and Dingwell 2013). However, step-to-step variability of module recruitment timing, while greater compared with young adults, was not significantly different between older fallers and nonfallers. We interpret this outcome to suggest that larger variability of module recruitment timing is not associated with a history of falls in older adults and thus neither with their risk of falls, at least in the population studied here.
Motor Module Complexity and Timing in the Presence of Optical Flow Perturbations
Finally, regardless of its origin, we posit that the reduced motor repertoire in older adult fallers may constrain their ability to appropriately respond to balance challenges encountered during walking. Neither measure of module complexity was altered in the presence of optical flow perturbations, suggesting that fallers may not recruit the appropriate motor modules necessary for well-coordinated walking balance control even in the presence of perturbations. This outcome cannot be explained by a lack of effect of optical flow perturbations; indeed, such perturbations consistently destabilize walking balance, with larger effects due to aging and a history of falls (Franz et al. 2015; Qiao et al. 2018a, 2018b). Rather, this is consistent with prior work suggesting that module complexity is preserved across a variety of walking conditions with different task demands, even in stroke survivors with mobility deficits who exhibit a reduced motor repertoire (Routson et al. 2014). In that prior study, when faced with different walking task demands (e.g., changing speed, step length, and step height), individuals recruited the same motor modules as those during habitual walking. There only the timing of motor module recruitment changed to meet the additional task demands, and those who normally recruited fewer modules performed those tasks more poorly. In our study, we find a similar outcome; only motor module recruitment timing changed in the presence of perturbations designed to elicit the visual perception of falling. Specifically, we found that step-to-step variability of recruitment timing increased in response to increasing perturbation amplitude in all subjects. This outcome is highly consistent with an increased demand for corrective motor adjustments from one step to the next to preserve walking balance and prevent a fall. Interestingly, even when faced with a challenging balance task, older adult fallers continue to recruit fewer motor modules than older nonfallers and young adults. An exploratory post hoc analysis revealed that individuals with lower motor module complexity had increased local dynamic instability in the presence of optical flow perturbations but not during normal walking (Fig. 7). Thus the reduced peripheral motor repertoire in older adults with a history of falls may explain their disproportionate response to balance perturbations and could thus be considered a neuromuscular mechanism associated with a risk of falls.
Fig. 7.
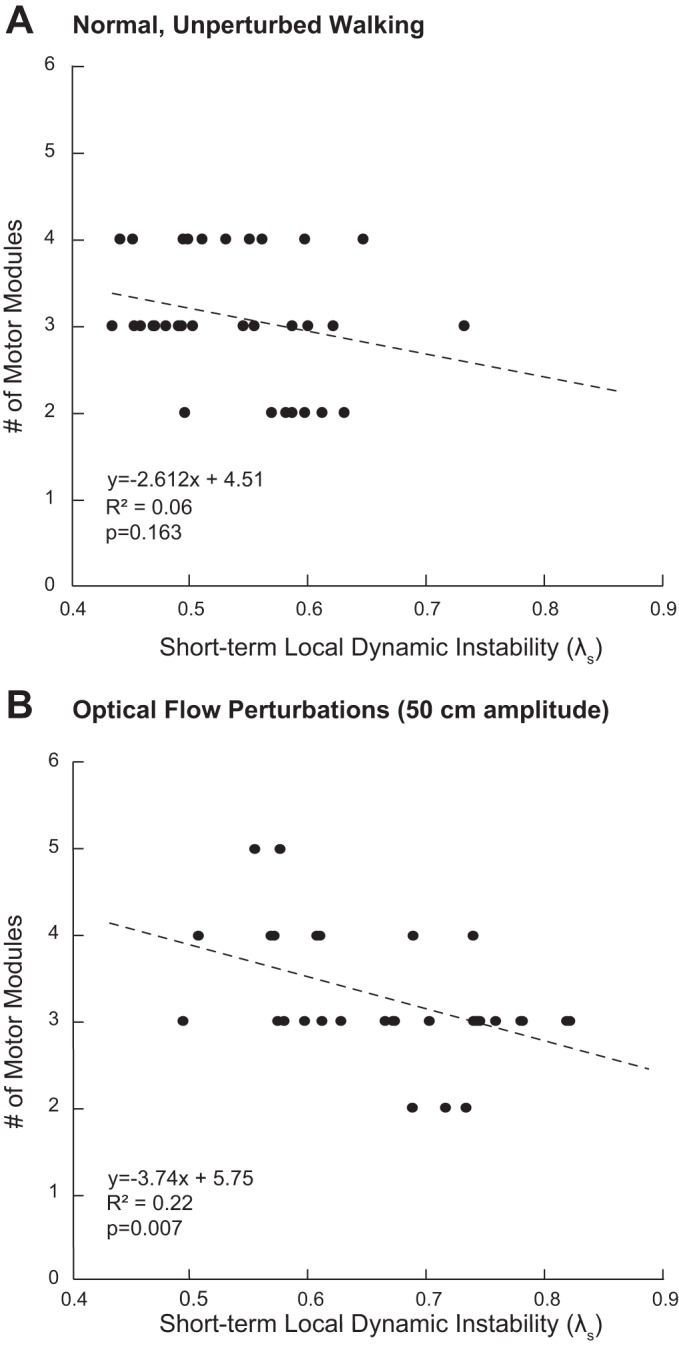
Results of an exploratory post hoc analysis of the association between motor module complexity (i.e., # of modules) and previously published values of local dynamic instability, quantified using short-term maximum divergence exponents (λs, 0–1 stride) (Qiao et al. 2018b). We refer the reader to the original manuscript for a detailed description of our data reduction and analysis procedures. Briefly, in that paper, we used a state space constructed from the 3-dimensional velocity of subject’s 7th cervical vertebrae and their time-delayed copies to compute maximum rates of divergence of initially neighboring trajectories. There, larger values of λs signify larger local dynamic instability. We concluded there that, compared with their effects in young adults, optical flow perturbations were capable of revealing independent effects of aging and a history of falls on gait instability that are not otherwise apparent during normal, unperturbed walking. Here, we performed 2 linear regressions across our study cohort: 1 during normal, unperturbed walking (A) and 1 in the presence of the largest amplitude optical flow perturbations used in this study (B; i.e., 50 cm). Our findings reveal that individuals with less motor module complexity in this study (namely, older adults with a history of falls) had larger local dynamic instability in the presence of optical flow perturbations (r2 = 0.22, P = 0.007) but not during normal, unperturbed walking (r2 = 0.06, P = 0.163). This outcome provides additional evidence that a reduced motor repertoire in older adult fallers may be a constraint on their ability to appropriately respond to balance challenges during walking: the major finding of our present study.
GRANTS
This work was supported by National Institutes of Health Grants R56-AG-054797, UL1-TR-001111, and F32-NS-087775 and University of North Carolina at Chapel Hill and North Carolina State University Closed-Loop Engineering for Advanced Rehabilitation (CLEAR) Core.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
J.F. conceived and designed research; J.F. performed experiments; J.L.A. and J.F. analyzed data; J.L.A. and J.F. interpreted results of experiments; J.L.A. and J.F. prepared figures; J.L.A. and J.F. drafted manuscript; J.L.A. and J.F. edited and revised manuscript; J.L.A. and J.F. approved final version of manuscript.
ACKNOWLEDGMENTS
Present address for J. Allen: Department of Chemical and Biomedical Engineering, West Virginia University, 1306 Evansdale Dr., PO Box 6102, Morgantown, WV 26506-6102.
REFERENCES
- Allen JL, Kautz SA, Neptune RR. The influence of merged muscle excitation modules on post-stroke hemiparetic walking performance. Clin Biomech (Bristol, Avon) 28: 697–704, 2013. doi: 10.1016/j.clinbiomech.2013.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen JL, McKay JL, Sawers A, Hackney ME, Ting LH. Increased neuromuscular consistency in gait and balance after partnered, dance-based rehabilitation in Parkinson’s disease. J Neurophysiol 118: 363–373, 2017. doi: 10.1152/jn.00813.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen JL, Neptune RR. Three-dimensional modular control of human walking. J Biomech 45: 2157–2163, 2012. doi: 10.1016/j.jbiomech.2012.05.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold EM, Ward SR, Lieber RL, Delp SL. A model of the lower limb for analysis of human movement. Ann Biomed Eng 38: 269–279, 2010. doi: 10.1007/s10439-009-9852-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Artoni F, Monaco V, Micera S. Selecting the best number of synergies in gait: preliminary results on young and elderly people. IEEE Int Conf Rehabil Robot 2013: 6650416, 2013. doi: 10.1109/ICORR.2013.6650416. [DOI] [PubMed] [Google Scholar]
- Berniker M, Jarc A, Bizzi E, Tresch MC. Simplified and effective motor control based on muscle synergies to exploit musculoskeletal dynamics. Proc Natl Acad Sci USA 106: 7601–7606, 2009. doi: 10.1073/pnas.0901512106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowden MG, Clark DJ, Kautz SA. Evaluation of abnormal synergy patterns poststroke: relationship of the Fugl-Meyer Assessment to hemiparetic locomotion. Neurorehabil Neural Repair 24: 328–337, 2010. doi: 10.1177/1545968309343215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brach JS, Studenski S, Perera S, VanSwearingen JM, Newman AB. Stance time and step width variability have unique contributing impairments in older persons. Gait Posture 27: 431–439, 2008. doi: 10.1016/j.gaitpost.2007.05.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bugnariu N, Fung J. Aging and selective sensorimotor strategies in the regulation of upright balance. J Neuroeng Rehabil 4: 19, 2007. doi: 10.1186/1743-0003-4-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cappellini G, Ivanenko YP, Poppele RE, Lacquaniti F. Motor patterns in human walking and running. J Neurophysiol 95: 3426–3437, 2006. doi: 10.1152/jn.00081.2006. [DOI] [PubMed] [Google Scholar]
- Cheung VC, d'Avella A, Tresch MC, Bizzi E. Central and sensory contributions to the activation and organization of muscle synergies during natural motor behaviors. J Neurosci 25: 6419–6434, 2005. doi: 10.1523/JNEUROSCI.4904-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheung VC, d'Avella A, Bizzi E. Adjustments of motor pattern for load compensation via modulated activations of muscle synergies during natural behaviors. J Neurophysiol 101: 1235–1257, 2009. doi: 10.1152/jn.01387.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chvatal SA, Macpherson JM, Torres-Oviedo G, Ting LH. Absence of postural muscle synergies for balance after spinal cord transection. J Neurophysiol 110: 1301–1310, 2013. doi: 10.1152/jn.00038.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chvatal SA, Torres-Oviedo G, Safavynia SA, Ting LH. Common muscle synergies for control of center of mass and force in nonstepping and stepping postural behaviors. J Neurophysiol 106: 999–1015, 2011. doi: 10.1152/jn.00549.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cigolle CT, Ha J, Min LC, Lee PG, Gure TR, Alexander NB, Blaum CS. The epidemiologic data on falls, 1998-2010: more older Americans report falling. JAMA Intern Med 175: 443–445, 2015. doi: 10.1001/jamainternmed.2014.7533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark DJ, Ting LH, Zajac FE, Neptune RR, Kautz SA. Merging of healthy motor modules predicts reduced locomotor performance and muscle coordination complexity post-stroke. J Neurophysiol 103: 844–857, 2010. doi: 10.1152/jn.00825.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- d’Avella A, Bizzi E. Shared and specific muscle synergies in natural motor behaviors. Proc Natl Acad Sci USA 102: 3076–3081, 2005. doi: 10.1073/pnas.0500199102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis BL, Vaughan CL. Phasic behavior of EMG signals during gait: use of multivariate statistics. J Electromyogr Kinesiol 3: 51–60, 1993. doi: 10.1016/1050-6411(93)90023-P. [DOI] [PubMed] [Google Scholar]
- DeVita P, Hortobagyi T. Age causes a redistribution of joint torques and powers during gait. J Appl Physiol (1985) 88: 1804–1811, 2000. doi: 10.1152/jappl.2000.88.5.1804. [DOI] [PubMed] [Google Scholar]
- Dorfman LJ, Bosley TM. Age-related changes in peripheral and central nerve conduction in man. Neurology 29: 38–44, 1979. doi: 10.1212/WNL.29.1.38. [DOI] [PubMed] [Google Scholar]
- Eikema DJ, Hatzitaki V, Konstantakos V, Papaxanthis C. Elderly adults delay proprioceptive reweighting during the anticipation of collision avoidance when standing. Neuroscience 234: 22–30, 2013. doi: 10.1016/j.neuroscience.2012.12.053. [DOI] [PubMed] [Google Scholar]
- Fox EJ, Tester NJ, Kautz SA, Howland DR, Clark DJ, Garvan C, Behrman AL. Modular control of varied locomotor tasks in children with incomplete spinal cord injuries. J Neurophysiol 110: 1415–1425, 2013. doi: 10.1152/jn.00676.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis CA, Franz JR, O’Connor SM, Thelen DG. Gait variability in healthy old adults is more affected by a visual perturbation than by a cognitive or narrow step placement demand. Gait Posture 42: 380–385, 2015. doi: 10.1016/j.gaitpost.2015.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franz JR, Francis CA, Allen MS, O’Connor SM, Thelen DG. Advanced age brings a greater reliance on visual feedback to maintain balance during walking. Hum Mov Sci 40: 381–392, 2015. doi: 10.1016/j.humov.2015.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franz JR, Kram R. How does age affect leg muscle activity/coactivity during uphill and downhill walking? Gait Posture 37: 378–384, 2013. doi: 10.1016/j.gaitpost.2012.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goble DJ, Coxon JP, Wenderoth N, Van Impe A, Swinnen SP. Proprioceptive sensibility in the elderly: degeneration, functional consequences and plastic-adaptive processes. Neurosci Biobehav Rev 33: 271–278, 2009. doi: 10.1016/j.neubiorev.2008.08.012. [DOI] [PubMed] [Google Scholar]
- Hart CB, Giszter SF. A neural basis for motor primitives in the spinal cord. J Neurosci 30: 1322–1336, 2010. doi: 10.1523/JNEUROSCI.5894-08.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes HB, Chvatal SA, French MA, Ting LH, Trumbower RD. Neuromuscular constraints on muscle coordination during overground walking in persons with chronic incomplete spinal cord injury. Clin Neurophysiol 125: 2024–2035, 2014. doi: 10.1016/j.clinph.2014.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hortobágyi T, Solnik S, Gruber A, Rider P, Steinweg K, Helseth J, DeVita P. Interaction between age and gait velocity in the amplitude and timing of antagonist muscle coactivation. Gait Posture 29: 558–564, 2009. doi: 10.1016/j.gaitpost.2008.12.007. [DOI] [PubMed] [Google Scholar]
- Hunter LC, Hendrix EC, Dean JC. The cost of walking downhill: is the preferred gait energetically optimal? J Biomech 43: 1910–1915, 2010. doi: 10.1016/j.jbiomech.2010.03.030. [DOI] [PubMed] [Google Scholar]
- Ivanenko YP, Poppele RE, Lacquaniti F. Motor control programs and walking. Neuroscientist 12: 339–348, 2006. doi: 10.1177/1073858406287987. [DOI] [PubMed] [Google Scholar]
- Jeka JJ, Allison LK, Kiemel T. The dynamics of visual reweighting in healthy and fall-prone older adults. J Mot Behav 42: 197–208, 2010. doi: 10.1080/00222895.2010.481693. [DOI] [PubMed] [Google Scholar]
- Kemoun G, Thoumie P, Boisson D, Guieu JD. Ankle dorsiflexion delay can predict falls in the elderly. J Rehabil Med 34: 278–283, 2002. doi: 10.1080/165019702760390374. [DOI] [PubMed] [Google Scholar]
- Lee DD, Seung HS. Learning the parts of objects by non-negative matrix factorization. Nature 401: 788–791, 1999. doi: 10.1038/44565. [DOI] [PubMed] [Google Scholar]
- Lord SR, Webster IW. Visual field dependence in elderly fallers and non-fallers. Int J Aging Hum Dev 31: 267–277, 1990. doi: 10.2190/38MH-2EF1-E36Q-75T2. [DOI] [PubMed] [Google Scholar]
- Marques NR, LaRoche DP, Hallal CZ, Crozara LF, Morcelli MH, Karuka AH, Navega MT, Gonçalves M. Association between energy cost of walking, muscle activation, and biomechanical parameters in older female fallers and non-fallers. Clin Biomech (Bristol, Avon) 28: 330–336, 2013. doi: 10.1016/j.clinbiomech.2013.01.004. [DOI] [PubMed] [Google Scholar]
- Melzer I, Benjuya N, Kaplanski J. Postural stability in the elderly: a comparison between fallers and non-fallers. Age Ageing 33: 602–607, 2004. doi: 10.1093/ageing/afh218. [DOI] [PubMed] [Google Scholar]
- Menz HB, Morris ME, Lord SR. Foot and ankle characteristics associated with impaired balance and functional ability in older people. J Gerontol A Biol Sci Med Sci 60: 1546–1552, 2005. doi: 10.1093/gerona/60.12.1546. [DOI] [PubMed] [Google Scholar]
- Monaco V, Ghionzoli A, Micera S. Age-related modifications of muscle synergies and spinal cord activity during locomotion. J Neurophysiol 104: 2092–2102, 2010. doi: 10.1152/jn.00525.2009. [DOI] [PubMed] [Google Scholar]
- Nagai K, Yamada M, Uemura K, Tanaka B, Mori S, Yamada Y, Aoyama T, Ichihashi N, Tsuboyama T. Effects of fear of falling on muscular coactivation during walking. Aging Clin Exp Res 24: 157–161, 2012. doi: 10.3275/7716. [DOI] [PubMed] [Google Scholar]
- Neptune RR, Clark DJ, Kautz SA. Modular control of human walking: a simulation study. J Biomech 42: 1282–1287, 2009. doi: 10.1016/j.jbiomech.2009.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Connor KW, Loughlin PJ, Redfern MS, Sparto PJ. Postural adaptations to repeated optic flow stimulation in older adults. Gait Posture 28: 385–391, 2008. doi: 10.1016/j.gaitpost.2008.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Connor SM. Using MATLAB, Virtual Reality, and a Treadmill To Investigate how Humans Use Visual Input To Control Their Gait (Online) MathWorks, 2012. https://www.mathworks.com/company/newsletters/articles/using-matlab-virtual-reality-and-a-treadmill-to-investigate-how-humans-use-visual-input-to-control-their-gait.html. [Google Scholar]
- Overduin SA, d’Avella A, Carmena JM, Bizzi E. Microstimulation activates a handful of muscle synergies. Neuron 76: 1071–1077, 2012. doi: 10.1016/j.neuron.2012.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Overduin SA, d’Avella A, Roh J, Carmena JM, Bizzi E. Representation of muscle synergies in the primate brain. J Neurosci 35: 12615–12624, 2015. doi: 10.1523/JNEUROSCI.4302-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owings TM, Grabiner MD. Step width variability, but not step length variability or step time variability, discriminates gait of healthy young and older adults during treadmill locomotion. J Biomech 37: 935–938, 2004. doi: 10.1016/j.jbiomech.2003.11.012. [DOI] [PubMed] [Google Scholar]
- Peterson DS, Martin PE. Effects of age and walking speed on coactivation and cost of walking in healthy adults. Gait Posture 31: 355–359, 2010. doi: 10.1016/j.gaitpost.2009.12.005. [DOI] [PubMed] [Google Scholar]
- Piazza SJ, Okita N, Cavanagh PR. Accuracy of the functional method of hip joint center location: effects of limited motion and varied implementation. J Biomech 34: 967–973, 2001. doi: 10.1016/S0021-9290(01)00052-5. [DOI] [PubMed] [Google Scholar]
- Qiao M, Feld JA, Franz JR. Aging effects on leg joint variability during walking with balance perturbations. Gait Posture 62: 27–33, 2018a. doi: 10.1016/j.gaitpost.2018.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiao M, Truong KN, Franz JR. Does local dynamic stability during unperturbed walking predict the response to balance perturbations? An examination across age and falls history. Gait Posture 62: 80–85, 2018b. doi: 10.1016/j.gaitpost.2018.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rankin BL, Buffo SK, Dean JC. A neuromechanical strategy for mediolateral foot placement in walking humans. J Neurophysiol 112: 374–383, 2014. doi: 10.1152/jn.00138.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez KL, Roemmich RT, Cam B, Fregly BJ, Hass CJ. Persons with Parkinson’s disease exhibit decreased neuromuscular complexity during gait. Clin Neurophysiol 124: 1390–1397, 2013. doi: 10.1016/j.clinph.2013.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roh J, Cheung VC, Bizzi E. Modules in the brain stem and spinal cord underlying motor behaviors. J Neurophysiol 106: 1363–1378, 2011. doi: 10.1152/jn.00842.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roos PE, Dingwell JB. Using dynamic walking models to identify factors that contribute to increased risk of falling in older adults. Hum Mov Sci 32: 984–996, 2013. doi: 10.1016/j.humov.2013.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Routson RL, Clark DJ, Bowden MG, Kautz SA, Neptune RR. The influence of locomotor rehabilitation on module quality and post-stroke hemiparetic walking performance. Gait Posture 38: 511–517, 2013. doi: 10.1016/j.gaitpost.2013.01.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Routson RL, Kautz SA, Neptune RR. Modular organization across changing task demands in healthy and poststroke gait. Physiol Rep 2: e12055, 2014. doi: 10.14814/phy2.12055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawers A, Allen JL, Ting LH. Long-term training modifies the modular structure and organization of walking balance control. J Neurophysiol 114: 3359–3373, 2015. doi: 10.1152/jn.00758.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawers A, Pai YC, Bhatt T, Ting LH. Neuromuscular responses differ between slip-induced falls and recoveries in older adults. J Neurophysiol 117: 509–522, 2017. doi: 10.1152/jn.00699.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shuman B, Goudriaan M, Bar-On L, Schwartz MH, Desloovere K, Steele KM. Repeatability of muscle synergies within and between days for typically developing children and children with cerebral palsy. Gait Posture 45: 127–132, 2016. doi: 10.1016/j.gaitpost.2016.01.011. [DOI] [PubMed] [Google Scholar]
- Springer S, Giladi N, Peretz C, Yogev G, Simon ES, Hausdorff JM. Dual-tasking effects on gait variability: the role of aging, falls, and executive function. Mov Disord 21: 950–957, 2006. doi: 10.1002/mds.20848. [DOI] [PubMed] [Google Scholar]
- Steele KM, Rozumalski A, Schwartz MH. Muscle synergies and complexity of neuromuscular control during gait in cerebral palsy. Dev Med Child Neurol 57: 1176–1182, 2015. doi: 10.1111/dmcn.12826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stokes HE, Thompson JD, Franz JR. The neuromuscular origins of kinematic variability during perturbed walking. Sci Rep 7: 808, 2017. doi: 10.1038/s41598-017-00942-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sundermier L, Woollacott MH, Jensen JL, Moore S. Postural sensitivity to visual flow in aging adults with and without balance problems. J Gerontol A Biol Sci Med Sci 51A: M45–M52, 1996. doi: 10.1093/gerona/51A.2.M45. [DOI] [PubMed] [Google Scholar]
- Takei T, Confais J, Tomatsu S, Oya T, Seki K. Neural basis for hand muscle synergies in the primate spinal cord. Proc Natl Acad Sci USA 114: 8643–8648, 2017. doi: 10.1073/pnas.1704328114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang L, Li F, Cao S, Zhang X, Wu D, Chen X. Muscle synergy analysis in children with cerebral palsy. J Neural Eng 12: 046017, 2015. doi: 10.1088/1741-2560/12/4/046017. [DOI] [PubMed] [Google Scholar]
- Thelen DG, Schultz AB, Alexander NB, Ashton-Miller JA. Effects of age on rapid ankle torque development. J Gerontol A Biol Sci Med Sci 51A: M226–M232, 1996. doi: 10.1093/gerona/51A.5.M226. [DOI] [PubMed] [Google Scholar]
- Tinetti ME, Speechley M, Ginter SF. Risk factors for falls among elderly persons living in the community. N Engl J Med 319: 1701–1707, 1988. doi: 10.1056/NEJM198812293192604. [DOI] [PubMed] [Google Scholar]
- Ting LH, Macpherson JM. A limited set of muscle synergies for force control during a postural task. J Neurophysiol 93: 609–613, 2005. doi: 10.1152/jn.00681.2004. [DOI] [PubMed] [Google Scholar]
- Torres-Oviedo G, Ting LH. Muscle synergies characterizing human postural responses. J Neurophysiol 98: 2144–2156, 2007. doi: 10.1152/jn.01360.2006. [DOI] [PubMed] [Google Scholar]
- Tresch MC, Saltiel P, Bizzi E. The construction of movement by the spinal cord. Nat Neurosci 2: 162–167, 1999. doi: 10.1038/5721. [DOI] [PubMed] [Google Scholar]
- Whipple RH, Wolfson LI, Amerman PM. The relationship of knee and ankle weakness to falls in nursing home residents: an isokinetic study. J Am Geriatr Soc 35: 13–20, 1987. doi: 10.1111/j.1532-5415.1987.tb01313.x. [DOI] [PubMed] [Google Scholar]
- Whitmore MW, Hargrove LJ, Perreault EJ. Gait characteristics when walking on different slippery walkways. IEEE Trans Biomed Eng 63: 228–239, 2016. doi: 10.1109/TBME.2015.2497659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfson L, Judge J, Whipple R, King M. Strength is a major factor in balance, gait, and the occurrence of falls. J Gerontol A Biol Sci Med Sci 50: 64–67, 1995. [DOI] [PubMed] [Google Scholar]
- Yeh TT, Cluff T, Balasubramaniam R. Visual reliance for balance control in older adults persists when visual information is disrupted by artificial feedback delays. PLoS One 9: e91554, 2014. doi: 10.1371/journal.pone.0091554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zar JH. Biostatistical Analysis. Upper Saddle River, NJ: Prentice-Hall, 1999. [Google Scholar]
- Zeni JA Jr, Richards JG, Higginson JS. Two simple methods for determining gait events during treadmill and overground walking using kinematic data. Gait Posture 27: 710–714, 2008. doi: 10.1016/j.gaitpost.2007.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]