

Abstract
This study summarizes the response of a hot spring cyanobacterium Fischerella sp. strain HKAR-14, under simulated light conditions of ultraviolet radiation (UVR), photosynthetically active radiation (PAR), PAR + UV-A (PA) and PAR + UV-A + UV-B (PAB). Exposure to UVR caused a decline in growth and Chl a while total carotene content increased under PA and PAB. Maximum photochemical efficiency of photosystem II (Fv/Fm) and relative electron transport rate decreased significantly in PA and PAB exposure. Higher non-photochemical quenching and lower photochemical quenching values were observed in UVR-exposed samples as compared to the control. Levels of intracellular reactive oxygen species (ROS) increased significantly in PAB and PA. Fluorescence microscopic images showed an increase in green fluorescence, indicating the generation of ROS in UVR. The antioxidant machinery including superoxide dismutase, catalase and peroxidase showed an increase of 1.76-fold and 2.5-fold superoxide dismutase, 2.4-fold and 3.7-fold catalase, 1.83-fold and 2.5-fold peroxidase activities under PA and PAB, respectively. High-performance liquid chromatography equipped with photodiode array detector, electrospray ionization mass spectrometry, Fourier-transform infrared spectroscopy and nuclear magnetic resonance spectroscopy analyses reveal the occurrence of a single mycosporine-like amino acid, shinorine (λmax 332.3 ± 2 nm, m/z 333.1), with a retention time of 1.157 min. The electrochemical characterization of shinorine was determined by cyclic voltammetry. The shinorine molecule possesses electrochemical activity and represents diffusion-controlled process in 0.1 M (pH 7.0) phosphate buffer. An antioxidant assay of shinorine showed its efficient activity as antioxidant which increased in a dose-dependent manner.
Keywords: Cyanobacteria, Chlorophyll fluorescence, Mycosporine-like amino acids, Antioxidant, Ultraviolet radiation
Introduction
Cyanobacteria represent a unique assemblage of prokaryotes with chlorophyll (Chl) a that are able to perform photosynthesis-dependent oxygen evolution with water as source of protons, similar to eukaryotic algae and higher plants. These organisms led to the evolution of aerobic life on this planet (Nelson and Ben-Shem 2005; Fischer 2008). They are outstanding for their role in global carbon dioxide and nitrogen fixation (Rajneesh et al. 2017; Pathak et al. 2018). Cyanobacteria constitute a crucial microflora with respect to total biomass and productivity in aquatic as well as terrestrial ecosystems (Whitton 2012; Pathak et al. 2018). They can colonize a wide range of habitats including hot deserts, ice shelves, aquatic systems, dry rocks and soil. They are found in Arctic and Antarctic lakes, hot springs and occur as endosymbionts in several protists, lichens and plants (Vincent and Quesada 1994; Thajuddin and Subramanian 2005; Singh 2018). Cyanobacteria have a great potential for biotechnological applications as food and fuel, biofertilizers in paddy fields, mariculture and biomedicals (Singh et al. 2016). Besides their important role in biotechnology, they have a wide application in photosynthetic research as model organisms, partly due to their prokaryotic nature (Rastogi and Sinha 2009; Olsson-Francis et al. 2013; Ogawa et al. 2017).
Climate changes have strong interactions with stratospheric ozone depletion, caused primarily by an increase in greenhouse gas concentrations, which results in increased ultraviolet radiation (UVR) that adversely affects aquatic ecosystems (Häder et al. 2015). Due to the Montreal protocol, the level of chlorofluorocarbons (CFCs) has decreased, but the full recovery of the ozone layer is not expected within this century (Burkholder et al. 2015). As per the reports of national aeronautics and space administration (NASA) and national oceanic and atmospheric administration (NOAA) satellite data, in the years having normal weather conditions in early October or late September, the ozone hole grows to a maximum area of about 8 million square miles. But significant reduction of the ozone hole has been reported in September and October recently by NASA and NOAA scientists due to abnormal weather patterns in the upper atmosphere over Antarctica resulting in the smallest ozone hole observed since 1982 (https://www.nasa.gov/feature/goddard/2019/2019-ozone-hole-is-the-smallest-on-record-since-its-discovery). Nitrogen fixation and photosynthesis are energy-dependent, ecologically important processes which are based on harvesting solar radiation (Rajneesh et al. 2017). The cyanobacterial phycobilisome (PBS) (light-harvesting complex) collects photosynthetically active radiation (PAR; 400–700 nm) of the solar spectrum to fuel both photosystem (PS) I and PSII in cyanobacteria (Gutu and Kehoe 2012). Depending upon the ambient light quality, cyanobacteria can adjust their pigment composition in the PBS which is ecologically a very important phenomenon called complementary chromatic adaptation (Kehoe and Gutu 2006; Gutu and Kehoe 2012). Due to the obligate requirement of solar radiation for maintenance of normal biochemistry and physiology of the cell, cyanobacteria are often exposed to lethal doses of UVR (280–400 nm) that harms their photosynthetic machinery (Rastogi et al. 2014).
UVR, especially UV-B (280–315 nm) damages cyanobacteria in various ways. Direct effects include DNA damage by thymine dimer formation (Klisch et al. 2005), lipid peroxidation, DNA strand breaks and impairment of orientation and motility (He et al. 2002). These modifications are due to the production of free radicals or reactive oxygen species (ROS), such as hydroxyl radicals (HO·), hydrogen peroxide (H2O2), hydroperoxyl radicals (HO2·), and superoxide anion radicals (O2−) (He and Häder 2002a; Rastogi et al. 2010a, 2011; Lesser 2011; Rajneesh et al. 2019). In photosynthetic organisms, PSII is a prominent target of UVR (Barbato et al. 2000). UV-A radiation indirectly affects cells by photo-sensitizing reactions in the presence of Chl and phycobilins (He and Häder 2002b; Hargreaves et al. 2007; Thapar et al. 2008). To counteract the damaging effect of UVR, cyanobacteria have developed several lines of defense mechanisms (Richa and Sinha 2015; Pathak et al. 2017, 2019a, b; Rajneesh et al. 2019). Cyanobacteria adopt three different adjustment strategies to overcome UVR damage: avoidance, repair and protection. For instance, to protect themselves, they can synthesize quenching agents and UV-screening substances, or migrate to escape damage caused by UV-B radiation. They repair DNA strand breaks and damage of the photosynthetic apparatus induced by UV-B radiation (Garcia-Pichel and Belnap 1996; Quesada and Vincent 1997).
UVR can damage PSII directly by photoinactivating the oxygen-evolving complex or the D1 protein (Tyystjärvi 2008; Singh et al. 2010a). Several studies have established the fact that photosynthetic components are damaged by UVR, but limited reports are available on the UVR-induced effects on photosynthetic performance of cyanobacteria. Biosynthesis of photoprotective compounds, such as mycosporine-like amino acids (MAAs) and scytonemin, is one of the several defense strategies evolved by cyanobacteria to deal with the harmful effects of solar UVR (Garcia-Pichel et al. 1993; Sinha et al. 1999, 2007; Singh et al. 2013a, 2017; Pathak et al. 2019b). MAAs act as prominent UV-protectants that provide protection against lethal UVR (Garcia-Pichel et al. 1993; Richa and Sinha 2015; Wada et al. 2015). MAAs are a family of over 30 known, colorless (< 400 Da), polar compounds characterized by a cyclohexenone or cyclohexenimine ring which is covalently linked to the N substituent of an amino acid or its imino alcohol (Pope et al. 2015; Ahmed et al. 2019). These compounds absorb maximally at wavelengths between 310–362 nm (ɛ = 28,100–50,000 M−1 cm−1) and dissipate the absorbed energy in the form of heat without generation of ROS (Conde et al. 2000).
Several studies have demonstrated that consequences of UV-B for photosynthetic organisms are negative and extremely variable, including processes from molecular to ecophysiological scales (Bischof et al. 2006; Barufi et al. 2011). Some investigations showed that UV-A and UV-B radiations, though mostly being deleterious (Xu and Gao 2010; Roleda et al. 2012), likewise appeared to have some positive effects in alleviating photoinhibition or promoting the recovery of inhibited photosynthesis (Hanelt and Roleda 2009; Xu and Gao 2010). An effective bioindicator to evaluate the physiological status of cyanobacteria under UVR is in vivo Chl a fluorescence. Maximum quantum yield and electron transport rate are used as physiological stress indicators in algae (Figueroa et al. 2003).
In this study, we have analyzed the variation in photosynthetic performance as well as the presence of antioxidative enzymes and MAA induction caused by PAR and UVR in the hot spring cyanobacterium Fischerella sp. strain HKAR-14 to nullify the effects of UVR. Photosynthetic performance of the cyanobacterium was studied by monitoring photosynthetic parameters, such as photosynthetic pigment content, maximal quantum yield of dark-acclimated samples (optimum quantum yield: Fv/Fm), relative electron transport rate (rETR), non-photochemical quenching (NPQ) and photochemical quenching (qP), as determined by pulse amplitude modulated fluorescence (PAM) measurements. In vivo detection of ROS was done using 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA). Antioxidative enzymes, such as catalase (CAT), superoxide dismutase (SOD) and peroxidase (POD), were determined during exposure of the organisms to UVR. Apart from enzymatic defense mechanisms, induction of UV-screening compounds (MAAs) has been studied to test the non-enzymatic protection strategy of the cyanobacterium. The purified MAA was biochemically characterized through analytical techniques, such as ultraviolet–visible (UV–VIS) absorption spectroscopy, high-performance liquid chromatography equipped with photodiode array detector (HPLC–PDA), electrospray ionization mass spectrometry (ESI–MS), Fourier-transform infrared (FTIR) spectroscopy and nuclear magnetic resonance (NMR) spectroscopy. Electrochemical characterization of purified MAA was also carried out. The in vitro antioxidative potential of the purified MAA was determined using the 2,2-diphenyl-1-picryl-hydrazyl (DPPH) radical scavenging assay, ferric reducing antioxidant power (FRAP) assay, superoxide radical scavenging activity and 2,2-Azinobis (3-ethylbenzothiazoline 6-sulfonate) (ABTS) radical scavenging activity.
Materials and methods
Experimental organism and growth conditions
The cyanobacterium Fischerella sp. strain HKAR-14 was isolated from Chandra Kund, Rajgir, Bihar, India. The morphology of the cyanobacterium was studied using a light microscope (Olympus CX21i, USA). Fischerella sp. strain HKAR-14 was cultured by following standard microbiological methods (Sinha et al. 1995). Cultures were routinely grown photoautotrophically and axenically in an autoclaved BG-11 liquid medium (Rippka et al. 1979) without any N supplement at 20 ± 2 °C. Cultures were illuminated with fluorescent tubes at a photon flux density of 55.08 ± 9.18 μmol m−2 s−1 at the surface of the vessels with 14 h light followed by 10 h darkness. To avoid contamination, cultures were sub-cultured at regular intervals and hand-shaken five times daily. Logarithmically or exponentially growing (14 days) cultures were utilized for performing all experiments.
Sequencing of rRNA genes and phylogenetic analyses
DNA fragments (PCR-amplified) were sequenced in an ABI 3130 Genetic Analyzer (ABI, USA) using a Big Dye Terminator Cycle sequencing kit (version 3.1) and an Applied Biosystems automated DNA sequencer. The conditions for sequencing reactions were 25 cycle of 96 °C for 1 min (initial denaturation), 96 °C for 10 s (denaturation), 50 °C for 5 s (hybridization) and 60 °C for 4 min (elongation). Data analysis software (Seq Scape, version 5.2) was used for editing the electropherograms. Alignment of the sequences of 16S rRNA gene fragments against known sequences present in the GenBank database was done using the basic local alignment search tool.
(BLAST) program of national center for biotechnology information (NCBI) search (Altschul et al. 1990). CLUSTALW was used for producing multiple alignments. The 16S rRNA gene sequences of the cyanobacterium were classified into phylogenetic groups as proposed by Desikachary (Desikachary1959) to determine the genetic variability between and within the groups. A phylogenetic tree was constructed using the neighbor-joining algorithm (Saitou and Nei 1987) provided in MEGA 7 software (Kumar et al. 2016).
Source and mode of PAR and UV irradiation
The cells of exponentially growing Fischerella sp. strain HKAR-14 were transferred into sterile transparent Petri dishes (75 mm) and placed on a rotary shaker for uniform exposure of 1 Wm−2 UV-B (Philips Ultraviolet-B TL 15 W:12, Holland), 3.5 Wm−2 UV-A (Philips Ultraviolet-B TL 15 W:12, Holland) and PAR (55.08 ± 9.18 μmol m−2 s−1) (OSRAM L 36 W: 32 Lumilux de luxe warm white and Radium NL 36 W: 26 Universal white, Germany) for 72 h at a temperature of about 20 ± 2 °C to avoid excess heating of cells. Different cut-off filter foils, such as 395 nm and 295 (Ultraphan, Digefra, Munich, Germany) and 320 nm (Folex, Dreieich, Munich, Germany), were used for covering the Petri dishes to obtain transmittance of desired wavebands of UV-B (280–315 nm), UV-A (315–400 nm) radiation and photosynthetically active radiation (PAR) (400–700 nm), respectively. Subsequently, at regular time intervals of 0 (control), 12, 24, 36, 48, 60, and 72 h, equal amounts of samples were withdrawn from each filter treatment in triplicates for the analysis.
Determination of pigment contents
For determination of pigment contents, samples were homogenized in 100% methanol. Homogenized samples were incubated overnight at 4 °C followed by centrifugation at 10,000 × g for 10 min. Absorption spectra of supernatants were measured in the wavelength range of 200–700 nm in a double beam spectrophotometer (U-2910, UV/VIS, Hitachi, Tokyo, Japan) using quartz cuvettes. Chl a showed maximum absorbance at 666 nm and total carotenes at 470 nm. Chl a and total carotene contents were determined by the methods as described by Şükran et al. (1998).
where Ca = Chl a, Cb = Chl b, Cx+c = Total carotene and Cb = 0 (In cyanobacteria Chl b is absent).
Photosynthetic performance as in vivo chlorophyll fluorescence measurements
The photosynthetic performance of Fischerella sp. strain HKAR-14 under PAR, PA and PAB irradiation conditions was measured using a pulse-amplitude-modulation (PAM) fluorometer (PAM-2500, Walz, Effeltrich, Germany) after 0, 12, 24, 36, 48 and 72 h exposure. To monitor fluorescence parameters, such as Fv/Fm, NPQ, qP and relative electron transport rate (rETR), samples (2 mL) were dark-adapted for 15 min at room temperature. The dark-adapted samples were exposed to a modulated light beam with an intensity of 3–4 μmol photons m−2 s−1 to measure the initial fluorescence (F0). This was followed by an exposure to a saturating pulse of white light of 3000 μmol photons m−2 s−1 to induce the maximum fluorescence (Fm). Variable fluorescence (Fv) was determined by Fv = Fm − F0, and the optimum quantum yield (Fv/Fm) was calculated as follows:
where Fv/Fm represents the maximal quantum yield of the photosynthetic apparatus.
The effect of UVR was assessed by comparing the inhibition of Fv/Fm that was calculated as the percentage of decrease between the values measured in PAR + UV-A (PA) and PAR + UV-A + UV-B (PAB) and values measured in samples exposed only to PAR. The relative UV inhibition of PSII was calculated based on the following equation:
where PAR, PA and PAB represent Fv/Fm under respective treatments. Relative UV inhibition was only calculated when PAR, PA and PAB were significantly different. Then, the fluorescence parameters, including effective fluorescence parameters [effective quantum yield (ɸPSII), rETR, NPQ and qP], were automatically determined under a series of PAR gradients exposing the samples to ten light steps (each 30 s) of actinic light (0, 5, 34, 104, 201, 366, 622, 984, 1389, 2018, 2973 μmol photons m−2 s−1) and recorded by the pre-installed software routine.
Relative electron transport rate (rETR) was estimated according to Rascher et al. (2003)
where ɸPSII = effective quantum yield, 0.5 is a factor that accounts for the energy partitioning between PSII and PSI (PSI/PSII allocation factor). The absorption of the cell suspension was not calculated in this study and the light absorption fraction, 0.84 (generally used for leaves) was not included in the ETR calculations. The term rETR was therefore used.
NPQ and qP quenching coefficients were calculated based on a built-in functions of the PAM and the following equations:
The qP coefficient quantifies the actual fraction of the open PSII reaction centers. It reflects the balance between excitation of PSII centers that close them and the removal of electrons from PSII by the electron transport chain that reopens the centers.
Determination of antioxidant enzyme activity
For assaying antioxidant enzymes, cultures were exposed to PAR, PA and PAB radiations for desired time periods. The antioxidative enzymes were extracted by method described earlier (Tripathi and Srivastava 2001) with slight modifications. The irradiated cells (30 mL) were harvested by centrifugation (12,000 × g for 20 min at 4 °C) and the pellets were homogenized in extraction buffer containing 50 mM potassium phosphate buffer (pH 7.5), 1 mM ethylenediaminetetracetic acid (EDTA), 1% (w/v) polyvinyl polypyrollidone (PVP) and 2.5 mM phenylmethane sulfonyl fluoride (PMSF). An ultrasonicator (Sonic and Materials, USA) was used for sonicating the resulting suspension which was subsequently centrifuged at 10,000 × g for 30 min at 4 °C and supernatants were used for determination of the antioxidant enzymes activity of CAT (EC 1.11.1.6), SOD (EC 1.15.1.1) and POD (EC 1.11.1.7). Inhibition in reduction of nitro blue tetrazolium (NBT) was monitored for measuring the total activity of SOD (Fridovich 1974; Beyer and Fridovich 1987). CAT activity was determined by calculating the H2O2 absorbance at 240 nm (ε = 39.4 mM−1 cm−1) after 1 min incubation in darkness (Aebi 1984). The quantification of H2O2 was carried out by the Amplex Red assay (Invitrogen, Eugene, OR, USA) as described earlier (Halliwell 2006). For determining POD activity, absorbance at 420 nm was measured at every 20 s for 2 min. Reaction mixtures contained 0.1 M phosphate buffer pH 6.0, 5.33% pyrogallol solution (M/V), 3% H2O2 as substrate and 100 µL of enzyme extract. One unit of POD activity is defined as the amount of enzyme required to catalyze the production of 1 mg of purpurogallin from pyrogallol in 20 s at 20 °C under assay conditions (Maehly and Chance 1955).
In vivo detection of ROS using 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA)
Detection of ROS production was done by DCFH-DA after exposing the culture to PAR, PA and PAB. 5 μM (final concentration) of DCFH-DA solubilized in ethanol was added to the cultures and the samples were incubated at room temperature in the dark for 1 h on a shaker. UV irradiation quickly hydrolyzes DCFH or auto-oxidizes DCFH-DA (Burow and Valet 1987; He and Häder 2002b); hence, DCFH-DA is added only after irradiation. After 1 h of incubation, Fischerella sp. strain HKAR-14 was subjected to fluorescence microscopy (Nikon Eclipse 90I, Japan) and fluorescence spectrophotometric analysis (Agilent Technologies, Cary Eclipse, USA) with an excitation wavelength of 485 nm and an emission band at 525 nm (He and Häder 2002b; Rajneesh et al. 2017, 2019). To reduce the damage of cells, the exposure time was limited to 600 ms. After different durations of stress exposure, fluorescence was measured in terms of emitted fluorescence intensity.
MAA extraction and spectroscopic analysis
Extraction of the UV-absorbing compound MAA was performed as per method utilized by Rastogi et al. (2012). In brief, cyanobacterial cells were harvested by centrifugation (Mikro 220R, Hettich, Germany) and pellets were resuspended in 100% methanol (HPLC grade), incubated overnight under dark conditions at 4 °C followed by homogenization. The aliquots were then centrifuged (5000 × g, 5 min) and supernatants were transferred to new microtubes and subjected to spectroscopic analysis between 250–700 nm using a UV–VIS spectrophotometer (U-2910, 2J1-0012, Hitachi, Tokyo, Japan). Analysis of the raw spectra (peaks) was done using UV Probe version software (Shimadzu Corp., Kyoto, Japan). The obtained supernatant (methanolic extracts) was evaporated at 40 °C in a vacuum evaporator (SPD111V, Thermo Electron Corp.) after spectroscopic analysis. The remaining residue was re-dissolved in 600 µL ultra-pure water. Chloroform (75 µL) was added to this solution followed by gentle vortexing and centrifugation (5000 × g, 5 min). After centrifugation, the water phase (uppermost) was transferred very carefully into fresh Eppendorf tubes to remove contamination by photosynthetic pigments (lipophilic) from the MAA (water-soluble). Finally, the samples were filtered by sterilized 0.2 μm pore size syringe filters (Axiva Sichem Biotech., New Delhi) and subjected to the HPLC analysis.
HPLC–PDA analysis of UV-absorbing compound
Partially purified MAA was analyzed using HPLC (Waters, Elstree, UK), using a reverse-phase semi-preparative column (symmetry prep C18, 7 µm particle size, 7.8 mm × 300 mm long) connected to an asymmetry guard column equipped with a Waters Photodiode array detector. Samples (50 μL) were injected into the HPLC column and run at a flow rate of 1.0 mL min−1 using a mobile phase of 0.02% (v/v) acetic acid in ultra-pure water. The detection wavelength was 330 nm and the PDA scan wavelength was from 250 to 450 nm. The sharp peak, with a retention time (RT) of approximately 1.16 min was eluted and collected with the help of a fraction collector attached to the HPLC unit and quantification of MAA was performed using the peak area (Richa and Sinha 2015).
Biochemical characterization of MAA
Electrospray ionization-mass spectrometry (ESI–MS)
The HPLC purified fraction of MAA from Fischerella sp. strain HKAR-14, was subjected to electrospray ionization-mass spectrometry (ESI–MS) to produce protonated molecules. Mass spectra were recorded on an Amazon SL mass spectrometer (Bruker Daltonics Inc., Billerica, MA, USA). Cone voltage of 30 V was found to induce the formation of (M + H)1+ with a mass range of 100–1000 m/z. Data were analyzed using the software Data Analysis 4.0 (Bruker Daltonics Inc., Billerica, MA, USA).
Fourier-transform infrared (FTIR) spectroscopy
For identification of functional groups and analysis of structures of the unknown mixture, FTIR spectroscopy is considered to be the well-established, useful and informative method. Lyophilized MAA was fused with oven-dried KBr (stored in desiccators) in 1:100 ratio, and a transparent disk was prepared using a hydraulic press and inserted in a Perkin Elmer Infrared Spectrophotometer version 10.03.05 (Perkin Elmer, Waltham, MA, USA) to record the spectra (Richa and Sinha 2015).
Nuclear magnetic resonance (NMR) spectroscopy
Similarly, lyophilized MAA was dissolved in D2O (isotopic purity 99.5 atoms) at a concentration of 5 mg mL−1 and 1H and 13C-NMR spectra were carried out with a JEOL AL300 FTNMR spectrometer (JEOL Ltd., Tokyo, Japan) at room temperature (Richa and Sinha 2015).
Electrochemical characterization of MAA
Phosphate buffer (0.1 M) was prepared from potassium dihydrogen orthophosphate (KH2PO4) and dipotassium hydrogen orthophosphate (K2HPO4) purchased from Merck, India. The phosphate buffers of different pH (7.8, 7.0, 6.6, 6.2 and 5.8) were prepared by varying the amounts of KH2PO4 and K2HPO4 in distilled water. Aqueous solution of MAA (0.067 M) extracted from Fischerella sp. strain HKAR-14 was used for all the electrochemical experiments. Triple distilled water was used in the electrochemical experiments and for the solutions preparation. The electrochemical experiments were carried out using CH Instruments (CHI-660C, USA) electrochemical workstation. A glassy carbon (GC) electrode (surface area = 0.07 cm2) as working electrode, a platinum wire as a counter electrode, and a saturated calomel electrode (SCE) as a reference electrode were used in a three-electrode system.
Antioxidant activity of MAA
2,2-diphenyl-1-picryl-hydrazyl (DPPH) radical scavenging assay
The radical scavenging activity of the purified MAA was determined using the DPPH (2,2-diphenyl-1-picryl-hydrazyl) free-radical scavenging assay (Chaiklahan et al. 2013). Different concentrations of MAA (2.0 mL) were mixed with 2.5 mL of DPPH (100 μM in methanol) followed by incubating the mixture for 30 min. The absorbance of the mixtures was measured at 517 nm and ascorbate was chosen as a positive control. The scavenging activity was calculated according to the equation:
where A0 is the absorbance of DPPH solution (water instead of the sample), A1 is the absorbance of the sample and A2 is the absorbance of the sample without DPPH solution.
Ferric reducing antioxidant power (FRAP) assay
The FRAP assay was performed according to the method described by Benzie and Strain (1996). The working FRAP reagent was made by mixing 300 mM acetate buffer (pH 3.6), 2,4,6-tripyridyl-S-triazine (TPTZ) solution (10 mM), and FeCl3·6H2O (20 mM) in a 10:1:1 ratio and heated to 37 °C prior to use. Preparation of 300 mM acetate buffer was done by mixing 3.1 g of sodium acetate trihydrate (NaOAc·3H2O) with 16 mL glacial acetic acid and the final volume was maintained by adding it to 1 L with double-distilled water. Equal volumes of 10 mM TPTZ were mixed with 40 mM HCl for preparation of the TPTZ solution. For the actual assays, a 9:1 ratio of FRAP reagent and sample was incubated in the dark at 37 °C for 5 min to allow the reaction of MAA with the FRAP solution. The absorbance was measured at 593 nm against a blank reagent. Ascorbic acid was used as positive control sample.
Superoxide radical scavenging activity (SRSA) assay
The SRSA of the aqueous isolated MAA was evaluated using an improved pyrogallol (1,2,3-trihydroxybenzene) autoxidation method as described by Li (2012). Freshly prepared solution of 60 mM pyrogallol (prepared in 10 mM HCl) was used for the assay. Sample solution (100 μL) (at pH 7.4) was mixed with 50 mM Tris–HCl buffer (870 μL) at pH 8.2 containing 1 mM EDTA, and 30 μL pyrogallol solution. Quickly after the addition of pyrogallol solution, the absorbance of the reaction mixture was measured at 325 nm in time scan mode for 300 s. The SRSA percentage was calculated using the formula:
where ΔAcs and ΔAas indicate the changes in the absorbance of the reaction mixture of control and antioxidant samples, respectively.
2,2-Azinobis (3-ethylbenzothiazoline 6-sulfonate) (ABTS•+) radical scavenging assay
This method is based on the decolorization (decrease in the absorbance) of the radical cation of 2,2-azinobis (3-ethylbenzothiazoline 6-sulfonate; ABTS•+) produced due to oxidation of ABTS with potassium persulfate (Re et al. 1999). The cation has typical absorption peaks at 820, 734, 660, and 413 nm, and the addition of antioxidants (MAA) to the pre-formed radical determines its reduction which can be observed by a decrease in absorbance. ABTS stock solution (7 mM) with 2.45 mM potassium persulfate (final concentration) reacts to produce the ABTS radical cation. The mixture was kept in the dark for 12–16 h at room temperature before use, for assuring the complete formation and stability of the radical.
ABTS·+ solution (0.9 mL) and 0.1 mL of various concentrations of purified MAA were mixed for 45 s and the absorbance was measured at 734 nm after 1 min and dose-dependent responses were analyzed. Ascorbic acid (AA) (variable amounts) was used as a positive control (Arnao et al. 1996; Miller and Rice-Evans 1997). After 1 min, absorbance measurements were recorded since the decolorization reaction with the reference antioxidant (ascorbic acid) was practically completed by this time (Re et al. 1999). The ABTS radical scavenging activity was calculated with the formula:
where A0: absorbance of the control (without MAA) and A1: the absorbance of the samples.
Statistical analysis
All the experiments were conducted with three replicates to evaluate the means and standard deviation (mean ± SD). One-way analysis of variance (ANOVA) was used to evaluate the significance of the data. Multiple comparisons were done utilizing the Tukey’s HSD post hoc test. For all tests, the level of significance was set at 0.05. Microsoft Excel (2010), Sigma Plot 11 and SPSS-16 software were used for performing all statistical analyses.
Results
Morphological and phylogenetic analyses
The isolated cyanobacterium was heterocystous, autotrophically growing and characterized by a thallus which was felt-like and composed from creeping uni- or multiserial filaments. The identification of the isolated taxon Fischerella sp. HKAR-14 was confirmed by 16S rRNA gene sequencing and the gene sequence was submitted in National Center for Biotechnology Information (NCBI) database with accession number KT150975. Further identification was done through phylogenetic tree mapping utilizing the maximum likelihood method (Fig. 1) (Tamura et al. 2011). The isolated cyanobacterium was identified as Fischerella sp. strain HKAR-14.
Fig. 1.

Phylogenetic relationship of the isolated cyanobacterium Fischerella sp. strain HKAR-14 among cyanobacteria and its analysis inferred from 16S rRNA gene sequences. The evolutionary distances of isolated cyanobacteria were computed using the maximum composite likelihood method. The sequence obtained in the present study is indicated by a black circle
Growth and photosynthetic pigments
The change in growth (absorbance at 750 nm) of Fischerella sp. strain HKAR-14 was measured under different radiation conditions of PAR, PA and PAB which has been summarized in Table 1. PAR-irradiated cells showed strongest growth; in PA- and PAB-irradiated cells, significant decrease in growth was observed with pronounced inhibition in PAB. The amount of photosynthetic pigments in the cyanobacterium was almost constant (P > 0.05) under PAR irradiation. The Chl a content of Fischerella showed 28% and 60% depletion after 60 and 72 h of PA and PAB exposure, respectively, as compared to the controls. A slight increase was observed in PAR-treated samples up to 72 h of treatment. Total carotene content increased to approximately 33 and 52% following 60 h and 48 h in PA- and PAB-treated samples, respectively, in comparison to the controls.
Table 1.
Growth, Chlorophyll a and total carotene contents of Fischerella sp. strain HKAR-14 exposed to PAR, PA or PAB
| Time (h) | Growth at 750 nm | Chl a (mg mL−1) | Total carotene (mg mL−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| PAR | PA | PAB | PAR | PA | PAB | PAR | PA | PAB | |
| 0 | 0.36 ± 0.018 | 0.36 ± 0.018 | 0.36 ± 0.018 | 0.025 ± 0.001 | 0.025 ± 0.001 | 0.025 ± 0.001 | 3.35 ± 0.1 | 3.35 ± 0.1 | 3.35 ± 0.1 |
| 12 | 0.388 ± 0.0232 | 0.369 ± 0.0295 | 0.351 ± 0.0238 | 0.029 ± 0.002 | 0.027 ± 0.0011 | 0.024 ± 0.008 | 3.39 ± 0.26 | 3.72 ± 0.16 | 4.29 ± 0.25 |
| 24 | 0.445 ± 0.0320 | 0.395 ± 0.0229 | 0.345 ± 0.0224 | 0.033 ± 0.0014 | 0.03 ± 0.0023 | 0.022 ± 0.0012 | 3.62 ± 0.32 | 4.18 ± 0.21 | 5.43 ± 0.15 |
| 36 | 0.523 ± 0.0407 | 0.459 ± 0.0230 | 0.335 ± 0.0221 | 0.038 ± 0.0013 | 0.028 ± 0.0016 | 0.019 ± 0.0018 | 3.8 ± 0.15 | 4.61 ± 0.22 | 6.15 ± 0.37 |
| 48 | 0.565 ± 0.0452 | 0.435 ± 0.0307 | 0.312 ± 0.0212 | 0.04 ± 0.002 | 0.024 ± 0.002 | 0.016 ± 0.0021 | 4.19 ± 0.3 | 4.86 ± 0.19 | 6.9 ± 0.32 |
| 60 | 0.648 ± 0.0388 | 0.426 ± 0.0232 | 0.219 ± 0.0172 | 0.038 ± 0.0008 | 0.021 ± 0.0009 | 0.012 ± 0.0007 | 3.85 ± 0.1 | 4.98 ± 0.28 | 5.52 ± 0.27 |
| 72 | 0.692 ± 0.0567 | 0.409 ± 0.0318 | 0.253 ± 0.0169 | 0.035 ± 0.0018 | 0.018 ± 0.0011 | 0.01 ± 0.0024 | 3.99 ± 0.3 | 4.52 ± 0.41 | 5.09 ± 0.18 |
Values represent means ± SD
Photosynthetic performance
The changes in maximal quantum yield of the photosynthetic apparatus (Fv/Fm) of Fischerella were measured under PAR, PA and PAB as shown in Fig. 2A. A significant reduction in Fv/Fm was observed when the cultures were exposed to both PA and PAB compared to PAR alone (P < 0.05), with exposure to PAB causing maximum inhibition (Fig. 2B). The photosynthetic performance of Fischerella was determined as a function of in vivo Chl a fluorescence after 12, 24, 36, 48, 60 or 72 h exposure under PAR, PA or PAB. A decrease in optimum photosynthetic yield was observed under PA and PAB irradiation in comparison to the controls. Maximum decrease was observed in the sample exposed in PAB for 36 h.
Fig. 2.
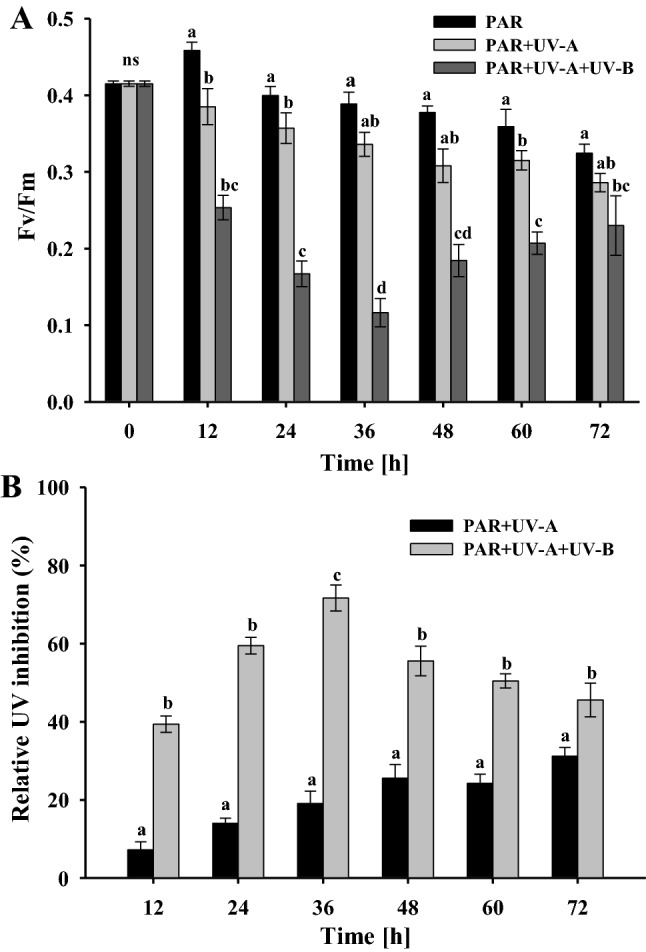
Photosynthetic yields (Fv/Fm) of Fischerella sp. strain HKAR-14 (A), Relative inhibition of maximum quantum yield of PSII (Fv/Fm) (B), after exposure to photosynthetically active radiation (PAR), PAR + UV-A (PA) and PAR + UV-A + UV-B (PAB) at different time intervals up to 72 h. A horizontal line over bars indicates no significant difference (P > 0.05) among treatments. The error bars denote standard deviations of means (means ± SD, n = 3). Similar letters over bar represent homogeneous mean groups (P > 0.05)
Initially, there was a slight increase in Fv/Fm after 12 h of exposure to PAR with no significant differences (P > 0.05). Fv/Fm declined significantly (P > 0.05) to about 15% under PAR between 24 and 72 h. Similarly, Fv/Fm declined up to 31% with significant differences between 36 and 72 h of PA exposure while a non-significant (P > 0.05) decline was observed during the exposure between 12 and 36 h. In PAB exposure, Fv/Fm declined significantly up to 71% following 36 h exposure. The same was increased significantly, up to 45% between 48 and 60 h of treatment compared to the controls. All the three irradiation treatments differed significantly from each other (P < 0.05) between treatments. The cyanobacterium exhibited a decrease in Fv/Fm between 12 and 72 h, and those treated with PAB showed a significantly sharper transient drop.
A significant negative correlation was observed between growth, Chl a and Fv/Fm over time and ROS generation in the studied organism with PAB exposure. This may be because with increasing exposure of UVR, growth, Chl a and Fv/Fm decreased initially which increased on subsequent exposure. Conversely, a positive correlation was observed between total carotene and MAA production with respect to time and ROS generation in the studied cyanobacterium (Table 2).
Table 2.
Time course and concentration-dependent correlation between growth
| Growth (730 nm) | Chl a | Total carotenes | Fv/Fm | MAA | |
|---|---|---|---|---|---|
| Time course (R) | − 0.955 | − 0.989 | 0.604 | − 0.512 | 0.994 |
| P value | 0.000805 | 0.0000248 | 0.151 | 0.240 | 0.00000554 |
| Concentration course (R) | − 0.880 | − 0.952 | 0.744 | − 0.624 | 0.984 |
| P value | 0.00900 | 0.000934 | 0.0552 | 0.134 | 0.0000620 |
Chl a, total carotene, Fv/Fm and MAA in PAB, R correlation coefficient
Quenching coefficients
Curves of quenching coefficients, NPQ and qP showed variations with different irradiations (i.e. under PAR, PA and PAB). With increasing levels of actinic irradiance, the NPQ values increased in the treated samples. Significant increase in the NPQ was observed after 36 h of treatment under PAR which continued up to 72 h as compared to the controls (Fig. 3A). Contrastingly, NPQ was found to be higher in the PA-treated samples and increased continuously upto 72 h of exposure. Approximately twofold increase in NPQ value was observed in 72 h-treated samples as compared to the controls (Fig. 3B). Similarly, exposure to PAB greatly increased the NPQ between 12 and 36 h. Further, irradiation decreased NPQ between 48 and 72 h of PAB exposure as compared to the controls and maximum NPQ was reported after 36 h (approximately 3.5-fold) of treatment in comparison to the controls (Fig. 3C).
Fig. 3.

Variation in non-photochemical quenching (NPQ) (A–C), photochemical quenching (qP) (D–F), and relative electron transfer rate (rETR) (G–I) of Fischerella sp. strain HKAR-14 cultures exposed to photosynthetically active radiation (PAR) (a), PAR + UV-A (PA) (b) and PAR + UV-A + UV-B (PAB) (c) irradiation for varying time periods. The error bar represents standard deviation of mean (means ± SD, n = 3)
The irradiance-response curve of qP showed a decline under increasing irradiance in the PAR, PA and PAB treatments. The qP value ranging between 0 and 1 reflects the proportion of open PSII in the thylakoid membrane with the value of 1 indicating a normal photosystem performance. A slight gradual decrease was observed in qP under PAR-treated samples and maximum decline was reported after 72 h (Fig. 3D). A similar trend was observed in PA-treated samples and maximal decrease was observed in 72 h-exposed samples (Fig. 3E). The results showed that PAB radiation led to a significant decrease in qP values which continuously decreased up to 72 h and maximal decrease was observed after 72 h in comparison to the controls (Fig. 3F).
The plots of rETR increased with increasing duration of exposure to irradiation followed by a plateau on reaching a saturation point, and in the control samples, rETR values were found to be lower in all treated samples. The rETR values under PAR were notably lower during actinic light exposure, and the maximum rETR declined after 72 h of PAR treatment (Fig. 3G). However, in samples receiving 12 h PAR exposure, a significant increase was observed. For the PA treatment, there was a gradual decrease which was continuing up to 72 h exposure, and the maximal reduction was about twofold which was found for the controls under an actinic irradiation of 72 h (Fig. 3H). Curves of rETR showed lower initial saturation in PAB-treated samples than in the control samples. After 12 h of exposure, rETR values were drastically reduced which continued up to 36 h followed by an increase between 48 and 72 h (Fig. 3I). These results clearly indicate that even higher levels of PAR inhibit the photosynthesis in Fischerella, and exposure of UVR further augments this effect.
Effects on antioxidant enzyme activity
Exposure of the cyanobacterium to PAR, PA and PAB showed significant alterations in the activity of SOD, CAT and POD (Fig. 4). Figure 4A shows that SOD activity increased from 0.1683 ± 0.0026 to 0.2577 ± 0.0305 U mg protein−1 under PAR which continued up to 72 h. The activity of SOD increased (from 0.1683 ± 0.0026 to 0.2977 ± 0.03 U mg protein−1) 1.8-fold with significant (P < 0.05) differences between 36 and 72 h exposure in PA-treated samples as compared to the controls. Similarly, PAB induced the activity of SOD up to 72 h with significant differences. Its activity increased between 36 and 48 h (from 0.168 ± 0.0026 to 0.424 ± 0.0215 and 0.388 ± 0.0165 U mg protein−1, respectively) in PAB-treated samples in comparison to the controls. However, maximum SOD activity was observed up to 2.5-fold after 36 h of treatment.
Fig. 4.

Activity of antioxidative enzymes in cultures exposed to photosynthetically active radiation (PAR), PAR + UV-A (PA) and PAR + UV-A + UV-B (PAB) irradiation for varying time periods. Superoxide dismutase (SOD) (A), catalase (CAT) (B) and peroxidase (POD) (C). The error bar represents standard deviation of mean (means ± SD, n = 3). A horizontal line over bars indicates no significant difference (P > 0.05) among treatments. Similar letters over the bars represent homogeneous mean groups (P > 0.05)
In PAR-treated samples, CAT activity increased but remained almost equal to the level in the controls (Fig. 4B). Upon PA and PAB exposure, CAT activity increased significantly (P < 0.05) as compared to the controls. CAT activity increased from 0.0920 ± 0.004 to 0.2237 ± 0.013 µmol min−1 mg−1 of protein (2.4-fold) up to 72 h of treatment in PA. Similarly, there was a significant increase (P < 0.05) in CAT activity under PAB, which reached a maximum of 0.3393 ± 0.016 µmol min−1 mg−1 of protein (3.7-fold) after 48 h of treatment compared to the controls (0.0920 ± 0.004 µmol min−1 mg−1 of protein) and then the activity declined after 60 h of PAB exposure.
POD activity (Fig. 4C) was 0.0180 ± 0.0035 U POD mg−1 of protein in the controls and slightly increased from 0.0180 ± 0.0035 to 0.0289 ± 0.0016 following 72 h of PAR exposure. The POD activity enhanced gradually in PA exposure and 1.8-fold (P < 0.05) induction was observed after 72 h of PA treatment. Similarly, POD activity increased significantly under PAB exposure and maximum induction was observed: 2- and 2.5-fold (P < 0.05) following 36 h and 48 h of treatment, respectively. Further irradiation induced a decline in POD activity. Although SOD activity was found to be dominant in the cyanobacterium, induction of CAT appeared to be higher than SOD and POD under PAB exposure.
In vivo detection of ROS
In vivo free radicals in the filaments of Fischerella were detected using fluorescence microscopy after different duration of exposure to PAR, PA and PAB radiations (Fig. 5A). Production of free radicals as indicated by green fluorescence was enhanced with significant differences in PAR, PA and PAB radiations in comparison to the controls.
Fig. 5.

A Fluorescence images after irradiation with photosynthetically active radiation (PAR), PAR + UV-A (PA) and PAR + UV-A + UV-B (PAB) showing intracellular green 2′,7′-dichlorodihydrofluorescein (DCF) fluorescence after various duration of exposure as a result of reactive oxygen species (ROS) production. DCF fluorescence at 0 h was used as a control. B Emission spectra of DCF fluorescence intensities after irradiation with PAR, PA and PAB after different duration of exposure. The error bar represents standard deviation of mean (means ± SD, n = 3). A horizontal line over the bars indicates no significant difference (P > 0.05) among treatments. Similar letters over the bars represent homogeneous mean groups (P > 0.05)
Figure 5B illustrates the emission spectra of samples incubated with DCFH-DA after different duration of exposure to PAR, PA and PAB showing the emission peak at 525 nm after excitation at 485 nm. Maximum generation of ROS occurred in PAB-exposed samples as suggested by the emission spectra. The DCF-fluorescence was significantly (P < 0.05) higher in the 72 h PAB-exposed samples in comparison to PAR or samples exposed to shorter durations of radiation, indicating highest ROS production in the PAB-exposed samples. There was no significant (P > 0.05) difference in DCF fluorescence between PAR- and PA-treated samples; however, the level of ROS was significantly higher (P < 0.05) in these samples as compared to the controls. Hence, data from both fluorescence spectroscopy and microscopy supported each other and clearly suggest that PAB radiation has a pronouncedly higher effect on ROS generation than PAR or PA radiation.
Under UVR (PAB) stress physiological processes, such as ROS generation, SOD, CAT and POD, activities were found to be positively correlated with exposure time in the studied cyanobacterium. Similarly, a positive correlation was observed between activities of SOD, CAT and POD against ROS generation (Table 3). This may be due to increased antioxidative enzymatic activities in response to increasing exposure of UVR.
Table 3.
Time course and concentration-dependent correlation between ROS generation, SOD, CAT and POD activity in PAB
| ROS | SOD | CAT | POD | |
|---|---|---|---|---|
| Time course (R) | 0.978 | 0.689 | 0.817 | 0.605 |
| P value | 0.000134 | 0.0869 | 0.024 | 0.150 |
| Concentration course (R) | – | 0.806 | 0.912 | 0.748 |
| P value | – | 0.0287 | 0.00425 | 0.0531 |
R correlation coefficient
Biosynthesis of UV-absorbing compounds
Spectroscopic analysis of methanolic extracts of Fischerella showed absorption maxima for MAA at 332.3 ± 2 nm (Fig. 6A). Figure 6B shows the HPLC chromatograms obtained from HPLC coupled with a PDA detector revealing that the purified MAA exhibited an RT of 1.16 min and a UV absorption maximum (UVλmax) at 332.3 nm (Fig. 6C). This compound was further investigated for its UV-screening potentials following its induction under PAR, PA and PAB radiation. Time-dependent induction of MAA synthesis was observed under PAB. With increasing exposure to PAB (up to 72 h), the MAA content in the cells increased in comparison to the controls (Fig. 7A).
Fig. 6.

Absorption spectrum of the methanolic extract showing ultraviolet (UV) absorption maximum at 332.3 ± 2 nm indicating the presence of a UV-absorbing mycosporine-like amino acid (MAA) (A), high performance liquid chromatography (HPLC) chromatogram of the partially purified MAA, showing a typical peak at a retention time of 1.157 min (B) and absorption maximum at 332.3 nm (C)
Fig. 7.
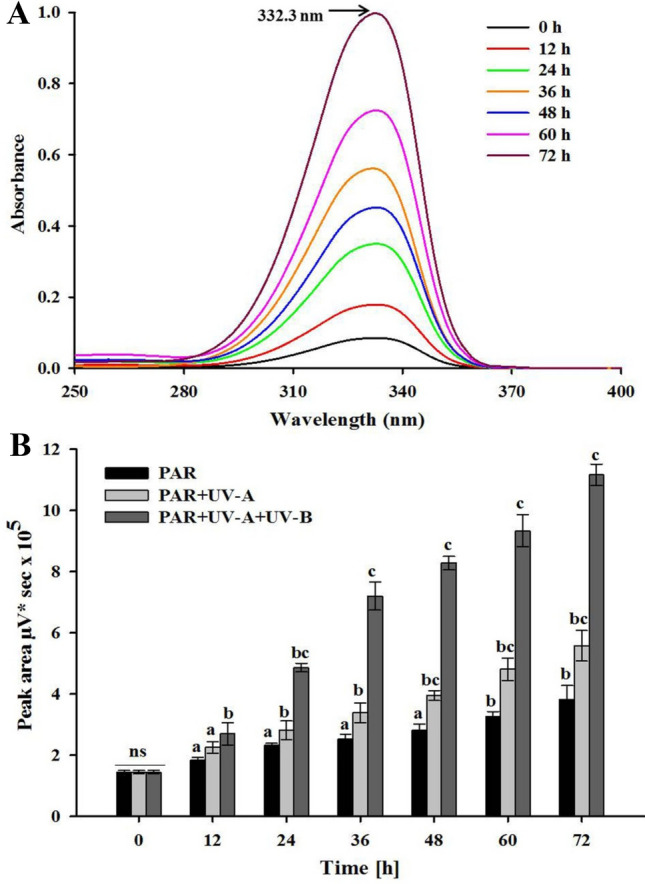
Absorption spectra showing the induction of shinorine after different durations of photosynthetically active radiation (PAR), PAR + UV-A (PA) and PAR + UV-A + UV-B (PAB) in Fischerella sp. strain HKAR-14 (A) and biosynthesis of shinorine under PAR, PA or PAB irradiation for 72 h (B). The error bars represent standard deviation of the means (means ± SD, n = 3). A horizontal line over bars indicates no significant difference (P > 0.05) among treatments. Similar letters over the bars represent homogeneous mean groups (P > 0.05)
Figure 7B shows the induction of MAA biosynthesis under PAR, PA and PAB exposure up to 72 h. Significant (P < 0.05) induction in MAA content was observed in the samples exposed to PAB, followed by those kept in PA in comparison to the controls. Similarly, in samples exposed to PAB, significant (P < 0.05) induction in the MAA content was observed from their initial value as compared to the controls following 12 h of exposure and this increase in MAA content further continued up to 72 h of exposure. MAA induction was significantly higher (P < 0.05) under PAR up to 72 h of exposure as compared to the controls. An increase of 3.8-fold in MAA induction was observed up to 72 h of exposure in PA. In PAB-treated samples, significant (P < 0.05) induction of MAA was observed in comparison to the controls; after 12 h of exposure 1.8-fold induction was observed which continued up to 72 h with eightfold increase in induction of MAA.
There was no significant (P > 0.05) difference in the content of MAA between 12 and 24 h in the PA-exposed samples. In contrast to the PAB treatment, induction of MAA was very low in the cells irradiated under PAR or PA cut-off filters indicating that PAR or PA were less effective in induction and synthesis of MAA in Fischerella sp. strain HKAR-14.
Characterization of MAA by ESI–MS, FTIR and NMR
The identity of the MAA was determined from the ESI–MS analysis (Fig. 8A) of the purified MAA from Fischerella sp. strain HKAR-14 which showed a peak at a value of m/z 333.1 (positive ionization). Based on the RT of the HPLC and the ESI–MS analysis of the purified fragment, the MAA was identified as shinorine (RT 1.16 min, MW-333.1). For further analysis, we proceeded with the purified MAA having a sharp peak from Fischerella with an RT of 1.16 and m/z of 333.1 which was tentatively identified as “shinorine” and was subjected to FTIR analysis. A strong frequency pattern having bands of 3419.96, 2926.92, 2160, 1653.13, 1552, 1440.7, 1107.1 and 616.56 cm−1 was evident in the FTIR results (Fig. 8B). The results obtained showed similarity to the basic functional groups of the MAA shinorine. Bands of 3419.96 cm−1 may be assigned to the –OH group, 2926.92 cm−1 for side chain vibrations consisting of C–H stretching and reflecting the presence of a (NH2+) group and bands of 1653.13 and 1405.47 cm−1 may be assigned to the presence of –NH2 and carboxylic groups, respectively. NMR analysis was also carried out to characterize the compound. The positions of hydrogen (1H) and carbon (13C) indicated that the purified compound is indeed shinorine (Fig. 8C, D). Absence of aromatic and heteroaromatic protons in the compound were detected in proton NMR (1H) analysis. The proton NMR spectrum is expected to show a peak at δ 10.14 (due to presence two acidic proton of –COOH groups). The proton NMR spectrum showed a peak at δ 1.39, 3.40, and 3.50 (due to the presence of protons of a –CH2 group) and at δ 2.10 (due to presence of protons of the –OH group). The 13C-NMR spectrum also showed the presence of an aliphatic functional group (74.27), including a carboxyl carbon (δ 170.08), one imine carbon (δ 145.27), five different type aliphatic carbons (δ 31.61, 41.53, 52.4, 64.8, and 74.27), δ 122.21 with one ethylene (C=C) carbon.
Fig. 8.

Electro spray ionization-mass spectrometry (ESI–MS) of high performance liquid chromatography (HPLC) purified fractions giving a peak with a m/z value of 333.1 confirming the identity of the collected purified fraction as shinorine (A), Fourier Transform Infrared (FTIR) radiation spectroscopy showing basic functional groups present in the purified fraction of the mycosporine-like amino acid (MAA), (B) Nuclear magnetic resonance (NMR) spectra of purified fraction of shinorine, 1H (C) and 13C (d) 1D NMR spectrum of the purified fraction were recorded in D2O solution. NMR spectra revealed correlation with chemical shift data of shinorine
Electrochemical characterization of shinorine
An aqueous solution of shinorine (0.067 M) was added (1.0 mL) to 0.1 M phosphate buffer at pH 7.0 (4.0 mL) in an electrochemical cell. This solution was characterized by the cyclic voltammetry () technique (Fig. 9A) using a three-electrode system at a scan rate of 50 mV s−1. The cyclic voltammetry (CV) scan represents an irreversible oxidation peak (Epa) at 0.85 V. CV scans at different scan rates (20–300 mV s−1) were recorded and are shown in Fig. 9B. The oxidation peak current (Ipa) increases on increasing the scan rate with a slight positive shift in the peak potential. The Ipa of shinorine represents a linear relationship with the square root of the scan rate, indicating a diffusion-controlled process (Fig. 9C). The Epa value observed for shinorine (850 mV) represents a slightly higher value. The characteristics of shinorine were also tested at different pH values (7.8, 7.0, 6.6, 6.2 and 5.8) using 0.1 M phosphate buffers (Fig. 9D). The Ipa observed for shinorine shows a maximum Ipa value at pH 7.0 compared to all other pH values.
Fig. 9.
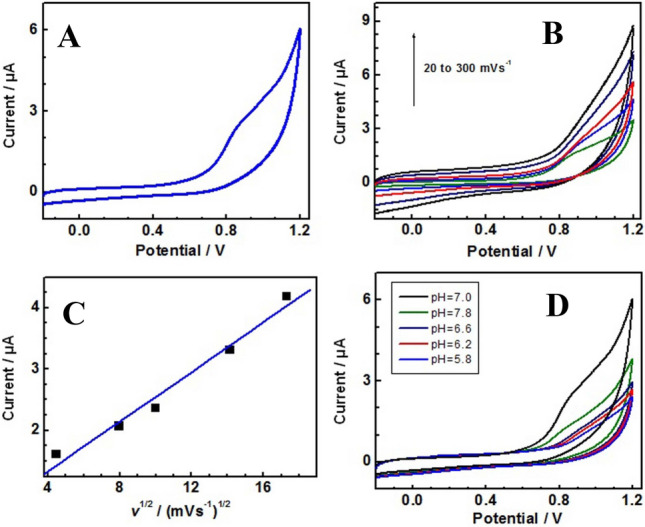
Cyclic voltammetry (CV) scan of the shinorine molecule in 0.1 M pH 7.0 phosphate buffer at the scan rate of 20 mV s−1 (A). CV of shinorine at different scan rates (20, 50, 100, 200, and 300 mVs−1) in 0.1 M pH 7.0 phosphate buffer (B). Plot of anodic peak currents (measured from Fig. 1b) against the square root of scan rates (C). CV scans of shinorine molecule in 0.1 M of different pH buffers (5.8, 6.2, 6.6, 7.0, and 7.8) at the scan rate of 50 mV s−1 (D)
In vitro antioxidant assay of MAA
DPPH radical scavenging assay
Figure 10A shows significant scavenging potentials of shinorine against DPPH radicals, although the percentages of scavenging activities were lower than that of ascorbate within the test dosage range. The percentages of MAA scavenging activities against DPPH radicals increased slightly with increasing concentrations. However, the percentage of MAA scavenging activity was lower. Activity increased from 51.07 ± 2.6061 to 81.20 ± 5.5210% when MAA concentration was increased from 60 to 100 μg mL−1, and it exhibited a concentration-dependent activity (10–100 μg mL−1). Scavenging activity of ascorbate was in the range of 18.2 ± 0.918 to 98.1 ± 6.259% at the tested concentrations.
Fig. 10.

Free radical scavenging capacity of partially purified shinorine as determined by the 2, 2-diphenyl-1-picryl-hydrazyl (DPPH) assay (A), ferric reducing antioxidant power (FRAP) assay (B) and superoxide radical scavenging activity (SRSA) assay (C) and 2, 2-Azinobis (3-ethylbenzothiazoline 6-sulfonate) (ABTS·+) radical scavenging assay (d). The radical scavenging capacity of shinorine was determined and correlated with that of ascorbic acid (AA). The error bars denote standard deviations of means (means ± SD, n = 3)
Determination of the antioxidant activity by FRAP assay
The reducing ability of shinorine showed a linear correlation with the absorbance change at 593 nm and the substance concentration up to a maximum of 1.4 absorbance units, for different concentrations of MAA (10, 20, 40, 60, 80 and 100 μg mL−1) (Fig. 10B). Shinorine exhibited higher reducing potential than ascorbate.
Superoxide radical scavenging activity
Superoxide radical scavenging potential of various concentrations of MAA is presented in Fig. 10C. All of the MAA fractions studied exhibited a concentration-dependent behavior. The percentage scavenging rate was 7.12 ± 0.3987, 15.36 ± 0.4761, 29.7 ± 0.8316, 53.5 ± 1.444, 75.8 ± 1.591 and 82.5 ± 2.640%, at concentrations of 10, 20, 40, 60, 80 and 100 μg mL−1 MAA, respectively. Highest superoxide radical scavenging (82.50 ± 2.640%) activity was exhibited by 100 μg mL−1 MAA.
ABTS radical scavenging activity
ABTS radicals scavenging activity of shinorine in terms of percentage has been shown in Fig. 10D. The scavenging activities were found to be 5.3 ± 0.2599, 9.8 ± 0.6304, 17.9 ± 1.064, 36.33 ± 3.019, 51.33 ± 2.4362 and 62.3 ± 4.544 at concentrations of 10, 20, 40, 60, 80 and 100 μg mL−1 MAA, respectively. The ABTS radical scavenging ability of 100 μg mL−1 MAA was very close to that of ascorbate and higher than of other fractions. While the ABTS radical scavenging ability of MAA exhibited concentration dependence at concentration ranging from 10–100 μg mL−1. The result demonstrated that MAA shinorine from Fischerella showed significant ABTS radicals scavenging activity.
Discussion
Cyanobacteria are affected by both the quality and quantity of the predominant light conditions in their natural environment. UVR is detrimental component of solar radiation which reaches the Earth’s surface and induces marked consequences for morphology, growth, photosynthesis, genome integrity and intracellular ROS production in cyanobacteria (Singh et al. 2013a). In cyanobacteria, photosynthetic performance was found to be adversely affected due to damage in the photosynthetic apparatus (Bhandari and Sharma 2006). In living organisms, UV-B radiation is very harmful (Sinha et al. 1996, 2008; Häder and Sinha 2005; Rastogi et al. 2011). In cyanobacteria, different antioxidant non-enzymatic and enzymatic protective strategies have been observed to counterbalance the detrimental effects of acute short wavelength UVR (Singh et al. 2013a).
The present investigation deals with PAR-, PA- and PAB-induced changes in growth, photosynthetic pigments, maximal quantum yield of PSII (Fv/Fm), NPQ, qP and rETR as well as activity of antioxidative enzymes, such as CAT, SOD and POD, and non-enzymatic mediated MAA production. In different species of cyanobacteria, the effects of UVR may vary (Helbling et al. 2006; Karsten et al. 2007; Pattanaik et al. 2008; Fiorda Giordanino et al. 2011). The growth of Fischerella increased in PAR, while in PA and PAB treatments, a significant decrease in growth was observed. Similar findings were reported by Rastogi et al. (2014) in Anabaena siamensis TISTR-8012 and these growth responses are probably results of varying levels of damage caused by UV-B indirectly or directly on DNA, proteins and the photosynthetic apparatus, which are responsible for the observed growth inhibition (Friso et al. 1994; Callaghan et al. 2004).
UVR damaged the photosynthetic pigments in Fischerella which varied significantly with respect to exposure time as depicted in Table 1 and it was observed that the content of photosynthetic pigment Chl a varied after different durations of PAR, PA and PAB exposure. A significant reduction in Chl a was observed in PA while a stronger decrease was reported in PAB. The intensive irradiation of photosynthetic pigments with PA and PAB shows rapid bleaching and the same represses photosynthetic processes in cyanobacteria (Bhandari and Sharma 2006; Singh et al. 2013a; Kannaujiya and Sinha 2017). A progressive increment in Chl a was found to a certain degree in cyanobacteria exposed to PAR and PA radiation in contrast to PAB exposure. Similarly, significant reduction in Chl a was observed in PAB in A. siamensis TISTR-8012 (Rastogi et al. 2014). A decrease in Chl a concentration may be attributed to photoreduction of protochlorophyllide to chlorophyllide under UV-B stress (Marwood and Greenberg 1996). With regard to the total carotene content after PA exposure, it can be observed that PA exposure resulted in enhanced production of carotenes, but PAB strongly enhanced total carotene production up to 48 h of exposure followed by a decrease in subsequent hours of exposure. Carotenes are well-established photoprotective compounds with high ROS-scavenging potential (Young 1991; Pattanaik et al. 2008). Enhancement in total carotene content due to UV-B in Aphanothece sp. might be ascribed to their role as scavenger of singlet oxygen (Vincent and Quesada 1994).
The measurement of Fv/Fm is widely used for revealing the maximum light-capturing potential occurring within the PS at any time in the cell. In this study, the photosynthetic efficiency of Fischerella was found to be greatly affected in PAB- and PA-exposed samples; also under PAR, the decrement in photosynthetic yield has been found (Bhandari and Sharma 2006; Singh et al. 2013b).The observed decrease in photosynthetic performance in Fischerella under UVR was in accordance with previous reports which have reported reduction in Fv/Fm, growth and pigments under UVR in several species of cyanobacteria (Gao et al. 2007; Singh et al. 2013b; Fragoso et al. 2014). The photosynthetic yield was extremely repressed even after 10 min of high PAR and UVR in Anabaena variabilis PCC 7937; however, the photosynthetic yield recovered after 2 h of exposure, which proceeded during the following 24 h (Singh et al. 2013b), showing the presence of photoprotective strategies in the cyanobacteria which may enable their survival under severe environmental conditions of increased UVR (Sinha et al. 2008). Furthermore, UV-A damage to PSII is usually less than in UV-B, although both have similar effects on PSII (Vass 2012). Additionally, the turnover of D1 and D2 proteins of the PSII reaction center in cyanobacteria can be increased, when exposed to UV-B radiation ameliorating the stress (Sass et al. 1997). A few investigations have revealed the harmful effects of UVR (long-term exposure with mild PAR and UVR) on photosynthetic performance and its recovery after damage (Karsten et al. 2007).
The results from the present study reveal that the induction of NPQ confer some protection to PSII as reflected by the inverse relationship between photoinhibition of photosynthesis under PA and PAB (Fig. 2) and the ability to induce NPQ (Fig. 3). NPQ mechanisms help in dissipating the absorbed solar energy within the antenna system before reaching the reaction center of PSII (Müller et al. 2001). These mechanisms include phycobilisome-dependent NPQ in cyanobacteria, that is mediated by the orange carotenoid protein, which plays a role in photoprotection in diverse cyanobacteria (Wilson et al. 2006). The NPQ mechanism assists in diminishing the excitation pressure on PSII and helps in providing protection against triplet chlorophyll-dependent and/or electron transport-dependent photodamage (Vass 2011). In cyanobacteria, non-photochemical quenching is dominated by state transitions (Campbell et al. 1998) which have been rectified by recent experiments demonstrating carotenoid-triggered energy dissipation in phycobilisomes (Kirilovsky 2007).
Cyanobacterial species manage changing light conditions use different D1 forms within the PSII reaction center in response to low and high light conditions (Golden 1995). Recent studies showed that the light-dependent exchange of D1 protein forms is identified with ideal photoprotection. In this process, the regulation of non-radiative charge recombination by means of altering the redox potential of Phe via the exchange of Glu and Gln residues at the 130th amino acid position in the D1 protein plays a crucial role (Tichý et al. 2003; Cser and Vass 2007; Kós et al. 2008; Sander et al. 2010; Sugiura et al. 2010). The irradiance-response curve of qP varied with respect to qualitative changes in radiations, i.e. PAR, PA, PAB in this study. A decline under increasing irradiances of PAR, PA and PAB was observed; but qP strongly declined under PAB as the exposure duration increased. The exposure to PAR had little effect on qP. Moreover, a decrease in qP is related to an increase in NPQ; nevertheless, the NPQ-protecting mechanism failed to save cyanobacterial cells from death (Wang et al. 2016).
In this study, Fischerella exhibited an increase in NPQ and a decrease in qP with increasing duration of irradiance. A crucial factor which determines the probability of PSII photochemistry in cyanobacteria is the redox state of QA, i.e., the proportion of PSII reaction centers in the thylakoid membrane that are open and capable of doing photochemistry. Frequently, the PSII efficiency factor (or the mathematically equivalent qP) is employed to estimate the redox state of QA (Baker 2008).
Our results show that rETR in PAR, PA and PAB differ significantly. rETR values increased with the increasing irradiation until it reached a saturation point, and rETR values were found to be lower in PA- and PAB-exposed samples than the PAR-treated samples (Fig. 3g–i). With the increasing exposure to PAR, the rETR increased initially, and afterwards a slight decrease was observed, indicating that the electron transporting efficiency of the photosynthetic system was down-regulated when exposure to PAR was excessive. Response of rETR was similar as the quantum yield of PSII and was found to be highest in the samples receiving only PAR, but in samples receiving PAB, rETR values were lower. Thus, it is clear that even higher levels of PAR inhibited the photosynthesis in accordance with observations in A. variabilis PCC 7937, and exposure of UVR further augmented this process (Singh et al. 2013b).
Our study indicates the presence of enzymatic as well as non-enzymatic defense strategies against UVR stress in Fischerella. These defense strategies ensure their survival and growth under UVR-induced oxidative stress conditions. Multifold induction of antioxidative enzymes activity, such as SOD, CAT and POD, was observed upon PA and PAB irradiation in comparison to PAR. However, maximum induction was observed in PAB-treated samples in all studied antioxidative enzymes. Our findings are consistent with previous reports by Singh et al. (Singh et al. 2013b) in Anabaena doliolum. To counteract the deleterious effects of UV-B radiation, significant induction of antioxidative enzymes was observed in A. doliolum and Anabaena strain L31. Increased activity of ascorbate peroxidases (APX) and SOD was reported in Phormidium corium and Nostoc spongiaeforme under high-light exposure (Bhandari and Sharma 2006). Earlier studies have also shown the induction of transcripts of antioxidative enzyme systems by UV-B radiation (Agrawal et al. 2009; Singh et al. 2010b). The activity of SOD, CAT and POD was found to be enhanced after 15 and 60 min of UV-B exposure in three cyanobacteria (Zeeshan and Prasad 2009). The greater increase in the activity of SOD, CAT and POD defense systems helps in protecting the cyanobacterial cells by scavenging excessive ROS resulting inthe protection of PSII under enhanced UVR. Exogenous supplementation of antioxidants, such as N-acetylcysteine, ascorbic acid and sodium nitroprusside, enhanced survival rates of Anabaena sp. (He and Häder 2002b) and Nostoc sp. (Wang et al. 2008) by decreasing Chl a bleaching, damage to the photosynthetic apparatus, photoinhibition of photosynthesis, lipid peroxidation and DNA damage following UVR exposure.
The present study and earlier findings strongly support the fact that Fischerella produces endogenous antioxidant molecules under UVR exposure to counteract oxidative stress induced by UV-mediated ROS. Apart from enzymatic antioxidants, the UV-screening compounds MAAs and scytonemin are also synthesized by several cyanobacteria, which can likewise work as non-enzymatic antioxidant biomolecules ameliorating the damaging effects of UVR (Singh et al. 2013b). A literature survey revealed that Fischerella muscicola TISTR8215, isolated from Thailand, produced a single MAA, M-332, in response to UVR (Rastogi and Incharoensakdi 2015). Several organisms including cyanobacteria inhabiting diverse habitats have been found to synthesize different UV-screening MAAs to counterbalance the damaging effects of UVR (Rastogi et al. 2010b; Sinha et al. 2007; Pathak et al. 2017). In the last decades, several MAAs have been identified and characterized biochemically (Carreto et al. 2005; Carreto and Carignan 2011; Singh et al. 2013b; Richa and Sinha 2015), however, an extensive number of MAAs still has to be biochemically characterized. We have demonstrated for the first time that Fischerella sp. strain HKAR-14 isolated from hot springs of Chandra Kund, Bihar, India, synthesizes and accumulates a single MAA with λmax 332.3 nm and with an RT of 1.16 min (Fig. 8b). FTIR data of the purified MAA resembled the previously reported shinorine and hence, the purified MAA was confirmed as shinorine (Whitehead and Hedges 2003; Oyamada et al. 2008). The NMR spectra suggested an acceptable correlation with earlier published chemical shift data for shinorine (Oyamada et al. 2008). In this study, all the data of biochemical characterization of the purified MAA confirmed that the main UV-screening compound synthesized under UVR stress is indeed shinorine.
The present investigation showed that the Fischerella sp. strain HKAR-14 is capable to induce MAA biosynthesis in response to UVR within 24 h which may allow it to acclimatize under harsh environmental conditions of UVR, exhibiting a protective mechanism against high PAR and UVR that it faces in its natural habitat. The presence of high concentrations of UV-absorbing compounds in cells is also supposed to provide protection under UVR in A. siamensis TISTR 8012 (Rastogi et al. 2014).
Findings of the present study also showed that MAA may protect the cell by scavenging ROS and this was confirmed by an in vitro antioxidant assay of shinorine allowing the cyanobacterium to resist UVR-induced oxidative stress. Similar findings were reported by de la Coba et al. (2009) and Torres et al. (2018) of in vitro antioxidant capacities of MAAs.
Electrochemical properties of shinorine were characterized by the CV technique which showed an irreversible oxidation peak (Epa) at 0.85 V displaying a very similar electrochemical behavior as gadusol (a similar molecule as shinorine) (Plack et al. 1981; Arbeloa et al. 2012). The oxidation peak current (Ipa) of shinorine increases on increasing the scan rate with a slight positive shift in the peak potential representing a linear relationship with the square root of the scan rate, indicating a diffusion-controlled process (Bard et al. 1980; Sonkar and Ganesan 2015; Sonkar et al. 2017). The Epa value observed for shinorine (850 mV) represents a slightly higher value than that of similar molecules, such as gadusol and gadusolate (Plack et al. 1981; Blasco et al. 2007; Arbeloa et al. 2012). Thus, shinorine is a more powerful reducing agent than gadusol and gadusolate and therefore efficient antioxidant properties are expected at physiological pH values (pH 7.0) (Arbeloa et al. 2012). The characteristics of shinorine were also tested at different pH values and it was observed that the oxidation peak current (Ipa) showed a maximum Ipa value at pH 7.0 compared to all other pH values. Hence, the physiological pH (pH 7.0) is the best pH for the redox activity of the shinorine molecule and its reducing and antioxidant behaviors (Arbeloa et al. 2012). There has been clear evidence that biosynthesis and induction of MAAs helps in protection of vital functions in several organisms including cyanobacteria against deleterious short wavelength UVR (Garcia-Pichel and Castenholz 1993; Garcia-Pichel et al. 1993; Klisch et al. 2001).
Conclusion
Based on the results obtained from this study, it was clearly demonstrated that PA and PAB affect the growth, photosynthetic pigments and damage Fv/Fm and rETR in the cyanobacterium Fischerella sp. strain HKAR-14. In comparison with PAR, UVR significantly changed the distribution of energy. NPQ may be the main pathway for the excessive energy dissipation in PSII exposed to UVR, while a crucial role is played by photochemical quenching in preventing photoinhibition. These results indicate that the damage caused by UVR is possibly due to the formation of ROS. For mitigating the ROS-induced oxidative stress, cyanobacteria have developed efficient antioxidant systems, including antioxidant molecules, such as carotenoids and antioxidant enzymes, such as SOD, CAT and POD. In addition to the enzymatic antioxidants, the cyanobacterium also synthesizes and accumulates the MAA shinorine that acts as an active UVR photoprotectant. Shinorine also protects against ROS through scavenging free radicals. Results indicate that there is a dynamic balance between damage and repair in the studied cyanobacterium which helps its long-term survival after intensive UVR exposure. In addition, MAAs are able to convert the absorbed energy to heat without the formation of free oxygen species and to antagonize the free radical-induced oxidative damage. The shinorine molecule has been characterized with cyclic voltammetry technique. It shows optimum antioxidant characteristics in 0.1 M in pH 7.0 phosphate buffer. The electrochemical behavior represents the diffusion-controlled process on the electrode surface. This feature of the MAA can be used to develop novel biodegradable sunscreens and other bioactive products in pharmaceutical/cosmetics industries.
Accession numbers The identification of the cyanobacterium was confirmed by 16S rRNA gene sequencing with Accession number: KT150975.
Acknowledgements
Haseen Ahmed [UGC-JRF-21/12/2014 (ii) EU-V] thankfully acknowledges University Grant Commission, New Delhi, India, for providing fund in the form of fellowship. Jainendra Pathak (09/013/0515/2013-EMR-I) is thankful to Council of Scientific and Industrial Research (CSIR), New Delhi, India, for the financial support in the form of fellowships. Department of Biotechnology (DBT), Govt. of India, (DBTJRF/13/AL/143/2158) is gratefully acknowledged for providing fellowship to Rajneesh. Authors are also thankful to the Interdisciplinary School of Life Sciences (ISLS), BHU, Varanasi, India, for providing access to the ESI-MS and fluorescence microscopy facility. Department of Chemistry, BHU, Varanasi, India, is acknowledged for providing FTIR and NMR facilities.
Author contributions
HA designed and conducted the experiments, analyzed the data and wrote the manuscript. JP and R helped in performing experiments and in writing the manuscript. PKS and VG helped in performing experiments. RPS provided laboratory facilities and with D-PH edited the manuscript. The final manuscript was read and approved by all the authors.
Compliance with ethical standards
Conflict of interest
Authors declare no conflict of interest.
Research does not involve human participants and/or animals
No Informed consent is required.
Contributor Information
Haseen Ahmed, Email: haseenahmedbhu@gmail.com.
Jainendra Pathak, Email: gpathakvns@gmail.com.
Rajneesh, Email: rajshr2@gmail.com.
Piyush K. Sonkar, Email: piyush.sonkar37@gmail.com
Vellaichamy Ganesan, Email: velganesh@yahoo.com.
Donat-P. Häder, Email: donat@dphaeder.de
Rajeshwar P. Sinha, Email: rpsinhabhu@gmail.com
References
- Aebi H (1984) Catalase in vitro. Methods in enzymology. In: Packer L (ed) Academic Press Inc, San Diego, pp 121–126. https://doi.org/10.1016/S0076-6879(84)05016-3 [DOI] [PubMed]
- Agrawal S, Singh S, Agrawal M. Ultraviolet-B induced changes in gene expression and antioxidants in plants. Adv Bot Res. 2009;52:47–86. doi: 10.1016/S0065-2296(10)52003-2. [DOI] [Google Scholar]
- Ahmed H, Pathak J, Rajneesh, Pandey A, Singh PR, Sinha RP (2019) Genomics and proteomics of photoprotective compound mycosporine-like amino acids in cyanobacteria. Innovations in Life Science Research. Nova Science Publishers, Hauppauge, pp 97–128
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- Arbeloa EM, Ramírez CL, Procaccini RA, Churio MS. Electrochemical characterization of the marine antioxidant gadusol. Nat Prod Commun. 2012 doi: 10.1177/1934578x1200700928. [DOI] [PubMed] [Google Scholar]
- Arnao M, Cano A, Hernandez-Ruiz J, Garcıa-Cánovas F, Acosta M. Inhibition byl-ascorbic acid and other antioxidants of the 2,2′-azino-bis (3-ethylbenzthiazoline-6-sulfonic Acid) oxidation catalyzed by peroxidase: a new approach for determining total antioxidant status of foods. Anal Biochem. 1996;236:255–261. doi: 10.1006/abio.1996.0164. [DOI] [PubMed] [Google Scholar]
- Baker NR. Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu Rev Plant Biol. 2008;59:89–113. doi: 10.1146/annurev.arplant.59.032607.092759. [DOI] [PubMed] [Google Scholar]
- Barbato R, Bergo E, Szabo I, della Vecchia F, Giacometti GM. Ultraviolet B exposure of whole leaves of barley affects structure and functional organization of photosystem II. J Biol Chem. 2000;275:10976–10982. doi: 10.1074/jbc.275.15.10976. [DOI] [PubMed] [Google Scholar]
- Bard AJ, Faulkner LR, Leddy J, Zoski CG. Electrochemical methods: fundamentals and applications. New York: Wiley; 1980. [Google Scholar]
- Barufi JB, Korbee N, Oliveira MC, Figueroa FL. Effects of N supply on the accumulation of photosynthetic pigments and photoprotectors in Gracilaria tenuistipitata (Rhodophyta) cultured under UV radiation. J Appl Phycol. 2011;23:457–466. doi: 10.1007/s10811-010-9603-x. [DOI] [Google Scholar]
- Benzie IF, Strain JJ. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: the FRAP assay. Anal Biochem. 1996;15:70–76. doi: 10.1006/abio.1996.0292. [DOI] [PubMed] [Google Scholar]
- Beyer WF, Jr, Fridovich I. Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Anal Biochem. 1987;161:559–566. doi: 10.1016/0003-2697(87)90489-1. [DOI] [PubMed] [Google Scholar]
- Bhandari R, Sharma PK. High-light-induced changes on photosynthesis, pigments, sugars, lipids and antioxidant enzymes in freshwater (Nostoc spongiaeforme) and marine (Phormidium corium) cyanobacteria. Photochem Photobiol. 2006;82:702–710. doi: 10.1562/2005-09-20-RA-690. [DOI] [PubMed] [Google Scholar]
- Bischof K, Gómez I, Molis M, Hanelt D, Karsten U, Lüder U, Roleda MY, Zacher K, Wiencke C. Ultraviolet radiation shapes seaweed communities. Rev Environ Sci Bio. 2006;5:141–166. doi: 10.1007/s11157-006-0002-3. [DOI] [Google Scholar]
- Blasco AJ, González Crevillén A, González MC, Escarpa A. Direct electrochemical sensing and detection of natural antioxidants and antioxidant capacity in vitro systems. Electroanal Int J Devoted Fundam Pract Asp Electroanal. 2007;19:2275–2286. doi: 10.1002/elan.200704004. [DOI] [Google Scholar]
- Burkholder JB, Cox RA, Ravishankara AR. Atmospheric degradation of ozone depleting substances, their substitutes, and related species. Chem Rev. 2015;115:3704–3759. doi: 10.1021/cr5006759. [DOI] [PubMed] [Google Scholar]
- Burow S, Valet G. Flow-cytometric characterization of stimulation, free radical formation, peroxidase activity and phagocytosis of human granulocytes with 2,7-dichloroflorescein (DCF) Eur J Cell Biol. 1987;43:128–133. [PubMed] [Google Scholar]
- Callaghan TV, Björn LO, Chernov Y, Chapin T, Christensen TR, Huntley B, Ims RA, Johansson M, Jolly D, Jonasson S. Responses to projected changes in climate and UV-B at the species level. Ambio. 2004;33:418–435. doi: 10.1579/0044-7447-33.7.418. [DOI] [PubMed] [Google Scholar]
- Campbell D, Hurry V, Clarke AK, Gustafsson P, Öquist G. Chlorophyll fluorescence analysis of cyanobacterial photosynthesis and acclimation. Microbiol Mol Biol Rev. 1998;62:667–683. doi: 10.1128/mmbr.62.3.667-683.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carreto JI, Carignan MO. Mycosporine-like amino acids: relevant secondary metabolites. Chemical and ecological aspects. Marine Drugs. 2011;9:387–446. doi: 10.3390/md9030387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carreto JI, Carignan MO, Montoya NG. A high-resolution reverse-phase liquid chromatography method for the analysis of mycosporine-like amino acids (MAAs) in marine organisms. Mar Biol. 2005;146:237–252. doi: 10.1007/s00227-004-1447-y. [DOI] [Google Scholar]
- Chaiklahan R, Chirasuwan N, Triratana P, Loha V, Tia S, Bunnag B. Polysaccharide extraction from Spirulina sp. and its antioxidant capacity. Int J Biol Macromol. 2013;58:73–78. doi: 10.1016/j.ijbiomac.2013.03.046. [DOI] [PubMed] [Google Scholar]
- Conde FR, Churio MS, Previtali CM. The photoprotector mechanism of mycosporine-like amino acids. Excited-state properties and photostability of porphyra-334 in aqueous solution. J Photochem Photobiol B Biol. 2000;56:139–144. doi: 10.1016/S1011-1344(00)00066-X. [DOI] [PubMed] [Google Scholar]
- Cser K, Vass I. Radiative and non-radiative charge recombination pathways in Photosystem II studied by thermoluminescence and chlorophyll fluorescence in the cyanobacterium Synechocystis 6803. Biochim Biophys Acta. 2007;1767:233–243. doi: 10.1016/j.bbabio.2007.01.022. [DOI] [PubMed] [Google Scholar]
- De la Coba F, Aguilera J, Figueroa F, De Gálvez M, Herrera E. Antioxidant activity of mycosporine-like amino acids isolated from three red macroalgae and one marine lichen. J Appl Phycol. 2009;21:161–169. doi: 10.1007/s10811-008-9345-1. [DOI] [Google Scholar]
- Desikachary TV. Cyanophyta. New Delhi: Indian Council of Agriculture Research; 1959. pp. 222–223. [Google Scholar]
- Figueroa FL, Conde-Alvarez R, Gómez I. Relations between electron transport rates determined by pulse amplitude modulated chlorophyll fluorescence and oxygen evolution in macroalgae under different light conditions. Photosynth Res. 2003;75:259–275. doi: 10.3354/meps151081. [DOI] [PubMed] [Google Scholar]
- Fiorda Giordanino M, Strauch SM, Villafañe VE, Helbling EW. Influence of temperature and UVR on photosynthesis and morphology of four species of cyanobacteria. J Photochem Photobiol B Biol. 2011;103:68–77. doi: 10.1016/j.jphotobiol.2011.01.013. [DOI] [PubMed] [Google Scholar]
- Fischer WF. Life before the rise of oxygen. Nature. 2008;455:1051–1052. doi: 10.1038/4551051a. [DOI] [PubMed] [Google Scholar]
- Fragoso GM, Neale PJ, Kana TM, Pritchard AL. Kinetics of photosynthetic response to ultraviolet and photosynthetically active radiation in Synechococcus WH8102 (cyanobacteria) Photochem Photobiol. 2014;90:522–532. doi: 10.1111/php.12202. [DOI] [PubMed] [Google Scholar]
- Fridovich I. Superoxide dismutase. Adv Enzymol Relat Areas Mol Biol. 1974;41:35–97. doi: 10.1002/9780470122860.ch2. [DOI] [PubMed] [Google Scholar]
- Friso G, Spetea C, Giacometti GM, Vass I, Barbato R. Degradation of photosystem II reaction center D1-protein induced by UVB radiation in isolated thylakoids. Identification and characterization of C- and N-terminal breakdown products. Biochim Biophys Acta. 1994;1184:78–84. doi: 10.1016/0005-2728(94)90156-2. [DOI] [Google Scholar]
- Gao K, Yu H, Brown MT. Solar PAR and UV radiation affects the physiology and morphology of the cyanobacterium Anabaena sp. PCC 7120. J Photochem Photobiol B Biol. 2007;89:117–124. doi: 10.1016/j.jphotobiol.2007.09.006. [DOI] [PubMed] [Google Scholar]
- Garcia-Pichel F, Belnap J. Microenvironments and microscale productivity of cyanobacterial desert crusts. J Phycol. 1996;32:774–782. doi: 10.1111/j.0022-3646.1996.00774.x. [DOI] [Google Scholar]
- Garcia-Pichel F, Castenholz RW. Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Appl Environ Microbiol. 1993;59:163–169. doi: 10.1128/aem.59.1.163-169.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Pichel F, Wingard CE, Castenholz RW. Evidence regarding the UV sunscreen role of a mycosporine-like compound in the cyanobacterium Gloeocapsa sp. Appl Environ Microbiol. 1993;59:170–176. doi: 10.1128/aem.59.1.170-176.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golden SS. Light-responsive gene expression in cyanobacteria. J Bact. 1995;177:1651–1654. doi: 10.1128/jb.177.7.1651-1654.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray E, Stein T (2019) Ozone hole is the smallest on record since its discovery https://www.nasa.gov/feature/goddard/2019/2019-ozone-hole-is-the-smallest-on-record-since-its-discovery. Accessed 21 Oct 2019
- Gutu A, Kehoe DM. Emerging perspectives on the mechanisms, regulation, and distribution of light color acclimation in cyanobacteria. Mol Plant. 2012;5:1–13. doi: 10.1093/mp/ssr054. [DOI] [PubMed] [Google Scholar]
- Häder D-P, Sinha RP. Solar ultraviolet radiation-induced DNA damage in aquatic organisms: potential environmental impact. Mut Res. 2005;571:221–233. doi: 10.1016/j.mrfmmm.2004.11.017. [DOI] [PubMed] [Google Scholar]
- Häder D-P, Williamson CE, Wängberg S-A, Rautio M, Rose KC, Gao K, Helbling EW, Sinha RP, Worrest R. Effects of UV radiation on aquatic ecosystems and interactions with other environmental factors. Photochem Photobiol Sci. 2015;14:108–126. doi: 10.1039/C4PP90035A. [DOI] [PubMed] [Google Scholar]
- Halliwell B. Reactive species and antioxidants: redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006;141:312–322. doi: 10.1104/pp.106.077073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanelt D, Roleda MY. UVB radiation may ameliorate photoinhibition in specific shallow-water tropical marine macrophytes. Aquat Bot. 2009;91:6–12. doi: 10.1016/j.aquabot.2008.12.005. [DOI] [Google Scholar]
- Hargreaves A, Taiwo FA, Duggan O, Kirk SH, Ahmad SI. Near-ultraviolet photolysis of β-phenylpyruvic acid generates free radicals and results in DNA damage. J Photochem Photobiol B Biol. 2007;89:110–116. doi: 10.1016/j.jphotobiol.2007.09.007. [DOI] [PubMed] [Google Scholar]
- He YY, Häder D-P. Reactive oxygen species and UV-B: effect on cyanobacteria. Photochem Photobiol Sci. 2002;1:729–736. doi: 10.1039/B110365M. [DOI] [PubMed] [Google Scholar]
- He YY, Häder D-P. UV-B-induced formation of reactive oxygen species and oxidative damage of the cyanobacterium Anabaena sp.: protective effects of ascorbic acid and N-acetyl-l-cysteine. J Photochem Photobiol B Biol. 2002;66:115–124. doi: 10.1016/S1011-1344(02)00231-2. [DOI] [PubMed] [Google Scholar]
- He YY, Klisch M, Häder D-P. Adaptation of cyanobacteria to UV-B stress correlated with oxidative stress and oxidative damage. Photochem Photobiol. 2002;76:188–196. doi: 10.1562/0031-8655(2002)076<0188:aoctub>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Helbling EW, Gao K, Ai H, Ma Z, Villafañe VE. Differential responses of Nostoc sphaeroides and Arthrospira platensis to solar ultraviolet radiation exposure. J Appl Phycol. 2006;18:57–66. doi: 10.1007/s10811-005-9015-5. [DOI] [Google Scholar]
- Kannaujiya VK, Sinha RP. Impacts of diurnal variation of ultraviolet-B and photosynthetically active radiation on phycobiliproteins of the hot-spring cyanobacterium Nostoc sp. strain HKAR-2. Protoplasma. 2017;254:423–433. doi: 10.1007/s00709-016-0964-0. [DOI] [PubMed] [Google Scholar]
- Karsten U, Kartha KK, Lembcke S, Schumann R. The effects of ultraviolet radiation on photosynthetic performance, growth and sunscreen compounds aeroterrestrial biofilm algae isolated from building facades. Planta. 2007;225:991–1000. doi: 10.1007/s00425-006-0406-x. [DOI] [PubMed] [Google Scholar]
- Kehoe DM, Gutu A. Responding to color: the regulation of complementary chromatic adaptation. Annu Rev Plant Biol. 2006;57:127–150. doi: 10.1146/annurev.arplant.57.032905.105215. [DOI] [PubMed] [Google Scholar]
- Kirilovsky D. Photoprotection in cyanobacteria: the orange carotenoid protein (OCP)-related non-photochemical-quenching mechanism. Photosynth Res. 2007;93:7–16. doi: 10.1007/s11120-007-9168-y. [DOI] [PubMed] [Google Scholar]
- Klisch M, Sinha RP, Richter PR, Häder D-P. Mycosporine-like amino acids (MAAs) protect against UV-B-induced damage in Gyrodinium dorsum Kofoid. J Plant Physiol. 2001;158:1449–1454. doi: 10.1078/0176-1617-00552. [DOI] [Google Scholar]
- Klisch M, Sinha RP, Helbling EW, Häder D-P. Induction of thymine dimers by solar radiation in natural freshwater phytoplankton assemblages in Patagonia, Argentina. Aquat Sci. 2005;67:72–78. doi: 10.1007/s00027-004-0770-1. [DOI] [Google Scholar]
- Kós PB, Deák Z, Cheregi O, Vass I. Differential regulation of psbA and psbD gene expression, and the role of the different D1 protein copies in the cyanobacterium Thermosynechococcus elongatus BP-1. Biochim Biophys Acta. 2008;1777:74–83. doi: 10.1016/j.bbabio.2007.10.015. [DOI] [PubMed] [Google Scholar]
- Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–1874. doi: 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lesser MP. Oxidative stress in tropical marine ecosystems. In: Abele D, Vázquez-Medina JP, Zenteno-Savín T, editors. Oxidative stress in aquatic ecosystems. Hoboken: Wiley-Blackwell; 2011. pp. 7–19. [Google Scholar]
- Li X. Improved pyrogallol autoxidation method: a reliable and cheap superoxide-scavenging assay suitable for all antioxidants. J Agric Food Chem. 2012;60:6418–6424. doi: 10.1021/jf204970r. [DOI] [PubMed] [Google Scholar]
- Maehly A, Chance B. Assay of catalases and peroxidases, in methods in enzymology. Methods Enzymol. 1955;2:764–775. doi: 10.1016/s0076-6879(55)02300-8. [DOI] [Google Scholar]
- Marwood CA, Greenberg BM. Effect of supplementary UVB radiation on chlorophyll synthesis and accumulation of photosystems during chloroplast development in Spirodela oligorrhiza. Photochem Photobiol. 1996;64:664–670. doi: 10.1111/j.1751-1097.1996.tb03121.x. [DOI] [Google Scholar]
- Miller NJ, Rice-Evans C. Factor influencing the antioxidant activity determined by the ABTS+ radical cation assay. Free Radic Res. 1997;26:195–199. doi: 10.3109/10715769709097799. [DOI] [PubMed] [Google Scholar]
- Müller P, Li XP, Niyogi KK. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001;125:1558–1566. doi: 10.1104/pp.125.4.1558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson N, Ben-Shem A. The structure of photosystem I and evolution of photosynthesis. BioEssays. 2005;27:914–922. doi: 10.1002/bies.20278. [DOI] [PubMed] [Google Scholar]
- Ogawa T, Misumi M, Sonoike K. Estimation of photosynthesis in cyanobacteria by pulse-amplitude modulation chlorophyll fluorescence: problems and solutions. Photosynth Res. 2017;133:63–73. doi: 10.1007/s11120-017-0367-x. [DOI] [PubMed] [Google Scholar]
- Olsson-Francis K, Watson JS, Cockell CS. Cyanobacteria isolated from the high-intertidal zone: a model for studying the physiological prerequisites for survival in low Earth orbit. Int J Astrobiol. 2013;12:292–303. doi: 10.1017/S1473550413000104. [DOI] [Google Scholar]
- Oyamada C, Kaneniwa M, Ebitani K, Murata M, Ishihara K. Mycosporine-like amino acids extracted from scallop (Patinopecten yessoensis) ovaries: UV protection and growth stimulation activities on human cells. Mar Biotechnol. 2008;10:141–150. doi: 10.1007/s10126-007-9043-z. [DOI] [PubMed] [Google Scholar]
- Pathak J, Rajneesh, Ahmed H, Richa, Sinha RP. Metabolomic profiling of cyanobacterial UV-protective compounds. Curr Metab. 2017;5:138–163. doi: 10.2174/2213235X04666160829162640. [DOI] [Google Scholar]
- Pathak J, Rajneesh, Maurya PK, Singh SP, Häder D-P, Sinha RP. Cyanobacterial farming for environment friendly sustainable agriculture practices: innovations and perspectives. Frontiers Environ Sci. 2018;6:7. doi: 10.3389/fenvs.2018.00007. [DOI] [Google Scholar]
- Pathak J, Ahmed H, Singh SP, Häder D-P, Sinha RP. Genetic regulation of scytonemin and mycosporine-like amino acids (MAAs) biosynthesis in cyanobacteria. Plant Gene. 2019;17:100172. doi: 10.1016/j.plgene.2019.100172. [DOI] [Google Scholar]
- Pathak J, Singh PR, Häder DP, Sinha RP. UV-induced DNA damage and repair: a cyanobacterial perspective. Plant Gene. 2019;19:100194. doi: 10.1016/j.plgene.2019.100194. [DOI] [Google Scholar]
- Pattanaik B, Roleda MY, Schumann R, Karsten U. Isolate-specific effects of ultraviolet radiation on photosynthesis, growth and mycosporine-like amino acids in the microbial mat-forming cyanobacterium Microcoleus chthonoplastes. Planta. 2008;227:907–916. doi: 10.1007/s00425-007-0666-0. [DOI] [PubMed] [Google Scholar]
- Plack PA, Fraser NW, Grant PT, Middleton C, Mitchell AI, Thomson RH. Gadusol, an enolic derivative of cyclohexane-1,3-dione present in the roes of cod and other marine fish. Isolation, properties and occurrence compared with ascorbic acid. Biochem J. 1981;199:741–747. doi: 10.1042/bj1990741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pope MA, Spence E, Seralvo V, Gacesa R, Heidelberger S, Weston AJ, Dunlap WC, Shick JM, Long PF. O-Methyltransferase is shared between the pentose phosphate and shikimate pathways and is essential for mycosporine-like amino acid biosynthesis in Anabaena variabilis ATCC 29413. ChemBioChem. 2015;16:320–327. doi: 10.1002/cbic.201402516. [DOI] [PubMed] [Google Scholar]
- Quesada A, Vincent WF. Strategies of adaptation by Antarctic cyanobacteria to ultraviolet radiation. Eur J Phycol. 1997;32:335–342. doi: 10.1080/09670269710001737269. [DOI] [Google Scholar]
- Rajneesh, Singh SP, Pathak J, Sinha RP. Cyanobacterial factories for the production of green energy and value-added products: an integrated approach for economic viability. Renew Sustain Energ Rev. 2017;69:578–595. doi: 10.1016/j.rser.2016.11.110. [DOI] [Google Scholar]
- Rajneesh, Pathak J, Richa, Häder D-P, Sinha RP. Impacts of ultraviolet radiation on certain physiological and biochemical processes in cyanobacteria inhabiting diverse habitats. Environ Exp Bot. 2019;161:375–387. doi: 10.1016/j.envexpbot.2018.10.037. [DOI] [Google Scholar]
- Rascher U, Lakatos M, Büdel B, Lüttge U. Photosynthetic field capacity of cyanobacteria of a tropical inselberg of the Guiana Highlands. Eur J Phycol. 2003;38:247–256. doi: 10.1080/0967026031000121679. [DOI] [Google Scholar]
- Rastogi RP, Incharoensakdi A. Occurrence and induction of a ultraviolet-absorbing substance in the cyanobacterium Fischerellamuscicola TISTR8215. Phycol Res. 2015;63:51–55. doi: 10.1111/pre.12069. [DOI] [Google Scholar]
- Rastogi RP, Sinha RP. Biotechnological and industrial significance of cyanobacterial secondary metabolites. Biotechnol Adv. 2009;27:521–539. doi: 10.1016/j.biotechadv.2009.04.009. [DOI] [PubMed] [Google Scholar]
- Rastogi RP, Singh SP, Häder D-P, Sinha RP. Detection of reactive oxygen species (ROS) by the oxidant-sensing probe 2ʹ,7ʹ-dichlorodihydrofluorescein diacetate in the cyanobacterium Anabaena variabilis PCC 7937. Biochem Biophys Res Commun. 2010;397:603–607. doi: 10.1016/j.bbrc.2010.06.006. [DOI] [PubMed] [Google Scholar]
- Rastogi RP, Singh SP, Häder D-P, Sinha RP. Mycosporine-like amino acids profile and their activity under PAR and UVR in a hot-spring cyanobacterium Scytonema sp. HKAR-3. Aust J Bot. 2010;58:286–293. doi: 10.1071/bt10004. [DOI] [Google Scholar]
- Rastogi RP, Singh SP, Häder D-P, Sinha RP. Ultraviolet-B-induced DNA damage and photorepair in the cyanobacterium Anabaena variabilis PCC 7937. Environ Exp Bot. 2011;74:280–288. doi: 10.1016/j.envexpbot.2011.06.010. [DOI] [Google Scholar]
- Rastogi RP, Kumari S, Richa, Han T, Sinha RP. Molecular characterization of hot spring cyanobacteria and evaluation of their photoprotective compounds. Can J Microbiol. 2012;58:719–727. doi: 10.1139/w2012-044. [DOI] [PubMed] [Google Scholar]
- Rastogi RP, Incharoensakdi A, Madamwar D. Responses of a rice-field cyanobacterium Anabaena siamensis TISTR-8012 upon exposure to PAR and UV radiation. J Plant Physiol. 2014;171:1545–1553. doi: 10.1016/j.jplph.2014.07.011. [DOI] [PubMed] [Google Scholar]
- Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice-Evans C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol Med. 1999;26:1231–1237. doi: 10.1016/S0891-5849(98)00315-3. [DOI] [PubMed] [Google Scholar]
- Richa, Sinha RP. Biochemical characterization of sunscreening mycosporine-like amino acids from two Nostoc species inhabiting diverse habitats. Protoplasma. 2015;252:199–208. doi: 10.1007/s00709-014-0674-4. [DOI] [PubMed] [Google Scholar]
- Rippka R, Deruelles J, Wazterbury JB, Herdmann M, Stanier RY. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J Gen Microbiol. 1979;111:1–61. doi: 10.1099/00221287-111-1-1. [DOI] [Google Scholar]
- Roleda MY, Nyberg CD, Wulff A. UVR defense mechanisms in eurytopic and invasive Gracilaria vermiculophylla (Gracilariales, Rhodophyta) Physiol Plant. 2012;146:205–216. doi: 10.1111/j.1399-3054.2012.01615.x. [DOI] [PubMed] [Google Scholar]
- Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- Sander J, Nowaczyk M, Buchta J, Dau H, Vass I, Deák Z, Dorogi M, Iwai M, Rögner M. Functional characterization and quantification of the alternative PsbA copies in Thermosynechococcus elongatus and their role in photoprotection. J Biol Chem. 2010;285:29851–29856. doi: 10.1016/j.bbabio.2010.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sass L, Spetea C, Mate Z, Nagy F, Vass I. Repair of UV-B induced damage of Photosystem II via de novo synthesis of the D1 and D2 reaction centre subunits in Synechocystis sp. PCC 6803. Photosynth Res. 1997;54:55–62. doi: 10.1023/A:1005895924892. [DOI] [Google Scholar]
- Singh H. Desiccation and radiation stress tolerance in cyanobacteria. J Basic Microbiol. 2018;58:813–826. doi: 10.1002/jobm.201800216. [DOI] [PubMed] [Google Scholar]
- Singh SP, Häder D-P, Sinha RP. Cyanobacteria and ultraviolet radiation (UVR) stress: mitigation strategies. Ageing Res Rev. 2010;9:79–90. doi: 10.1016/j.arr.2009.05.004. [DOI] [PubMed] [Google Scholar]
- Singh A, Sarkar A, Singh S, Agrawal S. Investigation of supplemental ultraviolet-B-induced changes in antioxidative defense system and leaf proteome in radish (Raphanussativus L. cv Truthful): an insight to plant response under high oxidative stress. Protoplasma. 2010;245:75–83. doi: 10.1007/s00709-010-0138-4. [DOI] [PubMed] [Google Scholar]
- Singh G, Babele PK, Sinha RP, Tyagi MB, Kumar A. Enymatic and non-enzymatic defense mechanisms against ultraviolet-B radiation in two Anabaena species. Process Biochem. 2013;48:796–802. doi: 10.1016/j.procbio.2013.04.022. [DOI] [Google Scholar]
- Singh SP, Rastogi RP, Sinha RP, Häder D-P. Photosynthetic performance of Anabaena variabilis PCC 7937 under simulated solar radiation. Photosynthetica. 2013;51:259–266. doi: 10.1007/s11099-013-0012-7. [DOI] [Google Scholar]
- Singh JS, Kumar A, Rai AN, Singh DP. Cyanobacteria: a precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front Microbiol. 2016;7:529. doi: 10.3389/fmicb.2016.00529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh A, Tyagi MB, Kumar A. Cyanobacteria growing on tree barks possess high amount of sunscreen compound mycosporine-like amino acids (MAAs) Plant Physiol Biochem. 2017;119:110–120. doi: 10.1016/j.plaphy.2017.08.020. [DOI] [PubMed] [Google Scholar]
- Sinha RP, Kumar HD, Kumar A, Häder D-P. Effects of UV-B irradiation on growth, survival, pigmentation and nitrogen metabolism enzymes in cyanobacteria. Acta Protozool. 1995;34:187–192. [Google Scholar]
- Sinha RP, Singh N, Kumar A, Kumar HD, Häder M, Häder D-P. Effects of UV irradiation on certain physiological and biochemical processes in cyanobacteria. J Photochem Photobiol B Biol. 1996;32:107–113. doi: 10.1016/1011-1344(95)07205-5. [DOI] [Google Scholar]
- Sinha RP, Klisch M, Vaishampayan A, Häder D-P. Biochemical and spectroscopic characterization of the cyanobacterium Lyngbya sp. inhabiting mango (Mangifera indica) trees: presence of an ultraviolet-absorbing pigment, scytonemin. Acta Protozool. 1999;38:291–298. [Google Scholar]
- Sinha RP, Singh SP, Häder D-P. Database on mycosporines and mycosporine-like amino acids (MAAs) in fungi, cyanobacteria, macroalgae, phytoplankton and animals. J Photochem Photobiol B Biol. 2007;89:29–35. doi: 10.1016/j.jphotobiol.2007.07.006. [DOI] [PubMed] [Google Scholar]
- Sinha RP, Rastogi RP, Ambasht NK, Häder D-P. Live of wetland cyanobacteria under enhancing solar UV-B radiation. Proc Natl Acad Sci India B. 2008;78:53–65. [Google Scholar]
- Sonkar PK, Ganesan V. Synthesis and characterization of silver nanoparticle-anchored amine-functionalized mesoporous silica for electrocatalytic determination of nitrite. J Solid State Electrochem. 2015;19:2107–2115. doi: 10.1007/s10008-014-2725-3. [DOI] [Google Scholar]
- Sonkar PK, Prakash K, Yadav M, Ganesan V, Sankar M, Gupta R, Yadav DK. Co(II)-porphyrin-decorated carbon nanotubes as catalysts for oxygen reduction reactions: an approach for fuel cell improvement. J Mater Chem A. 2017;5:6263–6276. doi: 10.1039/c6ta10482g. [DOI] [Google Scholar]
- Sugiura M, Kato Y, Takahashi R, Suzuki H, Watanabe T, Noguchi T, Rappaport F, Boussac A. Energetics in Photosystem II from Thermosynechococcus elongatus with a D1 protein encoded by either the psbA1 or psbA3 gene. Biochim Biophys Acta. 2010;1797:1491–1499. doi: 10.1016/j.bbabio.2010.03.022. [DOI] [PubMed] [Google Scholar]
- Şükran D, Güneş T, Sivaci R. Spectrophotometric determination of chlorophyll-A, B and total carotenoid contents of some algae species using different solvents. Turk J Bot. 1998;22:13–18. [Google Scholar]
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28:2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thajuddin N, Subramanian G. Cyanobacterial biodiversity and potential applications in biotechnology. Curr Sci. 2005;89:47–57. [Google Scholar]
- Thapar R, Srivastava AK, Bhargava P, Mishra Y, Rai LC. Impact of different abiotic stresses on growth, photosynthetic electron transport chain, nutrient uptake and enzyme activities of Cu-acclimated Anabaena doliolum. J Plant Physiol. 2008;165:306–316. doi: 10.1016/j.jplph.2007.05.002. [DOI] [PubMed] [Google Scholar]
- Tichý M, Lupınková L, Sicora C, Vass I, Kuvikova S, Prášil O, Komenda J. Synechocystis 6803 mutants expressing distinct forms of the photosystem II D1 protein from Synechococcus 7942: relationship between the psbA coding region and sensitivity to visible and UV-B radiation. Biochim Biophys Acta. 2003;1605:55–66. doi: 10.1016/S0005-2728(03)00064-1. [DOI] [PubMed] [Google Scholar]
- Torres P, Santos JP, Chow F, Ferreira MJP, dos Santos DY. Comparative analysis of in vitro antioxidant capacities of mycosporine-like amino acids (MAAs) Algal Res. 2018;34:57–67. doi: 10.1016/j.algal.2018.07.007. [DOI] [Google Scholar]
- Tripathi S, Srivastava P. Presence of stable active oxygen scavenging enzymes superoxide dismutase, ascorbate peroxidase and catalase in a desiccation-tolerant cyanobacterium Lyngbya arboricola under dry state. Curr Sci. 2001;81:197–200. [Google Scholar]
- Tyystjärvi E. Photoinhibition of photosystem II and photodamage of the oxygen evolving manganese cluster. Coord Chem Rev. 2008;252:361–376. doi: 10.1016/j.ccr.2007.08.021. [DOI] [Google Scholar]
- Vass I. Role of charge recombination processes in photodamage and photoprotection of the photosystem II complex. Physiol Plant. 2011;142:6–16. doi: 10.1111/j.1399-3054.2011.01454.x. [DOI] [PubMed] [Google Scholar]
- Vass I. Molecular mechanisms of photodamage in the photosystem II complex. Biochim Biophys Acta. 2012;1817:209–217. doi: 10.1016/j.bbabio.2011.04.014. [DOI] [PubMed] [Google Scholar]
- Vincent WF, Quesada A. Ultraviolet radiation effects on cyanobacteria: implication for Antarctic microbial ecosystems. In: Weiler CS, Penhale PA, editors. Antarctic research series: ultraviolet radiation in antarctica: measurements and biological effects. Washington: American Geophysical Union; 1994. pp. 111–124. [Google Scholar]
- Wada N, Sakamoto T, Matsugo S. Mycosporine-like amino acids and their derivatives as natural antioxidants. Antioxidants. 2015;4:603–646. doi: 10.3390/antiox4030603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang G, Chen K, Chen L, Hu C, Zhang D, Liu Y. The involvement of the antioxidant system in protection of desert cyanobacterium Nostoc sp. against UV-B radiation and the effects of exogenous antioxidants. Ecotoxicol Environ Saf. 2008;69:150–157. doi: 10.1016/j.ecoenv.2006.03.014. [DOI] [PubMed] [Google Scholar]
- Wang Y, Qu T, Zhao X, Tang X, Xiao H, Tang X. A comparative study of the photosynthetic capacity in two green tide macroalgae using chlorophyll fluorescence. Springer Plus. 2016;5:775. doi: 10.1186/s40064-016-2488-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitehead K, Hedges JI. Electrospray ionization tandem mass spectrometric and electron impact mass spectrometric characterization of mycosporine-like amino acids. Rapid Commun Mass Spectrom. 2003;17:2133–2138. doi: 10.1002/rcm.1162. [DOI] [PubMed] [Google Scholar]
- Whitton BA. Ecology of cyanobacteria II: Their Diversity In Space And Time. Dordrecht: Springer Science & Business Media; 2012. [Google Scholar]
- Wilson A, Ajlani G, Verbavatz JM, Vass I, Kerfeld CA, Kirilovsky D. A soluble carotenoid protein involved in phycobilisome-related energy dissipation in cyanobacteria. Plant Cell. 2006;18:992–1007. doi: 10.1105/tpc.105.040121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu J, Gao K. UV-A enhanced growth and UV-B induced positive effects in the recovery of photochemical yield in Gracilaria lemaneiformis (Rhodophyta) J Photochem Photobiol B Biol. 2010;100:117–122. doi: 10.1016/j.jphotobiol.2010.05.010. [DOI] [PubMed] [Google Scholar]
- Young AJ. The photoprotective role of carotenoids in higher plants. Physiol Plant. 1991;83:702–708. doi: 10.1111/j.1399-3054.1991.tb02490.x. [DOI] [Google Scholar]
- Zeeshan M, Prasad SM. Differential response of growth, photosynthesis, antioxidant enzymes and lipid peroxidation to UV-B radiation in three cyanobacteria. S Afr J Bot. 2009;75:466–474. doi: 10.1016/j.sajb.2009.03.003. [DOI] [Google Scholar]