

Abstract
Cryptorchidism is one of the main risk factors for infertility and testicular cancer. Orchiopexy surgery corrects cryptorchidism effects. Different models of cryptorchidism developed in the rat include surgery. We assessed testicular alterations in rats submitted to surgical cryptorchidism and examined their potential for reversibility at different time points in order to verify time dependency effect(s) on the recovery of the undescended testes. Cryptorchidism was induced in 3‐week‐old rats. Animals were euthanized 3, 6 or 11 weeks after surgery to evaluate the morphological progression of cryptorchidism‐induced germinative epithelial alterations. Other groups underwent orchiopexy 3, 5 or 9 weeks after surgical cryptorchidism, before or after puberty. Animals were euthanized 3 or 8 weeks after orchiopexy. Controls underwent sham surgery at the same time points as the surgical groups. Cryptorchid testes showed decreased weight, germinative epithelial degeneration, apoptosis and vacuolation, corresponding to impairment of spermatogenesis and of Sertoli cells. Some tubules has a Sertoli cell‐only pattern and atrophy. The intensity of damage was related to the duration of cryptorchidism. After orchiopexy, spermatogenesis completely recovered only when testicular relocation occurred before puberty and the interval for recovery was extended. These results indicate that age, sexual maturity and extension of germ cell damage were relevant for producing germ cell restoration and normal spermatogenesis. We provide original observations on the time dependency of testicular alterations induced by cryptorchidism and their restoration using morphologic, morphometric and immunohistochemical approaches. It may be useful to study germ cell impairment, progression and recovery in different experimental settings, including exposure to exogenous chemicals.
Keywords: cryptorchidism, germ cell damage, orchiopexy, testicular damage
1. INTRODUCTION
Failure of testicular descent, cryptorchidism, is a common anomaly that affects 2% to 4% of newborn boys, involving both testes in approximately 18% of cases. The incidence of this condition has increased in several countries in recent years and have been associated with risk factors such as parental lifestyle and/or exposure to environmental contaminants, possibly resulting in genetic abnormalities. 1 , 2 , 3 Accordingly, it has been proposed that cryptorchidism, low sperm counts, hypospadias and testicular germ cell tumours have a common origin during foetal life and comprise, individually or jointly, what has been termed the testicular dysgenesis syndrome (TDS). 4 , 5 An undescended testis exposed to the inner body temperature may induce heat‐induced changes in testicular cell populations and hormones, leading to perturbed spermatogenesis. 6 The most conspicuous morphological alteration in cryptorchid testes that has been observed in young men is atrophy of seminiferous tubules and variable depletion of germ cells via apoptosis, with apparent hyperplasia of Leydig cells in the interstitium. 7 , 8 Intratubular Sertoli cells, critically involved with spermatogenesis, may also be variably compromised in cryptorchidism; they may become vacuolated, reduced in number and/or appear to be immature in long‐term cryptorchid testes. 9 , 10
There are a variety of methods to induce experimental cryptorchidism in rodents that have provided better understanding of the physiopathology of the condition. 11 , 12 Exogenous anti‐androgens like flutamide 13 , 14 and dibutyl‐phthalate (DBP) administered during gestation 15 or hormones such as oestradiol subcutaneously applied after birth 16 can induce cryptorchidism in rodents. DBP has recently been reported to impair expression levels of essential genes for testicular normal development. 17 Besides chemical procedures, congenital and genetically engineered models have also been used to study undescended testis. 18 , 19 In transgenic mice, over‐expression of aromatase (CYP19A1) and insulin‐like 3 (INSL‐3) hormone, and in knockout mice for the luteinizing hormone receptor (LHR) gene and the thyroid‐specific enhancer/binding protein T/ebp/Nkx2.1 transcription factor, cryptorchidism seems to occur either as a single manifestation or in combination with other abnormalities such as hormonal imbalances. 19 The Long‐Evans/Cryptorchid (LE/Orl) rat 20 , 21 and the Trans‐scrotal (TS) rat 22 are natural congenital models of cryptorchidism. These natural and chemically or hormonally induced models of cryptorchidism have the potential disadvantage of complex systemic perturbation of animal homeostasis and physiological pathways.
To study specifically the testicular alterations related to the male gonad dystopia, some postnatal surgical models of cryptorchidism have been adopted. 23 , 24 , 25 Our laboratory established a successful surgical model for bilateral cryptorchidism by fixing the testes to the inner abdominal wall at the earliest moment possible after birth, postnatal day (PND) 21. 26 The testicular descent in rodents is not complete until after 2‐3 weeks of age in the mouse and 3‐4 weeks in rats, in contrast to humans. Therefore, the onset of a potential thermal injury begins at puberty, rather than at birth, in the rodent. 27 , 28 Alterations observed in these models of experimental cryptorchidism are similar to those found in the human condition and include reduction in testicular weight, decreased seminiferous tubule diameter, degeneration of germinative epithelium and dysfunction of Sertoli and Leydig cells. 12 , 26 , 29 , 30 , 31 Increased incidence of vacuoles within the seminiferous epithelium, a morphological indication of Sertoli cell disturbance, have also been observed in cryptorchid testes of rats. 32 Furthermore, positive immunostaining for CK18, a biomarker of immature Sertoli cells, as observed in the testes of Rhesus monkeys made cryptorchid, suggests lack of differentiation of these cells in the context of gonadal dystopia. 33
Experimentally cryptorchid rats should undergo orchiopexy (surgery that relocates the testes to the scrotum) as early as possible to re‐establish normal spermatogenesis and maintain fertility. 23 , 30 , 34 However, a comprehensive and sequential description of the rat seminiferous tubule alterations resulting from cryptorchidism and orchiopexy is lacking in the literature. The present experimental study with rats was developed to characterize the progression of changes induced by cryptorchidism in the seminiferous tubules, utilizing a model of surgical cryptorchidism and orchiopexy established by our laboratory. 26 Additionally, three different time points for performing orchiopexy and two later intervals for evaluation of gonadal recovery were assessed. The effectiveness of the cryptorchid and orchiopexy procedures and the influence of orchiopexy timing were evaluated by qualitative and semi‐quantitative estimates of the seminiferous tubule components. The influences of the surgical procedures on apoptosis and cell proliferation were also evaluated quantitatively. This study provides original observations on the time dependency of testicular alterations resulting from surgical cryptorchidism and their restoration by orchiopexy using morphological, morphometrical and immunohistochemical approaches.
2. MATERIALS AND METHODS
2.1. General
Six‐week‐old Sprague‐Dawley rats weighing between 150 and 180 g were obtained from the Multidisciplinary Center for Biological Investigation (CEMIB, UNICAMP, Campinas, São Paulo, Brazil) and placed in the animal facility under a 12‐hours light/dark cycle and controlled temperature (22 ± 2°C) and humidity (55 ± 10%). Standard pelleted food (Presence; Evialis) and tap water were provided ad libitum at all times during the study. After a 4‐week acclimation period, two adult female rats were paired overnight to one male in the home cage of the male. The day of mating evidence (sperm‐positive vaginal smear or copulation plug) was considered as day 0 of gestation. All females were allowed to deliver naturally. The day that parturition was judged to be complete was designated postnatal day 0 (PND 0). The offspring were weaned at PND 21 when they were operated for cryptorchidism and randomly allocated to the experimental groups.
2.2. Experimental design
At weaning (PND 21), bilateral abdominal cryptorchidism (CPT) was performed on male rats as described below (Figure 1). Selected cryptorchid animals were euthanized 3, 6 or 11 weeks later (groups CPT + 3, CTP + 6 and CPT + 11, respectively). Other groups underwent orchiopexy at the 3rd (ORC3), 5th (ORC5) or 9th (ORC9) week after the cryptorchidism procedure. Therefore, the surgeries were performed at PND 42, PND 56 and PND 84, respectively. Considering that rat puberty occurs in the range from PND 38 to PND 55, and sexual maturity at approximately the 6th week (PND 42), 28 it can be assumed that the ORC3 animals were at puberty, and the ORC5 and ORC9 rats were young adult animals, ie, underwent orchiopexies after puberty. These animals were euthanized at two different times during their adulthood, either three (groups ORC3 + 3, ORC5 + 3 and ORC9 + 3) or eight (groups ORC3 + 8, ORC5 + 8 and ORC9 + 8) weeks after the orchiopexies. Control animals were sham‐operated at the same times as the cryptorchidism or orchiopexy surgical procedures. For each surgical treatment and control group, at least 10 and 5 animals were used, respectively. For euthanasia, the rats were anaesthetized with ketamine (30 mg/kg i.p.) and xylazine (4 mg/kg i.p.) between 0800 and 1000 hours and exsanguinated by heart puncture. The testes and epididymides were immediately removed, weighed and placed in modified Davidson's fixative for 24 hours. 35 , 36 After fixation, the organs were processed and embedded in paraffin.
FIGURE 1.
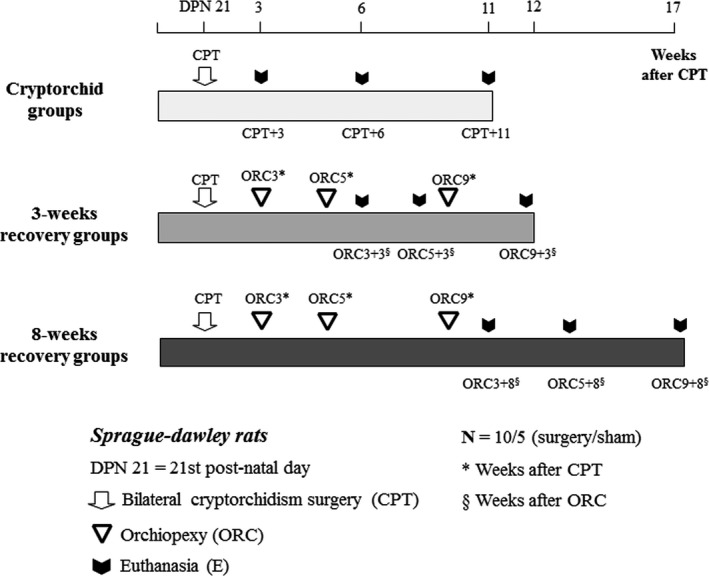
Experimental design. Sprague‐Dawley rats were surgically made cryptorchid at the weaning (3rd week of age or DPN 21st postnatal day). Some cryptorchid groups (CPT) were euthanized 3, 6 or 11 wks after the surgeries (CPT + 3, CPT + 6, CPT + 11, respectively). For evaluation of reversibility of testicular alterations induced by cryptorchidism, other cryptorchid groups were submitted to orchiopexy (ORC) 3, 5 or 9 wks after cryptorchidism (ORC3, ORC5 and ORC9, respectively) and euthanized after 3 or 8 wks. Corresponding sham‐operated controls (not shown) were euthanized simultaneously
2.3. Surgery to establish the cryptorchidism and orchiopexy
The procedures described by Souza et al 26 were followed. Briefly, on PND 21, rats weighing between 60 and 70 g were anaesthetized with ketamine 30 mg/kg i.p. and xylazine 4 mg/kg i.p. Abdominal wall anaesthesia and analgesic effects were achieved with local injections of lidocaine (7 mg/kg, s.c.) and ketoprofen (10 mg/kg, s.c.), respectively. The abdomen was aseptically opened and the testes were translocated from the scrotum into the abdominal cavity through the inguinal rings using tweezers. Each testis and the respective epididymis were separately attached to the inner dorsolateral abdominal wall with two stitches at the cranial and caudal regions, respectively, passing a 5‐0 blunt needle with non‐absorbable suture material (NL50CR13 Nylon, Bioline) with great care through the tunica albuginea. The abdominal wall and skin were closed, and the animals were kept at 30°C for 30 minutes to minimize the deleterious effects of hypothermia induced by anaesthesia.
For orchiopexy, we used the same pre‐ and postsurgery anaesthetic, analgesic and asepsis procedures as used for the cryptorchidism surgery. At each of the scheduled times, a midline abdominal incision was made and the sutures which held the testes attached to the inner posterior abdominal wall were carefully cut. Tweezers were introduced into the scrotum through the inguinal ring so the inner surface of the scrotum wall was clamped and reversed to facilitate the manipulation. The testes were sutured (5‐0 blunt needle, non‐absorbable suture material, NL50CR13 Nylon, Bioline, Ltda.) through the tunica albuginea to the inner surface of the scrotum wall. Finally, using tweezers, the testes were carefully guided into the scrotum. The abdominal wall was closed with 5‐0 triangular tipped needles with non‐absorbable suture material (Nylon, Bioline, Fios Cirúrgicos Ltda.). All animals received antibiotic (enrofloxacin 5 mg/kg s.c.) during the 3 days following surgery.
Control animals underwent sham surgery with the same anaesthesia, analgesia and aseptic conditions as described above, and their abdomens were similarly opened and closed by sutures through the muscular and skin layers.
2.4. Classification of histological alterations
Histological sections of 5 µm thickness were cut from paraffin blocks in such a way that a transverse central section of each testis was obtained and stained with haematoxylin and eosin (H&E). The histologic analyses were systematically and blindly performed by one of the authors (APFC). The seminiferous tubules were analysed as previously described, 26 using a four‐grade classification system based on the most frequent morphologic testicular alterations observed in the H&E sections. All round seminiferous tubules were classified according to the most differentiated predominating germ cell type: Class 1—normal spermatogenesis, containing spermatids and sperms; Class 2—tubules with spermatids and sperms but also showing alterations such as epithelial vacuoles, apoptotic bodies and/or giant germ cells; Class 3—tubules presenting spermatocytes and spermatogonia with the same elements of epithelial damage as Class 2, but more severe and more extensive; and Class 4—Sertoli cell‐only (SCO) tubules. The results obtained by this classification system were then adjusted by an adapted H‐score, the same used for semi‐quantitative evaluations in immunohistochemistry analyses. 37 , 38 The frequency of tubules (%) in each class was calculated by multiplying the number of tubules in that class by 100 and dividing by the total number of tubules analysed. The final H‐score for each group was the result of adding the number of tubules in class 1 times 1, the number of tubules in class 2 times 2, the number of tubules in class 3 times 3 and the number of tubules in class 4 times 4, and dividing each result by the total number of tubules counted in the respective section. In the surgery groups, the minimum, maximum and average number of tubule sections counted was 166, 534 and 358, respectively. In the control groups, these values were 199, 592 and 434, respectively.
2.5. Quantitative evaluations of seminiferous tubule alterations
Quantitative estimates of the seminiferous tubule diameter, number of vacuoles within the seminiferous epithelium and the number of Sertoli cells were performed. Fifty round seminiferous tubules were randomly chosen from each testis for estimating tubular diameters and the incidence of vacuoles. The seminiferous tubule diameters were measured with the use of Panoramic Viewer software (3DHISTECH Ltd). Two perpendicularly opposed measures from the outer face of the tubule basement membrane to the other side of the same tubule section were obtained for each tubule, and the mean value was used to calculate the mean seminiferous tubule diameter (MSTD). 39 Vacuoles were considered present when one or more vacuoles ≥16 μm in greatest diameter occurred within the seminiferous epithelium. 40 Finally, one hundred round seminiferous tubules were randomly selected from each testis to estimate the Sertoli cell number. The number of nuclei was assumed to equate to the number of Sertoli cells. Therefore, Sertoli cells were identified and counted based on their nuclear morphological characteristics. 9
2.6. Mitotic indices (MIs)
The MI of the germinative epithelium was assessed on H&E sections. Five samples from each surgical treatment group and three from each control group were analysed. The MI was calculated by dividing the total number of cells undergoing mitosis by the total number of seminiferous tubules × 100. Immunohistochemical analyses of proliferation markers such as Ki‐67 or PCNA were not performed due to technical limitations, related to the use of Davidson's fixative.
2.7. Caspase‐3 and cytokeratin 18 evaluations
The apoptotic index (caspase‐3) and the presence of cytokeratin 18 (CK18) were immunohistochemically evaluated in testicular sections from five animals from the surgical groups and three animals from the control groups. Sections were deparaffinized, rehydrated and peroxidase, and protein was blocked with peroxidazed 1 (Biocare Medical) and background sniper (Biocare Medical). Tissue sections underwent heat‐induced antigen retrieval. In sequence, tissue sections were incubated overnight at approximately 4°C with the primary antibody anti‐caspase‐3 (rabbit monoclonal, Cell Signalling), or anti‐CK18 (mouse monoclonal, ABCAM, AB668) diluted at 1:10 and 1:400, respectively. The sections were then incubated with a biotinylated second antibody followed by incubation with a peroxidase‐conjugated biotin‐avidin complex (VECTASTAIN Elite ABC Kit, Vector Laboratories) for caspase‐3 and with MACH4 Universal HRP Polymer (Biocare Medical) for CK18 followed by a DAB display system (DAB Substrate kit, Vector Laboratories). A section of human tonsil was used as positive control for caspase staining, and Sertoli cells from a prepubertal rat testis were used as the positive control for CK18. For caspase‐3, the total number of positive cells and the total number of seminiferous tubules in the section were counted. The labelling index (LI) was calculated by dividing the number of caspase‐positive cells by the number of seminiferous tubules x 100. For CK18, the presence or absence of positive Sertoli cells was noted for all seminiferous tubules in the section.
2.8. Statistical analyses
All statistical analyses (except histological analyses) were performed using SigmaStat 3.5 (Aspire Software International). Parameters obtained from the cryptorchidism/orchiopexy groups were compared to their respective control groups. Tests of normality and variance were followed by Student's t test or the Mann‐Whitney test. For histological analyses (IBM SPSS 22.0 Statistics software (Statistical Package for Social Science, SPSS Inc), generalized linear mixed‐effects model with gamma distribution followed by Sidak correction was used. The Wilcoxon rank‐sum test was used for caspase‐3 and for the MI. The ANOVA test was performed among the cryptorchidism/orchiopexy subgroups: 3‐week recovery and 8‐week recovery groups. Statistically significant differences were assumed when P < .05.
3. RESULTS
The surgical procedures for both cryptorchidism and orchiopexy were well tolerated by the animals. There was no mortality due to the surgeries, and no local inflammation and infection were noted afterwards. At necropsy, 4 of the 39 (10%) cryptorchid animals had one atrophic testis, and in 6 of 39 (15%) cryptorchid rats, one of the testes spontaneously returned to the scrotum. During orchiopexy, no adhesion between the abdominal wall and the testes was observed. Gonadal removal and relocation into the scrotum were readily accomplished.
3.1. Body, testes and epididymis weights
No significant differences in body weights were found between groups at any of the experimental time points (data not shown). The mean testicular absolute weights of the control (CTL) sham‐operated animals increased from the 6th week (pubertal animals) to the 9th week of age, when it reached a plateau that was maintained to the end of the experiment at the 20th week of postnatal life (Figure 2A). The mean absolute weights of cryptorchid testes of animals euthanized after three (CPT + 3), six (CPT + 6) or eleven (CPT + 11) weeks after the establishment of the cryptorchidism were, respectively, 50%, 69% and 71% lower than their respective control groups (P < .05) (Figure 2A), indicating that the testicular growth impairment was more severe as the period after the surgery for cryptorchidism lengthened. Three weeks after orchiopexy (or at the 3rd week of recovery), the testes weights of the ORC3 + 3, ORC5 + 3 and ORC9 + 3 groups were similar and significantly (P < .05) lower than their respective control groups, irrespective of whether the surgery was performed 3, 5 or 9 weeks after the cryptorchidism. Eight weeks after orchiopexy, the testes weights of ORC3 + 8, ORC5 + 8 and ORC9 + 8 animals were not reduced as much as the cryptorchid testes, being 20%, 25% and 40% lower than the respective control groups. Therefore, orchiopexy was effective in allowing the testes to almost reach the anticipated weights for the age. It was clearly observable that a shorter duration of cryptorchidism and a longer period of recovery allowed a better recovery of testes weights, as indicated by the data from ORC3 + 8 group.
FIGURE 2.
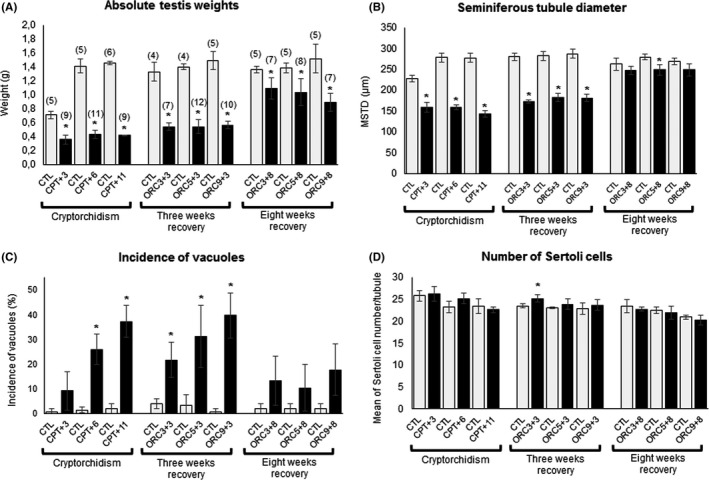
Groups submitted to cryptorchidism (CPT) only or to recovery of cryptorchidism alterations through orchiopexy (ORC). A, Absolute testes weights (g). B, Seminiferous tubule diameter. C, Incidence of vacuoles within the seminiferous epithelium. D, Number of Sertoli cells per tubule. Cryptorchid groups euthanized 3, 6 or 11 wks after the surgery: CPT + 3, CTP + 6 and CPT + 11, respectively. Orchiopexy at the 3rd (ORC3), 5th (ORC5) or 9th (ORC9) week after the cryptorchidism procedure. These animals were euthanized either 3 (groups ORC3 + 3, ORC5 + 3 and ORC9 + 3) or 8 (groups ORC3 + 8, ORC5 + 8 and ORC9 + 8) weeks after orchiopexy. CTL: Control group. Comparisons were performed using Student's t test, (*P < .05). Values are presented as mean ± SD. Numbers in parentheses: number of animals per group
The epididymides mean weights went through a similar process of decreased weights and recovery (data not shown).
3.2. Histology of the seminiferous tubules
Irrespective of the animal age at each time point during the study, almost all control (CTL) animals presented seminiferous tubules of about the same size (Figure 2B), with complete spermatogenesis (Score 1) (Table 1; Figure 3A,B). At the different time points, control animals showed a general picture of normal spermatogenesis with only rare seminiferous tubules with vacuoles (usually one vacuole/tubule) (Figure 2C) or germ cells going through apoptosis (not shown). In contrast, rats subjected only to cryptorchidism (CPT group) presented severe impairment of spermatogenesis (scores 3.3, 3.1 and 3.2 for CPT + 3, CPT + 6 and CPT + 11, respectively), with tubules containing spermatogonia and spermatocytes, intra‐epithelial vacuoles, apoptotic cells and giant multinucleated cells (Figure 3A,B) irrespective of the length of the different periods of cryptorchidism (Table 1). It should be noted that these observations did not include any immunohistochemistry staining, which could limit our analysis quantitatively but still provides an adequate qualitative and semi‐quantitative assessment. Normal seminiferous tubules were not observed in cryptorchid animals at any of the time points examined (3, 6 or 11 weeks after surgery), resulting in a score significantly higher (P < .003) than the control. The CPT groups also presented some SCO‐pattern tubules, sometimes atrophic, and the Sertoli cells appeared immature, with round or elongated nuclei (Figure 3C,D) compared to the control (Figure 3B).
Table 1.
Testicular histological scores of rats submitted to cryptorchidism (CPT) and orchiopexy (ORC) surgeries
| Groups a | n b | Histological score c | |
|---|---|---|---|
| Surgery d | Control e | ||
| Cryptorchidism + euthanasia after 3, 6 or 8 wks | |||
| CPT + 3 | 9/5 | 3.3 [3.1‐3.4]* | 1.0 [1.0‐1.0] |
| CPT + 6 | 10/5 | 3.1 [3.1‐3.3]* | 1.0 [1.0‐1.0] |
| CPT + 11 | 9/6 | 3.2 [3.3‐3.5]* | 1.0 [1.0‐1.0] |
| Orchiopexy at the 3rd, 5th and 9th weeks; euthanasia 3 wks later | |||
| ORC3 + 3 | 6/4 | 2.7 [2.5‐3.1]* | 1.0 [1.0‐1.0] |
| ORC5 + 3 | 12/4 | 2.5 [2.8‐2.9]* | 1.0 [1.0‐1.0] |
| ORC9 + 3 | 9/5 | 1.3 [1.1‐1.7]* | 1.0 [1.0‐1.1] |
| Orchiopexy at the 3rd, 5th and 9th weeks; euthanasia 8 wks later | |||
| ORC3 + 8 | 6/5 | 1.1 [1.1‐1.6] | 1.1 [1.0‐1.1] |
| ORC5 + 8 | 7/4 | 2.7 [2.5‐2.9]* | 1.0 [1.0‐1.0] |
| ORC9 + 8 | 6/5 | 1.3 [1.0‐1.7]* | 1.0 [1.0‐1.0] |
Animals submitted to cryptorchidism (CPT) or to CPT/orchiopexy (ORC).
n = number of animals underwent surgery/sham.
Values are expressed as the median (Q1‐Q3). Statistical analysis performed by generalized linear mixed‐effects model with gamma distribution followed by Sidak correction.
Animals submitted to cryptorchidism (CPT) or to CPT/orchiopexy (ORC).
Animals were sham‐operated at same time point as animals submitted to cryptorchidism or orchiopexy.
Significantly different from the respective control, P < .05.
FIGURE 3.
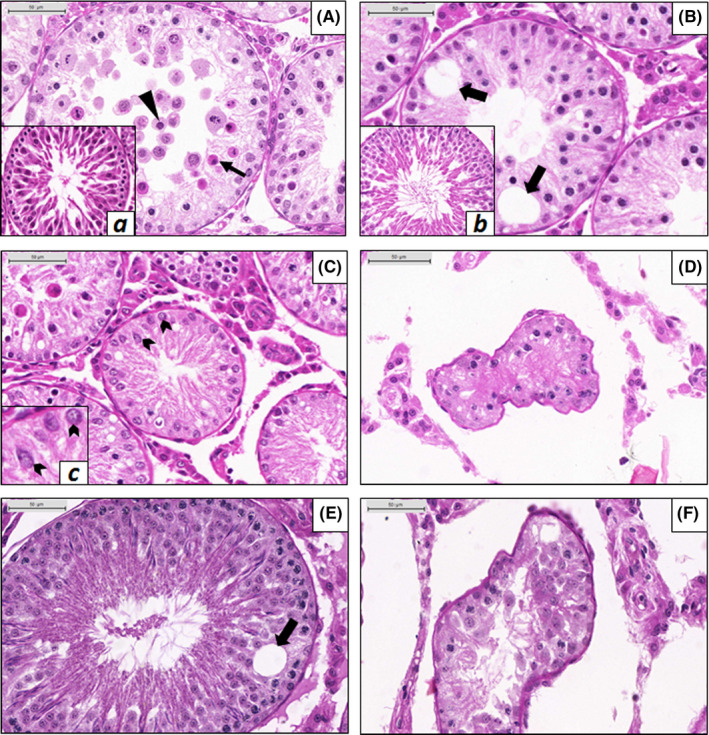
Histology of the testes from animals submitted to cryptorchidism (CPT) or to cryptorchidism followed by orchiopexy (ORC); testes from control animals are shown in the insets a, b and c. A, Seminiferous epithelium of a CPT + 3 animal. Several apoptotic germ cells are present (black narrow arrow), some located in the centre of the tubule, detached from the epithelium, characterizing exfoliation (black arrowhead). a, Seminiferous tubule in a control animal at the 6th week. B, Seminiferous epithelium of a CPT + 6 animal showing vacuoles (black wide arrows). b, Seminiferous tubule of a CPT + 6 control animal. C, Sertoli cell–only (SCO) tubule of a CPT + 3 animal. Sertoli cells show round nuclei; some of them appear to be fixed to the epithelium base, indicating possible dysfunction in the cell (black chevron arrows). c, SCO tubule showing immature Sertoli cells (black chevron arrows). D, Atrophic tubule in a CPT + 11 animal showing deformation of its normal conformation and severe impaired spermatogenesis (Sertoli cell–only pattern) tubule. E, Seminiferous tubule of an ORC3 + 8 animal. Spermatogenesis is complete, although vacuoles may be found in some tubules (black wide arrow). F, Atrophic seminiferous tubule of an ORC9 + 8 animal
Regarding the animals that underwent orchiopexy (ORC) after 3 weeks, a slight recovery of seminiferous tubules was observed, resulting in a score around 2.5 for ORC3 + 3 and ORC5 + 3 and a score of 1.3 for ORC9 + 3. Nevertheless, these scores were significantly higher than the respective controls (P ≤ .05) (Table 1). In general, when we analysed the 8‐week recovery groups (ORC3 + 8, ORC5 + 8, ORC9 + 8), more normal tubules were observed compared to the 3‐week recovery groups (ORC3 + 3, ORC5 + 3, ORC9 + 3). However, compared to the control groups, ORC5 + 8 and ORC9 + 8 groups showed significantly (P < .03) higher scores (scores 2.7 and 1.3, respectively). The ORC3 + 8 was the only group that showed a score of 1.1, similar to control (score 1.1) (Table 1, Figure 3E), highlighting that orchiopexy was effective in restoring the normal seminiferous epithelium when performed before puberty (or 3 weeks after the surgery for cryptorchidism). The interval of 3 or 8 weeks influenced the recovery only in ORC3 groups, suggesting that a long interval of recovery is successful only if the germ cells of the cryptorchid testis had not been damaged and lost.
Most of the rats submitted to orchiopexy after puberty (9 weeks after cryptorchidism) and euthanized 8 weeks after surgery (ORC9 + 8) presented atrophic tubules (Figure 3F), a condition not seen in other groups that had the testes relocated earlier to the scrotum.
3.3. Seminiferous tubule diameter, incidence of vacuoles, Sertoli cell number and expression of CK‐18
All groups that underwent cryptorchidism (CPT + 3, CPT + 6 and CPT + 11) and the 3‐week recovery groups (ORC3 + 3, ORC5 + 3 and ORC9 + 3) presented a significantly decreased (P < .05) mean seminiferous tubule diameter (MSTD) compared to the respective controls (Figure 2B). In the ORC3 + 8 and ORC9 + 8 groups, euthanized 8 weeks after orchiopexy, the MSTD estimates did not differ from their respective controls. A small but significant difference was still observed in the ORC5 + 8 group. ‘By ANOVA, the three 8‐week recovery groups (ORC3 + 8, ORC5 + 8, ORC9 + 8) showed no significant differences among the groups, indicating that they had similar MSTD re‐establishment.
The incidences of intraepithelial vacuoles increased progressively from the 3rd to the 11th week after establishment of cryptorchidism but were significantly higher (P < .05) than the respective controls only after 6 and 11 weeks (CPT + 6, CPT + 11) (Figure 2C). Orchiopexy had no influence on the progressive increase of vacuoles when evaluated only 3 weeks after surgery (ORC3 + 3, ORC5 + 3 and ORC9 + 3), and irrespective of the duration of cryptorchidism, but it was associated with reduced levels of vacuoles in the epithelium when the period of observation was extended to 8 weeks (ORC3 + 8, ORC5 + 8 and ORC9 + 8) (Figure 2C).
The number of Sertoli cells within the seminiferous tubules was similar among the groups, irrespective of the treatment the animals received (Figure 2D), although a slight but significant (P < .05) increase in the mean number of Sertoli cells was observed in the ORC3 + 3 group. Since ANOVA did not indicate significant differences among the three 3‐week recovery groups, that observation was assumed to be due to biological variation between the animals and not a response to cryptorchidism. The lack of immunohistochemical markers for Sertoli cells limits the precision of these determinations, but the overall lack of difference between groups suggests that there is little influence on Sertoli cells in this model. None of the CPT or ORC operated animals had any CK18‐positive immunostained cells within the seminiferous tubules (Figure 4).
FIGURE 4.
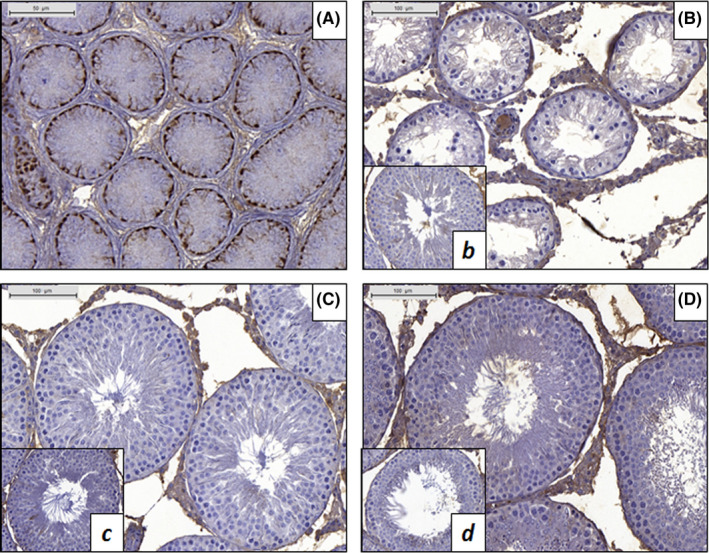
Immunohistochemistry for cytokeratin 18 (CK18). A, Immature testis of a normal 5‐d‐old rat showing positive CK18 staining in the cytoplasm of Sertoli cells. B, Seminiferous tubules of a CPT + 11 animal. No staining was observed in Sertoli cells. b, Control CPT + 11 animal. C, Testis of an ORC3 + 8 animal showing no CK18‐positive Sertoli cells. c, Control ORC3 + 3 animal. D, Seminiferous tubules of an ORC9 + 8 animal showing no staining for CK18. d, Control ORC9 + 8 animal
3.4. Mitotic and apoptotic indices
MIs in the control groups varied between 15%‐30%, but in the cryptorchid groups (CPT + 3, CPT + 5, CPT + 11), they were significantly (P < .05) decreased to <5% (Figure 5A,E). Three weeks after orchiopexy, the MI of groups ORC3 + 3, OEC5 + 3 and ORC9 + 3 was higher than that observed in the cryptorchid animals (CPT + 3, CPT + 5, CPT + 11), but still lower although not significantly compared to their respective controls. Eight weeks after orchiopexy, the MI in the ORC3 + 8, OEC5 + 8 and ORC9 + 8 orchiopexy groups was similar to that observed in the control groups (Figure 5C,E). Although a more precise and direct determination of the MIs for each cell type would be achieved by performing double‐labelling immunohistochemistry, our evaluation of H&E sections indicates that the germinative epithelium proliferative process is continuously depressed during cryptorchidism and that the relocation of the testes into the scrotum reverses this depression.
FIGURE 5.
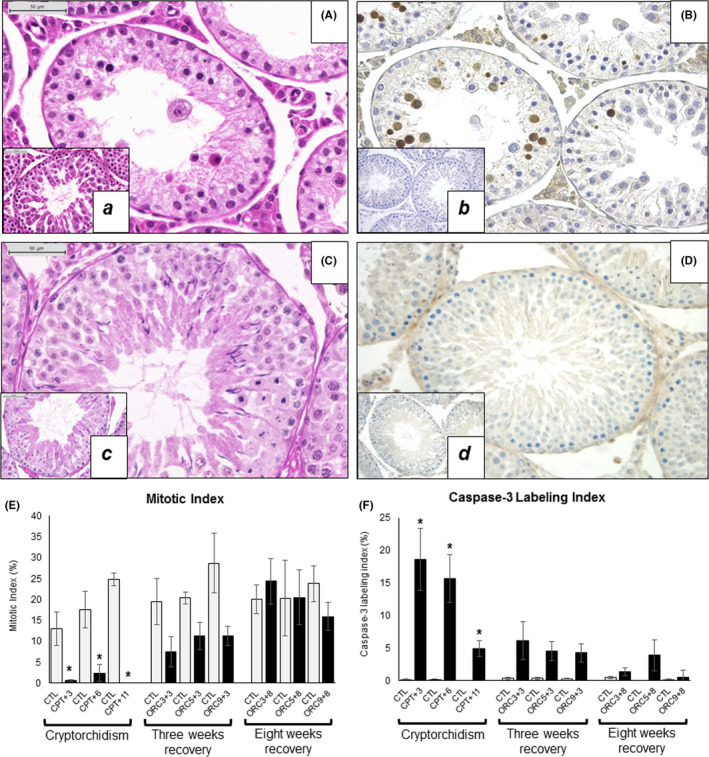
Mitosis and caspase‐3 expression in the rat testis. CPT + 3 animal showing no mitosis (A) and high number of caspase‐3‐positive cells (B), 400×. ORC9 + 8 animal with cells undergoing division (C) and no caspase‐3 expression (D), 400×. a‐d, Inset for each figure shows its respective control. E, Mitotic Index (%). F, Caspase‐3 Labeling Index (%). Comparisons were performed using Wilcoxon rank‐sum test (*P < .05)
As observed by caspase‐3 immunohistochemical expression, normally apoptosis occurs at very low levels in the germinative epithelium, <1% of the cells (Figure 5F). After the animals were made cryptorchid, this process increased significantly, particularly after 3 and 6 weeks (CPT + 3, CPT + 6), when the caspase‐3 LI reached values between 15% and 18% (Figure 5B,F). After this, the caspase‐3 LI decreased, as observed in the group that was euthanized 11 weeks after cryptorchidism (around 5% of the cells), although still significantly higher than the respective control. Orchiopexy induced a progressive reversal of the apoptotic activity in such a way that at the 8th week after the relocation of the testes into the scrotum (ORC9 + 8 group), the caspase‐3 LI practically returned to normal levels (Figure 5D,F).
4. DISCUSSION
Experimental models of cryptorchidism have been used mostly to evaluate testicular changes potentially associated with infertility. 11 , 12 These models reproduce severe spermatogenesis failure involving all structural components, with restoration of testis structure and function by performing orchiopexy. However, a comprehensive study, as presented here, that systematically characterizes the testicular changes related to surgical cryptorchidism after different periods and their reversion by orchiopexies performed at different rat ages is not available in the literature. The present study provides original information on the rat testes susceptibility to this experimental gonadal dystopia and the potential reversibility of the changes induced by cryptorchidism.
In the current study, surgical cryptorchidism was established in the immature rat. The induced testicular changes were similar to those described in other experimental rat models of cryptorchidism: decreased testicular weights, decreased tubular diameters, germ cell loss and spermatogenesis disruption. 23 , 24 , 26 , 29 , 41 In addition to showing that our procedure for orchiopexy performed at different times after the cryptorchidism procedure induced germ cell replenishment and spermatogenesis recovery, as observed in other studies, 23 , 26 , 30 we were able to document that reversibility of the changes was an age‐ and time‐dependent process, ie, the earlier the orchiopexy and the longer the recovery time, the more restored the testes were.
Apoptosis is a common process during normal spermatogenesis in mammals, which supports cellular homeostasis and a fine balance between maturing germinative cells and Sertoli cells. However, the apoptotic process of testicular germ cells can be enhanced by a variety of physical and chemical stresses, such as trauma, heat, radiation or withdrawal of hormonal support. 42 , 43 The significant decrease of testes weights and tubular diameters observed in the cryptorchid groups appears to be mostly related to the extensive germ cell loss, in parallel to decreased production of testicular fluid by the Sertoli cells. 29 As the duration of cryptorchidism was extended, there was progressive loss of germ cells and increased incidence of SCO tubules (Class 4). The number of germ cells expressing caspase‐3 gradually declined as the interval between cryptorchidism and termination increased. Exposure of the rat testis to mild heat for 6 hours resulted in activation of apoptosis by redistribution of the pro‐apoptotic regulator Bax protein from a cytoplasmic to a paranuclear localization in the heat‐susceptible germ cells. 44 The relocation of Bax is associated with activation of the initiator caspase‐9 and the executioner caspases 3, 6 and 7. 45 Therefore, the present findings suggest early activation of caspase‐3 and gradual germ cell disappearance due to apoptosis leading to extensive germinative epithelial loss by the 11th week after cryptorchidism. Although cell death appeared to be predominantly due to apoptosis, there is a possibility that some cell death also occurred due to necrosis. Nevertheless, there clearly was increased cell death following cryptorchidism and regeneration with orchiopexy.
In contrast to the apoptotic indices, the MIs were reduced in all three cryptorchid groups, but this was more accentuated in the CPT + 11 animals probably due to increased cell death and lack of germ cells. The progressive return of the MI to the levels of the controls as the interval after orchiopexy increased, in parallel with progressively less epithelial alterations, demonstrates the potential for recovery of the germinative epithelium.
The significant increase of vacuoles within the germinative epithelium in the cryptorchid groups suggests that the Sertoli cells were also compromised 46 , 47 by the gonadal dystopic condition. However, involvement of Sertoli cells apparently did not modify their differentiation status or their elimination, since in the cryptorchid groups there was no immuno‐expression of CK18 in Sertoli cells or changes in their number. In fact, CK18 antigen was not immunohistochemically expressed in Sertoli cells in groups that underwent cryptorchidism only or in those that also underwent orchiopexy, although it was positive in a 5‐day‐old control rat (Figure 5). Similar observations were made in human cryptorchid testes, which presented weak or absent CK18 immuno‐expression in SCO tubules. 10 , 48 These observations suggest that despite the morphological involvement (vacuoles), there was no compromise of Sertoli cell differentiation based on CK‐18 expression. However, besides CK18, other immunohistologic markers of cell immaturity, such as anti‐Mullerian hormone (AMH) and sialoglycoprotein (M2A antigen), have been used to verify the differentiation of Sertoli cells, not only in humans but also in experimental animals. 10 , 33 , 49 The expression of CK18 by Sertoli cells in human cryptorchid testes was negative, although expression was positive for AMH and M2A antigens. 10 Other studies have demonstrated that CK18 immuno‐expression and AMH immuno‐expression are both positive in human testis with spermatogenic arrest or Sertoli cell–only (SCO) tubules. 50 , 51 In addition, Rhesus monkeys made cryptorchid by interfering with the gubernaculum testis showed CK18‐positive Sertoli cells, suggesting that the differentiation of these cells was blocked. 33 Overall, in the present study, Sertoli cells were impaired by the altered micro‐environment, and the use of biomarkers other than CK18 for immature cells might provide a better assessment of their maturation status.
In humans, it has been suggested that early orchiopexy (between 6‐12 months of age) is preferable for reversing or preventing abnormal germ cell development, avoiding later infertility and increased risk of testicular cancer. 52 , 53 Experimentally, cryptorchid rats should also undergo orchiopexy as early as possible to avoid degeneration and enhance recovery of the seminiferous epithelium. 26 , 29 , 33 However, most of the reported experimental studies did not assess the interval for complete reversibility of testicular damage caused by cryptorchidism. In the present study, orchiopexy was performed at pre‐ and postpubertal ages and two intervals of testicular recovery from cryptorchidism—three and 8 weeks—were evaluated. Animals with a 3‐week recovery period (ORC3 + 3, ORC5 + 3 and ORC9 + 3) showed partial recovery of the germinative epithelium with the presence of spermatids and sperm (Class 2), along with some cells expressing caspase‐3 and partial recovery of the proliferative rate (MI) and almost complete recovery of testes weights and tubular diameters. The regeneration process naturally starts with germ cell proliferation (proliferative rate returning to control levels) originating from persisting spermatogonial stem cells, leading to cellular replenishment. However, the 3‐week interval was not sufficient to induce a complete recovery of the seminiferous tubules and spermatogenesis, or even a complete recovery of testes weights and tubular diameters. The incidence of vacuoles was still increased in all 3‐week recovery groups. This increase was time‐dependent and was more prominent in the group with the longest period between cryptorchidism and its reversion (ORC9 + 3), with a proportionally smaller increase in the other two groups (OCR3 + 3, ORC5 + 3). This observation suggests that the sooner the orchiopexy was performed, the lower were the number of vacuoles in the seminiferous epithelium, and possibly the Sertoli cell damage was less. The mean number of Sertoli cells per tubule in two of the three 3‐week recovery groups was not different from the control animals, even with increased incidence of vacuoles, suggesting that the population of Sertoli cells was not quantitatively involved in the process. The slight but statistically significant increase in the mean number of Sertoli cells in the ORC3 + 3 group was most likely due to biologic variation among the animals, rather than indicative of a response to cryptorchidism.
At the 8th week after orchiopexy, both the testes weights and the tubular diameters of the three recovery groups (ORC3 + 8, ORC5 + 8 and ORC9 + 8) were similar to the respective controls. Also, all groups had caspase‐3 expression and MI levels similar to the control, indicating complete recovery of the seminiferous tubules. However, only the animals of the ORC3 + 8 group that underwent orchiopexy before puberty showed morphologic normal spermatogenesis, despite the presence of some vacuoles and apoptotic bodies. Considering the testicular histologic picture, the most optimum time point to perform orchiopexy was the 3rd week after cryptorchidism since no morphologic differences were observed when compared to the control group (Table 1, Figure 3E). Although we have not experimentally addressed animal fertility in the current study, it would be informative to evaluate whether the ORC animals reached a sexually mature age in subsequent work, but that is beyond the scope of this study.
In the present study, experimental cryptorchidism led to decreased testes and epididymis weights, decreased tubular diameters, spermatogenesis disruption with germ cell loss and increased incidence of vacuoles in the seminiferous epithelium, and tubular atrophy. The intensity of testicular damage was proportional to the duration of the abdominal cryptorchidism. Spermatogenesis was completely recovered only when testicular relocation occurred before puberty (at the 6th week of age, 3 weeks after the surgery for cryptorchidism) and the interval for recovery was long (8 weeks), indicating that age and sexual maturity were relevant for restoration of morphologically normal spermatogenesis. The systematic description of the alterations induced by cryptorchidism and the degree of its reversion by orchiopexy performed at different times as presented here provides a useful model for studying germinative epithelial impairment and recovery in different experimental settings, including exposure to exogenous chemicals. However it is of course important to take into account that, like in all models, there are limitations to this model for possible extrapolation to humans, especially regarding differences in ages and maturity for germ cell development and descent of the testis into the scrotum.
ETHICAL APPROVAL
This study was approved by the Committee for Ethics in Animal Experimentation of the UNESP Medical School, SP, Brazil, protocol no. 926/2012.
CONFLICT OF INTEREST
There are no potential conflicts of interest. During the development of this study, authors APFC, LMMG, NPS and MGNP were enrolled in the Graduate Program of Pathology at the UNESP Medical School, Botucatu, SP, Brazil.
ACKNOWLEDGEMENTS
Paulo Roberto Cardoso and Paulo Cesar Georgete provided helpful technical assistance. The authors are grateful to Lynette M. Smith, University of Nebraska Medical Center, Omaha, NE, USA, for assistance with statistical analyses of the mitotic and caspase‐3 labelling data.
Ferragut Cardoso AP, Gomide LMM, Souza NP, et al. Time response of rat testicular alterations induced by cryptorchidism and orchiopexy. Int J Exp Path.2021;102:57–69. 10.1111/iep.12384
Ana P. Ferragut Cardoso and Lígia M. M. Gomide contributed equally to this publication and serve as co‐first authors.
Funding information
This work was supported by the São Paulo Research Foundation—FAPESP (grant no. 2012/09873‐4), the Brazilian Federal Agency for Support and Evaluation of Graduate Education—CAPES (fellowship 99999.001280/2014‐07) and the Center for Evaluation of Environmental Impact of Human Health (TOXICAM), Botucatu Medical School, São Paulo State Univ (UNESP), Botucatu, SP, Brazil.
REFERENCES
- 1. Toppari J, Virtanen HE, Main KM, Skakkebaek NE. Cryptorchidism and hypospadias as a sign of testicular dysgenesis syndrome (TDS): environmental connection: cryptorchidism and hypospadias. Birth Defects Res A Clin Mol Teratol. 2010;88:910‐919. 10.1002/bdra.20707 [DOI] [PubMed] [Google Scholar]
- 2. Hutson JM, Balic A, Nation T, Southwell B. Cryptorchidism. Semin Pediatr Surg. 2010;19:215‐224. 10.1053/j.sempedsurg.2010.04.001 [DOI] [PubMed] [Google Scholar]
- 3. Ferguson L, Agoulnik AI. Testicular cancer and cryptorchidism. Front Endocrinol. 2013;4:32 10.3389/fendo.2013.00032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Skakkebaek NE, Rajpert‐De Meyts E, Main KM. Testicular dysgenesis syndrome: an increasingly common developmental disorder with environmental aspects. Hum Reprod. 2001;16:972‐978. [DOI] [PubMed] [Google Scholar]
- 5. Sharpe RM, Skakkebaek NE. Testicular dysgenesis syndrome: mechanistic insights and potential new downstream effects. Fertil Steril. 2008;89:e33‐e38. 10.1016/j.fertnstert.2007.12.026 [DOI] [PubMed] [Google Scholar]
- 6. Durairajanayagam D, Agarwal A, Ong C. Causes, effects and molecular mechanisms of testicular heat stress. Reprod Biomed Online. 2015;30:14‐27. 10.1016/j.rbmo.2014.09.018 [DOI] [PubMed] [Google Scholar]
- 7. AbouZeid AA, Mousa MH, Soliman HA, Hamza AF, Hay SA. Intra‐abdominal testis: histological alterations and significance of biopsy. J Urology. 2011;185:269‐274. 10.1016/j.juro.2010.09.026 [DOI] [PubMed] [Google Scholar]
- 8. Koni A, Ozseker HS, Arpali E, et al. Histopathological evaluation of orchiectomy specimens in 51 late postpubertal men with unilateral cryptorchidism. J Urology. 2014;192:1183‐1188. 10.1016/j.juro.2014.05.048 [DOI] [PubMed] [Google Scholar]
- 9. Kollin C, Stukenborg JB, Nurmio M, et al. Boys with undescended testes: endocrine, volumetric and morphometric studies on testicular function before and after orchidopexy at nine months or three years of age. J Clin Endocrinol Metab. 2012;97:4588‐4595. 10.1210/jc.2012-2325 [DOI] [PubMed] [Google Scholar]
- 10. Nistal M, Gonzalez‐Peramato P, De Miguel MP. Sertoli cell dedifferentiation in human cryptorchidism and gender reassignment shows similarities between fetal environmental and adult medical treatment estrogen and antiandrogen exposure. Reprod Toxicol. 2013;42:172‐179. 10.1016/j.reprotox.2013.08.009 [DOI] [PubMed] [Google Scholar]
- 11. Watts LM, Hasthorpe S, Farmer PJ, Hutson JM. Apoptotic cell death and fertility in three unilateral cryptorchid rat models. Urol Res. 2000;28:332‐337. [DOI] [PubMed] [Google Scholar]
- 12. Bergh A, Söder O. Studies of cryptorchidism in experimental animal models. Acta Paediatr. 2007;96:617‐621. 10.1111/j.1651-2227.2007.00295.x [DOI] [PubMed] [Google Scholar]
- 13. Goh DW, Momose Y, Middlesworth W, Hutson JM. The relationship among calcitonin gene‐related peptide, androgens and gubernacular development in 3 animal models of cryptorchidism. J Urology. 1993;150:574‐576. 10.1016/S0022-5347(17)35554-4 [DOI] [PubMed] [Google Scholar]
- 14. van der Schoot P. Disturbed testicular descent in the rat after prenatal exposure to the antiandrogen flutamide. J Reprod Fertil. 1992;96:483‐496. [DOI] [PubMed] [Google Scholar]
- 15. Mahood IK, Hallmark N, McKinnell C, Walker M, Fisher JS, Sharpe RM. Abnormal leydig cell aggregation in the fetal testis of rats exposed to Di ( n ‐Butyl) phthalate and its possible role in testicular dysgenesis. Endocrinology. 2005;146:613‐623. 10.1210/en.2004-0671 [DOI] [PubMed] [Google Scholar]
- 16. Lein M, Fahlenkamp D, Schönberger B, Prollius S, Loening S. The pharmacological effect of the gonadotrophin‐releasing hormone on experimental cryptorchidism in rats. Scand J Urol Nephrol. 1996;30:185‐191. [DOI] [PubMed] [Google Scholar]
- 17. Souza NP, Arnold LL, Pennington KL, et al. Isolation and molecular characterization of spermatogonia from male Sprague‐Dawley rats exposed in utero and postnatally to dibutyl phthalate or acrylamide. Toxicol Mech Metho. 2019;29:488‐498. 10.1080/15376516.2019.1611981 [DOI] [PubMed] [Google Scholar]
- 18. Dündar M, Koçak I, Culhaci N. A new experimental model for cryptorchidism: inguinoscrotal approach. Urol Res. 2001;29:178‐181. [DOI] [PubMed] [Google Scholar]
- 19. Huhtaniemi I, Poutanen M. Transgenic and knockout mouse models for aberrant pituitary‐testicular function: relevance to the pathogenesis of cryptorchidism. Turk J Pediatr. 2004;46(Suppl):28‐34. [PubMed] [Google Scholar]
- 20. Mouhadjer N, Pointis G, Malassine A, Bedin M. Testicular steroid sulfatase in a cryptorchid rat strain. J Steroid Biochem. 1989;34:555‐558. [DOI] [PubMed] [Google Scholar]
- 21. Lugg JA, Penson DF, Sadeghi F, et al. Prevention of seminiferous tubular atrophy in a naturally cryptorchid rat model by early surgical intervention. J Androl. 1996;17:726‐732. [PubMed] [Google Scholar]
- 22. Park WH, Hutson JM. The gubernaculum shows rhythmic contractility and active movement during testicular descent. J Pediatr Surg. 1991;26:615‐617. [DOI] [PubMed] [Google Scholar]
- 23. Jegou B, Peake RA, Irby DC, de Kretser DM. Effects of the induction of experimental cryptorchidism and subsequent orchidopexy on testicular function in immature rats. Biol Reprod. 1984;30:179‐187. [DOI] [PubMed] [Google Scholar]
- 24. Rossi LM, Pereira LAV, de Santis L, et al. Sperm retrieval techniques in rats with suppressed spermatogenesis by experimental cryptorchidism. Hum Reprod. 2005;20:443‐447. 10.1093/humrep/deh549 [DOI] [PubMed] [Google Scholar]
- 25. Garcia PV, Arrotéia KF, Joazeiro PP, de Fátima Paccola Mesquita S, de Grava Kempinas W, Pereira LAV. Orchidopexy restores morphometric‐stereologic changes in the caput epididymis and daily sperm production in cryptorchidic mice, although sperm transit time and fertility parameters remain impaired. Fertil Steril. 2011;96:739‐744. 10.1016/j.fertnstert.2011.06.036 [DOI] [PubMed] [Google Scholar]
- 26. Souza N, Ferragut Cardoso AP, Gomide LM, et al. Experimental cryptorchidism enhances testicular susceptibility to dibutyl phthalate or acrylamide in Sprague‐ Dawley rats. Human Exper Toxicol. 2019;38(8):899‐913. [DOI] [PubMed] [Google Scholar]
- 27. Loebenstein M, Hutson J, Li R. Gonocyte transformation in a congenitally cryptorchid rat is normal and may be similar to the situation reported in human acquired cryptorchidism. J Pediatr Surg. 2018;53:1770‐1775. 10.1016/j.jpedsurg.2017.12.026 [DOI] [PubMed] [Google Scholar]
- 28. Sengupta P. The laboratory rat: relating its age with human’s. Int J Prev Med. 2013;4:624‐630. [PMC free article] [PubMed] [Google Scholar]
- 29. Jégou B, Le Gac F, Irby DC, de Kretser DM. Studies on seminiferous tubule fluid production in the adult rat: effect of hypophysectomy and treatment with FSH, LH and testosterone. Int J Androl. 1983;6:249‐260. [DOI] [PubMed] [Google Scholar]
- 30. Mizuno K, Hayashi Y, Kojima Y, Kurokawa S, Sasaki S, Kohri K. Early orchiopexy improves subsequent testicular development and spermatogenesis in the experimental cryptorchid rat model. J Urology. 2008;179:1195‐1199. 10.1016/j.juro.2007.10.029 [DOI] [PubMed] [Google Scholar]
- 31. Liu F, Huang H, Xu Z‐L, Qian X‐J, Qiu W‐Y. Germ cell removal after induction of cryptorchidism in adult rats. Tissue Cell. 2012;44:281‐287. 10.1016/j.tice.2012.04.005 [DOI] [PubMed] [Google Scholar]
- 32. Lanning LL, Creasy DM, Chapin RE, et al. Recommended approaches for the evaluation of testicular and epididymal toxicity. Toxicol Pathol. 2002;30:507‐520. 10.1080/01926230290105695 [DOI] [PubMed] [Google Scholar]
- 33. Zhang Z‐H, Hu Z‐Y, Song X‐X, et al. Disrupted expression of intermediate filaments in the testis of rhesus monkey after experimental cryptorchidism. Int J Androl. 2004;27:234‐239. 10.1111/j.1365-2605.2004.00477.x [DOI] [PubMed] [Google Scholar]
- 34. Quinn FM, Crockard AD, Brown S. Reversal of degenerative changes in the scrotal testis after orchidopexy in experimental unilateral cryptorchidism. J Pediatr Surg. 1991;26:451‐454. [DOI] [PubMed] [Google Scholar]
- 35. Latendresse JR, Warbrittion AR, Jonassen H, Creasy DM. Fixation of testes and eyes using a modified Davidson’s fluid: comparison with Bouin’s fluid and conventional Davidson’s fluid. Toxicol Pathol. 2002;30:524‐533. 10.1080/01926230290105721 [DOI] [PubMed] [Google Scholar]
- 36. Kittel B, Ruehl‐Fehlert C, Morawietz G, et al. Revised guides for organ sampling and trimming in rats and mice–Part 2. A joint publication of the RITA and NACAD groups. Exp Toxicol Pathol. 2004;55:413‐431. [DOI] [PubMed] [Google Scholar]
- 37. Detre S, Saclani Jotti G, Dowsett M. A “quickscore” method for immunohistochemical semiquantitation: validation for oestrogen receptor in breast carcinomas. J Clin Pathol. 1995;48:876‐878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Ishibashi H, Suzuki T, Suzuki S, et al. Sex steroid hormone receptors in human thymoma. J Clin Endocrinol Metab. 2003;88:2309‐2317. 10.1210/jc.2002-021353 [DOI] [PubMed] [Google Scholar]
- 39. Tenorio BM, Jimenez GC, Morais RN, Torres SM, Albuquerque Nogueira R, Silva Junior VA. Testicular development evaluation in rats exposed to 60 Hz and 1 mT electromagnetic field. J Appl Toxicol. 2011;31:223‐230. 10.1002/jat.1584 [DOI] [PubMed] [Google Scholar]
- 40. Markelewicz RJ, Hall SJ, Boekelheide K. 2,5‐hexanedione and carbendazim coexposure synergistically disrupts rat spermatogenesis despite opposing molecular effects on microtubules. Toxicol Sci. 2004;80:92‐100. 10.1093/toxsci/kfh140 [DOI] [PubMed] [Google Scholar]
- 41. Zakaria O, Shono T, Imajima T, Suita S. Fertility and histological studies of the contralateral testes in two different intra‐ and extra‐abdominal rat models of unilateral cryptorchidism. Br J Urol. 1998;82:574‐577. [DOI] [PubMed] [Google Scholar]
- 42. Kierszenbaum AL. Apoptosis during spermatogenesis: the thrill of being alive. Mol Reprod Dev. 2001;58:1‐3. 10.1002/1098-2795(200101)58:1<1:AID-MRD1>3.0.CO;2-0 [DOI] [PubMed] [Google Scholar]
- 43. Gautam DK, Misro MM, Chaki SP, Chandra M, Sehgal N. hCG treatment raises H2O2 levels and induces germ cell apoptosis in rat testis. Apoptosis. 2007;12:1173‐1182. 10.1007/s10495-007-0060-1 [DOI] [PubMed] [Google Scholar]
- 44. Yamamoto CM, Sinha Hikim AP, Huynh PN, et al. Redistribution of Bax is an early step in an apoptotic pathway leading to germ cell death in rats, triggered by mild testicular hyperthermia. Biol Reprod. 2000;63:1683‐1690. [DOI] [PubMed] [Google Scholar]
- 45. Hikim APS, Lue Y, Yamamoto CM, et al. Key apoptotic pathways for heat‐induced programmed germ cell death in the testis. Endocrinology. 2003;144:3167‐3175. 10.1210/en.2003-0175 [DOI] [PubMed] [Google Scholar]
- 46. Kerr JB, Rich KA, de Kretser DM. Effects of experimental cryptorchidism on the ultrastructure and function of the Sertoli cell and peritubular tissue of the rat testis. Biol Reprod. 1979;21:823‐838. [DOI] [PubMed] [Google Scholar]
- 47. Kanter M, Aktas C, Erboga M. Heat stress decreases testicular germ cell proliferation and increases apoptosis in short term: an immunohistochemical and ultrastructural study. Toxicol Indust Health. 2013;29:99‐113. 10.1177/0748233711425082 [DOI] [PubMed] [Google Scholar]
- 48. Rogatsch H, Jezek D, Hittmair A, Mikuz G, Feichtinger H. Expression of vimentin, cytokeratin, and desmin in Sertoli cells of human fetal, cryptorchid, and tumour‐adjacent testicular tissue. Virchows Arch. 1996;427:497‐502. [DOI] [PubMed] [Google Scholar]
- 49. Hutchison GR, Scott HM, Walker M, et al. Sertoli cell development and function in an animal model of testicular dysgenesis syndrome. Biol Reprod. 2008;78:352‐360. 10.1095/biolreprod.107.064006 [DOI] [PubMed] [Google Scholar]
- 50. Steger K, Rey R, Kliesch S, Louis F, Schleicher G, Bergmann M. Immunohistochemical detection of immature Sertoli cell markers in testicular tissue of infertile adult men: a preliminary study. Int J Androl. 1996;19:122‐128. [DOI] [PubMed] [Google Scholar]
- 51. Steger K, Rey R, Louis F, et al. Reversion of the differentiated phenotype and maturation block in Sertoli cells in pathological human testis. Hum Reprod. 1999;14:136‐143. [DOI] [PubMed] [Google Scholar]
- 52. Chan E, Wayne C, Nasr A. Ideal timing of orchiopexy: a systematic review. Pediatr Surg Int. 2014;30:87‐97. 10.1007/s00383-013-3429-y [DOI] [PubMed] [Google Scholar]
- 53. Hutson JM, Vikraman J, Li R, Thorup J. Undescended testis: What paediatricians need to know: undescended testis. J Paediatr Child Health. 2017;53:1101‐1104. 10.1111/jpc.13744 [DOI] [PubMed] [Google Scholar]