

Abstract
As the organ with the highest demand for oxygen, the brain has a poor tolerance to ischemia and hypoxia. Despite severe ischemia/hypoxia induces the occurrence and development of various central nervous system (CNS) diseases, sublethal insult may induce strong protection against subsequent fatal injuries by improving tolerance. Searching for potential measures to improve brain ischemic/hypoxic is of great significance for treatment of ischemia/hypoxia related CNS diseases. Ischemic/hypoxic preconditioning (I/HPC) refers to the approach to give the body a short period of mild ischemic/hypoxic stimulus which can significantly improve the body's tolerance to subsequent more severe ischemia/hypoxia event. It has been extensively studied and been considered as an effective therapeutic strategy in CNS diseases. Its protective mechanisms involved multiple processes, such as activation of hypoxia signaling pathways, anti‐inflammation, antioxidant stress, and autophagy induction, etc. As a strategy to induce endogenous neuroprotection, I/HPC has attracted extensive attention and become one of the research frontiers and hotspots in the field of neurotherapy. In this review, we discuss the basic and clinical research progress of I/HPC on CNS diseases, and summarize its mechanisms. Furthermore, we highlight the limitations and challenges of their translation from basic research to clinical application.
Keywords: hypoxia, ischemia, neurological diseases, neuroprotection, preconditioning
IPC/HPC/RIPC exert neurprotective effects by several molecular mechanisms, including activating HIF‐1α signaling, reducing ROS, activating autophagy, anti‐apoptosis, anti‐inflammation, reducing excitotoxicity, and enhancing BBB integrity. BBB, blood brain barrier; HIF, hypoxia inducible factor; HPC, hypoxic preconditioning; IPC, ischemic preconditioning; RIPC, remote ischemic preconditioning; ROS, reactive oxygen species.

1. INTRODUCTION
Based on the idea that sublethal insult may induce strong protection against subsequent fatal injuries, the first hypoxic preconditioning (HPC) study took place in 1964, which confirmed HPC‐afforded tolerance of the brain against subsequent cerebral ischemic injury. 1 In 1990, ischemic preconditioning (IPC) was reported and proved to elicit protective effects on ischemic damage. 2 Compared with severe or pathogenic ischemic/hypoxic events, I/HPC reverses the pathological process through a milder and appropriate degree of stimulation. The brain is extremely sensitive to oxygen levels. I/HPC has been demonstrated to allow for resistance of various cerebral injuries, such as stroke, neonatal hypoxia/ischemia, and neurodegenerative diseases. 3 Interestingly, repeated transient limb ischemia, termed “remote ischemic preconditioning (RIPC),” can also alleviate the ischemic injury of a distant organ, such as brain. RIPC has been widely studied in clinical trials in recent years and seems to be of more clinical value, because it avoids the direct ischemic/hypoxic insults to important organs. 4 In this review, we focus on the potential therapeutic effects of I/HPC and RIPC in central nervous system (CNS) diseases, discuss the underlying protective mechanisms, and highlight the challenges of their translation from basic research to clinical application.
2. RESEARCH PROGRESS OF I/HPC IN CNS DISEASES
Central nervous system diseases are comprised of cerebrovascular diseases, neurodegenerative diseases, multiple sclerosis, spinal cord injury, and others. As a potential therapeutic strategy, the protective effects of I/HPC and RIPC in CNS diseases have been extensively studied in multiple layers including in vitro cell cultures, ex vivo brain slices, in vivo experimental animal models, and clinical patients (Tables 1 and 2, Figure 1). In this section, we will introduce the research progress of I/HPC and RIPC in both clinical and preclinical CNS diseases.
TABLE 1.
Basic research cases of neuroprotection of IPC/HPC/RIPC
| Method | Subjects | Hypoxia dosage | Outcome | References |
|---|---|---|---|---|
| IPC | tMCAO rats | 10 min of tMCAO, followed by 24 h of recovery and reperfusion |
Neurological outcomes ↑ Lesion volume ↓ Apoptosis ↓ |
132 |
| 30 min of tMCAO, followed by 72 h of recovery and reperfusion |
Neurological outcomes ↑ Lesion volume ↓ ER stress ↓ |
51, 117 | ||
| 5 cycles of 3 min transient occlusion of the bilateral common carotid arteries with each followed by 5 min of reperfusion |
Neurological outcomes ↑ Lesion volume ↓ |
133 | ||
| pMCAO rats | 10 min of tMCAO, followed by 24 h of recovery and reperfusion |
Neurological outcomes ↑ Lesion volume ↓ Brain edema ↓ Autophagy ↑ |
113, 114, 134 | |
| tMCAO mice | 5 min of tMCAO, followed by 24 h of recovery and reperfusion | Lesion volume ↓ | 135 | |
| 12 min of tMCAO, followed by 72 h of recovery and reperfusion |
Lesion volume ↓ BBB integrity ↑ Oxidative stress ↓ |
60, 61 | ||
| 15 min of tMCAO, followed by 72 h of recovery and reperfusion |
Lesion volume ↓ HIF−1α level ↑ |
53, 79 | ||
| pMCAO mice | 7 min of tMCAO, followed by 96 h of recovery and reperfusion |
Lesion volume ↓ BBB integrity ↑ |
124 | |
| ICH rats | 15 min of tMCAO, followed by 72 h of recovery and reperfusion |
Brain edema ↓ Blood coagulation ↓ |
15 | |
| forebrain ischemia gerbils | 5 min forebrain ischemia, followed by 72 h of recovery and reperfusion |
Neuronal apoptosis↓ Dendritic integrity ↑ |
136 | |
| HPC | tMCAO rats | altitude 5000 m for 3 h daily for 14 days |
Lesion volume↓ Cognitive function↑ Inflammation↓ |
77 |
| tMCAO mice | 8% or 11% O2 for 2 h or 4 h daily for 14 days |
Lesion volume↓ Inflammation↓ |
92, 93 | |
| tMCAO mice | 8% O2 for 4 h, followed by 48 or 72 h of recovery |
Lesion volume↓ Integrity of BBB ↑ |
121, 137 | |
| Propofol‐treated rat pups | 8% O2 for 10 min, followed by room air for a 10 min, five cycles | Apoptosis ↓ | 138, 139 | |
| H‐I injury piglet | 8% O2 for 3 h or 24 h |
Brain damage ↓ HIF−1α level ↑ VEGF ↑ |
46 | |
| tGCI rats | 8% O2 for 30 min, followed by 24 h of recovery |
Neurological outcomes ↑ Autophagy ↑ Apoptosis↓ Mitochondrial function↑ |
109, 140 | |
| EAE mice | 8% or 10% O2 for 14d |
Integrity of BBB ↑ Inflammation↓ |
26, 27, 29 | |
| RIPC | tMCAO rats | Both hind limbs 4 cycles of 5 min ischemia followed by 5 min of reperfusion |
Neurological outcomes ↑ Lesion volume ↓ Splenic immune response↑ |
9 |
| Left hind limb 4 cycles of 5 min ischemia followed by 5 min of reperfusion daily for 3 days |
Neurological outcomes ↑ Lesion volume ↓ Apoptosis ↓ |
84 | ||
| Both hind limbs 3 cycles of 10 min ischemia followed by 10 min of reperfusion |
Lesion volume ↓ Neurological outcomes ↑ Inflammation↓ HIF−1α and HIF−2α↓ |
80, 141 | ||
| tMCAO diabetic mice | Both hind limbs 3 cycles of 10 min ischemia followed by 10 min of reperfusion |
Lesion volume ↓ Neurological outcomes ↑ Inflammation↓ Apoptosis↓ |
94, 142 | |
| tGCI mice | left hind limb 4 cycles of 5 min ischemia followed by 5 min of reperfusion |
Lesion volume ↓ Neurological outcomes ↑ Vascular dementia↓ Apoptosis↓ Oxidative stress↓ |
62 |
TABLE 2.
Clinical study cases of neuroprotection of RIPC
| Method | Subjects | Hypoxia dosage | Outcome | References |
|---|---|---|---|---|
| RIPC | Carotid artery stenting patients | Bilateral upper limb 5 cycles consisting of 5 min ischemia and 5 min reperfusion, twice daily for 14 days | Secondary ischemic brain injury ↓ | 143 |
| Intracranial arterial stenosis patients | Bilateral upper limb 5 cycles consisting of 5 min ischemia and 5 min reperfusion, twice daily for 300 days |
Cerebral perfusion ↑ Incidence of recurrent stroke ↓ Fazekas and Scheltens scores ↓ |
144, 145, 146 | |
| Subarachnoid hemorrhage patients | The upper arm 3 cycles consisting of 5 min ischemia and 5 min reperfusion for 14 days | Safe and well tolerated | 13 | |
| Lower limb 4 cycles consisting of 5 min ischemia and reperfusion for 4 times |
Incidence of stroke ↓ Mortality ↓ |
14 | ||
| Acute ischemic stroke patients | The upper arm 5 cycles consisting of 3 min ischemia and 5 min reperfusion, twice daily for 5 days |
Lesion volume ↓ Functional recovery↑ |
147 | |
| Subcortical ischemic vascular dementia patients | Bilateral upper limb 5 cycles consisting of 5 min ischemia and 5 min reperfusion, twice daily for 180 days | Cognitive function↑ | 148 | |
| Ischemic moyamoya disease patients | Bilateral upper limb 5 cycles consisting of 5 min ischemia and 5 min reperfusion, three times daily for 720 days |
Ischemic events ↓ Cerebral perfusion ↑ |
149 | |
| Small vessel disease patients | Bilateral upper limb 5 cycles consisting of 5 min ischemia and 5 min reperfusion, twice daily for 360 days |
Mean flow velocity of the middle cerebral artery ↑ White matter lesion volume↓ |
150 | |
| Brain tumor patients | The upper arm 3 cycles consisting of 5 min ischemia and 5 min reperfusion |
Incidence of postoperative Ischemic Damage ↓ Lesion volume↓ |
151 | |
| Healthy young men and women | The upper arm 4 cycles consisting of 5 min ischemia and 5 min reperfusion |
Plasmic BDNF and VEGF↑ Microvascular endothelial function↑ |
152 |
FIGURE 1.
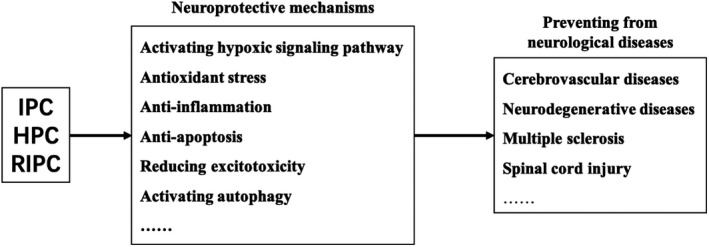
Neuroprotective mechanisms of IPC/HPC/RIPC treatment in neurological diseases. IPC/HPC/RIPC could prevent from several neurological diseases, such as cerebrovascular diseases, neurodegenerative diseases, multiple sclerosis, and spinal cord injury. There protective machenisms including activating hypoxic signaling pathway, antioxidant stress, anti‐inflammation, anti‐apoptosis, reducing excitotoxicity, and activating autophagy. HPC, hypoxic preconditioning; IPC, ischemic preconditioning; RIPC, remote ischemic preconditioning
2.1. Cerebrovascular diseases
Ischemic stroke is caused by cerebral vascular occlusion, accounting for 80% of stroke cases. Thrombolytic tissue plasminogen activator is the best strategy, but its narrow therapeutic window limits its clinical usage. 5 Since the 1990s, several data on I/HPC have been collected in animal models of focal and global cerebral ischemia, consistently proving that regional brief ischemic/hypoxic episode exerts subsequent neuroprotection against subsequent major ischemia/hypoxia event. 6 In rats, hypoxia exposure significantly decreased the infarct volume induced by focal permanent ischemia. 7 Clinically, transient ischemic attack (TIA) can be regarded as a kind of IPC in situ. Patients with TIA history before an ischemic stroke were observed to have better prognosis than those without TIA history, 8 confirming the protective effects of cerebral preconditioning. Similarly, RIPC with limb has also been found to protect against ischemic stroke in several clinical studies. 9 , 10 , 11
Hemorrhagic stroke accounts for about 20% of stroke, with very limited treatment options. Rupture of intracranial aneurysms is one of the most critical reasons for subarachnoid hemorrhage (SAH), it frequently resulted in subsequent vasospasm leading to delayed cerebral ischemia (DCI) and focal neurological deficits. HPC can reduce vasospasm and DCI after SAH. 12 RIPC was safe and well tolerated for patients with SAH, 13 and decreased the incidence of stroke and death. 14 IPC also protected against brain edema and blood hypo‐coagulation in intra‐cerebral hemorrhage (ICH) rats. 15
Stem cell transplantation therapy is a hot topic in the treatment of stroke. IPC improved the curative effect of stem cell transplantation in ischemic stroke model induced by transient middle cerebral artery occlusion (tMCAO). 16 HPC in neural stem cells and bone marrow mesenchymal stem cells (BMSCs) enhanced efficacy of stem cell therapy by promoting grafted‐cell survival in the ICH models. 17 , 18 Mechanistically, HPC‐treated BMSC significantly increased the expression of some key survival and regeneration factors, such as B‐cell lymphoma‐2 (Bcl‐2), brain‐derived neurotrophic factor (BDNF), and VEGF, to promote functional recovery. 18 Taken together, these studies indicate that I/HPC is a promising strategy for therapy or combination therapy of cerebrovascular diseases, while their exact mechanism remains to be explored.
2.2. Neurodegenerative diseases
Neurodegenerative diseases refer to progressive dysfunction and death of selective neuronal subsets. Alzheimer's disease (AD) and Parkinson's disease (PD) are the most common neurodegenerative diseases, hypoxia also participates in their development. I/HPC is a potential approach to prevent neurodegeneration. 19 In experimental AD animals, intermittent hypoxic training (IHT) could alleviate AD pathology and improve cognitive function by preventing neuronal loss. 20 , 21 This was associated with preserved cerebrovascular function through reduced oxidative stress. 20 The idea of I/HPC was tested clinically in elderly patients with mild cognitive impairment, a precursor of AD. IHT was proved to improve cognitive function and delay the development of AD. 22 Currently there is little evidence on the effects of I/HPC in PD, although other sorts of preconditioning such as cross‐hemispheric preconditioning seems to confer a favorable outcome in PD. 23 Since hypoxia is closely associated with various pathogenic mechanisms of PD, we believe it is worth investigating, and is a potential direction for PD management.
2.3. Multiple sclerosis
Multiple sclerosis (MS) is an autoimmune disease characterized by white matter inflammatory demyelination, it is common in young and middle‐aged people. 24 Experimental allergic encephalomyelitis (EAE) is a wildly used preclinical MS model, with similar immune pathogenesis and lesions to MS. 25 HPC can prevent the development of EAE by decreasing leukocyte infiltration to the CNS, 26 and microglia may play a critical role on its protective mechanisms. 27 Furthermore, increased levels of regulatory T cells (Tregs) and anti‐inflammatory cytokine interleukin (IL)‐10 may also involve in the neuroprotective effects of HPC on EAE models. 28 In addition to its anti‐inflammatory effects, HPC also promotes EAE recovery by promoting vascular remodeling response and enhancing blood brain barrier (BBB) integrity. 29
2.4. Spinal cord injury
Spinal cord injury lead to serious dysfunction of the limbs and trunk below the injured segment. Previous studies on its treatment mainly focused on treatment timing, drug treatment, and complication treatment. In recent years, the application of physical intervention of I/HPC has attracted much. 30 IPC reduced paraplegia incidence and neuronal damage induced by spinal cord ischemia reperfusion injury in various models by attenuating blood spinal cord barrier (BSCB) disruption, 31 triggering spinal cord autoregulation, 32 and upregulating endogenous antioxidant enzymes. 33 The combination of HPC and stem cell therapy has a high translational value. HPC‐treated BMSC showed better cell survival rate and migration, along with increased neuron differentiation, enhanced paracrine effect, increased nutritional support, and improved functional recovery. 34 , 35 Mechanistically, HPC‐treated stem cells help shift microglial M1 to M2 polarization. 36 A recent study also suggests that activation of hypoxia inducible factor (HIF)‐1α played a critical for the survival of BMSCs after transplantation. 37 RIPC also attenuated motor deficits and histologic damage induced by ischemia reperfusion injury through various protective mechanisms, including suppressing BSCB disruption, 38 upregulating antioxidant enzyme activity 39 and preventing the increase of extracellular glutamate and subsequent excitotoxicity. 40
2.5. Others
In addition to the above diseases, the protective effects of I/HPC have also been studied in various other CNS diseases. Hypobaric HPC protected animals from stress‐related depression and anxiety. 41 HPC‐mediated molecular adaptation improved brain resistance to glutamate excitotoxicity in ethanol withdrawal. 42 HPC can also reduce brain edema induced by alginic acid‐induced status epilepticus in rats, which may be due to stress‐related transcription factors and effector proteins. 43 In addition, serial HPC can improve the cognitive functions in mice exposed to hypoxia. 44
3. NEUROPROTECTIVE MECHANISMS OF I/HPC
Conditional stimulations trigger protective responses through different sensors and signaling molecules, resulting in protective phenotypes in the brain. The mechanisms include interrelated biological pathways that minimize neuronal damage and promote the recovery through cascade of reaction. In this section, we will discuss the possible neuroprotective mechanisms related to I/HPC from the multiple aspects (Figure 2).
FIGURE 2.

Molecular mechanisms of IPC/HPC/RIPC treatment. Various critical molecules and mechanisms are involved in neuroprotective effects of IPC/HPC/RIPC treatment. AKT, protein kinase B; BAX, Bcl‐2‐associated X; BBB, blood brain barrier; Bcl‐2, B‐cell lymphoma‐2; CAT, catalase; EPO, erythropoietin; GLT, glutamate transporter; GPx, glutathione peroxidase; HIF, hypoxia inducible factor; HPC, hypoxic preconditioning; HSP70, heat‐shock protein 70; IFN, interferon; IL, interleukin; IPC, ischemic preconditioning; NCX, Na+–Ca2+ exchanger; NF‐κB, nuclear factor‐kappa B; Nrf2, erythroid 2‐related factor 2; NO, nitric oxide; PI3K, phosphatidylinositol 3‐kinase; PKC, protein kinase C; S1P, sphingosine‐1‐phosphate; SOD, superoxide dismutase; Sphk1, sphingosine kinase; Rab, ras‐related in brain; RIPC, remote ischemic preconditioning; TLR, toll‐like receptor; TNF, tumor necrosis factor; TRAIL, TNF‐related apoptosis inducing ligand; VEGF, vascular endothelial growth factor
3.1. Activating hypoxic signaling pathway
Hypoxia inducible factor‐1 is the major molecular of hypoxic response in the brain, it composed of oxygen sensitive α subunit and structurally stable β subunit. Under physiological conditions, HIF‐1α subunit is hydroxylated by proline hydroxylase (PHDs), which further promotes its binding with Von Hippel‐Lindau (VHL) complex, resulting in its ubiquitination and proteasomal degradation. Under hypoxic conditions, HIF‐1α combines with HIF‐1β to form a complex, which translocates to the nucleus and binds to the hypoxia response element on the target gene, resulting in the transcriptional activation of multiple genes, such as erythropoietin (EPO) and VEGF (Figure 3). 45
FIGURE 3.
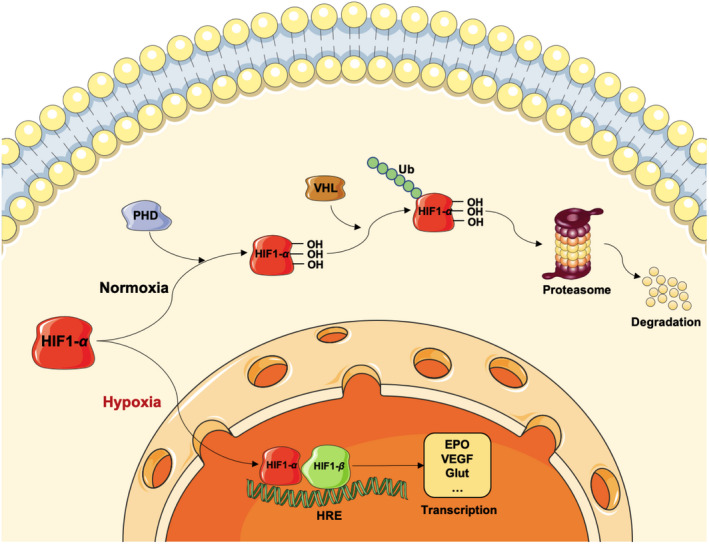
Molecular mechanisms of HIF‐1α mediated hypoxia response. Under normoxic conditions, HIF‐1α subunit is hydroxylated by PHD, which further promotes its binding with VHL complex, resulting in its ubiquitin and proteasomal degradation. Under hypoxic conditions, HIF‐1α combines with HIF‐1β to form a complex, which translocates to the nucleus and binds to HRE resulting in the transcription of multiple genes, such as EPO, VEGF, and Glut. EPO, erythropoietin; HIF, hypoxia inducible factor; HRE, hypoxia response element; PHD, proline hydroxylase; VEGF, vascular endothelial growth factor
HPC could significantly increase HIF‐1α level and its nuclear translocation, and whereby increase the expression of its target gene VEGF in neurons, endothelial cells, and astrocytes. 46 Some other HIF‐1 target genes were also required for HPC‐induced tolerance, such as cyclin‐dependent kinase inhibitor p21, whose deficiency abolished the neuroprotection of HPC. 47 Maintaining intracellular Ca2+ homeostasis is crucial to prevents Ca2+‐associated cell damage, IPC increases the expression of Na+‐Ca2+ exchanger (NCX) 1, which helps in this regard through HIF‐1 signaling. 48 Another mechanism via which IPC modulates Ca2+ homeostasis is through NCX1 and NCX3 upregulation mediated by nitric oxide (NO)/phosphatidylinositol 3‐kinase (PI3K)/protein kinase B (Akt) signaling. 49 , 50 Sumoylation of NCX3 stabilizes NCX3, and is regarded as a potential target in IPC‐induced neuroprotection. 51 Implantation of HPC‐treated hematopoietic stem cells improved stroke outcomes through promoting neuroplasticity mediated by HIF‐1α induction. 52 Interestingly, unlike hypoxia‐dependent mechanism in neurons, astrocytes enhance HIF‐1α expression through P2X7‐receptor‐dependent mechanism. 53
3.2. Antioxidant stress
Under normal circumstances, the body has an effective endogenous antioxidant defense system. Oxidative stress is a state of imbalance between oxidation and antioxidation in the body, it can be induced by excessive production of reactive oxygen species (ROS) or decreased ability of scavenging ROS. I/HPC and RIPC could decreases the levels of ROS and increases the levels of antioxidant enzymes to prevent from neuronal injury. IPC could increase catalase (CAT), glutathione peroxidase (GPx) and thioredoxin 2 activities to eliminate the excessive ROS in the hippocampal cornu ammonis (CA) 1 region. 54 Similarly, HPC increases activities of superoxide dismutase (SOD) and GPx in ischemic brain injury model. 55 RIPC reduces cerebral oxidative damage by increasing activity of CAT and reducing methane dicarboxylic aldehyde levels. 56 , 57 RIPC improve memory and cognitive function by enhancing SOD activity after hippocampal ischemia. 58
Despite regulating the above antioxidant enzymes, I/HPC and RIPC also reduced cerebral injury through antioxidant stress via various critical signaling pathways. Transcription factor erythroid 2‐related factor 2 (Nrf2) is a master redox regulator. HPC protects the brain against traumatic damage by upregulating Nrf2 level and suppressing oxidative stress damage. 59 In MCAO model, IPC alleviated motor deficits and cognitive impairment, accompanied by Nrf2 pathway activation, while these protective effects of IPC were abolished in Nrf2 knockout mice. 60 Nrf2 also played a critical role in IPC‐mediated blood‐brain barrier (BBB) preservation and neuroprotection. 61 Similarly, RIPC prevented mice from vascular dementia by increasing Nrf2 level to decrease oxidative stress. 62 Heat‐shock protein 70 (HSP70) is a cellular defense factor under stress, which can be upregulated by IHPC stimulus. 63 , 64 RIPC could mediate brain ischemic tolerance through activation of p38 mitogen‐activated protein kinase by upregulating HSP70 expression 65 and HIF‐1α/AMPK/HSP70 pathway. 66
In addition to the above critical molecules, I/HPC also played an antioxidant role through other ways. Mitochondrial respiratory chain is the main source of cellular ROS. In astrocytes, IPC promotes localization of Nrf2 on the mitochondrial outer membrane, thus preventing abnormal supercomplex formation and maintaining mitochondrial function. 67 IPC also regulate mitochondrial NAD+/NADH ratio through regulating nicotinamide phosphoribosyltransferase activity via protein kinase C (PKC) ɛ activation. 68 IPC facilitates the repair of oxidative DNA damage induced by ischemic injury through inducible DNA base‐excision repair. 69 RIPC protected neurons and mitochondria from oxidative damage in the porcine model of hypothermic ischemic insult. 70 It also reduced systemic oxidative stress by about 80% represented by lymphocytic DNA damage, and reduced circulating glutamate levels in rodents. 71 Furthermore, plasma from RIPC donor rabbits could also protect neural stem cells from oxidative stress and apoptosis through induction of thioredoxin, 72 and the involvement of adenosine A1 receptors also play a role. 73
3.3. Anti‐inflammation
Neuroinflammation is a double‐edged sword, appropriate duration and extent facilitate clearance of dead tissue and restoration of homeostasis, but excessive inflammatory response aggravates brain damage and affect long‐term neurological outcome. I/HPC and RIPC modulated immune response at various layers, including molecular, cellular, and systemic mechanisms to prevent from secondary neural injury (Figure 4). 74
FIGURE 4.

IPC/HPC/RIPC relieve neuroinflammation induced through central and peripheral immune cells. Neuroinflammation is involved in the pathogenesis of many neurological diseases. In the CNS, microglia or astrocytes activation could result in the release of inflammatory factors, such as TNF‐α, IL‐1β, and IL‐6. In addition, peripheral immune cells such as T lymphocytes and monocytes also infiltrate into CNS through BBB, which is usually destructive in most neurological diseases. The above process could be relieved by IPC/HPC/RIPC. BBB, blood brain barrier; CNS, central nervous system; HPC, hypoxic preconditioning; IL, interleukin; IPC, ischemic preconditioning; RIPC, remote ischemic preconditioning; TNF, tumor necrosis factor
As the brain resident immune cells, microglia are among the most important cells which orchestrate neuroinflammatory response. 75 , 76 HPC could suppress microglia abnormal activation and subsequent inflammatory responses after hypoxia‐ischemia insults. 77 In addition, conditioned medium from HPC‐treated BMSCs could switch microglia toward anti‐inflammatory polarization and alleviate microglia‐induced injury by inhibiting the levels of pro‐inflammatory cytokines, such tumor necrosis factor (TNF)‐α, and upregulating anti‐inflammatory cytokines, such as IL‐10. 78 Interestingly, IPC could induce cortical microglial proliferation dependent on fractalkine signaling. 79 RIPC inhibited inflammation by decreasing the levels of IL‐1β, IL‐6, and interferon‐γ in the ischemic brain. 80 Astrocytes are another type of glial cells which also exert immune regulation, 81 it mediate inflammatory effects by releasing neurotransmitters such as glutamate, and cytokines such as TNF‐α. IPC could reduce the damage of ischemia reperfusion effectively by reducing the release of astrocytic glutamate, which was further enhanced with astrocytic gap junction blockade. 82
Aside from cellular mediators, several signaling pathways also participate in the anti‐inflammatory effects of preconditioning treatment. Nuclear factor‐kappa B (NF‐κB) is a key player in mediating inflammation, exhibits a significant role in cerebral ischemic tolerance induced by I/HPC and RIPC. IPC could activate PKCε and ERK1/2 to promoted NF‐κB translocation to nucleus, 83 and RIPC‐mediated ischemic tolerance by activating NF‐κB pathway through interaction with Notch1 pathway. 84 On the contrary, there were also lots of studies suggested IPC suppressed NF‐κB activation. 85 , 86 IPC downregulated NF‐κB expression through inhibiting PI3K/Akt and ERK1/2 signaling pathways. As master regulators of innate immunity, toll‐like receptors (TLRs) play a critical role in CNS inflammatory response. 87 IPC reduces cerebral ischemic injury by inhibiting of the TLR4/NF‐κB signaling pathway. 88 Astrocytic TLR3 reprogramming also participates in IPC‐induced anti‐inflammation and ischemic tolerance. 89
I/HPC and RIPC also creates anti‐inflammatory effects through affecting chemotaxis of the peripheral immune cells. 90 HPC‐induced ischemic tolerance by decreasing early leukocyte infiltration dependent on a delay in C‐C motif chemokine ligand (CCL) 2 expression. 91 In addition, HPC‐induced chemokine (C‐X‐C motif) ligand 12 upregulation which suppressed leukocyte infiltration in tMCAO mice. 92 HPC also activated a novel immunosuppressed B‐cell phenotype to exert anti‐inflammatory effects in tMCAO mice model. 93 RIPC increased B cell in peripheral blood 90 and reduced T‐cell infiltration into the ischemic brain, accompanied by increased p‐ERK expression. 94 Another study also suggested RIPC exerted neuroprotection against cerebral ischemia mainly by modulating the spleen‐derived lymphocytes. 9
3.4. Anti‐apoptosis
Apoptosis is an active and orderly cell death process, 95 I/HPC and RIPC could protect against brain injury by inhibiting apoptosis. The expression of anti‐apoptotic genes increased after HPC treatment in hippocampal slice cultures. 96 IPC could downregulate pro‐apoptosis protein BCL‐2‐associated X (Bax) level, but upregulate anti‐apoptosis protein Bcl‐2 level, and further decrease cleaved caspase‐9 and caspase‐3. 85 , 97 It also blocked the ischemia‐induced mitochondrial translocation of Bad, a Bcl‐2 family member, via PI3K/Akt signaling, inhibiting apoptosis of CA1 pyramidal cells. 98 IPC prevented the opening of mitochondrial permeability transition pore (mPTP) and the releasing of cytochrome c mediated by nitrite. 99 Similarly, RIPC also decreased apoptosis of hippocampal neurons by improving the integrity of the mitochondrial membrane and inhibiting mPTP opening. 100 Endoplasmic reticulum (ER) stress is a strong inducer of apoptosis. IPC inhibited ER stress‐induced apoptosis through protein kinase RNA (PKR) like ER kinase pathway. 101 IPC could also downregulate TNF‐related apoptosis inducing ligand, a critical death receptor. 102
3.5. Reducing excitotoxicity
Excitotoxicity is a toxic process mainly caused by excessive excitatory neurotransmitter glutamate. 103 Thorase is important to maintain mitochondrial function and regulate surface glutamate receptor activity. IPC‐induced thorase expression to provide neuroprotection against N‐methyl‐D‐aspartic acid (NMDA) receptor‐mediated excitotoxicity. 104 IPC‐treated astrocytes could also confer ischemic tolerance to neurons associated with increased neuronal tolerance to NMDA. 105 Glutamate homeostasis in the CNS is maintained through uptake of excessive glutamate by excitatory amino acid transporter known as glial glutamate transporter (GLT)‐1. GLT‐1 is mainly located on astrocytes, and has been regarded as a potential therapeutic target in the treatment of brain ischemic injury, IPC can reduce glutamate excitotoxicity by upregulation of GLT‐1 activity in glial cells, thus inducing cerebral ischemic tolerance. 106 HPC also reversed the downregulation of GLT‐1 protein caused by global cerebral ischemia. 107
3.6. Activating autophagy
Autophagy is an evolutionarily conservative process crucial for cell survival, in the circumstances of hunger, infection and stress, autophagy contributes to homeostasis by removing aggregated proteins and damaged organelles, rapidly providing fuel supply for energy, and delaying cell death. In ischemia model, HPC increased the production and degradation of autophagosomes and resisted to subsequent fatal injury. 108 HPC also promoted autophagosome maturation by activating ras‐related in brain 7, a lysosome‐related protein, to protect against global cerebral ischemia‐induced injury. 109 HIF‐1α/Beclin1 signaling pathway activation was also involved in autophagy induction during HPC treatment. 110 Conventional PKCγ signaling molecules especially PKCγ‐synapsin pathway were proved to facilitate HPC‐mediated protection, 111 and PKCγ could modulate neuron‐specific autophagy through the Akt‐mTOR pathway MCAO mice model. 112 Similarly, IPC increased the levels of autophagy related proteins, including microtubule‐associated protein 1 light chain (LC) 3 II and beclin1, which were suppressed by autophagy inhibitors 3‐methyladenine and bafilomycin A1, and enhanced by autophagy agonist rapamycin, confirming the critical role of autophagy in IPC. 113 IPC also protected against neuronal injury via ER stress‐induced autophagy, proved by abolished neuroprotection with ER stress inhibitor salubrinal. 114
3.7. Others
Besides the above protective mechanisms, other mechanisms associated with I/HPC include but not limited to improving synaptic plasticity, modulating Ca2+ homeostasis, and preserving BBB function. IPC improved synaptic plasticity by increasing BDNF mRNA expression dependent on PKCɛ activation. 115 Meanwhile, IPC‐induced PKCɛ activation enhances GABA release, contribute to monoamine balance of the brain. 116 Ca2+ homeostasis was maintained by IPC through modulating the interaction of the ER‐located Ca2+ sensor stromal interacting molecule 1 with the plasma membrane channel ORAI1, 117 as well as activation of the NO/PI3K/Akt pathway. 49 Impairment of BBB integrity is a critical event in the pathogenesis of ischemic/hypoxic injury. 118 HPC could induce de novo formation of cerebral collaterals, which lessens the severity of a subsequent stroke event. 119 HPC also upregulates the expression of vascular genes, whereby increasing vascular density and cerebral blood flow. 120 HPC could also active sphingosine kinase 2 in a region‐specific manner, whose products sphingosine‐1‐phosphate is critical for vascular functioning. 121 , 122 , 123 Astrocytes are proved to be major mediators in IPC‐mediated BBB preservation in vitro. 124 HPC could also help restore the maturation capacity in oligodendrocyte precursor cells in neonatal rats subjected to hypoxia/ischemia insults. 125
In addition to the proteins involved in the above mechanisms, some other molecules have also been proposed as potential targets of I/HPC. Heme oxygenase 1 (HO1), an anti‐oxidant enzyme, is significantly increased by IPC, and HO1 knockout could abolish IPC‐induced protective effects on ischemic brain injury, indicating its critical role in IPC. 126 Considerably, IPC protects hippocampal pyramidal neurons from ischemic injury by HO1‐mediated suppression of oxidative damage. 127 An apoptotic inhibitory molecule, cellular inhibitor of apoptosis 1, was also implicated in IPC in neurons and endothelial cells. 128 Epigenetic studies by microarray analysis suggested that methyl‐CpG binding protein 2 was also a prominent target in IPC‐induced tolerance. 129
4. LIMITATIONS AND CHALLENGES
Although both of I/HPC and RIPC have shown considerable protective effects in lots of CNS diseases, there are still many limitations and challenges to translate their clinical applications from basic research. Firstly, heterogeneity of population subgroups needs to be considered. Different individual factors, such as age, gender, race, and comorbid medical conditions require different degrees of conditioning treatment to induce optimal stimulate. In addition to these individual factors, different diseases should also be treated in different ways. In other words, the optimal duration and frequency of pretreatment stimuli vary from disease to disease, and different hypoxic levels, duration, and onset cycle may have very different effects. Secondly, timeliness is also a main limitation of I/HPC and RIPC, because their benefits usually last only a few days, while the onset of diseases is unpredictable. Therefore, patients should be stratified based on their risk for each individual disease, and different therapeutic strategy should depend on the pathogenesis individually. The combination therapy with preconditioning and postconditioning may be a promising direction. 130 In addition, specific biomarkers that respond to preconditioning treatment may be useful to guide optimal therapeutic strategy and further assess efficacy. Furthermore, cross adaptation or cross tolerance is also a growing field of interest, it refers to the phenomenon that one type of conditioning could establish tolerance toward another type of injury. 131 It suggests that I/HPC may play a role in more fields, which need to be further explored. Finally, although application of RIPC is practical in high‐risk population, while it maybe not practical to induce ischemia/hypoxia in healthy humans, so more mechanistic studies can help us achieve the protective effect through other methods, such as pharmacological treatment.
5. SUMMARY
Due to the complicated etiology of neurological diseases, the current treatment strategies are still limited. In addition, many neurological diseases come on gradually, so once diagnosed, the best treatment window is missed. Therefore, preconditioning therapy is a promising direction in the treatment of neurological diseases. Specific molecular mechanisms of I/HPC or RIPC need to be further explored, and their limitations and challenges need to be addressed as well.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
Jia Liu and Yakun Gu contributed equally to this work.
Funding information
This review was supported by the Beijing Hundred Thousand and Ten Thousand Talent Project (Grant number: 2019A36), the National Key R&D Program of China (Grant number: 2016YFC1301502), and Beijing Municipal Health Commission (Grant number: 303–01–005‐0019).
DATA AVAILABILITY STATEMENT
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
REFERENCES
- 1. Dahl NA, Balfour WM. Prolonged anoxic survival due to anoxia pre‐exposure: brain Atp, lactate, and pyruvate. Am J Physiol. 1964;207:452‐456. 10.1152/ajplegacy.1964.207.2.452 [DOI] [PubMed] [Google Scholar]
- 2. Kitagawa K, Matsumoto M, Tagaya M, et al. ‘Ischemic tolerance’ phenomenon found in the brain. Brain Res. 1990;528(1):21‐24. 10.1016/0006-8993(90)90189-i [DOI] [PubMed] [Google Scholar]
- 3. Li S, Hafeez A, Noorulla F, et al. Preconditioning in neuroprotection: From hypoxia to ischemia. Prog Neurogibol. 2017;157:79‐91. 10.1016/j.pneurobio.2017.01.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Basalay MV, Davidson SM, Gourine AV, Yellon DM. Neural mechanisms in remote ischaemic conditioning in the heart and brain: mechanistic and translational aspects. Basic Res Cardiol. 2018;113(4):25. 10.1007/s00395-018-0684-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Grotta JC. tPA for stroke: important progress in achieving faster treatment. JAMA. 2014;311(16):1615‐1617. 10.1001/jama.2014.3322 [DOI] [PubMed] [Google Scholar]
- 6. Dirnagl U, Becker K, Meisel A. Preconditioning and tolerance against cerebral ischaemia: from experimental strategies to clinical use. Lancet Neurol. 2009;8(4):398‐412. 10.1016/S1474-4422(09)70054-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Bernaudin M, Nedelec AS, Divoux D, MacKenzie ET, Petit E, Schumann‐Bard P. Normobaric hypoxia induces tolerance to focal permanent cerebral ischemia in association with an increased expression of hypoxia‐inducible factor‐1 and its target genes, erythropoietin and VEGF, in the adult mouse brain. J Cereb Blood Flow Metab. 2002;22(4):393‐403. 10.1097/00004647-200204000-00003 [DOI] [PubMed] [Google Scholar]
- 8. Wang WW, Chen DZ, Zhao M, Yang XF, Gong DR. Prior transient ischemic attacks may have a neuroprotective effect in patients with ischemic stroke. Arch Med Sci. 2017;13(5):1057‐1061. 10.5114/aoms.2016.63744 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Chen C, Jiang W, Liu Z, et al. Splenic responses play an important role in remote ischemic preconditioning‐mediated neuroprotection against stroke. J Neuroinflammation. 2018;15(1):167. 10.1186/s12974-018-1190-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Yang J, Shakil F, Cho S. Peripheral mechanisms of remote ischemic conditioning. Cond Med. 2019;2(2):61‐68. [PMC free article] [PubMed] [Google Scholar]
- 11. Chong J, Bulluck H, Fw Ho A, Boisvert WA, Hausenloy DJ. Chronic remote ischemic conditioning for cardiovascular protection. Cond Med. 2019;2(4):164‐169. [PMC free article] [PubMed] [Google Scholar]
- 12. Koch S, Gonzalez N. Preconditioning the human brain: proving the principle in subarachnoid hemorrhage. Stroke. 2013;44(6):1748‐1753. 10.1161/STROKEAHA.111.000773 [DOI] [PubMed] [Google Scholar]
- 13. Koch S, Katsnelson M, Dong C, Perez‐Pinzon M. Remote ischemic limb preconditioning after subarachnoid hemorrhage: a phase Ib study of safety and feasibility. Stroke. 2011;42(5):1387‐1391. 10.1161/STROKEAHA.110.605840 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Laiwalla AN, Ooi YC, Liou R, Gonzalez NR. Matched cohort analysis of the effects of limb remote ischemic conditioning in patients with aneurysmal subarachnoid hemorrhage. Transl Stroke Res. 2016;7(1):42‐48. 10.1007/s12975-015-0437-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. He Y, Karabiyikoglu M, Hua Y, Keep RF, Xi G. Ischemic preconditioning attenuates brain edema after experimental intracerebral hemorrhage. Transl Stroke Res. 2012;3(1 Suppl 1):180‐187. 10.1007/s12975-012-0171-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Chen J, Yang Y, Shen L, et al. Hypoxic preconditioning augments the therapeutic efficacy of bone marrow stromal cells in a rat Ischemic Stroke Model. Cell Mol Neurobiol. 2017;37(6):1115‐1129. 10.1007/s10571-016-0445-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Wakai T, Narasimhan P, Sakata H, et al. Hypoxic preconditioning enhances neural stem cell transplantation therapy after intracerebral hemorrhage in mice. J Cereb Blood Flow Metab. 2016;36(12):2134‐2145. 10.1177/0271678X15613798 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Sun J, Wei ZZ, Gu X, et al. Intranasal delivery of hypoxia‐preconditioned bone marrow‐derived mesenchymal stem cells enhanced regenerative effects after intracerebral hemorrhagic stroke in mice. Exp Neurol. 2015;272:78‐87. 10.1016/j.expneurol.2015.03.011 [DOI] [PubMed] [Google Scholar]
- 19. Nalivaeva NN, Rybnikova EA. Editorial: brain hypoxia and ischemia: new insights into neurodegeneration and neuroprotection. Front Neurosci. 2019;13:770. 10.3389/fnins.2019.00770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Manukhina EB, Downey HF, Shi X, Mallet RT. Intermittent hypoxia training protects cerebrovascular function in Alzheimer's disease. Exp Biol Med. 2016;241(12):1351‐1363. 10.1177/1535370216649060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Meng SX, Wang B, Li WT. Intermittent hypoxia improves cognition and reduces anxiety‐related behavior in APP/PS1 mice. Brain Behav. 2020;10(2):e01513. 10.1002/brb3.1513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Serebrovska ZO, Serebrovska TV, Kholin VA, et al. Intermittent hypoxia‐hyperoxia training improves cognitive function and decreases circulating biomarkers of Alzheimer's disease in patients with mild cognitive impairment: a pilot study. Int J Mol Sci. 2019;20(21):5405. 10.3390/ijms20215405 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 23. Leak RK. Conditioning against the pathology of Parkinson's disease. Cond Med. 2018;1(3):143‐162. [PMC free article] [PubMed] [Google Scholar]
- 24. Lunemann JD, Ruck T, Muraro PA, Bar‐Or A, Wiendl H. Immune reconstitution therapies: concepts for durable remission in multiple sclerosis. Nat Rev Neurol. 2020;16(1):56‐62. 10.1038/s41582-019-0268-z [DOI] [PubMed] [Google Scholar]
- 25. Lassmann H, Bradl M. Multiple sclerosis: experimental models and reality. Acta Neuropathol. 2017;133(2):223‐244. 10.1007/s00401-016-1631-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Halder SK, Milner R. Chronic mild hypoxia accelerates recovery from preexisting EAE by enhancing vascular integrity and apoptosis of infiltrated monocytes. Proc Natl Acad Sci U S A. 2020;117(20):11126‐11135. 10.1073/pnas.1920935117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Halder SK, Milner R. A critical role for microglia in maintaining vascular integrity in the hypoxic spinal cord. Proc Natl Acad Sci U S A. 2019;116(51):26029‐26037. 10.1073/pnas.1912178116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Esen N, Katyshev V, Serkin Z, Katysheva S, Dore‐Duffy P. Endogenous adaptation to low oxygen modulates T‐cell regulatory pathways in EAE. J Neuroinflammation. 2016;13:13. 10.1186/s12974-015-0407-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Halder SK, Kant R, Milner R. Hypoxic pre‐conditioning suppresses experimental autoimmune encephalomyelitis by modifying multiple properties of blood vessels. Acta Neuropathol Commun. 2018;6(1):86. 10.1186/s40478-018-0590-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Baillieul S, Chacaroun S, Doutreleau S, Detante O, Pepin JL, Verges S. Hypoxic conditioning and the central nervous system: a new therapeutic opportunity for brain and spinal cord injuries? Exp Biol Med. 2017;242(11):1198‐1206. 10.1177/1535370217712691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Fang BO, Li X‐M, Sun X‐J, et al. Ischemic preconditioning protects against spinal cord ischemia‐reperfusion injury in rabbits by attenuating blood spinal cord barrier disruption. Int J Mol Sci. 2013;14(5):10343‐10354. 10.3390/ijms140510343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Liang CL, Lu K, Liliang PC, Chen TB, Chan SH, Chen HJ. Ischemic preconditioning ameliorates spinal cord ischemia‐reperfusion injury by triggering autoregulation. J Vasc Surg. 2012;55(4):1116‐1123. 10.1016/j.jvs.2011.09.096 [DOI] [PubMed] [Google Scholar]
- 33. Song W, Sun J, Su B, Yang R, Dong H, Xiong L. Ischemic postconditioning protects the spinal cord from ischemia‐reperfusion injury via modulation of redox signaling. J Thorac Cardiovasc Surg. 2013;146(3):688‐695. 10.1016/j.jtcvs.2012.11.039 [DOI] [PubMed] [Google Scholar]
- 34. Fan WL, Liu P, Wang G, Pu JG, Xue X, Zhao JH. Transplantation of hypoxic preconditioned neural stem cells benefits functional recovery via enhancing neurotrophic secretion after spinal cord injury in rats. J Cell Biochem. 2018;119(6):4339‐4351. 10.1002/jcb.26397 [DOI] [PubMed] [Google Scholar]
- 35. Wang W, Huang X, Lin W, et al. Hypoxic preconditioned bone mesenchymal stem cells ameliorate spinal cord injury in rats via improved survival and migration. Int J Mol Med. 2018;42(5):2538‐2550. 10.3892/ijmm.2018.3810 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 36. Liu W, Rong Y, Wang J, et al. Exosome‐shuttled miR‐216a‐5p from hypoxic preconditioned mesenchymal stem cells repair traumatic spinal cord injury by shifting microglial M1/M2 polarization. J Neuroinflammation. 2020;17(1):47. 10.1186/s12974-020-1726-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Luo Z, Wu F, Xue E, et al. Hypoxia preconditioning promotes bone marrow mesenchymal stem cells survival by inducing HIF‐1alpha in injured neuronal cells derived exosomes culture system. Cell Death Dis. 2019;10(2):134. 10.1038/s41419-019-1410-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Yu Q, Huang J, Hu J, Zhu H. Advance in spinal cord ischemia reperfusion injury: Blood‐spinal cord barrier and remote ischemic preconditioning. Life Sci. 2016;154:34‐38. 10.1016/j.lfs.2016.03.046 [DOI] [PubMed] [Google Scholar]
- 39. Dong H‐L, Zhang YI, Su B‐X, et al. Limb remote ischemic preconditioning protects the spinal cord from ischemia‐reperfusion injury: a newly identified nonneuronal but reactive oxygen species‐dependent pathway. Anesthesiology. 2010;112(4):881‐891. 10.1097/ALN.0b013e3181d0486d [DOI] [PubMed] [Google Scholar]
- 40. Mukai A, Suehiro K, Kimura A, et al. Protective effects of remote ischemic preconditioning against spinal cord ischemia‐reperfusion injury in rats. J Thorac Cardiovasc Surg. 2020. S0022‐5223(20):30793‐5. 10.1016/j.jtcvs.2020.03.094 [DOI] [PubMed] [Google Scholar]
- 41. Rybnikova E, Mironova V, Pivina S, et al. Antidepressant‐like effects of mild hypoxia preconditioning in the learned helplessness model in rats. Neurosci Lett. 2007;417(3):234‐239. 10.1016/j.neulet.2007.02.048 [DOI] [PubMed] [Google Scholar]
- 42. Jung ME, Mallet RT. Intermittent hypoxia training: Powerful, non‐invasive cerebroprotection against ethanol withdrawal excitotoxicity. Respir Physiol Neurobiol. 2018;256:67‐78. 10.1016/j.resp.2017.08.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Emerson MR, Nelson SR, Samson FE, Pazdernik TL. Hypoxia preconditioning attenuates brain edema associated with kainic acid‐induced status epilepticus in rats. Brain Res. 1999;825(1–2):189‐193. 10.1016/s0006-8993(99)01195-6 [DOI] [PubMed] [Google Scholar]
- 44. Shao G, Zhang R, Wang ZL, Gao CY, Huo X, Lu GW. Hypoxic preconditioning improves spatial cognitive ability in mice. Neurosignals. 2006;15(6):314‐321. 10.1159/000121368 [DOI] [PubMed] [Google Scholar]
- 45. Choudhry H, Harris AL. Advances in hypoxia‐inducible factor biology. Cell Metab. 2018;27(2):281‐298. 10.1016/j.cmet.2017.10.005 [DOI] [PubMed] [Google Scholar]
- 46. Ara J, Fekete S, Frank M, Golden JA, Pleasure D, Valencia I. Hypoxic‐preconditioning induces neuroprotection against hypoxia‐ischemia in newborn piglet brain. Neurobiol Dis. 2011;43(2):473‐485. 10.1016/j.nbd.2011.04.021 [DOI] [PubMed] [Google Scholar]
- 47. Mergenthaler P, Muselmann C, Sunwoldt J, et al. A functional role of the cyclin‐dependent kinase inhibitor 1 (p21(WAF1/CIP1)) for neuronal preconditioning. J Cereb Blood Flow Metab. 2013;33(3):351‐355. 10.1038/jcbfm.2012.213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Formisano L, Guida N, Valsecchi V, et al. Sp3/REST/HDAC1/HDAC2 complex represses and Sp1/HIF‐1/p300 complex activates ncx1 Gene transcription, in brain ischemia and in ischemic brain preconditioning, by epigenetic mechanism. J Neurosci. 2015;35(19):7332‐7348. 10.1523/JNEUROSCI.2174-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Sisalli MJ, Secondo A, Esposito A, et al. Endoplasmic reticulum refilling and mitochondrial calcium extrusion promoted in neurons by NCX1 and NCX3 in ischemic preconditioning are determinant for neuroprotection. Cell Death Differ. 2014;21(7):1142‐1149. 10.1038/cdd.2014.32 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Pignataro G, Boscia F, Esposito E, et al. NCX1 and NCX3: two new effectors of delayed preconditioning in brain ischemia. Neurobiol Dis. 2012;45(1):616‐623. 10.1016/j.nbd.2011.10.007 [DOI] [PubMed] [Google Scholar]
- 51. Cuomo O, Pignataro G, Sirabella R, et al. Sumoylation of LYS590 of NCX3 f‐Loop by SUMO1 participates in brain neuroprotection induced by ischemic preconditioning. Stroke. 2016;47(4):1085‐1093. 10.1161/STROKEAHA.115.012514 [DOI] [PubMed] [Google Scholar]
- 52. Lin CH, Lee HT, Lee SD, et al. Role of HIF‐1alpha‐activated Epac1 on HSC‐mediated neuroplasticity in stroke model. Neurobiol Dis. 2013;58:76‐91. 10.1016/j.nbd.2013.05.006 [DOI] [PubMed] [Google Scholar]
- 53. Hirayama Y, Koizumi S. Hypoxia‐independent mechanisms of HIF‐1alpha expression in astrocytes after ischemic preconditioning. Glia. 2017;65(3):523‐530. 10.1002/glia.23109 [DOI] [PubMed] [Google Scholar]
- 54. Lee J‐C, Park JH, Kim IH, et al. Neuroprotection of ischemic preconditioning is mediated by thioredoxin 2 in the hippocampal CA1 region following a subsequent transient cerebral ischemia. Brain Pathol. 2017;27(3):276‐291. 10.1111/bpa.12389 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Alkan T, Goren B, Vatansever E, Sarandol E. Effects of hypoxic preconditioning in antioxidant enzyme activities in hypoxic‐ischemic brain damage in immature rats. Turk Neurosurg. 2008;18(2):165‐171. [PubMed] [Google Scholar]
- 56. Mehrjerdi FZ, Aboutaleb N, Pazoki‐Toroudi H, et al. The protective effect of remote renal preconditioning against hippocampal ischemia reperfusion injury: role of KATP channels. J Mol Neurosci. 2015;57(4):554‐560. 10.1007/s12031-015-0636-0 [DOI] [PubMed] [Google Scholar]
- 57. Yan Y, Tong F, Chen J. Endogenous BMP‐4/ROS/COX‐2 mediated IPC and resveratrol alleviated brain damage. Curr Pharm Des. 2019;25(9):1030‐1039. 10.2174/1381612825666190506120611 [DOI] [PubMed] [Google Scholar]
- 58. Zare Mehrjerdi F, Aboutaleb N, Habibey R, et al. Increased phosphorylation of mTOR is involved in remote ischemic preconditioning of hippocampus in mice. Brain Res. 2013;1526:94‐101. 10.1016/j.brainres.2013.06.018 [DOI] [PubMed] [Google Scholar]
- 59. Shu L, Wang C, Wang J, et al. The neuroprotection of hypoxic preconditioning on rat brain against traumatic brain injury by up‐regulated transcription factor Nrf2 and HO‐1 expression. Neurosci Lett. 2016;611:74‐80. 10.1016/j.neulet.2015.11.012 [DOI] [PubMed] [Google Scholar]
- 60. Yang T, Sun Y, Li Q, et al. Ischemic preconditioning provides long‐lasting neuroprotection against ischemic stroke: the role of Nrf2. Exp Neurol. 2020;325:113142. 10.1016/j.expneurol.2019.113142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Yang T, Sun Y, Mao L, et al. Brain ischemic preconditioning protects against ischemic injury and preserves the blood‐brain barrier via oxidative signaling and Nrf2 activation. Redox Biol. 2018;17:323‐337. 10.1016/j.redox.2018.05.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. He JT, Li H, Yang L, Cheng KL. Involvement of endothelin‐1, H2S and Nrf2 in beneficial effects of remote ischemic preconditioning in global cerebral ischemia‐induced vascular dementia in mice. Cell Mol Neurobiol. 2019;39(5):671‐686. 10.1007/s10571-019-00670-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Racay P. Ischaemia‐induced protein ubiquitinylation is differentially accompanied with heat‐shock protein 70 expression after naive and preconditioned ischaemia. Cell Mol Neurobiol. 2012;32(1):107‐119. 10.1007/s10571-011-9740-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Ge PF, Luo TF, Zhang JZ, Chen DW, Luan YX, Fu SL. Ischemic preconditioning induces chaperone hsp70 expression and inhibits protein aggregation in the CA1 neurons of rats. Neurosci Bull. 2008;24(5):288‐296. 10.1007/s12264-008-0623-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Sun X‐C, Xian X‐H, Li W‐B, et al. Activation of p38 MAPK participates in brain ischemic tolerance induced by limb ischemic preconditioning by up‐regulating HSP 70. Exp Neurol. 2010;224(2):347‐355. 10.1016/j.expneurol.2010.04.009 [DOI] [PubMed] [Google Scholar]
- 66. Xia M, Ding Q, Zhang Z, Feng Q. Remote limb ischemic preconditioning protects rats against cerebral ischemia via HIF‐1alpha/AMPK/HSP70 pathway. Cell Mol Neurobiol. 2017;37(6):1105‐1114. 10.1007/s10571-016-0444-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Narayanan SV, Dave KR, Perez‐Pinzon MA. Ischemic preconditioning protects astrocytes against oxygen glucose deprivation via the Nuclear Erythroid 2‐Related Factor 2 pathway. Transl Stroke Res. 2018;9(2):99‐109. 10.1007/s12975-017-0574-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Morris‐Blanco KC, Cohan CH, Neumann JT, Sick TJ, Perez‐Pinzon MA. Protein kinase C epsilon regulates mitochondrial pools of Nampt and NAD following resveratrol and ischemic preconditioning in the rat cortex. J Cereb Blood Flow Metab. 2014;34(6):1024‐1032. 10.1038/jcbfm.2014.51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Li W, Luo Y, Zhang F, et al. Ischemic preconditioning in the rat brain enhances the repair of endogenous oxidative DNA damage by activating the base‐excision repair pathway. J Cereb Blood Flow Metab. 2006;26(2):181‐198. 10.1038/sj.jcbfm.9600180 [DOI] [PubMed] [Google Scholar]
- 70. Arvola O, Haapanen H, Herajärvi J, et al. Remote ischemic preconditioning reduces cerebral oxidative stress following hypothermic circulatory arrest in a Porcine Model. Semin Thorac Cardiovasc Surg. Spring. 2016;28(1):92‐102. 10.1053/j.semtcvs.2016.01.005 [DOI] [PubMed] [Google Scholar]
- 71. Jachova J, Gottlieb M, Nemethova M, Macakova L, Bona M, Bonova P. Neuroprotection mediated by remote preconditioning is associated with a decrease in systemic oxidative stress and changes in brain and blood glutamate concentration. Neurochem Int. 2019;129:104461. 10.1016/j.neuint.2019.05.005 [DOI] [PubMed] [Google Scholar]
- 72. Motomura A, Shimizu M, Kato A, et al. Remote ischemic preconditioning protects human neural stem cells from oxidative stress. Apoptosis. 2017;22(11):1353‐1361. 10.1007/s10495-017-1425-8 [DOI] [PubMed] [Google Scholar]
- 73. Hu S, Dong H, Zhang H, et al. Noninvasive limb remote ischemic preconditioning contributes neuroprotective effects via activation of adenosine A1 receptor and redox status after transient focal cerebral ischemia in rats. Brain Res. 2012;1459:81‐90. 10.1016/j.brainres.2012.04.017 [DOI] [PubMed] [Google Scholar]
- 74. McDonough A, Weinstein JR. Neuroimmune response in ischemic preconditioning. Neurotherapeutics. 2016;13(4):748‐761. 10.1007/s13311-016-0465-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Yenari MA. Microglia, the brain’s double agent. J Cereb Blood Flow Metab. 2020;40(1_suppl):S3‐S5. 10.1177/0271678X20968993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Zhang W, Bhatia TN, Leak RK. Functional diversities of myeloid cells in the central nervous system. CNS Neurosci Ther. 2020;26(12):1205‐1206. 10.1111/cns.13525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Huang L, Wu S, Li H, Dang Z, Wu Y. Hypoxic preconditioning relieved ischemic cerebral injury by promoting immunomodulation and microglia polarization after middle cerebral artery occlusion in rats. Brain Res. 2019;1723:146388. 10.1016/j.brainres.2019.146388 [DOI] [PubMed] [Google Scholar]
- 78. Yu H, Xu Z, Qu G, et al. Hypoxic preconditioning enhances the efficacy of mesenchymal stem cells‐derived conditioned medium in switching microglia toward anti‐inflammatory polarization in ischemia/reperfusion. Cell Mol Neurobiol. 2021;41(3):505‐524. 10.1007/s10571-020-00868-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. McDonough A, Noor S, Lee RV, et al. Ischemic preconditioning induces cortical microglial proliferation and a transcriptomic program of robust cell cycle activation. Glia. 2020;68(1):76‐94. 10.1002/glia.23701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Du X, Yang J, Liu C, et al. Hypoxia‐inducible factor 1alpha and 2alpha have beneficial effects in remote ischemic preconditioning against stroke by modulating inflammatory responses in aged rats. Front Aging Neurosci. 2020;12:54. 10.3389/fnagi.2020.00054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Zhou B, Zuo YX, Jiang RT. Astrocyte morphology: Diversity, plasticity, and role in neurological diseases. CNS Neurosci Ther. 2019;25(6):665‐673. 10.1111/cns.13123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Ma D, Feng L, Cheng Y, et al. Astrocytic gap junction inhibition by carbenoxolone enhances the protective effects of ischemic preconditioning following cerebral ischemia. J Neuroinflammation. 2018;15(1):198. 10.1186/s12974-018-1230-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Kim EJ, Raval AP, Hirsch N, Perez‐Pinzon MA. Ischemic preconditioning mediates cyclooxygenase‐2 expression via nuclear factor‐kappa B activation in mixed cortical neuronal cultures. Transl Stroke Res. 2010;1(1):40‐47. 10.1007/s12975-009-0006-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Liang W, Lin C, Yuan L, et al. Preactivation of Notch1 in remote ischemic preconditioning reduces cerebral ischemia‐reperfusion injury through crosstalk with the NF‐kappaB pathway. J Neuroinflammation. 2019;16(1):181. 10.1186/s12974-019-1570-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Shi S, Yang W, Tu X, Chen C, Wang C. Ischemic preconditioning reduces ischemic brain injury by suppressing nuclear factor kappa B expression and neuronal apoptosis. Neural Regen Res. 2013;8(7):633‐638. 10.3969/j.issn.1673-5374.2013.07.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Liang J, Luan Y, Lu B, Zhang H, Luo YN, Ge P. Protection of ischemic postconditioning against neuronal apoptosis induced by transient focal ischemia is associated with attenuation of NF‐kappaB/p65 activation. PLoS One. 2014;9(5):e96734. 10.1371/journal.pone.0096734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Gesuete R, Kohama SG, Stenzel‐Poore MP. Toll‐like receptors and ischemic brain injury. J Neuropathol Exp Neurol. 2014;73(5):378‐386. 10.1097/NEN.0000000000000068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Wang PF, Xiong XY, Chen J, Wang YC, Duan W, Yang QW. Function and mechanism of toll‐like receptors in cerebral ischemic tolerance: from preconditioning to treatment. J Neuroinflammation. 2015;12:80. 10.1186/s12974-015-0301-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Pan L‐N, Zhu W, Li Y, et al. Astrocytic Toll‐like receptor 3 is associated with ischemic preconditioning‐induced protection against brain ischemia in rodents. PLoS One. 2014;9(6):e99526. 10.1371/journal.pone.0099526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Liu Z‐J, Chen C, Li X‐R, et al. Remote ischemic preconditioning‐mediated neuroprotection against stroke is associated with significant alterations in peripheral immune responses. CNS Neurosci Ther. 2016;22(1):43‐52. 10.1111/cns.12448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Stowe AM, Wacker BK, Cravens PD, et al. CCL2 upregulation triggers hypoxic preconditioning‐induced protection from stroke. J Neuroinflammation. 2012;9(1):33. 10.1186/1742-2094-9-33 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Selvaraj UM, Ortega SB, Hu R, et al. Preconditioning‐induced CXCL12 upregulation minimizes leukocyte infiltration after stroke in ischemia‐tolerant mice. J Cereb Blood Flow Metab. 2017;37(3):801‐813. 10.1177/0271678X16639327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Monson NL, Ortega SB, Ireland SJ, et al. Repetitive hypoxic preconditioning induces an immunosuppressed B cell phenotype during endogenous protection from stroke. J Neuroinflammation. 2014;11(1):22. 10.1186/1742-2094-11-22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Liu C, Yang J, Zhang C, Geng X, Zhao H. Remote ischemic conditioning reduced cerebral ischemic injury by modulating inflammatory responses and ERK activity in type 2 diabetic mice. Neurochem Int. 2020;135:104690. 10.1016/j.neuint.2020.104690 [DOI] [PubMed] [Google Scholar]
- 95. Singh R, Letai A, Sarosiek K. Regulation of apoptosis in health and disease: the balancing act of BCL‐2 family proteins. Nat Rev Mol Cell Biol. 2019;20(3):175‐193. 10.1038/s41580-018-0089-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Bickler PE, Fahlman CS. Expression of signal transduction genes differs after hypoxic or isoflurane preconditioning of rat hippocampal slice cultures. Anesthesiology. 2009;111(2):258‐266. 10.1097/ALN.0b013e3181a8647f [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Ding Z‐M, Wu B, Zhang W‐Q, et al. Neuroprotective effects of ischemic preconditioning and postconditioning on global brain ischemia in rats through the same effect on inhibition of apoptosis. Int J Mol Sci. 2012;13(5):6089‐6101. 10.3390/ijms13056089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Miyawaki T, Mashiko T, Ofengeim D, et al. Ischemic preconditioning blocks BAD translocation, Bcl‐xL cleavage, and large channel activity in mitochondria of postischemic hippocampal neurons. Proc Natl Acad Sci U S A. 2008;105(12):4892‐4897. 10.1073/pnas.0800628105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Shiva S, Sack MN, Greer JJ, et al. Nitrite augments tolerance to ischemia/reperfusion injury via the modulation of mitochondrial electron transfer. J Exp Med. 2007;204(9):2089‐2102. 10.1084/jem.20070198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Zhou X, Yong L, Huang Y, et al. The protective effects of distal ischemic treatment on apoptosis and mitochondrial permeability in the hippocampus after cardiopulmonary resuscitation. J Cell Physiol. 2018;233(9):6902‐6910. 10.1002/jcp.26459 [DOI] [PubMed] [Google Scholar]
- 101. Hu YQ, Chen W, Yan MH, Lai JJ, Tang N, Wu L. Ischemic preconditioning protects brain from ischemia/reperfusion injury by attenuating endoplasmic reticulum stress‐induced apoptosis through PERK pathway. Eur Rev Med Pharmacol Sci. 2017;21(24):5736‐5744. 10.26355/eurrev_201712_14020 [DOI] [PubMed] [Google Scholar]
- 102. Cantarella G, Pignataro G, Di Benedetto G, et al. Ischemic tolerance modulates TRAIL expression and its receptors and generates a neuroprotected phenotype. Cell Death Dis. 2014;5:e1331. 10.1038/cddis.2014.286 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Ge Y, Chen W, Axerio‐Cilies P, Wang YT. NMDARs in cell survival and death: implications in stroke pathogenesis and treatment. Trends Mol Med. 2020;26(6):533‐551. 10.1016/j.molmed.2020.03.001 [DOI] [PubMed] [Google Scholar]
- 104. Zhang J, Yang J, Wang H, et al. The AAA + ATPase Thorase is neuroprotective against ischemic injury. J Cereb Blood Flow Metab. 2019;39(9):1836‐1848. 10.1177/0271678X18769770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Narayanan SV, Perez‐Pinzon MA. Ischemic preconditioning treatment of astrocytes transfers ischemic tolerance to neurons. Cond Med. 2017;1(1):2‐8. [PMC free article] [PubMed] [Google Scholar]
- 106. Gong J, Gong S, Zhang M, et al. Cerebral ischemic preconditioning reduces glutamate excitotoxicity by up‐regulating the uptake activity of GLT‐1 in rats. Amino Acids. 2014;46(6):1537‐1545. 10.1007/s00726-014-1723-1 [DOI] [PubMed] [Google Scholar]
- 107. Gong S‐J, Chen L‐Y, Zhang M, et al. Intermittent hypobaric hypoxia preconditioning induced brain ischemic tolerance by up‐regulating glial glutamate transporter‐1 in rats. Neurochem Res. 2012;37(3):527‐537. 10.1007/s11064-011-0639-3 [DOI] [PubMed] [Google Scholar]
- 108. Park H‐K, Chu K, Jung K‐H, et al. Autophagy is involved in the ischemic preconditioning. Neurosci Lett. 2009;451(1):16‐19. 10.1016/j.neulet.2008.12.019 [DOI] [PubMed] [Google Scholar]
- 109. Zhan L, Chen S, Li K, et al. Autophagosome maturation mediated by Rab7 contributes to neuroprotection of hypoxic preconditioning against global cerebral ischemia in rats. Cell Death Dis. 2017;8(7):e2949. 10.1038/cddis.2017.330 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110. Lu N, Li X, Tan R, et al. HIF‐1alpha/Beclin1‐mediated autophagy is involved in neuroprotection induced by hypoxic preconditioning. J Mol Neurosci. 2018;66(2):238‐250. 10.1007/s12031-018-1162-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Zhang N, Yin Y, Han S, et al. Hypoxic preconditioning induced neuroprotection against cerebral ischemic injuries and its cPKCgamma‐mediated molecular mechanism. Neurochem Int. 2011;58(6):684‐692. 10.1016/j.neuint.2011.02.007 [DOI] [PubMed] [Google Scholar]
- 112. Wei H, Li Y, Han S, et al. cPKCgamma‐modulated autophagy in neurons alleviates ischemic injury in brain of mice with ischemic stroke through Akt‐mTOR pathway. Transl Stroke Res. 2016;7(6):497‐511. 10.1007/s12975-016-0484-4 [DOI] [PubMed] [Google Scholar]
- 113. Sheng R, Zhang LS, Han R, Liu XQ, Gao B, Qin ZH. Autophagy activation is associated with neuroprotection in a rat model of focal cerebral ischemic preconditioning. Autophagy. 2010;6(4):482‐494. 10.4161/auto.6.4.11737 [DOI] [PubMed] [Google Scholar]
- 114. Gao BO, Zhang X‐Y, Han R, et al. The endoplasmic reticulum stress inhibitor salubrinal inhibits the activation of autophagy and neuroprotection induced by brain ischemic preconditioning. Acta Pharmacol Sin. 2013;34(5):657‐666. 10.1038/aps.2013.34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Neumann JT, Thompson JW, Raval AP, Cohan CH, Koronowski KB, Perez‐Pinzon MA. Increased BDNF protein expression after ischemic or PKC epsilon preconditioning promotes electrophysiologic changes that lead to neuroprotection. J Cereb Blood Flow Metab. 2015;35(1):121‐130. 10.1038/jcbfm.2014.185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. DeFazio RA, Raval AP, Lin HW, Dave KR, Della‐Morte D, Perez‐Pinzon MA. GABA synapses mediate neuroprotection after ischemic and epsilonPKC preconditioning in rat hippocampal slice cultures. J Cereb Blood Flow Metab. 2009;29(2):375‐384. 10.1038/jcbfm.2008.126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Secondo A, Petrozziello T, Tedeschi V, et al. ORAI1/STIM1 interaction intervenes in stroke and in neuroprotection induced by ischemic preconditioning through store‐operated calcium entry. Stroke. 2019;50(5):1240‐1249. 10.1161/STROKEAHA.118.024115 [DOI] [PubMed] [Google Scholar]
- 118. Chen W, Ju XZ, Lu Y, Ding XW, Miao CH, Chen JW. Propofol improved hypoxia‐impaired integrity of blood‐brain barrier via modulating the expression and phosphorylation of zonula occludens‐1. CNS Neurosci Ther. 2019;25(6):704‐713. 10.1111/cns.13101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Zhang H, Rzechorzek W, Aghajanian A, Faber JE. Hypoxia induces de novo formation of cerebral collaterals and lessens the severity of ischemic stroke. J Cereb Blood Flow Metab. 2020;40(9):1806‐1822. 10.1177/0271678X20924107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Gustavsson M, Mallard C, Vannucci SJ, Wilson MA, Johnston MV, Hagberg H. Vascular response to hypoxic preconditioning in the immature brain. J Cereb Blood Flow Metab. 2007;27(5):928‐938. 10.1038/sj.jcbfm.9600408 [DOI] [PubMed] [Google Scholar]
- 121. Wacker BK, Freie AB, Perfater JL, Gidday JM. Junctional protein regulation by sphingosine kinase 2 contributes to blood‐brain barrier protection in hypoxic preconditioning‐induced cerebral ischemic tolerance. J Cereb Blood Flow Metab. 2012;32(6):1014‐1023. 10.1038/jcbfm.2012.3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Wacker BK, Park TS, Gidday JM. Hypoxic preconditioning‐induced cerebral ischemic tolerance: role of microvascular sphingosine kinase 2. Stroke. 2009;40(10):3342‐3348. 10.1161/STROKEAHA.109.560714 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Lv MH, Li S, Jiang YJ, Zhang W. The Sphkl/SlP pathway regulates angiogenesis via NOS/NO synthesis following cerebral ischemia‐reperfusion. CNS Neurosci Ther. 2020;26(5):538‐548. 10.1111/cns.13275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Gesuete R, Orsini F, Zanier ER, et al. Glial cells drive preconditioning‐induced blood‐brain barrier protection. Stroke. 2011;42(5):1445‐1453. 10.1161/STROKEAHA.110.603266 [DOI] [PubMed] [Google Scholar]
- 125. Xu MY, Wang YF, Wei PJ, Gao YQ, Zhang WT. Hypoxic preconditioning improves long‐term functional outcomes after neonatal hypoxia‐ischemic injury by restoring white matter integrity and brain development. CNS Neurosci Ther. 2019;25(6):734‐747. 10.1111/cns.13102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Zeynalov E, Shah ZA, Li RC, Dore S. Heme oxygenase 1 is associated with ischemic preconditioning‐induced protection against brain ischemia. Neurobiol Dis. 2009;35(2):264‐269. 10.1016/j.nbd.2009.05.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Lee J‐C, Kim IH, Park JH, et al. Ischemic preconditioning protects hippocampal pyramidal neurons from transient ischemic injury via the attenuation of oxidative damage through upregulating heme oxygenase‐1. Free Radic Biol Med. 2015;79:78‐90. 10.1016/j.freeradbiomed.2014.11.022 [DOI] [PubMed] [Google Scholar]
- 128. Lin WY, Chang YC, Ho CJ, Huang CC. Ischemic preconditioning reduces neurovascular damage after hypoxia‐ischemia via the cellular inhibitor of apoptosis 1 in neonatal brain. Stroke. 2013;44(1):162‐169. 10.1161/STROKEAHA.112.677617 [DOI] [PubMed] [Google Scholar]
- 129. Lusardi TA, Farr CD, Faulkner CL, et al. Ischemic preconditioning regulates expression of microRNAs and a predicted target, MeCP2, in mouse cortex. J Cereb Blood Flow Metab. 2010;30(4):744‐756. 10.1038/jcbfm.2009.253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Liang D, He X‐B, Wang Z, et al. Remote limb ischemic postconditioning promotes motor function recovery in a rat model of ischemic stroke via the up‐regulation of endogenous tissue kallikrein. CNS Neurosci Ther. 2018;24(6):519‐527. 10.1111/cns.12813 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Lee BJ, Gibson OR, Thake CD, Tipton M, Hawley JA, Cotter JD. Editorial: cross adaptation and cross tolerance in human health and disease. Front Physiol. 2018;9:1827. 10.3389/fphys.2018.01827 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Chen LI, Huang K, Wang R, et al. Neuroprotective effects of cerebral ischemic preconditioning in a rat middle cerebral artery occlusion model: the role of the notch signaling pathway. Biomed Res Int. 2018;2018:8168720. 10.1155/2018/8168720 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Geng J, Zhang Y, Li S, et al. Metabolomic profiling reveals that reprogramming of cerebral glucose metabolism is involved in ischemic preconditioning‐induced neuroprotection in a rodent model of ischemic stroke. J Proteome Res. 2019;18(1):57‐68. 10.1021/acs.jproteome.8b00339 [DOI] [PubMed] [Google Scholar]
- 134. Sheng R, Liu X‐Q, Zhang L‐S, et al. Autophagy regulates endoplasmic reticulum stress in ischemic preconditioning. Autophagy. 2012;8(3):310‐325. 10.4161/auto.18673 [DOI] [PubMed] [Google Scholar]
- 135. Faraco G, Blasi F, Min W, Wang ZQ, Moroni F, Chiarugi A. Brain ischemic preconditioning does not require PARP‐1. Stroke. 2010;41(1):181‐183. 10.1161/STROKEAHA.109.567826 [DOI] [PubMed] [Google Scholar]
- 136. Lee TH, Yang JT, Lin JR, et al. Protective effects of ischemic preconditioning against neuronal apoptosis and dendritic injury in the hippocampus are age‐dependent. J Neurochem. 2020. 155(4):430‐447. 10.1111/jnc.15029 [DOI] [PubMed] [Google Scholar]
- 137. Yung LM, Wei Y, Qin T, Wang Y, Smith CD, Waeber C. Sphingosine kinase 2 mediates cerebral preconditioning and protects the mouse brain against ischemic injury. Stroke. 2012;43(1):199‐204. 10.1161/STROKEAHA.111.626911 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Guan R, Lv J, Xiao F, Tu Y, Xie Y, Li L. Potential role of the cAMP/PKA/CREB signalling pathway in hypoxic preconditioning and effect on propofolinduced neurotoxicity in the hippocampus of neonatal rats. Mol Med Rep. 2019;20(2):1837‐1845. 10.3892/mmr.2019.10397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Lv J, Liang Y, Tu Y, Chen J, Xie Y. Hypoxic preconditioning reduces propofol‐induced neuroapoptosis via regulation of Bcl‐2 and Bax and downregulation of activated caspase‐3 in the hippocampus of neonatal rats. Neurol Res. 2018;40(9):767‐773. 10.1080/01616412.2018.1477545 [DOI] [PubMed] [Google Scholar]
- 140. Zhan L, Lu Z, Zhu X, et al. Hypoxic preconditioning attenuates necroptotic neuronal death induced by global cerebral ischemia via Drp1‐dependent signaling pathway mediated by CaMKIIalpha inactivation in adult rats. FASEB J. 2019;33(1):1313‐1329. 10.1096/fj.201800111RR [DOI] [PubMed] [Google Scholar]
- 141. Yang J, Liu C, Du X, et al. Hypoxia inducible factor 1alpha plays a key role in remote ischemic preconditioning against stroke by modulating inflammatory responses in rats. J Am Heart Assoc. 2018;7(5):e007589. 10.1161/JAHA.117.007589 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Liu C, Zhang C, Du H, Geng X, Zhao H. Remote ischemic preconditioning protects against ischemic stroke in streptozotocin‐induced diabetic mice via anti‐inflammatory response and anti‐apoptosis. Brain Res. 2019;1724:146429. 10.1016/j.brainres.2019.146429 [DOI] [PubMed] [Google Scholar]
- 143. Zhao W, Meng R, Ma C, et al. Safety and efficacy of remote ischemic preconditioning in patients with severe carotid artery stenosis before carotid artery stenting: a proof‐of‐concept, randomized controlled trial. Circulation. 2017;135(14):1325‐1335. 10.1161/CIRCULATIONAHA.116.024807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. Zhou DA, Ding J, Ya J, et al. Efficacy of remote ischemic conditioning on improving WMHs and cognition in very elderly patients with intracranial atherosclerotic stenosis. Aging. 2019;11(2):634‐648. 10.18632/aging.101764 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Meng R, Asmaro K, Meng L, et al. Upper limb ischemic preconditioning prevents recurrent stroke in intracranial arterial stenosis. Neurology. 2012;79(18):1853‐1861. 10.1212/WNL.0b013e318271f76a [DOI] [PubMed] [Google Scholar]
- 146. Meng R, Ding Y, Asmaro K, et al. Ischemic conditioning is safe and effective for octo‐ and nonagenarians in stroke prevention and treatment. Neurotherapeutics. 2015;12(3):667‐677. 10.1007/s13311-015-0358-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Poalelungi A, Turiac E, Tulba D, Stoian D, Popescu BO. Remote ischemic conditioning in acute ischemic stroke ‐ a clinical trial design. J Med Life. 2020;13(2):156‐159. 10.25122/jml-2020-0049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Liao Z, Bu Y, Li M, et al. Remote ischemic conditioning improves cognition in patients with subcortical ischemic vascular dementia. BMC Neurol. 2019;19(1):206. 10.1186/s12883-019-1435-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Ding J‐Y, Shang S‐L, Sun Z‐S, et al. Remote ischemic conditioning for the treatment of ischemic moyamoya disease. CNS Neurosci Ther. 2020;26(5):549‐557. 10.1111/cns.13279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Mi T, Yu F, Ji X, Sun Y, Qu D. The interventional effect of remote ischemic preconditioning on cerebral small vessel disease: a pilot randomized clinical trial. Eur Neurol. 2016;76(1–2):28‐34. 10.1159/000447536 [DOI] [PubMed] [Google Scholar]
- 151. Sales AHA, Barz M, Bette S, et al. Impact of ischemic preconditioning on surgical treatment of brain tumors: a single‐center, randomized, double‐blind, controlled trial. BMC Med. 2017;15(1):137. 10.1186/s12916-017-0898-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Rytter N, Carter H, Piil P, et al. Ischemic preconditioning improves microvascular endothelial function in remote vasculature by enhanced prostacyclin production. J Am Heart Assoc. 2020;9(15): e016017. 10.1161/JAHA.120.016017 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.