

Abstract
Drug resistance of gastric carcinoma (GC) is a burning question in the medical field. This study aimed to investigating the ameliorative effect of apatinib (Apa) on paclitaxel (PTX) resistance in GC. In this research, PTX-resistant MGC803 cells were intervened by Apa. Cell proliferation was detected by cell counting kit-8 (CCK-8) assay, and cell migration and invasion was determined by Transwell assays. The levels of apoptosis-related proteins (Bcl-2, Bax), drug resistance-related proteins (MDR1, P-gp) and VEGFR2 protein were measure by Western blot, and the mRNA expression of VEGFR2 was tested by real-time quantitative polymerase chain reaction (RT-qPCR). Then VEGFR2 was overexpressed to examine the role of Apa in PTX-resistant MGC803 cells. The results identified a significantly reduced growth rate of MGC803/PTX cells after PTX induction, obviously increased invasive and migrated cells, and evidently enhanced proliferation capacity of MGC803/PTX cells as compared to MGC803 cells. In MGC803/PTX cells, VEGFR2, MDR1, P-gp and Bcl-2 were all up-regulated while Bax was down-regulated. After Apa intervention, PTX-resistant MGC803 cells showed decreased cell migration, invasion and proliferation, reduced MDR1, P-gp and VEGFR2 levels, and increased Bax protein level. Overexpression of VEGFR2 can offset the rescue effect of Apa on PTX-induced drug resistance of MGC803 cells. Taken together, Apa may inhibit PTX resistance of MGC803 cells via the VEGFR2 signaling pathway.
Keywords: Apatinib, paclitaxel, VEGFR2, gastric carcinoma, tumor resistance
Introduction
Gastric carcinoma (GC) is one of the common digestive system malignancies [1]. At present, the treatment of early GC mainly relies on surgical treatment combined with adjuvant treatments such as chemotherapy, radiotherapy and targeted therapy. Perioperative and postoperative chemotherapy and other adjuvant therapies can significantly improve the postoperative survival of patients [2,3]. Unfortunately, most patients with GC are in the advanced stage at their diagnosis, and the tumor has often invaded the serosa or even spread and underwent distant metastasis [4]. When there is metastasis or peripheral invasion, chemotherapy-based comprehensive treatment becomes the preferred choice [5].
Traditional chemotherapy drugs for GC include platinum [6], fluorouracil [7] and taxanes [8], among which taxanes mainly affects cell mitosis and inhibits cell growth [9]. These drugs are also the common first-line drugs for the treatment of GC. However, drug resistance has always been the bottleneck limiting their extensive clinical application. The research on the mechanism of paclitaxel (PTX) resistance has become a hotspot in clinical research. PTX was first discovered as a microtubule-specific drug for the treatment of tumors [10]. It has been reported that PTX can play an anti-tumor role by activating the PI3K/AKT pathway, mitochondrial pathway, EGFR pathway, and MEK-ERK signaling pathway, etc. The curative effect is better in early stage of tumors, but the overall response of PTX therapy decreased significantly in advanced and recurrent tumors [11,12]. Some studies have shown that PTX resistance is associated with tubulin gene mutation [13], high expression of multidrug resistance 1 (MDR1) gene [14], and apoptosis-related proteins Caspases and Bcl-2 family [15]. Apatinib (Apa) is a small molecule vascular endothelial growth factor receptor-tyrosine kinase inhibitor developed in China. Its mechanism of action includes highly selective competition for the ATP binding site of VEGFR2 in cells, blocking downstream signal transduction and inhibiting angiogenesis in tumor tissues [16]. Apa has anti-tumor activity against various tumor cells such as cholangiocarcinoma cells, GC cells, liver cancer cells, and non-small cell lung cancer cells [17-19], which can obviously inhibit tumor cell proliferation and induce their apoptosis. Apa has been reported to reduce P-glycolprotein (P-gp) in transplanted tumors [20], and can reduce the risk of death by nearly 40% in patients with advanced GC after clinically proven failure of second-line treatment [21]. The current clinical application of Apa is mainly through selective inhibition of VEGFR2-mediated budding angiogenesis to exert its anti-tumor effect. PTX is an effective drug to treat GC, and the PTX resistance is related to multidrug resistance protein 1 (MDRl). It has been reported that Apa can reverse ABCB1 and ABCG2-mediated multiple-drug resistance (MDR) by competitively inhibiting the transport function [22]. Although Apa, a tyrosine kinase inhibitor targeting vascular endothelial growth factor 2 (VEGFR-2), has been approved by the Food and Drug Administration (FDA) for the treatment of advanced GC; However, the mechanism of the reversal effect of Apa on PTX-resistance in GC cells has not been fully elucidated. Consequently, the innovation of this study is mainly to explore the mechanism of the reversal effect of Apa on PTX-resistance in GC cells, which we hope to provide insights into the reversal of PTX resistance in GC.
Methods
Cell cultivation
Human GC MGC803 cells purchased from the American Type Culture Collection (ATCC) were cultured in the complete medium composed of Roswell Park Memorial Institute 1640 (RPMI 1640, Gibco; Thermo Fisher Scientific, Inc.), 10% foetal bovine serum (FBS, Thermo Fisher Scientific, Inc.), 100 U/mL penicillin (Thermo Fisher Scientific, Inc.) and 100 U/mL streptomycin (Thermo Fisher Scientific, Inc.) in a sterile incubator. The cells were digested with 0.15% trypsin prepared by phosphate buffer saline (PBS) for passage twice a week. MGC803 cells in the logarithmic growth phase (LGP) were collected for experimental study.
Drug-resistant cell line construction
With MGC803 as parent cell line and PTX as the induction drug, drug-resistant cell line was established by drug gradient induction method. The half maximal inhibitory concentration (IC50) of PTX to MGC803 was determined by cell counting kit-8 (CCK-8) assay. First, MGC803 cells were inoculated in a Petri dish. When the cells adhered to the culture dish and the fusion degree was 60%-70%, they were treated with 20 nM PTX (about 10 times that of IC50). After 24 h, the drug-containing medium was removed, and the dead cells were removed by adding the drug-free medium and changing the solution every day, while the living cells were allowed to proliferate. After the cells resumed growth, they continued to be passaged until the cells were in good condition. The above procedure was repeated, and the induction of PTX concentration was gradually increased, for a total of 10 months. Finally, the drug-resistant cell line MGC803/PTX was obtained.
Cell transfection
MGC-803/PTX cells were inoculated in the wells of 6-well plates. The VEGFR2 sequences were designed and then amplified by RT-PCR using the following primers: F: 5’-CCGGCTCCCGAGTTCTGGGCATTTC-3’, R: 5’-CCAGGAGTTGGGGGTGTGGATGCTTC-3’. The sequences were then ligated with a pcDNA3.0 plasmid to construct a recombinant pcDNA3.0-VEGFR2 vector (Medicilon, Shanghai, China). The cells were randomized into blank control group (Control, normal culture without any treatment), negative control group (oe-NC, transfected with empty pcDNA3.0 vector) and VEGFR2 overexpression group (oe-VEGFR2, transfected with pcDNA3.0-VEGFR2) by Lipofectamine 2000 liposome (Thermo Fisher Scientific, Inc.) according to the manufacturer’s instructions.
CCK-8
Drug-resistant cell establishment: Cells in LGP were digested and spread on 96-well plates at 1×103 cells/well. Different concentrations of PTX were configured when cell adherence was observed, and the medium was sucked out. Then, 200 µL of drug-containing culture medium with different concentrations was added to wells, while drug-free culture medium was added to the blank control group and negative control group, for 48 h of incubation. Subsequently, into each well, 6 µL CCK-8 solution was added for 3 h of incubation at 37°C, followed by 10 s of slight oscillation with an oscillating bed. For cell proliferation test, cells were tested at the time of 0, 24, 48, 72, 96, and 120 h of culture. For the selection of Apa concentration, the concentrations ranged from 0 μM to 16 μM. To determine the effect of Apa on proliferation of drug-resistant cells overexpressing VEGFR2, the concentrations of Apa were selected to be 0 μM, 0.5 μM, 1.0 μM and 2.0 μM. Finally, a Model 550 Microplate reader (Bio-Rad Laboratories, Berkeley, CA, USA) was used to detect the absorbance of each well at 450 nm. Three parallel wells were set under each condition. Drug resistance index (RI = IC50 value of drug-resistant cells/IC50 value) of parent cells was calculated. The detection correction value, which was the comparison value between each well’s detection value and the detection value at 0 h, was connected to draw a cell growth curve for growth curve analysis. Cell inhibition rate (%) = [1- (the experimental well’s OD-the blank control group’s OD)/(the negative control well’s OD-the blank control group’s OD)] ×100%.
Plate clone formation experiment
MGC-803 parent cells, drug-resistant cells, drug-resistant cells overexpressing VEGFR2 and drug-resistant cells with different concentrations of Apa were inoculated into 6-well plates at 2×102/well and cultured in an incubator until the cell colonies were visible to the naked eye. Then, the cells were processed for 3 PBS rinses, 0.5 h of fixing with 4% paraformaldehyde, and 30 min of immersion in prepared Giemsa staining solution (500 μL). Then, the staining solution was washed away with running water. The number of cell clones (more than 50 cells were counted as one clone) was counted under the microscope (Olympus Life Science, Tokyo, Japan), and the clone formation rate was calculated as (number of clones/number of inoculated cells) ×100%.
Real-time quantitative polymerase chain reaction (RT-qPCR)
Nano-Drop2000 ultraviolet spectrophotometer (Keyu Xingye Technology, Beijing, China) was used to determine the concentration and purity of proteins isolated from cells. Then, inverse transcription of RNA into cDNA was conducted with the Takara reverse transcription kit (Invitrogen, Carlsbed, CA, USA) according to the kit instructions, and the synthesized cDNA was stored at -20°C for later use. The primers (Table 1) were designed and synthesized by GenePharma (Shanghai, China). qPCR reaction procedure (40 cycles): pre-denaturation: 95°C, 10 min; denaturation: 95°C, 30 s; annealing: 60°C, 30 s; extension: 72°C, 1 min, and terminal extension: 72°C, 5 min. Each sample was repeatedly determined for 3 times. The gene’s relative expression was normalized by 2-ΔΔCT (internal reference gene: GAPDH).
Table 1.
Primer sequences
| Gene name | Primer sequence (5’-3’) | |
|---|---|---|
| VEGRF2 | Forward | GGACTCTCTCTGCCTACCTCAC |
| Reverse | GGCTCTTTCGCTTACTGTTCTG | |
| GAPDH | Forward | AGAAGGCTGGGGCTCATTTG |
| Reverse | AGGGGCCATCCACAGTCTTC |
Western blots
MGC-803 and MGC-803/PA cells in LGP were collected, placed on ice and immersed in RIPA lysate (Beijing Solarbio Science & Technology Co., Ltd.) to extract the total protein of each group. Protein concentration was measured by bicinchoninic acid (BCA) method and leveled. After sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), the protein was moved to a polyvinylidene fluoride (PVDF) membrane, which was then cleaned with TRIS-buffered saline with 0.5% Tween 20 (TBST) and blocked with 5% milk. Then the PVDF membrane was slowly shaken on a plate shaking table and closed for 2 h. Thereafter, the diluted primary antibodies were added for overnight incubation at 4°C. After TBST rinsing, the PVDF membrane was immersed in the diluted secondary antibody to incubate at ambient temperature for 2 h, followed by another TBST rinsing. After luminescence, tabletting, developing, fixation, and gray scanning, the gray value of protein expression was calculated to analyze the relative expression intensity of the protein. The expression of protein to be measured = the gray value of the band to be measured divided by that of the β-actin band. Primary antibodies, including P-gp (dilution ratio: 1:1,000), MDR1 (dilution ratio: 1:1,000), Bax (dilution ratio: 1:1,000), Bcl-2 (dilution ratio: 1:1,000), VEGFR2 (dilution ratio: 1:1,000), and β-actin (dilution ratio: 1:1,000), as well as the second antibody goat anti-rabbit (HRP cross-linked, dilution ratio 1:2,000) were all offered by Cell Signaling Technology.
Transwell
Spread on the upper chamber of Transwell chamber, the diluted Matrigel matrix glue (50 μL) was left standing in an incubator at 37°C for 30 min. The transfected cells of each group were re-selected by serum-free medium. Then, 700 μL RPMI 1640 supplemented with 20% serum was added to the wells (lower chamber) of the 24-well plate. The chamber was put into the wells, with approximately 5000 cells into each cell, and the volume was increased to 200 μL with serum-free medium and medium with different drug concentrations. After 48 h of culture, the medium in the chamber was discarded, and the cells that did not invade into the upper inner membrane were wiped off with Q-tips and washed with PBS three times. Then the cells were subjected to 20 min of immobilization with 4% paraformaldehyde, PBS rinsing, and 20-25 min of dyeing (indoor temperature) with 0.1% crystal violet. The unstained cells were washed off with running water, dried, and placed under an inverted microscope (Olympus Life Science, Tokyo, Japan) to take pictures of different fields for cell counting. The same method was used to detect the cell migration capacity. The other steps were the same as the invasion assay except that the upper compartment of Transwell chamber was not coated with Matrigel matrix adhesive. The experiment was repeated 3 times and the mean value was obtained.
Statistical processing
SPSS20.0 was used for data analysis. The measurement data were expressed as mean ± SD. The intra-group and multi-group comparisons were conducted using independent samples T test and ANOVA followed by Tukey’s post-hoc test, respectively. GraphPad Prism8.0 was used for figure rendering. P<0.05 was regarded as statistically significant.
Results
Establishment of drug-resistant human GC cell line MGC803/PTX
The IC50 values of PTX against MGC803 and MGC803/PTX were detected by CCK-8 assay, and the RI was calculated. As shown in Figure 1A, the IC50 of MGC803/PTX increased from 2.87 nM of parent cells to 28.04 nM, indicating that MGC803/PTX cells developed drug resistance to PTX, with a RI of 9.7 (moderate drug resistance). Compared with the parent cells, the growth rate of drug-resistant cells slowed down significantly, and the growth slope decreased under the same condition (P<0.05), Figure 1B.
Figure 1.
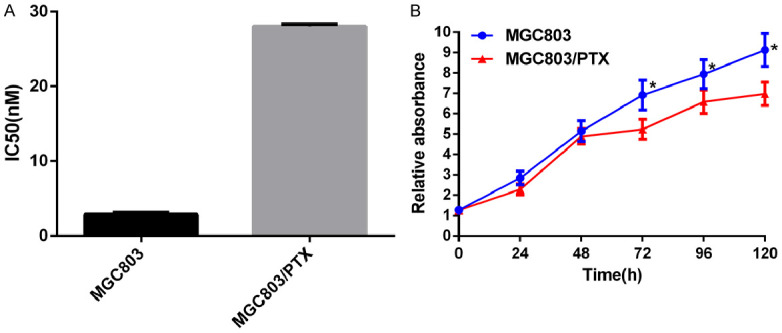
Establishment of drug-resistant human GC cell line MGC803/PTX. A: IC50 value and drug resistance index of PTX to MGC803 cells and MGC803/PTX cells; B: Growth curve of MGC80 parent cells and drug-resistant cells. *P<0.05 vs MGC803 cells.
Proliferation, invasion and migration of MGC803 and MGC803/PTX
The result of plate clone formation revealed significantly enhanced proliferation capacity of MGC803/PTX cells compared with MGC803 cells (P<0.05), as shown in Figure 2A. The results of Transwell assay determined evidently enhanced migration and invasion of MGC803 cells after PTX induction (P<0.05), as shown in Figure 2B.
Figure 2.
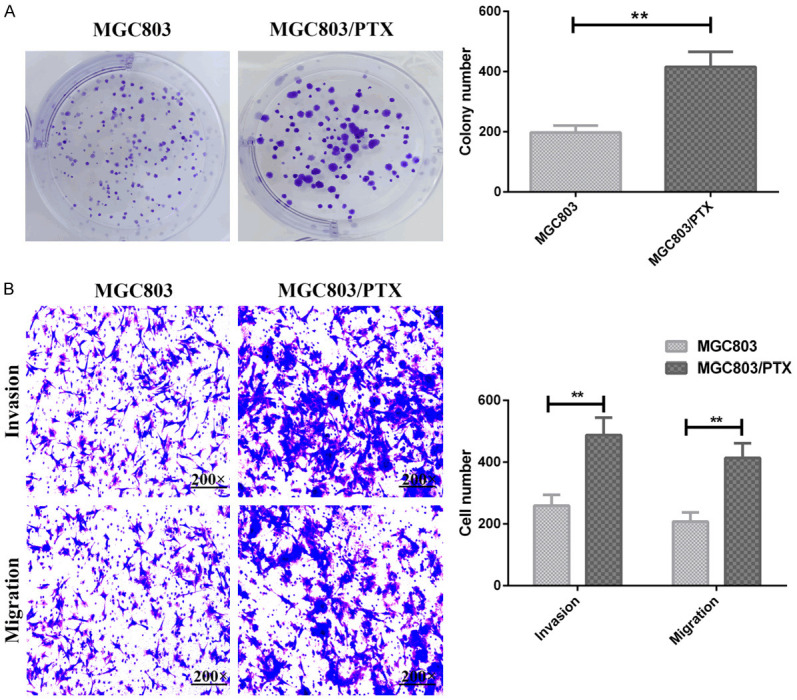
Proliferation, invasion and migration of MGC803 cells and MGC803/PTX cells. A: Number of clone formation of MGC803 cells and MGC803/PTX cells; B: Invasion and migration of MGC803 cells and MGC803/PTX cells; **P<0.01.
mRNA and protein expression of VEGFR2 in MGC803 and MGC803/PTX
MGC803/PTX cells presented significantly higher mRNA and protein expression of VEGFR2 than MGC803 cells (P<0.05), as shown in Figure 3.
Figure 3.
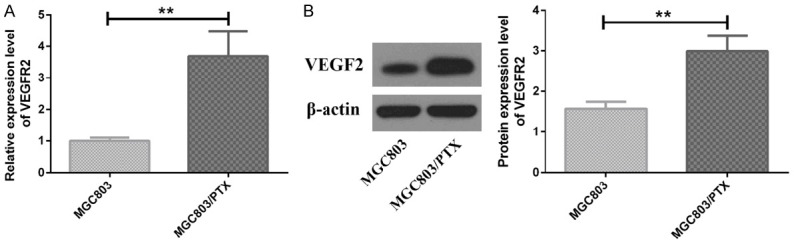
mRNA and protein expression of VEGFR2 in MGC 803 cells and MGC803/PTX cells. A: VEGFR2 mRNA expression; B: VEGFR2 protein expression; **P<0.01.
Expression of drug resistance- and apoptosis-related proteins in MGC803 and MGC803/PTX cells
Western Blotting identified that protein expression of MDR1, P-gp and Bcl-2 increased significantly while Bax protein decreased in MGC803/PTX cells, as compared with MGC803 cells, as shown in Figure 4.
Figure 4.
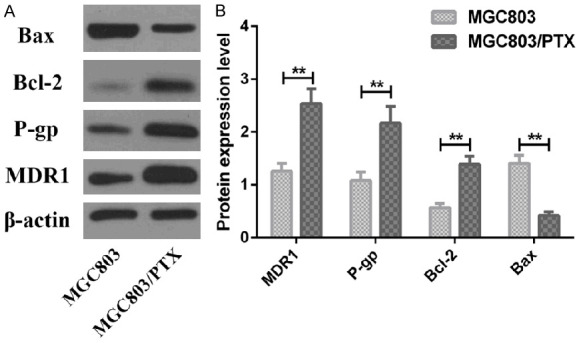
Expression of drug resistance- and apoptosis-related proteins in MGC 803 cells and MGC803/PTX cells. A: Western blot; B: Expression of related proteins; **P<0.01.
Selection of Apa concentration
In order to reduce the inherent cellular adverse reactions of drug resistance reversal agents in in vitro experiments, the concentration at which the inhibition rate of tumor cell proliferation does not exceed 10% is usually selected as the combined concentration of the drug resistance reversal agent. As shown in Figure 5, (0.1-16) μmol/L Apa inhibited the in vitro proliferation of MGC803 and MGC803/PTX in a concentration-dependent manner, and the inhibition rates of 0.5, 1.0, and 2 μmol/L Apa on MGC803/PTX cells were all less than 10%. Therefore, the three concentrations of Apa used to intervene MGC803/PTX were 0.5, 1.0, and 2.0 μmol/L, respectively.
Figure 5.
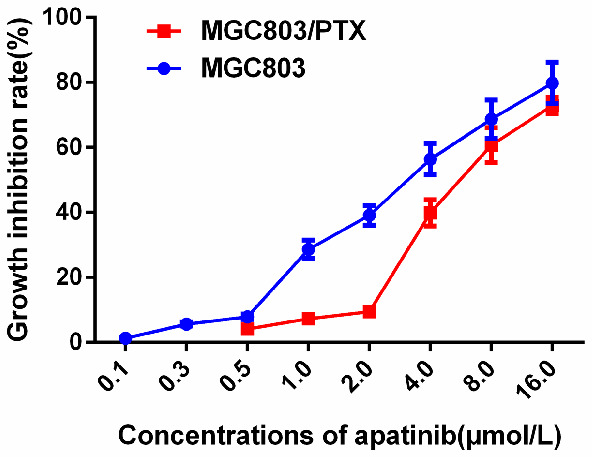
Selection of Apa concentration.
Effect of Apa on proliferation, invasion and migration of MGC803/PTX
After applying different concentrations of Apa to treat MGC803/PTX cells, the number of single cell clones, cell invasion and migration decreased statistically (all P<0.05), as shown in Figure 6.
Figure 6.
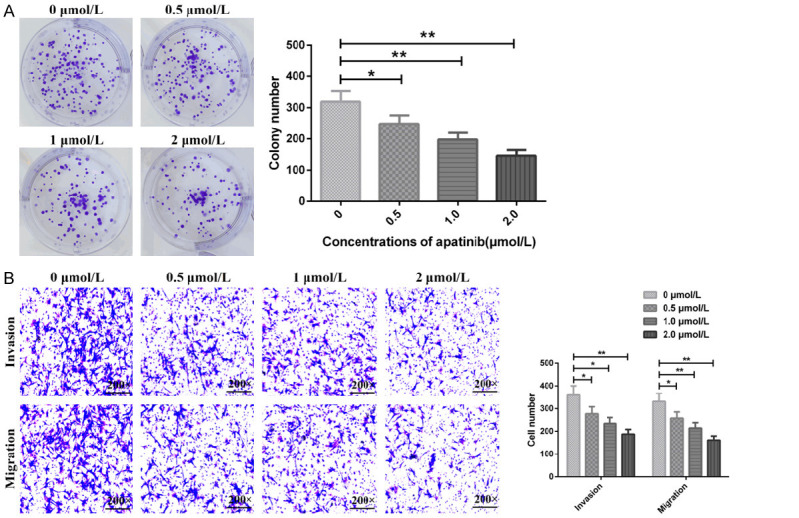
Effects of Apa on proliferation, invasion and migration of MGC803/PTX cells. A: Number of clone formation of MGC803/PTX cells; B: Number of invasion and migration of MGC803/PTX cells; *P<0.05, **P<0.01.
Effect of Apa on mRNA and protein expression of VEGFR2 in MGC803/PTX cells
As shown in Figure 7, mRNA and protein expression of VEGFR2 in MGC803/PTX cells decreased gradually as the Apa concentration increased; moreover, their levels were notably lower at the Apa concentration of 0.5, 1.0 and 2.0 μmol/L than that at the concentration of 0 μmol/L (all P<0.05).
Figure 7.

Effects of Apa on mRNA and protein expression of VEGFR2 in MGC803/PTX cells. A: VEGFR2 mRNA expression; B: VEGFR2 protein expression; *P<0.05, **P<0.01.
Effect of Apa on expression of drug resistance- and apoptosis-related proteins in MGC803/PTX cells
Western blotting identified that compared with the treatment with 0 μmol/L Apa, MDR1, P-gp and Bcl-2 protein elevated while Bax protein declined in MGC803/PTX cells as the Apa concentration increased; and compared with 0 μmol/L Apa, the differences in the above indexes were statistically significant when the Apa concentration was 0.5, 1.0, and 2.0 μmol/L respectively (all P<0.05), as shown in Figure 8.
Figure 8.
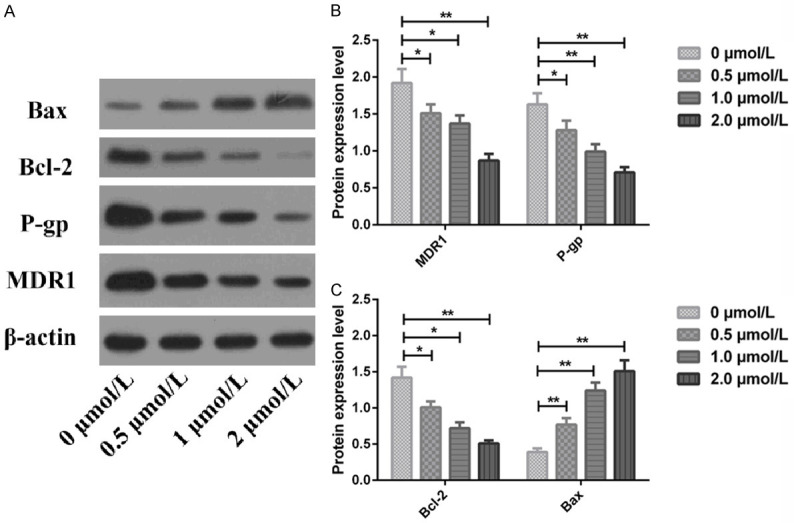
Effects of Apa on expression of drug resistance- and apoptosis-related proteins in MGC803/PTX cells. A: Western blots; B: MDR1 and P-pg protein expression; C: Bcl-2 and Bax protein expression; *P<0.05, **P<0.01.
Effects of Apa on MGC803/PTX cells overexpressing VEGFR2
The mRNA and protein expression of VEGFR2 in MGC803/PTX cells transfected with plasmid overexpressing VEGFR2 was significantly increased. After treatment with different concentrations of Apa, mRNA and protein expression of VEGFR2 declined obviously as the concentration of Apa increased (all P<0.05), as shown in Figure 9A, 9B. CCK-8 results showed that the proliferation rate of MGC803/PTX cells overexpressing VEGFR2 increased significantly but decreased markedly after Apa intervention (P<0.05), as shown in Figure 9C.
Figure 9.
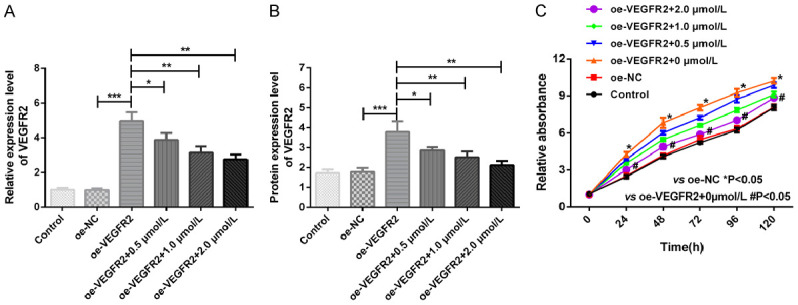
Effects of Apa on MGC803/PTX cells overexpressing VEGFR2. A: VEGFR2 mRNA expression; B: VEGFR2 protein expression; C: Growth curves of VEGFR2-overexpressing MGC803/PTX cells treated with different concentrations of Apa. *P<0.05, **P<0.01, ***P<0.001.
Discussion
MDR refers to the phenomenon that tumor cells develop drug resistance to anti-tumor drugs [23]. Once tumor cells acquire MDR, the effect of chemotherapy drugs is greatly compromised. So MDR is not only the major obstacle to the success of chemotherapy, but also a primary reason for cancer cell metastasis and recurrence [24]. This study focused on PTX resistance. There are many possible mechanisms of PTX resistance, such as microtubule protein mutation, microtubule binding protein, high expression of P-gp, and changes in Bcl-2 phosphorylation level [25-28]. In this study, the drug-resistant human GC cell line MGC803/PTX was successfully constructed using PTX as an inducer, and the results of the growth curve showed that the growth rate of the cells slowed down after drug resistance. This may be due to the long-term effects of the drug, which is a slow stress response of the cell in order to survive. In addition, in order to form clones, cells must be able to adhere to cell walls smoothly and be active in proliferation. The clone formation rate reflects two important characteristics: cell population dependence and proliferation ability. In this study, there was a significant difference in the clone formation rate between the two strains, indicating that the cell population dependence was reduced, and the ability of single cell adhesion and proliferation was enhanced after drug resistance. Therefore, the effect of Apa on the drug-resistant cells can be observed.
The results revealed that the migration, invasion and proliferation of PTX-resistant MGC803 cells decreased after treatment with Apa. Apa is a novel and effective inhibitor of vascular endothelial growth factor (VEGF) axis. Apa plays an anti-tumor role mainly through selective inhibition of VEGFR2-mediated budding angiogenesis, which has been verified in studies on arterial rings in rats [29]. In addition, we found that MDR1, P-gp and VEGFR2 decreased and Bax protein increased in PTX-resistant MGC803 cells intervened with Apa. Literature has shown that Apa can inhibit the P-gp transport function in drug-resistant solid tumors, thus reversing ABCB1 and ABCG2 mediated multi-resistance [30]. Zhang Q, et al. [31] found that Apa reversed PTX resistant lung cancer cells (A549) by blocking the function of ABCB1 transporter. P-gp is a drug pump protein encoded by MDR-1 gene, which belongs to the ABC transporter family. It is expressed in various tumors and normal tissues, and can bind to drugs to pump out many natural products such as taxanes, vinca alkaloids and anthracyclines [32]. P-gp is highly expressed in many drug-resistant cells, and the mechanism may be due to the amplification and mutation of MDR1 gene [33]. In PTX-resistant cell lines, MDR1 has been reported to predict resistance to PTX as high as 96% [34]. The Western blot results of this study also showed that the high expression of P-gp and MDR1 was the drug resistance mechanism of MGC803/PTX cells. Bei Y, et al. [33] found that Apa could weaken the resistance of GC cells to PTX by suppressing the JAK/STAT3 signaling pathway.
Angiogenesis is important for tumor growth and metastasis, and VEGF signal plays a vital role in angiogenesis [35]. VEGFR2 is the main receptor of VEGF-induced angiogenic signal transduction. Therefore, blocking the signal transduction of VEGFR2 pathway to inhibit angiogenesis and thus suppress tumor growth and metastasis may be a promising approach as a new target. Moreover, there is evidence that PTX shows natural anti-angiogenesis properties by directly affecting endothelial cell function and inhibiting VEGF induced angiogenesis both in vivo and in vitro [36]. In this research, we found that VEGFR2 overexpression can accelerate the proliferation of MGC803/PTX cells. Apa is one of the latest oral anti-angiogenic agents that can target VEGFR2 tyrosine kinase as a selective VEGFR2 inhibitor [37]. Then MGC803/PTX cells overexpressing VEGFR2 were treated with Apa, and it was found that Apa reversed the damage caused by overexpression of VEGFR2. Studies have shown that Apa inhibited the proliferation and migration of human umbilical vein endothelial cells by inhibiting the kinase activities of VEGFR2, c-kit and c-Src in vivo [38]. Our results also suggested that Apa could reverse the damage caused by overexpression of VEGFR2.
However, the present study still has limitation. This study is an in vitro experiment aiming to explore the possible mechanisms of Apa acting on chemotherapy drug resistance. Therefore, future randomized animal experimental and clinical studies with large sample size are needed to demonstrate the conclusions obtained in this study. We are looking forward to exploring whether Apa can successfully reverse drug resistance of GC in animals and human subjects through clinical trials.
In conclusion, Apa reverses PTX resistance in MGC803 cells through VEGFR2 pathway, which provides a possibility for improving the efficacy of chemotherapy and prolonging the survival of patients with GC.
Disclosure of conflict of interest
None.
References
- 1.Smyth EC, Nilsson M, Grabsch HI, van Grieken NC, Lordick F. Gastric cancer. Lancet. 2020;396:635–648. doi: 10.1016/S0140-6736(20)31288-5. [DOI] [PubMed] [Google Scholar]
- 2.Johnston FM, Beckman M. Updates on management of gastric cancer. Curr Oncol Rep. 2019;21:67. doi: 10.1007/s11912-019-0820-4. [DOI] [PubMed] [Google Scholar]
- 3.Machlowska J, Baj J, Sitarz M, Maciejewski R, Sitarz R. Gastric cancer: epidemiology, risk factors, classification, genomic characteristics and treatment strategies. Int J Mol Sci. 2020;21:4012. doi: 10.3390/ijms21114012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Tan Z. Recent advances in the surgical treatment of advanced gastric cancer: a review. Med Sci Monit. 2019;25:3537–3541. doi: 10.12659/MSM.916475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Reddavid R, Sofia S, Chiaro P, Colli F, Trapani R, Esposito L, Solej M, Degiuli M. Neoadjuvant chemotherapy for gastric cancer. Is it a must or a fake? World J Gastroenterol. 2018;24:274–289. doi: 10.3748/wjg.v24.i2.274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pan J, Xiang Z, Dai Q, Wang Z, Liu B, Li C. Prediction of platinum-resistance patients of gastric cancer using bioinformatics. J Cell Biochem. 2019;120:13478–13486. doi: 10.1002/jcb.28621. [DOI] [PubMed] [Google Scholar]
- 7.Cui XL, Li KJ, Ren HX, Zhang YJ, Liu XD, Bu BG, Wang L. Extract of cycas revoluta thunb. enhances the inhibitory effect of 5-fluorouracil on gastric cancer cells through the AKT-mTOR pathway. World J Gastroenterol. 2019;25:1854–1864. doi: 10.3748/wjg.v25.i15.1854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.De Vita F, Borg C, Farina G, Geva R, Carton I, Cuku H, Wei R, Muro K. Ramucirumab and paclitaxel in patients with gastric cancer and prior trastuzumab: subgroup analysis from RAINBOW study. Future Oncol. 2019;15:2723–2731. doi: 10.2217/fon-2019-0243. [DOI] [PubMed] [Google Scholar]
- 9.Weaver BA. How taxol/paclitaxel kills cancer cells. Mol Biol Cell. 2014;25:2677–2681. doi: 10.1091/mbc.E14-04-0916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Shi X, Sun X. Regulation of paclitaxel activity by microtubule-associated proteins in cancer chemotherapy. Cancer Chemother Pharmacol. 2017;80:909–917. doi: 10.1007/s00280-017-3398-2. [DOI] [PubMed] [Google Scholar]
- 11.Wang Y, Liu Y, Li G, Su Z, Ren S, Tan P, Zhang X, Qiu Y, Tian Y. Ephrin type-A receptor 2 regulates sensitivity to paclitaxel in nasopharyngeal carcinoma via the phosphoinositide 3-kinase/Akt signalling pathway. Mol Med Rep. 2015;11:924–930. doi: 10.3892/mmr.2014.2799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhang XY, Zhang YD. Enhanced antiproliferative and apoptosis effect of paclitaxel-loaded polymeric micelles against non-small cell lung cancers. Tumour Biol. 2015;36:4949–4959. doi: 10.1007/s13277-015-3142-7. [DOI] [PubMed] [Google Scholar]
- 13.Yin S, Zeng C, Hari M, Cabral F. Paclitaxel resistance by random mutagenesis of α-tubulin. Cytoskeleton (Hoboken) 2013;70:849–862. doi: 10.1002/cm.21154. [DOI] [PubMed] [Google Scholar]
- 14.Januchowski R, Wojtowicz K, Andrzejewska M, Zabel M. Expression of MDR1 and MDR3 gene products in paclitaxel-, doxorubicin-and vincristine-resistant cell lines. Biomed Pharmacother. 2014;68:111–117. doi: 10.1016/j.biopha.2013.09.004. [DOI] [PubMed] [Google Scholar]
- 15.Wang TH, Wang HS, Soong YK. Paclitaxel-induced cell death: where the cell cycle and apoptosis come together. Cancer. 2000;88:2619–2628. doi: 10.1002/1097-0142(20000601)88:11<2619::aid-cncr26>3.0.co;2-j. [DOI] [PubMed] [Google Scholar]
- 16.Liu K, Ren T, Huang Y, Sun K, Bao X, Wang S, Zheng B, Guo W. Apatinib promotes autophagy and apoptosis through VEGFR2/STAT3/BCL-2 signaling in osteosarcoma. Cell Death Dis. 2017;8:e3015. doi: 10.1038/cddis.2017.422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Peng H, Zhang Q, Li J, Zhang N, Hua Y, Xu L, Deng Y, Lai J, Peng Z, Peng B. Apatinib inhibits VEGF signaling and promotes apoptosis in intrahepatic cholangiocarcinoma. Oncotarget. 2016;7:17220–9. doi: 10.18632/oncotarget.7948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Scott LJ. Apatinib: a review in advanced gastric cancer and other advanced cancers. Drugs. 2018;78:747–758. doi: 10.1007/s40265-018-0903-9. [DOI] [PubMed] [Google Scholar]
- 19.Ni Y, Ye X. Apatinib for hepatocellular carcinoma. J Cancer Res Ther. 2019;15:741–742. doi: 10.4103/jcrt.JCRT_400_19. [DOI] [PubMed] [Google Scholar]
- 20.Hu X, Zhang J, Xu B, Jiang Z, Ragaz J, Tong Z, Zhang Q, Wang X, Feng J, Pang D. Multicenter phase II study of apatinib, a novel VEGFR inhibitor in heavily pretreated patients with metastatic triple-negative breast cancer. Int J Cancer. 2014;135:1961–1969. doi: 10.1002/ijc.28829. [DOI] [PubMed] [Google Scholar]
- 21.Wang B, Song L, Niu P. Clinical efficacy of apatinib in treatment of advanced gastric cancer. World Chin J Digestol. 2016;24:759–764. [Google Scholar]
- 22.Li F, Zhu T, Cao B, Wang J, Liang L. Apatinib enhances antitumour activity of EGFR-TKIs in non-small cell lung cancer with EGFR-TKI resistance. Eur J Cancer. 2017;84:184–192. doi: 10.1016/j.ejca.2017.07.037. [DOI] [PubMed] [Google Scholar]
- 23.Mi YJ, Liang YJ, Huang HB, Zhao HY, Wu CP, Wang F, Tao LY, Zhang CZ, Dai CL, Tiwari AK, Ma XX, To KK, Ambudkar SV, Chen ZS, Fu LW. Apatinib (YN968D1) reverses multidrug resistance by inhibiting the efflux function of multiple ATP-binding cassette transporters. Cancer Res. 2010;70:7981–7991. doi: 10.1158/0008-5472.CAN-10-0111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li YJ, Lei YH, Yao N, Wang CR, Hu N, Ye WC, Zhang DM, Chen ZS. Autophagy and multidrug resistance in cancer. Chin J Cancer. 2017;36:52. doi: 10.1186/s40880-017-0219-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bukowski K, Kciuk M, Kontek R. Mechanisms of multidrug resistance in cancer chemotherapy. Int J Mol Sci. 2020;21:3233. doi: 10.3390/ijms21093233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kavallaris M. Microtubules and resistance to tubulin-binding agents. Nat Rev Cancer. 2010;10:194–204. doi: 10.1038/nrc2803. [DOI] [PubMed] [Google Scholar]
- 27.Mooneyham A, Iizuka Y, Yang Q, Coombes C, McClellan M, Shridhar V, Emmings E, Shetty M, Chen L, Ai T, Meints J, Lee MK, Gardner M, Bazzaro M. UNC-45A is a novel microtubule-associated protein and regulator of paclitaxel sensitivity in ovarian cancer cells. Mol Cancer Res. 2019;17:370–383. doi: 10.1158/1541-7786.MCR-18-0670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Guo W, Dong W, Li M, Shen Y. Mitochondria P-glycoprotein confers paclitaxel resistance on ovarian cancer cells. Onco Targets Ther. 2019;12:3881–3891. doi: 10.2147/OTT.S193433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kwon CH, Park HJ, Choi Y, Won YJ, Lee SJ, Park DY. TWIST mediates resistance to paclitaxel by regulating Akt and Bcl-2 expression in gastric cancer cells. Tumor Biol. 2017;39:1010428317722070. doi: 10.1177/1010428317722070. [DOI] [PubMed] [Google Scholar]
- 30.De Vera AA, Gupta P, Lei Z, Liao D, Narayanan S, Teng Q, Reznik SE, Chen ZS. Immuno-oncology agent IPI-549 is a modulator of P-glycoprotein (P-gp, MDR1, ABCB1)-mediated multidrug resistance (MDR) in cancer: in vitro and in vivo. Cancer Lett. 2019;442:91–103. doi: 10.1016/j.canlet.2018.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhang Q, Song Y, Cheng X, Xu Z, Matthew OA, Wang J, Sun Z, Zhang X. Apatinib reverses paclitaxel-resistant lung cancer cells (A549) through blocking the function of ABCB1 transporter. Anticancer Res. 2019;39:5461–5471. doi: 10.21873/anticanres.13739. [DOI] [PubMed] [Google Scholar]
- 32.Robey RW, Pluchino KM, Hall MD, Fojo AT, Bates SE, Gottesman MM. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat Rev Cancer. 2018;18:452–464. doi: 10.1038/s41568-018-0005-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Washio I, Nakanishi T, Ishiguro N, Bister B, Tamai I. Effect of endogenous multidrug resistance 1 and P-glycoprotein expression on anticancer drug resistance in colon cancer cell lines. Biopharm Drug Dispos. 2019;40:32–43. doi: 10.1002/bdd.2167. [DOI] [PubMed] [Google Scholar]
- 34.Bei Y, Chen X, Xu Q, Lv J, Hu J, Yang S. Apatinib weakens resistance of gastric cancer cells to paclitaxel by suppressing JAK/STAT3 signaling pathway. Drug Dev Res. 2021 doi: 10.1002/ddr.21867. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 35.Carmeliet P. VEGF as a key mediator of angiogenesis in cancer. Oncology. 2005;69(Suppl 3):4–10. doi: 10.1159/000088478. [DOI] [PubMed] [Google Scholar]
- 36.Bocci G, Di Paolo A, Danesi R. The pharmacological bases of the antiangiogenic activity of paclitaxel. Angiogenesis. 2013;16:481–492. doi: 10.1007/s10456-013-9334-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Roviello G, Ravelli A, Fiaschi AI, Cappelletti MR, Gobbi A, Senti C, Zanotti L, Polom K, Reynolds AR, Fox SB, Generali D. Apatinib for the treatment of gastric cancer. Expert Rev Gastroenterol Hepatol. 2016;10:887–892. doi: 10.1080/17474124.2016.1209407. [DOI] [PubMed] [Google Scholar]
- 38.Tian S, Quan H, Xie C, Guo H, Lü F, Xu Y, Li J, Lou L. YN968D1 is a novel and selective inhibitor of vascular endothelial growth factor receptor-2 tyrosine kinase with potent activity in vitro and in vivo. Cancer Sci. 2011;102:1374–1380. doi: 10.1111/j.1349-7006.2011.01939.x. [DOI] [PMC free article] [PubMed] [Google Scholar]