

Abstract
Despite the attempts that have started since the 1960s, not even a single cell line of marine molluscs is available. Considering the vast contribution of marine bivalve aquaculture to the world economy, the prevailing viral threats, and the dismaying lack of advancements in molluscan virology, the requirement of a marine molluscan cell line is indispensable. This synthetic review discusses the obstacles in developing a marine molluscan cell line concerning the choice of species, the selection of tissue and decontamination, and cell culture media, with emphasis given on the current decade 2010–2020. Detailed accounts on the experiments on the virus cultivation in vitro and molluscan cell immortalization, with a brief note on the history and applications of the molluscan cell culture, are elucidated to give a holistic picture of the current status and future trends in molluscan cell line development.
Supplementary Information
The online version contains supplementary material available at 10.1007/s10616-022-00539-x.
Keywords: Molluscan cell culture, Culture medium, Decontamination, Cell line, Transformation, Transfection, Immortalization, Virus
Introduction
The molluscs, which comprise the invertebrates, represent the second-largest phylum of the animal kingdom, with more than 1,00,000 identified species (Halanych and Kocot 2017). As of 2018, aquaculture provides income to 20.53 million people. Global aquaculture production in 2018 was, overall, 82.1 million tonnes of food fish, of which 17.7 million tonnes was contributed by molluscs alone (FAO 2020). During the period from 2009 to 2014, 14% of the world-wide marine production came from bivalves, of which 89% was directly from aquaculture, i.e. mustering an annual of 20.6 billion US dollars (Wijsman et al. 2019). These statistics pinpoint the significance of sustainable production of bivalve mariculture globally. Despite this fact, bivalve mariculture is prone to frequent setbacks due to viral outbreaks that have resulted in mass mortalities since the early 1970s till date (Comps et al. 1976; Hine and Thorne 1997; Segarra et al. 2010; de Kantzow et al. 2017). Many viral families have been identified, primarily relying on ultramicroscopy. Only a few viruses, mainly belonging to the Herpesviridae family, have been studied in detail using molecular tools. To isolate, characterize and cultivate the viruses in vitro, and explore the host–pathogen interactions, the requirement of a suitable marine molluscan cell line is inevitable. The lack of the same is also the primary reason behind the lacunae in molluscan virology (Mialhe et al. 1988; Arzul et al. 2017).
Besides this, molluscan cell lines offer varied applications such as studies on ecotoxicology, and the effect of environmental stresses, transgenic studies, understanding of physiological processes, and innate immunity (reviewed by Rinkevich, 1999; Yoshino et al., 2013). While the attempts have started since the 1960s (Mialhe et al. 1988), not even a single cell line of marine molluscs could be developed. The only molluscan cell line available to date is the Bge cell line that had originated from the embryos of Biomphalaria glabrata (freshwater snail) through the untiring efforts of Dr. Eder L. Hansen (1976).
There could be various factors hindering the establishment of a marine molluscan cell line, and it is essential to solve each one of them to bring about a successful outcome. The crucial aspects to be considered are proper decontamination procedures, choice of tissue, and the formulation of an appropriate cell culture medium to optimize the growth and division of the molluscan cells in vitro. The appropriate intervention of gene transfer and cell hybridization techniques may also be relied upon to bring about fruitful endeavours.
History of molluscan cell culture
The attempts at initiating marine invertebrate cell culture had gained popularity in the 1970s, and over the decades, researchers focused primarily on six phyla—Porifera, Cnidaria, Crustacea, Mollusca, Echinodermata and Urochordata (Rinkevich 1999). The earliest attempt on molluscan organ culture was using the gills of the freshwater mussel, Anodonta sp. in which the vibratile epithelium survived for 63 days in diluted Ringer solution (Zweibaum 1925). The earliest attempt on marine molluscan organ culture was from the mantle of the oyster, Pinctada radiata (Bevelander and Martin 1949), which reported the deposition of conchiolin and subsequent formation of crystals in vitro. Despite the strenuous efforts over the decades (Brewster and Nicholson 1979; Stephens and Hetrick 1979; Hetrick et al. 1981; Wen et al. 1993a, b; Le Deuff et al. 1994; Renault et al. 1995; Chen and Wen 1999; Suja and Dharmaraj 2005; Suja et al. 2007, 2014; Daugavet and Blinova 2015; Maselli et al. 2018; Potts et al. 2020), no marine molluscan cell line could be developed till date. The earliest attempt on the cultivation of virus in vitro was recorded by Nakatsugawa et al. (1999), in which a virus of unknown aetiology was used to infect the primary culture of hemocytes from Japanese black abalones, Nordotis discus discus. A detailed account of the cultivation attempts of viruses in vitro is provided separately in this review (refer to “Attempts on virus-cultivation in vitro” section).
The Bge cell line was developed to study the host-parasite (B. glabrata—Schistosoma mansoni) interactions that were involved in the spreading of the infectious disease, schistosomiasis. It is a spontaneous cell line of adherent, fibroblastoid cells. The primary cell culture was obtained initially from trypsin-based cell dissociation, grown in the modified Schneider’s Drosophila medium and sub-cultured by gentle trypsinization (Hansen 1976). The cell line was characterized by immunoelectrophoresis, karyotyping, microscopic observations, and electrophoretic enzyme assays (Bayne et al. 1978). The cell line is difficult to propagate as the cells do not grow well as a monolayer or in suspension due to their tendency to form aggregates (Bayne 1989). Even though the cell line was originally diploid (n = 18), it has currently undergone extensive aneuploidy (Odoemelam et al. 2009). Both structural and sequential differences have been reported between the genome of Bge cells and the reference genome- B. glabrata BB02 (Wheeler et al. 2018).
The Bge cell line has been put to many application-oriented studies such as the demonstration of larval schistosome development within the snail host cells (Yoshino and Laursen 1995; Ivanchenko et al. 1999), genetic manipulation in vitro (Lardans et al. 1996; Laursen et al. 1997; Yoshino et al. 1998; Rinaldi et al. 2015) and as a model for understanding the biology of molluscs (Davids et al. 1999). A few notable attempts have been attained in the case of molluscan primary cell cultures. Cardiomyocyte and amoebocyte cultures, respectively developed from the heart of the oysters, Crassostrea gigas and Crassostrea virginica, and cardiomyocytes from the heart of clam, Meretrix lusoria could be maintained for durations ranging from 3 to 6 months (Brewster and Nicholson 1979; Domart-Coulon et al. 1994; Chen and Wen 1999), and larval cell cultures of C. virginica and Mercenaria mercenaria could be maintained up to 8 months (Ellis and Bishop 1989). Longer durations of cell culture have been reported from the heart and gonad of B. glabrata (Bayne et al. 1975), from the mantle of Pinctada margaritifera (Jayasankar et al. 2018), Mytilus edulis (Daugavet and Blinova 2015), and Haliotis varia (Suja and Dharmaraj 2005) (all of which survived for ≥ 1 year). The experiments on the transfection and immortalization of molluscan cells are dealt with separately in this review (refer to “Attempts on immortalization of molluscan cells in vitro” section).
Applications of molluscan cell culture
An overview of the applied studies of molluscan cell cultures is given here. Detailed reviews on the topic are available elsewhere (Rinkevich 1999; Yoshino et al. 2013; Ladhar-Chaabouni and Hamza-Chaffai 2016). The primary hemocyte culture has been used in studies concerning aquatic toxicology, and immunology (such as host–pathogen interactions) (Morga et al. 2011; Latire et al. 2012; Ji et al. 2017). The embryonic and larval cell cultures have been used in transgenic and physiological studies (Lardans et al. 1996; Yoshino et al. 1998; Boulo et al. 2000; Odintsova et al. 2010; Rinaldi et al. 2015; Boroda et al. 2019). The cell culture systems from the heart have been used in studies related to physiology, aquatic toxicology, and transgenesis (Cadoret et al. 1999; Domart-Coulon et al. 2000; Ödblom et al. 2000; Hanana et al. 2011; Droguet et al. 2012). The mantle cell culture has been used to study biomineralization processes, cell biology, and aquatic toxicology (Cornet 2007; Louzao et al. 2010; You et al. 2012b; Xiang et al. 2014; Jayasankar et al. 2018; Raghavan et al. 2019). The digestive gland and gill cell cultures have been subjected to studies such as aquatic cytotoxicity, genotoxicity, gene function analysis, and host–pathogen interactions (Mitchelmore et al. 1998; Domart-Coulon et al. 2000; Louzao et al. 2010; You et al. 2012a; Pichon et al. 2013). The neuronal cell culture has been used to demonstrate the physiology of nerve cells and factors affecting nerve function (Ridgway et al. 1991; Syed et al. 1992; Wildering et al. 1995; Leong et al. 2001). Gonad and labial palp cell cultures have been used for understanding molluscan physiology (Pazos and Mathieu 1999; Hanquet et al. 2011).
A decade review on the attempts on cell culture from marine molluscs
The various considerations to be undertaken to develop cell culture systems are discussed in the following sections, with particular attention given to the activities during the decade 2010–2020. In total, 48 publications during this decade (Table 1) have been selected based on the following criteria- adherence to the quality of the journals (based on SCOPUS indexed journals, SciMago Journal Rank indicator, and Clarivate Analytics-Journal Citation Reports-2019) and the omittance of unavailable manuscripts.
Table 1.
A decade review (2010–2020) on the attempts on cell culture from marine molluscs
| Class | Organism | Purpose of study | Remarks/conclusion | References |
|---|---|---|---|---|
| Bivalvia | Crassostrea gigas | Basic (cell isolation method) | A technique was developed for the isolation of spermatogonia and spermatocytes from the gonad | Franco et al. (2011) |
| Bivalvia | Crassostrea gigas | Applied (glucose uptake process) | The role of sodium glucose co-transporter (SGLT) in glucose uptake into vesicular cells was confirmed by the use of SGLT inhibitor, phlorizin | Hanquet et al. (2011) |
| Bivalvia | Crassostrea gigas | Applied (toxicology) | Tributyltin was found to be cytotoxic to the heart cells and affected cell viability, induced apoptosis, increased intracellular calcium level and decreased beating rhythm | Droguet et al. (2012) |
| Bivalvia | Crassostrea gigas | Applied (mitochondrial apoptosis) | Unlike in the vertebrates, APAF-1 protein and BH3-only subfamily proteins were not involved in the mitochondrial apoptosis pathway in oysters | Li et al. (2017) |
| Bivalvia | Crassostrea gigas | Applied (host-OsHV-1 pathology) | OsHV-1 could quickly initiate the replication in oyster hemocytes in vitro | Morga et al. (2017) |
| Bivalvia | Crassostrea gigas | Basic (cell culture development) | Cell culture initiated from the large explant method yielded cells with good viability | Potts et al. (2020) |
| Bivalvia | Crassostrea virginica | Applied (host-parasite pathology) | The initial infection by Perkinsus marinus induced caspase-independent apoptosis. The hemocytes were stimulated by the antigens on the parasite | Hughes et al. (2010) |
| Bivalvia | Crassostrea virginica | Applied (ocean acidification) | Elevated pCO2 resulted in the increased gene expression of four calcium-binding proteins involved in biomineralization | Richards et al. (2018) |
| Bivalvia | Crassostrea virginica | Applied (toxicology) | The phototoxicity assessment using dicloran and its intermediates showed decreased cell proliferation and decreased production of SCPB peptide | Xu et al. (2018) |
| Bivalvia | Pinctada fucata | Applied (biomineralization) | Cell-interactions involved in biomineralization were shown by way of histological and BrdU labeling methods | Awaji and Machii (2011) |
| Bivalvia | Pinctada fucata | Applied (biomineralization) | First report on the precipitation of amorphous calcium carbonate in vitro using primary mantle cell culture | Xiang et al. (2014) |
| Bivalvia | Pinctada margaritifera | Applied (biomineralization) | The expression of biomineralization gene nacrein, was obtained in mantle cell culture. Nacre was also secreted in vitro | Jayasankar et al. (2018) |
| Bivalvia | Mytilus galloprovincialis | Applied (toxicology) | A protocol for micronucleus cytome assay for the genotoxicity assessment in mussel was described | Bolognesi and Fenech (2012) |
| Bivalvia | Mytilus galloprovincialis | Applied (toxicology) | The toxicity of CdS quantum dots (used in anti-cancer therapy), bulk CdS and ionic Cd were reported in hemocyte and gill cell culture | Katsumiti et al. (2014) |
| Bivalvia | Mytilus galloprovincialis | Applied (toxicology) | BPA inhibited the lipid secretion of cells, and PFOS inhibited the lipid secretion and mitochondrial function | Balbi et al. (2017) |
| Bivalvia | Mytilus galloprovincialis | Applied (toxicology) | Okadaic acid was uptaken by the digestive gland cells through simple diffusion mechanism | Blanco et al. (2019) |
| Bivalvia | Mytilus edulis | Basic and Applied (cryopreservation; toxicology) | A protocol for the effective cryopreservation of cells was developed to be used for toxicological studies using the comet assay | Kwok et al. (2013) |
| Bivalvia | Mytilus edulis | Basic (functional characterization) | The motility and velocity of hemocytes were characterized by the use of videomicroscopy and cell tracking (using fluorescent probes) | Rioult et al. (2013) |
| Bivalvia | Mytilus edulis | Basic (cell culture development) | The cells remained viable for 22 months and could be cryopreserved using 5% DMSO with 50% cell survival post-thawing | Daugavet and Blinova (2015) |
| Bivalvia | Mytilus edulis | Basic (cell culture development) | The cells remained viable for 7 days, after which these were contaminated by mycoplasma and yeast. Three types of hemocytes were characterized | Barrick et al. (2018) |
| Bivalvia | Mytilus trossolus |
Basic and Applied (cell culture development; cell differentiation) |
Extracellular matrix was involved in the regulation of muscle proteins, leading to cell differentiation (myogenesis) in larval cell culture | Dyachuk (2013) |
| Bivalvia | Mytilus trossolus | Applied (cell adhesion) | β integrin-like protein was involved in cell-substrate adhesion and epithelial cell differentiation in larval primary cell culture | Maiorova and Odintsova (2015) |
| Bivalvia | Mytilus trossolus | Applied (apoptosis and necrosis) | Apoptosis occurred in 24% of the cryopreserved cells after thawing. The cells cryopreserved using 5% DMSO retained 60–65% viability post-thawing | Odintsova et al. (2017) |
| Bivalvia | Mytilus trossolus | Applied (apoptosis) | Apoptosis in the frozen-thawed cells was regulated by the use of apoptotic inducers or inhibitors, and staurosporine markedly increased apoptosis in the cells | Boroda et al. (2019) |
| Bivalvia | Perna viridis | Basic (cell culture development) | The cells showed the highest proliferative capacity in seawater medium. 10% FCS and 0.01% yeast extract could improve the cell size and proliferative capacity | Suja et al. (2017) |
| Bivalvia | Meretrix meretrix | Applied (gene function analysis) | First time report on the gene function analysis in bivalves using RNAi assay on ferritin gene | You et al. (2012a) |
| Bivalvia | Meretrix meretrix | Applied (apoptosis and cell adhesion) | The involvement of laminin receptor (MmeLR from M. meretrix) in cell apoptosis and adhesion was reported | You et al. (2012b) |
| Bivalvia | Ruditapes decussatus | Applied (toxicology) | The ionic currents in cardiomyocytes were identified. The pollutant, tributyltin, inhibited the potassium current, whereas okadaic acid did not | Hanana et al. (2011) |
| Bivalvia | Mesodesma mactroides | Applied (toxicology) | Copper inhibited carbonic anhydrase and Na+K+ATPase, competed with Na+ ions in ion transport, and affected anion regulation in mantle cells | Lopes et al. (2011) |
| Bivalvia | Mercenaria mercenaria | Applied (toxicology) | Acute hypercapnia had effects on the accumulation of trace metal pollutants (cadmium and copper) in mantle cells | Ivanina et al. (2013) |
| Bivalvia | Paphia malabarica | Basic (cryopreservation) | The cells cryopreserved with 15% glycerol retained enzyme activities (involved in biomineralization), besides showing good cell viability post thawing | Dessai (2018) |
| Bivalvia | Chlamys farreri | Applied (immunology) | Norepinephrine was involved in the regulation of nitric oxide production at different stages of immune response; nitric oxide exerted feedback regulation on the level of norepinephrine | Jiang et al. (2014) |
| Bivalvia | Chlamys farreri | Applied (toxicology) | Benzo(a)pyrene (BaP) at 0.8 µg L−1 caused significant DNA damage in digestive gland cells. GST and SOD were involved in the reduction of the BaP-induced toxicity | Cai et al. (2016) |
| Bivalvia | Chlamys farreri | Applied (host-OsHV-1/vibrio pathology) | The expression of immune response gene QM was significantly higher post-infection with Vibrio anguillarum and OsHV-1 in the hemocytes in vitro | Ji et al. (2017) |
| Bivalvia | Argopecten purpuratus | Applied (immunology) | Reactive oxygen intermediates acted as signaling molecules for the transcription of the antimicrobial peptide gene ApBD1 during an immune response | Oyanedel et al. (2016) |
| Gastropoda | Haliotis tuberculata | Applied (toxicology) | The pollutant, triclosan was found to be cytotoxic to hemocyte and gill cell cultures (IC50 6 µM) | Gaume et al. (2012) |
| Gastropoda | Haliotis tuberculata | Applied (biomineralization) | The expression of genes that were involved in biomineralization could be obtained in mantle cell culture but not in hemocyte culture | O’Neill et al. (2013) |
| Gastropoda | Haliotis tuberculata | Applied (host-vibrio pathology) | The cell interactions between host and vibrio were characterized by microscopic techniques (fluorescence and TEM) | Pichon et al. (2013) |
| Gastropoda | Haliotis tuberculata | Applied (toxicology) | Cadmium exposure resulted in changed morphology in hemocytes, but no significant toxicity was reported | Ladhar-Chaabouni et al. (2015) |
| Gastropoda | Haliotis discus hannai | Basic (cell culture development) | Growth factors and media salinity were crucial for the development of cell culture from radula | Kim et al. (2014) |
| Gastropoda | Haliotis discus hannai | Basic (cell culture development) | FBS and hemolymph were essential for the growth of ovary cells in vitro | Ryu et al. (2015) |
| Gastropoda | Haliotis asinina | Basic (cell culture development) | First report on the cell culture from H. asinina using gills and heart tissue | Suwattana et al. (2010) |
| Gastropoda | Haliotis midae | Basic (cell culture development) | Hemocyte culture was more favourable than larval cell culture due to lesser contamination | van der Merwe et al. (2010) |
| Gastropoda | Haliotis varia | Basic (depuration and decontamination of tissue) | More cell proliferation and lesser contamination in cell cultures from non-depurated abalones (than those from depurated ones), with tissues being treated with penicillin, streptomycin and fungizone prior to culture | Suja et al. (2014) |
| Gastropoda | Haliotis diversicolor | Basic (cell culture development) | First report on the role of IGFBP7 protein in the proliferation of hemocytes in vitro | Wang et al. (2015) |
| Gastropoda | Aplysia californica | Applied (neurophysiology) | The role of different PKMs in retaining persistent synaptic plasticity (that corresponds to long term memory) was identified | Hu et al. (2017) |
| Cephalopoda | Octopus vulgaris | Basic and Applied (cell characterization; antibacterial activity of hemocytes) | The hemocytes were morphologically and functionally characterized. The hemocytes possessed anti-bacterial activity against different strains | Troncone et al. (2015) |
| Cephalopoda | Octopus vulgaris | Basic (cell culture development) | The neuronal cell culture showed good survival and regenerative capacity when cultured in L-15 medium with FBS on poly-D-lysine coated substrate, following papain-based cell dissociation | Maselli et al. (2018) |
Of these 48 publications, 29% were purely basic studies on marine molluscan cell culture, 65% were purely applied studies, and 6% had basic as well as applied concepts in the studies undertaken. There is a rising trend towards applied studies using marine molluscan primary cell cultures and a lesser focus on developing cell lines from these.
Selection of suitable species
It has been reiterated by several authors that the choice of species has been limited over all these years (Rinkevich 2005; Yoshino et al. 2013). The criteria for the selection of species have been related to the following attributes: success previously attained in primary cell cultures (Rinkevich 2005), the commercial importance of the species (such as oysters, mussels, clams, and abalones), medical significance (such as intermediate hosts (e.g., snail) of human parasites) (Yoshino et al. 2013), pathological importance (such as hosts (e.g., oysters and abalones) of the pathogens- Bonamia sp., Perkinsus sp. and Vibrio sp.) (Alavi et al. 2009; Morga et al. 2009, 2011; Pichon et al. 2013; Ji et al. 2017), role as model organisms (such as gastropods used to study neurobiology) (Yoshino et al. 2013) and as bioindicators (such as oysters, mussels, clams, and abalones) (Domart-Coulon et al. 2000; Droguet et al. 2012; Latire et al. 2012; Ladhar-Chaabouni and Hamza-Chaffai 2016).
Our review of the 48 selected publications over the decade 2010–2020, listed in Table 1, indicated that the bivalves were the most-studied group (72.91%), followed by gastropods (22.92%). Among the bivalves, mussels (27.08%) were the most preferred group, followed by oysters (25%) (Fig. 1/Online resource Table 1). A detailed analysis (Table 2) revealed that all the species used for cell culture attempts during the decade were commercial (68.75%), bioindicators (29.17%), or model organisms (2.08%), representing a similar trend as those of past attempts.
Fig. 1.
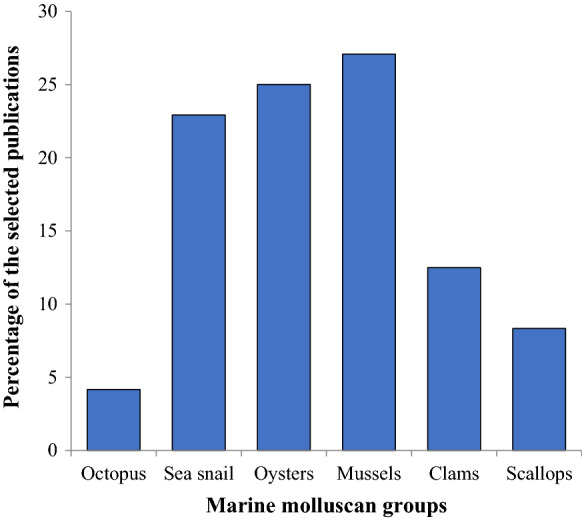
Marine molluscan groups used for cell culture development (In % of the selected publications 2010–2020)
Table 2.
The choice of marine molluscan species used for cell culture development and their status (commercial, bioindicator or model organism) (In % of the selected publications)
| Class | Category | Species | Status of species chosen for the studies | Total number of studies | References | |||
|---|---|---|---|---|---|---|---|---|
| Commercial | Bioindicator | Model organism | ||||||
| Pearl producer | Edible | |||||||
| Bivalvia | Oyster | Pinctada fucata | 2 | 12 | Hughes et al. (2010), Awaji and Machii (2011), Franco et al. (2011), Hanquet et al. (2011), Droguet et al. (2012), Xiang et al. (2014), Li et al. (2017), Morga et al. (2017), Xu et al. (2018), Jayasankar et al. (2018), Richards et al. (2018) and Potts et al. (2020) | |||
| Pinctada margaritifera | 1 | |||||||
| Crassostrea gigas | 5 | 1 | ||||||
| Crassostrea virginica | 1 | 2 | ||||||
| Mussel | Mytilus edulis | 3 | 1 | 13 | Bolognesi and Fenech (2012), Rioult et al. (2013), Dyachuk (2013), Kwok et al. (2013), Katsumiti et al. (2014), Daugavet and Blinova (2015), Maiorova and Odintsova (2015), Balbi et al. (2017), Odintsova et al. (2017); Suja et al. (2017), Barrick et al. (2018), Blanco et al. (2019) and Boroda et al. (2019) | |||
| Mytilus galloprovincialis | 4 | |||||||
| Mytilus trossolus | 4 | |||||||
| Perna viridis | 1 | |||||||
| Clam | Paphia malabarica | 1 | 6 | Hanana et al. (2011), Lopes et al. (2011), You et al. (2012b, a), Ivanina et al. (2013) and Dessai (2018) | ||||
| Ruditapes decussatus | 1 | |||||||
| Mercenaria mercenaria | 1 | |||||||
| Mesodesma mactroides | 1 | |||||||
| Meretrix meretrix | 2 | |||||||
| Scallop | Chlamys farreri | 2 | 1 | 4 | Jiang et al. (2014), Cai et al. (2016), Oyanedel et al. (2016) and Ji et al. (2017) | |||
| Argopecten purpuratus | 1 | |||||||
| Gastropoda | Sea snail | Haliotis tuberculata | 2 | 2 | 11 | Suwattana et al. (2010); van der Merwe et al. (2010), Gaume et al. (2012); O’Neill et al. (2013), Pichon et al. (2013), Kim et al. (2014), Suja et al. (2014), Wang et al. (2015), Ladhar-Chaabouni et al. (2015), Ryu et al. (2015) and Hu et al. (2017) | ||
| Haliotis discus hannai | 2 | |||||||
| Haliotis midae | 1 | |||||||
| Haliotis asinina | 1 | |||||||
| Haliotis diversicolor | 1 | |||||||
| Haliotis varia | 1 | |||||||
| Aplysia californica | 1 | |||||||
| Cephalopoda | Octopus | Octopus vulgaris | 2 | 2 | Troncone et al. (2015) and Maselli et al. (2018) | |||
| Total number of studies | 33 | 14 | 1 | 48 | ||||
| Percentage (%) | 68.75 | 29.17 | 2.08 | 100 | ||||
The species, for which physiological data are already available, offer an advantage in that the optimal osmolality, growth factors, and other components are known and could be applied in developing cell cultures (Bayne 1998). The advantage over the limited choice of species is that cell culture techniques would improve as the number of attempts on the same group of animals increase. Since researchers primarily focus on species that are of some importance, there will be a considerable advancement in the studies if cell lines of these species become available. The disadvantage is that there are chances of missing a suitable candidate species for molluscan cell line development when the choice is limited to a few.
Choice of organ/tissue and the life-stage
The choice of tissue for primary cell culture is usually influenced by the degree of obtaining aseptic cultures and the type of research question to be addressed (refer to “Applications of molluscan cell culture” section) (Mulcahy 2000). Stem cells are good candidates for cell line development. Fang’s research group (2008) had identified a proliferation hot spot of stem-like cells in the outer epithelia of the mantle central zone of the oyster, P. fucata. Fast-proliferating cells have been reported in hemocytes as well as epithelial cells of gills and digestive gland of the hard clam, M. mercenaria (Hanselmann et al. 2000), and in the digestive gland cells of the mussel, Mytilus galloprovincialis (Marigómez et al. 1999). Neoplastic cells are also favoured due to the ability to proliferate fast. Neoplastic cells with a high mitotic index (MI) of 3–12% were found in Mytilus trossolus and Mizuhopecten yessoensis (Odintsova et al. 2011). Cancerous clam hemocytes were successfully cultured from the clam Mya arenaria and maintained for three months (Walker et al. 2009). Neoplasia could be induced in M. arenaria using 5-bromodeoxyuridine (5-BrdU), and neoplastic hemocytes were isolated and cultured in vitro for four days (Oprandy and Chang 1983). Hemocytes are usually preferred for short-term studies as these are terminally differentiated and exhibit short longevity in vitro (2–3 days; maximum up to 2 weeks). Contamination-free cultures can be established easily since hemolymph that lies within the hemocoel tend to be aseptic. The cells from mantle tissue exhibit good proliferation and can be maintained long-term in vitro (~ 2–5 weeks) (reviewed by Yoshino et al., 2013).
The attempts on the genetic transformation in molluscan cells in vitro have been restricted to those derived from the heart and embryos/larvae. The heart, being enclosed separately in a pericardial sac, tend to be aseptic. It is easy to obtain axenic cell cultures from the embryos and larvae developed in vitro. Also, the embryonic/larval cells are noted for high proliferative potentials and the heart cells for retaining high viabilities. The heart and embryonic/larval cells have been maintained for extended periods (2–6 months in oysters; and 5 months in clam; 3–4 months in larvae) (Brewster and Nicholson 1979; Odintsova and Khomenko 1991; Domart-Coulon et al. 1994; Naganuma et al. 1994; Chen and Wen 1999).
While it is challenging to obtain axenic cell cultures from gills and digestive glands, a few have reported successful attempts on the short-term cultures of highly viable cells (Robledo and Cajaraville 1997; Faucet et al. 2004; Gómez-Mendikute et al. 2005).
It is to be noted that adult organisms mostly possess differentiated cells with limited capacity for proliferation. While it may be possible to develop a healthy primary cell culture system from adult cells, the chances are that cell proliferation will be limited. In this aspect, larval/embryonic cells outweigh the adult cells (except adult stem cells and neoplastic cells) as potential sources for continuous cell lines.
The analysis of the 48 publications (60 experiments) showed that the hemocytes were the most preferred for studies in cell culture (18 numbers; of which 11 were purely applied studies Hughes et al. 2010; Bolognesi and Fenech 2012; Gaume et al. 2012; O’Neill et al. 2013; Jiang et al. 2014; Katsumiti et al. 2014; Ladhar-Chaabouni et al. 2015; Oyanedel et al. 2016; Ji et al. 2017; Li et al. 2017; Morga et al. 2017), 5 were basic studies (van der Merwe et al. 2010; Rioult et al. 2013; Wang et al. 2015; Barrick et al. 2018; Potts et al. 2020), and 2 were a mix of both (Kwok et al. 2013; Troncone et al. 2015)). This was followed by the mantle tissue (13 studies; of which 8 were purely applied Awaji and Machii 2011; Lopes et al. 2011; You et al. 2012b; Ivanina et al. 2013; O’Neill et al. 2013; Xiang et al. 2014; Jayasankar et al. 2018; Richards et al. 2018), and 5 were purely basic (Suja et al. 2014, 2017; Daugavet and Blinova 2015; Dessai 2018; Potts et al. 2020)). Both applied studies and basic studies favoured the use of hemocytes, followed by the mantle tissue (Fig. 2/Online resource Table 2).
Fig. 2.

Tissues used for marine molluscan cell culture development and type of study. M mantle, L larva, DG digestive gland, HC hemocytes, H heart, GD gonad, G gills, N neuron, LP labial palps, R radula, AM adductor muscle (results from 60 experiments)
64% of the experiments dealt with adult animals (van der Merwe et al. 2010; Lopes et al. 2011; Franco et al. 2011; Hanana et al. 2011; Hanquet et al. 2011; Bolognesi and Fenech 2012; You et al. 2012a; Droguet et al. 2012; Gaume et al. 2012; Kwok et al. 2013; O’Neill et al. 2013; Pichon et al. 2013; Rioult et al. 2013; Kim et al. 2014; Suja et al. 2014, 2017; Ladhar-Chaabouni et al. 2015; Daugavet and Blinova 2015; Ryu et al. 2015; Wang et al. 2015; Cai et al. 2016; Oyanedel et al. 2016; Ji et al. 2017; Li et al. 2017; Hu et al. 2017; Jayasankar et al. 2018; Maselli et al. 2018; Richards et al. 2018; Dessai 2018; Potts et al. 2020), 14% with juveniles (Suwattana et al. 2010; Katsumiti et al. 2014; Xiang et al. 2014; Troncone et al. 2015; Balbi et al. 2017; Hu et al. 2017; Morga et al. 2017), 9% with larvae (van der Merwe et al. 2010; Dyachuk 2013; Maiorova and Odintsova 2015; Odintsova et al. 2017; Boroda et al. 2019) and the rest 13% (Hughes et al. 2010; Awaji and Machii 2011; You et al. 2012b; Ivanina et al. 2013; Jiang et al. 2014; Barrick et al. 2018; Xu et al. 2018; Blanco et al. 2019) used animals for which the life-stage was not specified. The preference for adult animals was evident during the decade because a larger quantity of hemolymph (hemocytes, being the most widely used source of cells during this period) could be aspirated from adult animals. Hemocytes, being short-lived and differentiated cells, have been used in short-term studies during this decade, as detailed out in the following section (refer to “Subculture and longevity” section).
Subculture and longevity
The cell culture systems derived from molluscs that could be maintained for relatively long durations have been already dealt with in “History of molluscan cell culture” section. In 2010–2020, 43 out of 60 cell-culture experiments did not proceed to sub-cultures as these studies were of short-term duration that focused on applications (Figs. 3 and 4/Online resource Tables 3 and 4) (Hughes et al. 2010; Suwattana et al. 2010; van der Merwe et al. 2010; Awaji and Machii 2011; Hanana et al. 2011; Hanquet et al. 2011; Lopes et al. 2011; Franco et al. 2011; Bolognesi and Fenech 2012; Droguet et al. 2012; Gaume et al. 2012; Ivanina et al. 2013; Kwok et al. 2013; O’Neill et al. 2013; Rioult et al. 2013; Jiang et al. 2014; Katsumiti et al. 2014; Kim et al. 2014; Suja et al. 2014, 2017; Ladhar-Chaabouni et al. 2015; Maiorova and Odintsova 2015; Troncone et al. 2015; Wang et al. 2015; Oyanedel et al. 2016; Cai et al. 2016; Balbi et al. 2017; Hu et al. 2017; Ji et al. 2017; Li et al. 2017; Morga et al. 2017; Odintsova et al. 2017; Barrick et al. 2018; Maselli et al. 2018; Richards et al. 2018; Blanco et al. 2019; Boroda et al. 2019). The maximum number of subcultures have been conducted with cells originating from the gonadal tissue (5 times) (Ryu et al. 2015). Mantle cell culture has been subcultured four times (Jayasankar et al. 2018), three times (Dessai 2018), and two times (Daugavet and Blinova 2015) (Fig. 3/Online resource Table 3).
Fig. 3.

Experiments indicating number of subcultures performed for each type of tissue. X-axis (numbers 0, 1, 2, 3, 4, 5) represent the number of subcultures (results from 60 experiments)
Fig. 4.

Experiments indicating longevity of cell culture derived from each type of tissue. M mantle, L larva, DG digestive gland, HC hemocytes, H heart, GD gonad, G gills, N neuron, LP labial palps, R radula, AM adductor muscle (results from 60 experiments)
With regard to the longevity of cell cultures, 6 experiments were cultured for a duration of less than 1 day (Franco et al. 2011; Hanquet et al. 2011; Lopes et al. 2011; Bolognesi and Fenech 2012; Blanco et al. 2019), 13 experiments for 1–2 days (Gaume et al. 2012; Ivanina et al. 2013; Katsumiti et al. 2014; Ladhar-Chaabouni et al. 2015; Cai et al. 2016; Oyanedel et al. 2016; Balbi et al. 2017; Li et al. 2017; Morga et al. 2017; Richards et al. 2018; Boroda et al. 2019), 5 experiments for 3–4 days (Hughes et al. 2010; Rioult et al. 2013; Wang et al. 2015; Maselli et al. 2018), 17 experiments for 1–2 weeks (van der Merwe et al. 2010; Awaji and Machii 2011; You et al. 2012b, a; Droguet et al. 2012; O’Neill et al. 2013; Jiang et al. 2014; Kim et al. 2014; Maiorova and Odintsova 2015; Hu et al. 2017; Suja et al. 2017; Barrick et al. 2018; Xu et al. 2018; Potts et al. 2020), 12 experiments for 1–3 months (2 cell cultures derived from mantle (Suja et al. 2014; Xiang et al. 2014), 1 from larva (Dyachuk 2013), 2 from hemocytes (Troncone et al. 2015; Ji et al. 2017), 3 from heart (Suwattana et al. 2010; Hanana et al. 2011; Potts et al. 2020), 1 from gonad (Ryu et al. 2015), and 2 from gills (Suwattana et al. 2010; Potts et al. 2020)), 2 experiments for more than a year (both of which were derived from the mantle tissue) (Daugavet and Blinova 2015; Jayasankar et al. 2018), and 5 experiments for which the longevity of the primary cell cultures have not been specified (Kwok et al. 2013; Odintsova et al. 2017; Dessai 2018; Potts et al. 2020) (Fig. 4/Online resource Table 4).
Depuration of animals and decontamination of tissue
Microbial contamination (protozoa, bacteria, and fungi) is a recurring issue in the development of primary cell culture systems, specifically in filter-feeders as the tissues are in direct contact with the surrounding water (Mialhe et al. 1988; Domart-Coulon et al. 1994; Bayne 1998; Chen and Wen 1999; van der Merwe et al. 2010). Of all the contaminants, thraustochytrids (Heterokont protists) are the most difficult to eliminate and often confused with the desired cells under culture (Rinkevich 1999). The thraustochytrids may be identified by fluorescence detection, electron microscopy, and enzyme assays (Mulcahy 2000).
Hetrick et al. (1981) observed that both the pumping activity of the oysters and several days of depuration were essential to keep down the protozoan population. For effective decontamination, Stephens and Hetrick (1979) recommended the depuration of animals for a week. However, abalones that were depurated for three days produced significantly lesser cell proliferation (24.96%) than the non-depurated ones (75.04%) (Suja et al., 2014) because prolonged periods of depuration would result in starved and stressed cells. Therefore, it is crucial to optimize the duration of depuration to obtain healthy cells free of contamination.
Decontamination of the tissues/ whole animal/ eggs is routinely carried out by employing chemicals (iodine (Hansen 1976), benzethonium chloride (Hansen 1976), hydrogen peroxide (Suwattana et al. 2010), sodium hypochlorite (Ellis and Bishop 1989; Barrick et al. 2018), ethanol (Stephens and Hetrick 1979; Chen and Wen 1999; Daugavet and Blinova 2015; Jayasankar et al. 2018), etc.), antibiotics such as penicillin and streptomycin, and antifungals such as amphotericin B. However, the continuous use of amphotericin B is not recommended (Perlman 1976) as it is reported to be toxic to cells (van der Merwe et al. 2010; Barrick et al. 2018). In fact, all antimicrobials are recommended to be used only in the wash steps and initial culture medium and removed in the subsequent steps (Mulcahy 2000; van der Merwe et al. 2010; You et al. 2012b, a; Suja et al. 2014, 2017).
In 2010–2020, the majority of the experiments (24 numbers) (Suwattana et al. 2010; Hughes et al. 2010; Lopes et al. 2011; Bolognesi and Fenech 2012; Rioult et al. 2013; Ivanina et al. 2013; Kwok et al. 2013; Jiang et al. 2014; Katsumiti et al. 2014; Kim et al. 2014; Troncone et al. 2015; Wang et al. 2015; Ladhar-Chaabouni et al. 2015; Oyanedel et al. 2016; Li et al. 2017; Morga et al. 2017; Odintsova et al. 2017; Hu et al. 2017; Maselli et al. 2018; Boroda et al. 2019; Potts et al. 2020) favoured cell culture initiated without depuration or antibiotic wash, of which 13 experiments (Hughes et al. 2010; Bolognesi and Fenech 2012; Rioult et al. 2013; Kwok et al. 2013; Jiang et al. 2014; Katsumiti et al. 2014; Troncone et al. 2015; Wang et al. 2015; Ladhar-Chaabouni et al. 2015; Oyanedel et al. 2016; Li et al. 2017; Morga et al. 2017; Potts et al. 2020) utilized hemocytes that could be easily obtained without contamination (refer to “Choice of organ/tissue and the life-stage” section). 21 experiments (van der Merwe et al. 2010; Awaji and Machii 2011; Franco et al. 2011; Hanana et al. 2011; Hanquet et al. 2011; Droguet et al. 2012; Dyachuk 2013; Katsumiti et al. 2014; Maiorova and Odintsova 2015; Ryu et al. 2015; Daugavet and Blinova 2015; Cai et al. 2016; Richards et al. 2018; Xu et al. 2018; Dessai 2018; Jayasankar et al. 2018; Potts et al. 2020) decontaminated the tissues using antibiotics without depurating the animals. Out of the 15 experiments that depurated the animals, only 3 conducted depuration for a duration of 6–12 h (You et al. 2012b, a; Suja et al. 2017), whereas the rest of the experiments (12 numbers) conducted depuration for 1–3 days (van der Merwe et al. 2010; Gaume et al. 2012; O’Neill et al. 2013; Pichon et al. 2013; Suja et al. 2014; Xiang et al. 2014; Balbi et al. 2017; Ji et al. 2017; Barrick et al. 2018; Blanco et al. 2019). 3 experiments were conducted by supplementing antibiotics during depuration (Xiang et al. 2014; Balbi et al. 2017; Suja et al. 2017). The sterilization of tissues using antibiotics is highly recommended and has been carried out by 55% (van der Merwe et al. 2010; Awaji and Machii 2011; Hanquet et al. 2011; Franco et al. 2011; Hanana et al. 2011; You et al. 2012b, a; Droguet et al. 2012; Gaume et al. 2012; O’Neill et al. 2013; Pichon et al. 2013; Dyachuk 2013; Katsumiti et al. 2014; Suja et al. 2014, 2017; Xiang et al. 2014; Maiorova and Odintsova 2015; Ryu et al. 2015; Daugavet and Blinova 2015; Cai et al. 2016; Jayasankar et al. 2018; Richards et al. 2018; Xu et al. 2018; Dessai 2018; Blanco et al. 2019; Potts et al. 2020) of the experiments (Fig. 5/Online resource Table 5).
Fig. 5.

Depuration of animals (with or without antibiotics) and sterilization of tissue for the initiation of marine molluscan cell culture. M mantle, L larva, DG digestive gland, HC hemocytes, H heart, GD gonad, G gills, N neuron, LP labial palps, R radula, AM adductor muscle, Depurated with Ab- depuration of animals in the presence of antibiotics, Ab wash- tissue washed with antibiotics (results from 60 experiments)
Cell culture media and supplements
The availability of information on the hemolymph components offers significant advantages for the development of de novo medium for cell culture or the optimization of existing cell culture media. Based on this concept, Machii and Wada (1989) developed a cell culture medium (Pf35) based on the composition of hemolymph from Pinctada fucata. The de novo medium has not produced results different from that of modified commercial media. Therefore, researchers tend to prefer the latter due to the relative ease of use. Preferred commercial media include Eagle’s MEM, Medium 199, Leibovitz’s L-15, etc. (Domart-Coulon et al. 1994).
The genome of several molluscan species is currently available (reviewed by Schell et al. 2017; Sigwart et al. 2021). The genomic information combined with transcriptomic data can be used to identify the species-specific and cell/tissue-specific genes and substrates that are required for cell survival and growth in vitro which in turn, can be used for the optimization of the cell culture medium. Pérez-Fernández et al. (2021) have simulated an optimized cell culture medium based on the genome-scale metabolic network of CHO-K1 (mammalian cell line). Though this is a relatively new field of research and so much yet to be explored, it offers feasible solutions towards the development of species-specific cell culture medium.
In 2010–2020, the majority of the experiments favoured Leibovitz’s L-15 as the cell culture medium (53%) (Fig. 6/Online resource Table 6) (Suwattana et al. 2010; Hanquet et al. 2011; You et al. 2012b, a; Gaume et al. 2012; O’Neill et al. 2013; Pichon et al. 2013; Dyachuk 2013; Jiang et al. 2014; Katsumiti et al. 2014; Kim et al. 2014; Maiorova and Odintsova 2015; Ryu et al. 2015; Troncone et al. 2015; Daugavet and Blinova 2015; Oyanedel et al. 2016; Cai et al. 2016; Balbi et al. 2017; Ji et al. 2017; Li et al. 2017; Odintsova et al. 2017; Hu et al. 2017; Barrick et al. 2018; Maselli et al. 2018; Richards et al. 2018; Xu et al. 2018; Dessai 2018; Blanco et al. 2019), followed by the use of seawater (natural or artificial) (15%) (Hughes et al. 2010; van der Merwe et al. 2010; Franco et al. 2011; Ivanina et al. 2013; Morga et al. 2017; Suja et al. 2017; Jayasankar et al. 2018; Boroda et al. 2019), a combination of L-15 and seawater (8%) (Hanana et al. 2011; Droguet et al. 2012; Potts et al. 2020), Hank’s balanced salt solution (5%) (Bolognesi and Fenech 2012; Kwok et al. 2013), and a combination of seawater and Opti-MEM® I reduced serum media (5%) (Potts et al. 2020). The use of other media (Marine Molluscan Balanced Salt Solution (MMBSS), Dulbecco's Modified Eagle Medium (DMEM), Medium 199, Basal Medium Eagle (BME), saline solution with a similar ionic composition of clam hemolymph, Marine Physiological Saline Solution (MPSS), a combination of L-15 and M-199, and a combination of seawater and M-199) were limited to an average of only one experiment each (Awaji and Machii 2011; Lopes et al. 2011; Rioult et al. 2013; Katsumiti et al. 2014; Kim et al. 2014; Suja et al. 2014; Xiang et al. 2014; Ladhar-Chaabouni et al. 2015; Wang et al. 2015). Cell culture systems that could be maintained for more than a year (in the decade 2010–2020) were cultured using the media-modified L-15 medium and sterilized seawater (supplemented with antibiotics and yeast extract) (Daugavet and Blinova 2015; Jayasankar et al. 2018).
Fig. 6.

Cell culture medium used for the development of marine molluscan cell culture. L-15-Leibovitz's L-15 Medium, SW-seawater, HBSS-Hank's Balanced Salt Solution, Opti-MEM-Opti-MEM® I reduced serum media (results from 60 experiments)
The serum is a source of growth factors, adhesion factors, vitamins, minerals, lipids, proteins, and hormones (Freshney 2010). Improved cell growth and viability have been observed by adding FBS/FCS and extract from molluscan hemolymph, larvae, and gonads (Hetrick et al. 1981; Domart-Coulon et al. 1994; Chen and Wen 1999). However, the use of FBS/FCS at a higher concentration (20%) has been reported to be toxic to cells (Domart-Coulon et al. 1994).
FBS/FCS being derived from a phylogentically different taxon, its functionality with respect to growth factors and hormones on cell culture derived from molluscs is questionable. However, sequence analysis of several growth factors, hormones and its receptors shows that these are homologous across different phyla (Ferkovich and Oberlander 1991; Reich 1992; van Kesteren et al. 1992; Herpin et al. 2004; Mimoto et al. 2007; Stout et al. 2010). Homologs of vertebrate growth factors have been also reported in invertebrates (reviewed by Ottaviani et al. 1996; Kipryushina et al. 2015) and the functional phylogenetics of these molecules have been discussed by Tettamanti et al. (2006). The positive influence of vertebrate growth factors on invertebrate cell culture systems has also been reported (reviewed by Ferkovich and Oberlander 1991; Mckay et al. 1999; Tettamanti et al. 2006). Moreover, researchers have discovered steroid hormones and its receptors in invertebrates (D’Aniello et al. 1996; Di Cosmo et al. 1998, 2002; Mimoto et al. 2007; Stout et al. 2010; Jones et al. 2017) followed by experiments revealing that steroid hormone and receptor functions are conserved in vertebrates and invertebrates (Di Cosmo et al. 2001; Mimoto et al. 2007; Stout et al. 2010; Jones et al. 2017).
In the decade 2010–2020, FBS/FCS was not used in 60% of the experiments (Fig. 7/Online resource Table 7), of which 72% involved applied studies. 82% of the experiments in the decade 2010–2020 did not use any extracts as supplements in the cell culture media, of which 61% (Hughes et al. 2010; Awaji and Machii 2011; Hanquet et al. 2011; Lopes et al. 2011; Hanana et al. 2011; Bolognesi and Fenech 2012; Droguet et al. 2012; Gaume et al. 2012; Ivanina et al. 2013; Kwok et al. 2013; O’Neill et al. 2013; Pichon et al. 2013; Dyachuk 2013; Jiang et al. 2014; Katsumiti et al. 2014; Xiang et al. 2014; Ladhar-Chaabouni et al. 2015; Maiorova and Odintsova 2015; Cai et al. 2016; Balbi et al. 2017; Li et al. 2017; Morga et al. 2017; Odintsova et al. 2017; Xu et al. 2018; Blanco et al. 2019; Boroda et al. 2019) involved applied studies.
Fig. 7.

FBS/FCS (in %) used as a supplement in cell culture medium for the development of marine molluscan cell culture (results from 60 experiments)
Sera and extracts are not used in applied studies to prevent interference in the results. However, it is also to be noted that three cell culture systems could be maintained for 1–2 months without the addition of any extracts or FBS/FCS (Pichon et al. 2013; Potts et al. 2020). 10% of the experiments in the decade 2010–2020 used hemolymph extract (Kim et al. 2014; Ryu et al. 2015; Troncone et al. 2015; Oyanedel et al. 2016; Hu et al. 2017; Ji et al. 2017), 7% used yeast extract (You et al. 2012a, b; Suja et al. 2017; Jayasankar et al. 2018) and 1% used chicken serum (Richards et al. 2018).
In the decade 2010–2020, 82% of the experiments have supplemented the cell culture media with antimicrobials (Suwattana et al. 2010; van der Merwe et al. 2010; Awaji and Machii 2011; Hanana et al. 2011; Hanquet et al. 2011; You et al. 2012b, a; Droguet et al. 2012; Gaume et al. 2012; Ivanina et al. 2013; O’Neill et al. 2013; Pichon et al. 2013; Rioult et al. 2013; Jiang et al. 2014; Katsumiti et al. 2014; Kim et al. 2014; Suja et al. 2014, 2017; Xiang et al. 2014; Ladhar-Chaabouni et al. 2015; Maiorova and Odintsova 2015; Ryu et al. 2015; Troncone et al. 2015; Wang et al. 2015; Daugavet and Blinova 2015; Oyanedel et al. 2016; Cai et al. 2016; Balbi et al. 2017; Ji et al. 2017; Li et al. 2017; Morga et al. 2017; Jayasankar et al. 2018; Barrick et al. 2018; Richards et al. 2018; Xu et al. 2018; Dessai 2018; Blanco et al. 2019; Boroda et al. 2019; Potts et al. 2020). However, the use of antimicrobials is to be limited, as described earlier in “Depuration of animals and decontamination of tissue” section.
Attempts on virus-cultivation in vitro
Only a few reports are available on the cultivation of viruses in molluscan cell culture. In a review article by Renault and Novoa (2004), cultivation of herpes-like virus in the primary embryonic cell culture of the oyster and cell lines of the fish have been reported. Even though cytopathic effects (CPE) could not be observed in the tested fish cell lines, viral DNA could be detected in the infected oyster primary cell culture by way of PCR and in situ hybridization. However, details on the experiments conducted are unavailable.
Nakatsugawa et al. (1999) could isolate a virus from the primary culture of hemocytes of the abalone, Nordotis discus discus affected by amyotrophia (atrophy of the foot muscle). The infected cell cultures showed CPE, such as cell aggregation, followed by the detachment of cells. The disease could not be reproduced in healthy juvenile abalones by injecting the virus-containing supernatant of the infected hemocyte cultures. However, it could be transmitted to healthy abalones by introducing the filtrate of the naturally infected abalones. The virus particles in the CPE-forming cultures were found to be icosahedral, 120 nm in diameter, and resembled retro-like virus particles when examined by transmission electron microscopy (TEM).
During the decade 2010–2020, only two reports are available on the cultivation of virus in molluscan cells in vitro. In a study by Ji et al. (2017), hemocyte culture of the scallop Chlamys farreri was challenged with the ostreid herpesvirus 1 (OsHV-1). The CPE that could be observed was cell aggregation, followed by the detachment of cells. The TEM examination of infected cells revealed polygonal nucleocapsids (virus particles) of 130–150 nm in diameter and empty capsids inside the cell nucleus. The gene expression study of QM (QM, an immune response gene) of the infected hemocytes showed a significantly higher level of expression at 12 h and 24 h post-challenge with OsHV-1. A significantly higher level of virus DNA could be detected in the cells 48 h post-infection by way of real-time PCR.
In another study by Morga et al. (2017), hemocyte culture from the spat of C. gigas was challenged with OsHV-1. Post-infection, a time-based quantification by real-time PCR showed a significant increase in the detection of virus DNA, differential patterns in the expression of 15 virus genes, and a significant upregulation of 2 host immune genes. These results suggested that the replication of OsHV-1 occurred in vitro. However, the CPE of the infected cells has not been specified. Moreover, virus particles could not be detected following the TEM examination.
Attempts on immortalization of molluscan cells in vitro
Normal cells usually have a finite life span. Still, certain cells under culture may show an enhanced growth rate and give rise to a continuous cell line, called spontaneous immortalization. Continuous cell lines are preferred over primary cultures as the former are homogeneous cultures, producing identical replicate cultures, and are relatively easier to handle. Immortalization can also be induced by the inhibition of cell cycle regulatory genes (like Rb and p53 genes) through transfection with the viral oncogenes (like SV 40 (Simian Virus 40) LT gene, human papillomavirus (HPV) E6 and E7 genes, and Epstein Barr Virus (EBV) gene). Other methods include transfection of the cells with telomerase gene and a regulatory promoter to overcome the Hayflick limit and fusion of target cells with that of the established cell line to produce hybrid cells (Bolton and Spurr 1996; Mayne et al. 1996; Peters et al. 1996; Bayne 1998; Bodnar et al. 1998; Vaziri and Benchimol 1998; Freshney 2010).
Successful immortalization in cell culture from marine invertebrates is yet to be reported (Rinkevich 2011). Four transfection techniques have been primarily used in the bivalves-microinjection, lipofection, electroporation, and particle bombardment, of which particle bombardment seems to be the most suitable (reviewed by Cadoret et al., 2000). There are also reports on the successful transfection of eggs and spermatozoa of bivalve molluscs using naked DNA (plasmid pGeneGrip containing GFP (green fluorescent protein)) (Guerra et al. 2005; Guerra and Esponda 2006).
In the case of expression vectors, integrative vectors are advantageous in that these can integrate the transfected DNA into host chromosomes and offer stable gene expression (Mialhe et al. 1995). While heterologous oncogenes could be used for immortalization experiments, homologous oncogenes have also been sought in the molluscs (Madaule and Axel 1985; Van Beneden et al. 1993; Van Beneden 1994; Kelley et al. 2001).
The experiments which have utilized heterologous promoters such as the Drosophila heat shock protein (hsp) 70, hsp82, and baculovirus IE1 promoter have shown efficiency in expressing transgenes in the molluscan cell cultures (in the literature cited by Cadoret et al., 2000). Researchers have also succeeded in expressing transgenes in the embryos and primary cell cultures of bivalves using homologous promoters (Cadoret et al. 1999; Yue et al. 2020).
The only cell culture systems that have been used to induce genetic transformation in molluscs were those that were derived from the heart and embryos/larvae. Transformation experiments have been conducted by Hetrick et al. (1981) on C. virginica by the injection of carcinogens (benzo(a)pyrene, 3-methylcholanthrene, and N-methyl-N’-nitrosoguanidine) into the animal as well as by the direct exposure of carcinogens on the oyster heart primary cell culture. Apart from observing large amoebocytes, no mitotically active cells could be attained. The primary heart cell culture from C. gigas was treated with the mutagens, ethyl methanesulfonate (EMS) and quercitin for different periods but yielded no fruitful results (Delsert and Cancela 1998). An induced culture of neoplastic hemocytes of M. arenaria has been reported, as described earlier in “Choice of organ/tissue and the life-stage” section (Oprandy and Chang 1983).
The successful transgene expression in the molluscan cells marks a critical event prior to plausible attempts on the immortalization of the cells. The first transgenic bivalve was produced through the electroporation-mediated transfection of the fertilized eggs of the clam, Mulinia lateralis with the pantropic retroviral vectors (LSRNL-(VSV-G) incorporated with the neomycin resistance gene (neo), and Geo 4.8 vector incorporated with the neo and β-galactosidase (β-gal) genes). The fertilized eggs were reared to adulthood, with the survival rate of the embryos being affected by stress due to electroporation rather than the introduction of retroviral vectors. Further experiments revealed the presence of an integrated, single copy of the provirus per transgenic offspring and that the transgenic germ-lines of the animals were mosaic for the provirus (Lu et al. 1996). Transfection experiments on the primary cell cultures of the heart and embryo of C. gigas were conducted using various heterologous promoters for the expression of luciferase gene. Success was achieved using hsp70 and retroviral promoters (MoMLV LTR and RSV LTR) in heart and embryo cell cultures, respectively (Boulo et al. 1996, 2000). Successful microinjection-mediated transfection in the fertilized eggs of C. gigas has been achieved with the use of homologous gene promoter, elongation factor-1-α (EF-1α) of C. gigas, cloned into a pEGFP-1 vector that drives the expression of EGFP (enhanced green fluorescence protein) (Yue et al. 2020).
The attempts on the excision and transposition of the transposable elements, hobo and hermes, in the primary heart cell culture of the oyster, C. gigas by lipofection resulted in the incorrect excision of the transposons from the donor plasmid. This was suspiciously attributed to the low quantity of the transposase enzyme expressed, and/or complete lack of or insufficient quantity of the co-factors required for the functionality of the transposase (Boulo 1997; Cadoret et al. 2000). The sperm-mediated transfection of the PiggyBac transposon vector with gGH (growth hormone gene from orange-spotted grouper) and GFP in the eggs of C. gigas gave rise to larvae which exhibited green fluorescence under an inverted phase contrast fluorescence microscope. The transgenesis was confirmed by PCR and genome walking analysis (Chen et al. 2018).
The electroporation-mediated co-transfection of the reporter genes (neo) and oncogenes (ras or myc) in the primary larval cell cultures from the oyster was carried out by Ellis (1991). Even though the transfection was successful, it failed to induce mitotically active cells.
The lipofection-mediated transfection of the mussel-specific expression vectors in the primary heart cell culture of C. gigas was undertaken by Delsert and Cancela (1998). The designed vectors contained mussel-actin promoter region (based on the genomic DNA library of M. galloprovincialis), and lacZ or luciferase reporter sequence incorporated into the commercial plasmid, pBgal-Basic vector. Similarly, transfection experiments using the insect expression vector, A3lacZ (containing lacZ or luciferase reporter sequence under the control of actin A3 gene promoter of Bombyx mori) and the mammalian expression vector (containing lacZ reporter sequence under the control of viral promoter CMV) were conducted on the primary culture of oyster heart cells. Their study concluded that the oyster cells could be transfected but with low efficiency. The attempts to transform oyster cells by placing SV40 LT oncogene under the control of mussel-actin promoter (using p.mac-4 T Ag vector) also failed to produce desirable results. The electroporation-mediated transfection of the heterologous vector A3lacZ in the oyster eggs resulted in transfection efficiency of 1% in the developed trochophore and veliger larvae.
Similar to the work of Delsert and Cancela (1998), a homologous actin Gia1 gene promoter (1670 bp) was constructed from the genomic library of C. gigas, and a specific expression vector, Gia-L3 containing this promoter and luciferase gene, was produced. The lipofection-mediated transfection of this plasmid in the primary heart cell culture of C. gigas resulted in successful expression of the transgene (Cadoret et al. 1999).
During the decade 2010–2020, experiments on the immortalization of molluscan cells in vitro have not been reported. Despite reports on the successful expression of transgenes in molluscan cells, the literature on immortalization-attempts of molluscan cells has been restricted to two failed attempts (Ellis 1991; Delsert and Cancela 1998). Other attempts on the immortalization of molluscan cells may not have been published due to the resistance from scientific journals in accepting failed experiments, which is the same concern expressed by Rinkevich (1999) earlier.
Perspectives on the development of marine molluscan cell line
The promising candidates for the future molluscan cell line would be natural or induced pluripotent stem cells (iPSc) (Takahashi and Yamanaka 2006), adult stem cells (reviewed by Rinkevich, 2011) and neoplastic cells (Walker et al. 2009; Odintsova et al. 2011). The identification of genes and factors involved in cellular quiescence (Yoshino et al. 2013) and totipotency or pluripotency of stem cells (Yamanaka 2008) shall aid in enhancing the proliferative capacity of the cells in vitro.
In addition to the use of extracellular matrix (ECM) components, identification of apt growth promoters (Cai et al. 2014), hemolymph and omics data analysis for the development of specific cell culture medium (Jayesh et al. 2012; 2013; 2015), experiments focusing on the immortalization of molluscan cells have to be taken forward. The identification of suitable promoter systems is crucial before attempting to transfect cells in vitro. The intron sequences in the candidate promoter sequence is not to be neglected wholly. For instance, in an experiment conducted by Yue et al. (2020), the EF-1α promoter sequence induced the expression of EGFP in the fertilized eggs of C. gigas only when the first intron sequence was also inserted. The search for suitable heterologous and homologous promoters is to be continued until stable transgene expression is acquired in molluscan cells that would lead to immortalization.
Retroviral vectors and transposons are integrative vectors that are highly likely to produce integrated transgenes in the host chromosome rather than existing as episomal elements. This integration would offer a stable expression of transgenes. In addition to this, the pseudotyped retroviral vectors (with the envelope glycoprotein of vesicular stomatitis virus (VSV-G)) (Emi et al. 1991) are preferable due to the broad host cell range.
In the case of non-viral vectors, plasmids can be facilitated to enter the nucleus of non-dividing cells by the use of DNA nuclear targeting sequences (DTS) such as the enhancer sequence (72 bp) of the SV 40 genome and transcription factors (which contain nuclear localization signals (NLS)) (Dean et al. 2005).
The fact that one out of the 700 primary embryonic cell cultures set up by Hansen (1976) had given rise to the spontaneous Bge cell line, the only molluscan cell line in the world to date, pinpoint the significance of consistent and solidarized efforts required for the development of a marine molluscan cell line.
Supplementary Information
Below is the link to the electronic supplementary material.
Author contributions
SB prepared the main manuscript, including all figures and tables. ISBS and JP conceptualized, reviewed, corrected and finally approved the manuscript.
Funding
The original research was financially supported by the Junior Research Fellowship of the Council for Scientific and Industrial Research (CSIR), Govt. of India (No. 09/239(0553)/2019-EMR-I). The first author thanks CSIR for the fellowship.
Data availability
All data generated or analyzed during this study are included in this published article.
Declarations
Competing interest
The authors declare that they have no competing interests.
Consent for publication
All authors approved the manuscript to be published.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
I. S. Bright Singh, Email: isbsingh@gmail.com.
Jayesh Puthumana, Email: jayesh@cusat.ac.in.
References
- Alavi MR, Fernández-Robledo JA, Vasta GR. Development of an in vitro assay to examine intracellular survival of Perkinsus marinus trophozoites upon phagocytosis by oyster (Crassostrea virginica and Crassostrea ariakensis) hemocytes. J Parasitol. 2009;95:900–907. doi: 10.1645/GE-1864.1. [DOI] [PubMed] [Google Scholar]
- Arzul I, Corbeil S, Morga B, Renault T. Viruses infecting marine molluscs. J Invertebr Pathol. 2017;147:118–135. doi: 10.1016/j.jip.2017.01.009. [DOI] [PubMed] [Google Scholar]
- Awaji M, Machii A. Fundamental studies on in vivo and in vitro pearl formation—contribution of outer epithelial cells of pearl oyster mantle and pearl sacs. Aqua-BioSci Monogr. 2011;4:1–39. doi: 10.5047/absm.2011.00401.0001. [DOI] [Google Scholar]
- Balbi T, Ciacci C, Grasselli E, et al. Utilization of Mytilus digestive gland cells for the in vitro screening of potential metabolic disruptors in aquatic invertebrates. Comp Biochem Physiol Part C Toxicol Pharmacol. 2017;191:26–35. doi: 10.1016/j.cbpc.2016.08.009. [DOI] [PubMed] [Google Scholar]
- Barrick A, Guillet C, Mouneyrac C, Châtel A. Investigating the establishment of primary cultures of hemocytes from Mytilus edulis. Cytotechnology. 2018;70:1205–1220. doi: 10.1007/s10616-018-0212-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayne CJ. Biotechnology in the study of molluscs. Bul Penelit Kesehat. 1989;17:141–147. [Google Scholar]
- Bayne CJ. Invertebrate cell culture considerations: insects, ticks, shellfish, and worms. In: Mather JP, Barnes D, editors. Methods in cell biology. New York: Academic Press; 1998. pp. 187–201. [DOI] [PubMed] [Google Scholar]
- Bayne CJ, Owczarzak A, Allen JR. Molluscan (Biomphalaria) cell line: serology, karyotype, behavioral, and enzyme electrophoretic characterization. J Invertebr Pathol. 1978;32:35–39. doi: 10.1016/0022-2011(78)90171-4. [DOI] [Google Scholar]
- Bayne CJ, Owczarzak A, Noonan WE. In vitro cultivation of cells and a microsporidian parasite of Biomphalaria glabrata (Pulmonata: Basommatophora) Ann New York Acad Sci. 1975;266:513–527. doi: 10.1111/j.1749-6632.1975.tb35127.x. [DOI] [PubMed] [Google Scholar]
- Bevelander G, Martin J. Culture of mantle tissue of marine molluscs. Anat Rec. 1949;105:614. [Google Scholar]
- Blanco J, Martín H, Mariño C, Rossignoli AE. Simple diffusion as the mechanism of okadaic acid uptake by the mussel digestive gland. Toxins. 2019;11:395–405. doi: 10.3390/toxins11070395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bodnar AG, Ouellette M, Frolkis M, et al. Extension of life-span by introduction of telomerase into normal human cells. Science. 1998;279:349–352. doi: 10.1126/science.279.5349.349. [DOI] [PubMed] [Google Scholar]
- Bolognesi C, Fenech M. Mussel micronucleus cytome assay. Nat Protoc. 2012;7:1125–1137. doi: 10.1038/nprot.2012.043. [DOI] [PubMed] [Google Scholar]
- Bolton BJ, Spurr NK. B-lymphocytes. In: Freshney RI, Freshney MG, editors. Culture of immortalized cells. New York: Wiley-Liss; 1996. pp. 283–297. [Google Scholar]
- Boroda AV, Kipryushina YO, Odintsova NA. Chemical modulation of apoptosis in molluscan cell cultures. Cell Stress Chaperones. 2019;24:905–916. doi: 10.1007/s12192-019-01014-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boulo V (1997) Transformation génétique chez les mollusques bivalves marins: analyses fonctionnelles in vitro et in vivo de vecteurs d’expression et d’intégration hétérologues. Thèse de doctorat de l’Ecole Pratique des Hautes Etudes
- Boulo V, Cadoret J, Le Marrec F, et al. Transient expression of luciferase reporter gene after lipofection in oyster (Crassostrea gigas) primary cell cultures. Mol Mar Biol Biotechnol. 1996;5:167–174. [PubMed] [Google Scholar]
- Boulo V, Cadoret J, Shike H, et al. Infection of cultured embryo cells of the pacific oyster, Crassostrea gigas, by pantropic retroviral vectors. Vitr Cell Dev Biol. 2000;36:395–399. doi: 10.1290/1071-2690(2000)036<0395:IOCECO>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Brewster F, Nicholson BL. In vitro maintenance of amoebocytes from the American oyster (Crassostrea virginica) J Fish Res Board Canada. 1979;36:461–467. doi: 10.1139/f79-064. [DOI] [Google Scholar]
- Cadoret JP, Bachère E, Roch P, et al. Genetic transformation of farmed marine bivalve molluscs. In: Fingerman M, Nagabhushanam R, et al., editors. Recent advances in marine biotechnology: aquaculture: seaweeds and invertebrates, Part A. Enfield: Science Publishers Inc; 2000. pp. 111–126. [Google Scholar]
- Cadoret JP, Debón R, Cornudella L, et al. Transient expression assays with the proximal promoter of a newly characterized actin gene from the oyster Crassostrea gigas. FEBS Lett. 1999;460:81–85. doi: 10.1016/s0014-5793(99)01319-8. [DOI] [PubMed] [Google Scholar]
- Cai X, Zhang Y, Publication AÁ, Cell ÁPÁ. Marine invertebrate cell culture: a decade of development. J Oceanogr. 2014;70:405–414. doi: 10.1007/s10872-014-0242-8. [DOI] [Google Scholar]
- Cai Y, Pan L, Miao J. In vitro study of the effect of metabolism enzymes on benzo(a)pyrene-induced DNA damage in the scallop Chlamys farreri. Environ Toxicol Pharmacol. 2016;42:92–98. doi: 10.1016/j.etap.2016.01.009. [DOI] [PubMed] [Google Scholar]
- Chen J, Wu C, Zhang B, et al. PiggyBac transposon-mediated transgenesis in the pacific oyster (Crassostrea gigas)—first time in mollusks. Front. 2018;9:811. doi: 10.3389/fphys.2018.00811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen S, Wen C. Establishment of cell lines derived from oyster, Crassostrea gigas Thunberg and hard clam, Meretrix lusoria Röding. Methods Cell Sci. 1999;21:183–192. doi: 10.1023/a:1009829807954. [DOI] [PubMed] [Google Scholar]
- Comps M, Bonami J-R, Vago C, Campillo A. Une virose de l’huître portugaise (Crassostea angulata LMK) Comptes Rendus Académie Des Sci Paris. 1976;282:1991–1993. [Google Scholar]
- Cornet M. Detection of genotoxicity in the marine environment: a preliminary feasibility study using primary mussel tissue culture. Sci Total Environ. 2007;382:22–29. doi: 10.1016/j.scitotenv.2007.03.033. [DOI] [PubMed] [Google Scholar]
- D’Aniello A, Di Cosmo A, Di Cristo C, et al. Occurrence of sex steroid hormones and their binding proteins in Octopus vulgaris Lam. Biochem Biophys Res Commun. 1996;227:782–788. doi: 10.1006/bbrc.1996.1585. [DOI] [PubMed] [Google Scholar]
- Daugavet MA, Blinova MI. Culture of mussel (Mytilus edulis L.) mantle cells. Cell Tissue Biol. 2015;9:233–243. doi: 10.1134/S1990519X15030037. [DOI] [PubMed] [Google Scholar]
- Davids BJ, Wu XJ, Yoshino TP. Cloning of a β integrin subunit cDNA from an embryonic cell line derived from the freshwater mollusc, Biomphalaria glabrata. Gene. 1999;228:213–223. doi: 10.1016/S0378-1119(99)00008-6. [DOI] [PubMed] [Google Scholar]
- de Kantzow MC, Hick PM, Dhand NK, Whittington RJ. Risk factors for mortality during the first occurrence of Pacific Oyster Mortality Syndrome due to Ostreid herpesvirus—1 in Tasmania, 2016. Aquaculture. 2017;468:328–336. doi: 10.1016/j.aquaculture.2016.10.025. [DOI] [Google Scholar]
- Dean DA, Strong DD, Zimmer WE. Nuclear entry of nonviral vectors. Gene Ther. 2005;12:881–890. doi: 10.1038/sj.gt.3302534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delsert C, da Cancela FL. Development of mussel-specific expression vectors suitable for transgenic organisms: implication on the establishment of mollusc continuous cell lines. Montpellier: IFREMER; 1998. [Google Scholar]
- Dessai SN. Cryopreservation of cultured mantle cells of Paphia malabarica for perennial availability. Cryobiology. 2018;82:93–98. doi: 10.1016/j.cryobiol.2018.04.002. [DOI] [PubMed] [Google Scholar]
- Le DR, Lipart C, Renault T. Primary culture of Pacific oyster, Crassostrea gigas, heart cells. J Tissue Cult Methods. 1994;16:67–72. doi: 10.1007/BF01404838. [DOI] [Google Scholar]
- Di Cosmo A, Di Cristo C, Paolucci M. A estradiol-17β receptor in the reproductive system of the female of Octopus vulgaris: characterization and immunolocalization. Mol Reprod Dev. 2002;61:367–375. doi: 10.1002/mrd.10014. [DOI] [PubMed] [Google Scholar]
- Di Cosmo A, Di Cristo C, Paolucci M. Sex steroid hormone fluctuations and morphological changes of the reproductive system of the female of Octopus vulgaris throughout the annual cycle. J Exp Zool. 2001;289:33–47. doi: 10.1002/1097-010X(20010101/31)289:1<33::AID-JEZ4>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- Di Cosmo A, Paolucci M, Di Cristo C, et al. Progesterone receptor in the reproductive system of the female of Octopus vulgaris: characterization and immunolocalization. Mol Reprod Dev. 1998;50:451–460. doi: 10.1002/(SICI)1098-2795(199808)50:4<451::AID-MRD9>3.0.CO;2-H. [DOI] [PubMed] [Google Scholar]
- Domart-Coulon I, Auzoux-Bordenave S, Doumenc D, Khalanski M. Cytotoxicity assessment of antibiofouling compounds and by-products in marine bivalve cell cultures. Toxicol Vitro. 2000;14:245–251. doi: 10.1016/s0887-2333(00)00011-4. [DOI] [PubMed] [Google Scholar]
- Domart-Coulon I, Doumenc D, Auzoux-Bordenave S, Le Fichant Y. Identification of media supplements that improve the viability of primarily cell cultures of Crassostrea gigas oysters. Cytotechnology. 1994;16:109–120. doi: 10.1007/BF00754613. [DOI] [PubMed] [Google Scholar]
- Droguet M, Devauchelle N, Pennec J-P, et al. Cultured heart cells from oyster: an experimental approach for evaluation of the toxicity of the marine pollutant tributyltin. Aquat Living Resour. 2012;25:185–194. doi: 10.1051/alr/2012017. [DOI] [Google Scholar]
- Dyachuk V. Extracellular matrix is required for muscle differentiation in primary cell cultures of larval Mytilus trossulus (Mollusca : Bivalvia) Cryotechnology. 2013;65:725–735. doi: 10.1007/s10616-013-9577-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis LL (1991) Electroporation of oyster cells. In: Meeting abstract-1991 world congress on cell and tissue culture. In vitro cellular & developmental biology—animal, 27, p 42A
- Ellis LL, Bishop SH. Isolation of cell lines with limited growth potential from marine bivalves. In: Mitsuhashi J, editor. Invertebrate cell system applications. Boca Raton: CRC Press; 1989. pp. 243–251. [Google Scholar]
- Emi N, Friedmann T, Yee JK. Pseudotype formation of murine leukemia virus with the G protein of vesicular stomatitis virus. J Virol. 1991;65:1202–1207. doi: 10.1128/jvi.65.3.1202-1207.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang Z, Feng Q, Chi Y, et al. Investigation of cell proliferation and differentiation in the mantle of Pinctada fucata (Bivalve, Mollusca) Mar Biol. 2008;153:745–754. doi: 10.1007/s00227-007-0851-5. [DOI] [Google Scholar]
- FAO . The State of World Fisheries and Aquaculture 2020. Sustainablity in action. Rome: Food and Agriculture Organization; 2020. [Google Scholar]
- Faucet J, Maurice M, Gagnaire B, et al. Isolation and primary culture of gill and digestive gland cells from the common mussel Mytilus edulis. Methods Cell Sci. 2004;25:177–184. doi: 10.1007/s11022-004-8227-4. [DOI] [PubMed] [Google Scholar]
- Ferkovich SM, Oberlander H. Growth factors in invertebrate in vitro culture. Vitr Cell Dev Biol. 1991;27:483–486. doi: 10.1007/BF02631149. [DOI] [PubMed] [Google Scholar]
- Franco A, Kellner K, Mathieu M, et al. Male germ cells of the Pacific oyster Crassostrea gigas: flow cytometry analysis, cell sorting and molecular expression. Aquat Living Resour. 2011;24:237–245. doi: 10.1051/alr/2011110. [DOI] [Google Scholar]
- Freshney RI. Culture of animal cells: a manual of basic technique and specialized applications. 6. Hoboken: Wiley-Blackwell; 2010. [Google Scholar]
- Gaume B, Bourgougnon N, Auzoux-Bordenave S, et al. In vitro effects of triclosan and methyl-triclosan on the marine gastropod Haliotis tuberculata. Comp Biochem Physiol Part C. 2012;156:87–94. doi: 10.1016/j.cbpc.2012.04.006. [DOI] [PubMed] [Google Scholar]
- Gómez-Mendikute A, Elizondo M, Venier P, Cajaraville MP. Characterization of mussel gill cells in vivo and in vitro. Cell Tissue Res. 2005;321:131–140. doi: 10.1007/s00441-005-1093-9. [DOI] [PubMed] [Google Scholar]
- Guerra R, Carballada R, Esponda P. Transfection of spermatozoa in bivalve molluscs using naked DNA. Cell Biol Int. 2005;29:159–164. doi: 10.1016/j.cellbi.2004.11.018. [DOI] [PubMed] [Google Scholar]
- Guerra R, Esponda P. Transfection of eggs in the bivalve mollusc Chamelea gallina (Bivalvia, Veneridae) J Submicrosc Cytol Pathol. 2006;38:5–10. [PubMed] [Google Scholar]
- Halanych KM, Kocot KM. Shellfish genes. Nat Ecol Evol. 2017;1:142. doi: 10.1038/s41559-017-0142. [DOI] [PubMed] [Google Scholar]
- Hanana H, Talarmin H, Pennec JP, et al. Establishment of functional primary cultures of heart cells from the clam Ruditapes decussatus. Cytotechnology. 2011;63:295–305. doi: 10.1007/s10616-011-9347-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanquet A-C, Jouaux A, Heude C, et al. A sodium glucose co-transporter (SGLT) for glucose transport into Crassostrea gigas vesicular cells: impact of alimentation on its expression. Aquaculture. 2011;313:123–128. doi: 10.1016/j.aquaculture.2011.01.015. [DOI] [Google Scholar]
- Hanselmann R, Smolowitz R, Gibson D. Identification of proliferating cells in hard clams. Biol Bull. 2000;199:199–200. doi: 10.2307/1542896. [DOI] [PubMed] [Google Scholar]
- Hansen EL. A cell line from embryos of Biomphalaria glabrata (Pulmonata): establishment and characteristics. In: Maramorosch K, editor. Invertebrate tissue culture: research applications. New York: Academic Press; 1976. pp. 75–99. [Google Scholar]
- Herpin A, Lelong C, Favrel P. Transforming growth factor-β related proteins: an ancestral and widespread superfamily of cytokines in metazoans. Dev Comp Immunol. 2004;28:461–485. doi: 10.1016/j.dci.2003.09.007. [DOI] [PubMed] [Google Scholar]
- Hetrick FM, Stephens E, Lomax N, Lutrell K (1981) Attempts to develop a marine molluscan cell line. University of Maryland Sea Grant College Program Technical Report UM-SG-TS-81-06, College Park
- Hine PM, Thorne T. Replication of herpes-like viruses in haemocytes of adult flat oysters Ostrea angasi: an ultrastructural study. Dis Aquat Org. 1997;29:189–196. doi: 10.3354/dao029189. [DOI] [Google Scholar]
- Hu J, Adler K, Farah CA, et al. Cell-specific PKM isoforms contribute to the maintenance of different forms of persistent long-term synaptic plasticity. J Neurosci. 2017;37:2746–2763. doi: 10.1523/JNEUROSCI.2805-16.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes FM, Foster B, Grewal S, Sokolova IM. Apoptosis as a host defense mechanism in Crassostrea virginica and its modulation by Perkinsus marinus. Fish Shellfish Immunol. 2010;29:247–257. doi: 10.1016/j.fsi.2010.03.003. [DOI] [PubMed] [Google Scholar]
- Ivanchenko MG, Lerner JP, Mccormick RS, et al. Continuous in vitro propagation and differentiation of cultures of the intramolluscan stages of the human parasite Schistosoma mansoni. Proc Natl Acad Sci USA. 1999;96:4965–4970. doi: 10.1073/pnas.96.9.4965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivanina AV, Beniash E, Etzkorn M, et al. Short-term acute hypercapnia affects cellular responses to trace metals in the hard clams Mercenaria mercenaria. Aquat Toxicol. 2013;140–141:123–133. doi: 10.1016/j.aquatox.2013.05.019. [DOI] [PubMed] [Google Scholar]
- Jayasankar V, Vasudevan SR, Poulose SC, Divipala I. Nacre formation by epithelial cell cultures from mantle of the black-lip pearl oyster, Pinctada margaritifera. Vitr Cell Dev Biol. 2018;54:477–485. doi: 10.1007/s11626-018-0269-z. [DOI] [PubMed] [Google Scholar]
- Jayesh P, Seena J, Singh ISB. Establishment of shrimp cell lines: perception and orientation. Indian J Virol. 2012;23:244–251. doi: 10.1007/s13337-012-0089-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jayesh P, Seena J, Philip R, Singh ISB. A novel medium for the development of in vitro cell culture system from Penaeus monodon. Cytotechnology. 2013;65:307–322. doi: 10.1007/s10616-012-9491-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jayesh P, Philip R, Singh ISB. Multifactorial interaction of growth factors on Penaeus monodon lymphoid cells and the impact of IGFs in DNA synthesis and metabolic activity in vitro. Cytotechnology. 2015;67:559–571. doi: 10.1007/s10616-014-9697-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ji A, Li X, Fang S, et al. Primary culture of Zhikong scallop Chlamys farreri hemocytes as an in vitro model for studying host-pathogen interactions. Dis Aquat Organ. 2017;125:217–226. doi: 10.3354/dao03145. [DOI] [PubMed] [Google Scholar]
- Jiang Q, Zhou Z, Wang L, et al. Mutual modulation between norepinephrine and nitric oxide in haemocytes during the mollusc immune response. Sci Rep. 2014;4:6963. doi: 10.1038/srep06963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones BL, Walker C, Azizi B, et al. Conservation of estrogen receptor function in invertebrate reproduction. BMC Evol Biol. 2017;17:65. doi: 10.1186/s12862-017-0909-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katsumiti A, Gilliland D, Arostegui I, Cajaraville MP. Cytotoxicity and cellular mechanisms involved in the toxicity of cds quantum dots in hemocytes and gill cells of the mussel Mytilus galloprovincialis. Aquat Toxicol. 2014;153:39–52. doi: 10.1016/j.aquatox.2014.02.003. [DOI] [PubMed] [Google Scholar]
- Kelley ML, Winge P, Heaney JD, et al. Expression of homologues for p53 and p73 in the softshell clam (Mya arenaria), a naturally-occurring model for human cancer. Oncogene. 2001;20:748–758. doi: 10.1038/sj.onc.1204144. [DOI] [PubMed] [Google Scholar]
- Kim MS, Nam YK, Kim DS, Gong SP. Initial culture conditions for primary cell populations derived from radula tissue in abalone Haliotis discus hannai. Fish Aquat Sci. 2014;17:385–390. doi: 10.5657/FAS.2014.0385. [DOI] [Google Scholar]
- Kipryushina YO, Yakovlev KV, Odintsova NA. Vascular endothelial growth factors: a comparison between invertebrates and vertebrates. Cytokine Growth Factor Rev. 2015;26:687–695. doi: 10.1016/j.cytogfr.2015.04.001. [DOI] [PubMed] [Google Scholar]
- Kwok A, Lyons BP, Hodges NJ, Bean TP. Cryopreservation and storage of mussel (Mytilus spp.) haemocytes for latent analysis by the Comet assay. Mutat Res Genet Toxicol Environ Mutagen. 2013;750:86–91. doi: 10.1016/j.mrgentox.2012.09.010. [DOI] [PubMed] [Google Scholar]
- Ladhar-Chaabouni R, Hamza-Chaffai A. The cell cultures and the use of haemocytes from marine molluscs for ecotoxicology assessment. Cytotechnology. 2016;68:1669–1685. doi: 10.1007/s10616-015-9932-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ladhar-Chaabouni R, Machreki-Ajmi M, Serpentini A, et al. Does a short-term exposure to cadmium chloride affects haemocyte parameters of the marine gastropod Haliotis tuberculata? Environ Sci Pollut Res. 2015;22:17343–17349. doi: 10.1007/s11356-014-3387-5. [DOI] [PubMed] [Google Scholar]
- Lardans V, Boulo V, Duclermortier P, et al. DNA transfer in a Biomphalaria glabrata embryonic cell line by DOTAP lipofection. Parasitol Res. 1996;82:574–576. doi: 10.1007/s004360050166. [DOI] [PubMed] [Google Scholar]
- Latire T, Le Pabic C, Mottin E, et al. Responses of primary cultured haemocytes from the marine gastropod Haliotis tuberculata under 10-day exposure to cadmium chloride. Aquat Toxicol. 2012;109:213–221. doi: 10.1016/j.aquatox.2011.09.017. [DOI] [PubMed] [Google Scholar]
- Laursen JR, Di Liu H, Wu XJ, Yoshino TP. Heat-shock response in a molluscan cell line: characterization of the response and cloning of an inducible HSP70 cDNA. J Invertebr Pathol. 1997;70:226–233. doi: 10.1006/jipa.1997.4686. [DOI] [PubMed] [Google Scholar]
- Leong CCW, Syed NI, Ca FLL. Retrograde degeneration of neurite membrane structural integrity of nerve growth cones following in vitro exposure to mercury. Membr Biophys Biochem. 2001;12:733–737. doi: 10.1097/00001756-200103260-00024. [DOI] [PubMed] [Google Scholar]
- Li Y, Zhang L, Qu T, et al. Conservation and divergence of mitochondrial apoptosis pathway in the Pacific oyster, Crassostrea gigas. Cell Death Dis. 2017;8:e2915. doi: 10.1038/cddis.2017.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopes TM, Barcarolli IF, de Oliveira CB, et al. Effect of copper on ion content in isolated mantle cells of the marine clam Mesodesma mactroides. Environ Toxicol Chem. 2011;30:1582–1585. doi: 10.1002/etc.528. [DOI] [PubMed] [Google Scholar]
- Louzao MC, Espiña B, Cagide E, et al. Cytotoxic effect of palytoxin on mussel. Toxicon. 2010;56:842–847. doi: 10.1016/j.toxicon.2010.02.027. [DOI] [PubMed] [Google Scholar]
- Lu JK, Chen TT, Allen SK, et al. Production of transgenic dwarf surfclams, Mulinia lateralis, with pantropic retroviral vectors. Proc Natl Acad Sci USA. 1996;93:3482–3486. doi: 10.1073/pnas.93.8.3482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Machii A, Wada TK. Some marine invertebrates tissue culture. In: Mitsuhashi J, editor. Invertebrate cell system applications. Boca Raton: CRC Press; 1989. pp. 225–233. [Google Scholar]
- Madaule P, Axel R. A novel ras-related gene family. Cell. 1985;41:31–40. doi: 10.1016/0092-8674(85)90058-3. [DOI] [PubMed] [Google Scholar]
- Maiorova MA, Odintsova NA. β integrin-like protein-mediated adhesion and its disturbances during cell cultivation of the mussel Mytilus trossulus. Cell Tissue Res. 2015;361:581–592. doi: 10.1007/s00441-015-2122-y. [DOI] [PubMed] [Google Scholar]
- Marigómez I, Lekube X, Cancio I. Immunochemical localisation of proliferating cells in mussel digestive gland tissue. Histochem J. 1999;31:781–788. doi: 10.1023/A:1003950003381. [DOI] [PubMed] [Google Scholar]
- Maselli V, Xu F, Syed NI, et al. A novel approach to primary cell culture for Octopus vulgaris neurons. Front Physiol. 2018;9:220. doi: 10.3389/fphys.2018.00220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayne LV, Price TNC, Moorwood K, Burke JF. Development of immortal human fibroblast cell lines. In: Freshney RI, Freshney MG, editors. Culture of immortalized cells. New York: Wiley-Liss; 1996. pp. 77–93. [Google Scholar]
- Mckay SE, Purcell AL, Carew TJ. Regulation of synaptic function by neurotrophic factors in vertebrates and invertebrates : implications for development and learning. Learn Mem. 1999;6:193–215. doi: 10.1101/lm.6.3.193. [DOI] [PubMed] [Google Scholar]
- Mialhe E, Bachere E, Boulo V, et al. Future of biotechnology-based control of disease in marine invertebrates. Mol Mar Biol Biotechnol. 1995;4:275–283. doi: 10.1007/BF02779021. [DOI] [PubMed] [Google Scholar]
- Mialhe E, Boulo V, Grizel H. Bivalve mollusc cell culture. Am Fish Soc. 1988;18:311–315. [Google Scholar]
- Mimoto A, Fujii M, Usami M, et al. Identification of an estrogenic hormone receptor in Caenorhabditis elegans. Biochem Biophys Res Commun. 2007;364:883–888. doi: 10.1016/j.bbrc.2007.10.089. [DOI] [PubMed] [Google Scholar]
- Mitchelmore CL, Birmelin C, Livingstone DR, Chipman JK. Detection of DNA strand breaks in isolated mussel (Mytilus edulis L.) digestive gland cells using the “Comet” assay. Ecotoxicol Environ Saf. 1998;41:51–58. doi: 10.1006/eesa.1998.1666. [DOI] [PubMed] [Google Scholar]
- Morga B, Arzul I, Chollet B, Renault T. Infection with the protozoan parasite Bonamia ostreae modifies in vitro haemocyte activities of flat oyster Ostrea edulis. Fish Shellfish Immunol. 2009;26:836–842. doi: 10.1016/j.fsi.2009.03.018. [DOI] [PubMed] [Google Scholar]
- Morga B, Faury N, Guesdon S, et al. Haemocytes from Crassostrea gigas and OsHV-1: a promising in vitro system to study host/virus interactions. J Invertebr Pathol. 2017;150:45–53. doi: 10.1017/CBO9781107415324.004. [DOI] [PubMed] [Google Scholar]
- Morga B, Renault T, Faury N, et al. Cellular and molecular responses of haemocytes from Ostrea edulis during in vitro infection by the parasite Bonamia ostreae. Int J Parasitol. 2011;41:755–764. doi: 10.1016/j.ijpara.2011.01.013. [DOI] [PubMed] [Google Scholar]
- Mulcahy MF. Culture of molluscan cells. In: Mothersill C, Austin B, editors. Aquatic invertebrate cell culture. Chichester: Springer-Praxis; 2000. pp. 165–181. [Google Scholar]
- Naganuma T, Degnan BM, Horikoshi K, Morse DE. Myogenesis in primary cell cultures from larvae of the abalone, Haliotis rufescens. Mol Mar Biol Biotechnol. 1994;3:131–140. [PubMed] [Google Scholar]
- Nakatsugawa T, Nagai T, Hiya K, et al. A virus isolated from juvenile Japanese black abalone Nordotis discus discus affected with amyotrophia. Dis Aquat Org. 1999;36:159–161. doi: 10.3354/dao036159. [DOI] [Google Scholar]
- O’Neill M, Gaume B, Denis F, Auzoux-Bordenave S. Expression of biomineralisation genes in tissues and cultured cells of the abalone Haliotis tuberculata. Cytotechnology. 2013;65:737–747. doi: 10.1007/s10616-013-9576-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ödblom MP, Williamson R, Jones MB. Ionic currents in cardiac myocytes of squid, Alloteuthis subulata. J Comp Physiol B. 2000;170:11–20. doi: 10.1007/s003600050002. [DOI] [PubMed] [Google Scholar]
- Odintsova NA, Boroda AV, Maiorova MA, Yakovlev KV. The death pathways in mussel larval cells after a freeze-thaw cycle. Cryobiology. 2017;77:41–49. doi: 10.1016/j.cryobiol.2017.05.009. [DOI] [PubMed] [Google Scholar]
- Odintsova NA, Dyachuk VA, Nezlin LP. Muscle and neuronal differentiation in primary cell culture of larval Mytilus trossulus (Mollusca: Bivalvia) Cell Tissue Res. 2010;339:625–637. doi: 10.1007/s00441-009-0918-3. [DOI] [PubMed] [Google Scholar]
- Odintsova NA, Khomenko AV. Primary cell culture from embryos of the Japanese scallop Mizuchopecten yessoensis (Bivalvia) Cytotechnology. 1991;6:49–54. doi: 10.1007/BF00353702. [DOI] [PubMed] [Google Scholar]
- Odintsova NA, Usheva LN, Yakovlev KV, Kiselev KV. Naturally occurring and artificially induced tumor-like formations in marine invertebrates: a search for permanent cell lines. J Exp Mar Bio Ecol. 2011;407:241–249. doi: 10.1016/j.jembe.2011.06.020. [DOI] [Google Scholar]
- Odoemelam E, Raghavan N, Miller A, et al. Revised karyotyping and gene mapping of the Biomphalaria glabrata embryonic ( Bge ) cell line. Int J Parasitol. 2009;39:675–681. doi: 10.1016/j.ijpara.2008.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oprandy JJ, Chang PW. 5-bromodeoxyuridine induction of hematopoietic neoplasia and retrovirus activation in the soft-shell clam, Mya arenaria. J Invertebr Pathol. 1983;42:196–206. doi: 10.1016/0022-2011(83)90062-9. [DOI] [PubMed] [Google Scholar]
- Ottaviani E, Franchini A, Kletsas D, Franceschi C. Presence and role of cytokines and growth factors in invertebrates. Ital J Zool. 1996;63:317–323. doi: 10.1080/11250009609356152. [DOI] [Google Scholar]
- Oyanedel D, Gonzalez R, Brokordt K, et al. Insight into the messenger role of reactive oxygen intermediates in immunostimulated hemocytes from the scallop Argopecten purpuratus. Dev Comp Immunol. 2016;65:226–230. doi: 10.1016/j.dci.2016.07.015. [DOI] [PubMed] [Google Scholar]
- Pazos AJ, Mathieu M. Effects of five natural gonadotropin-releasing hormones on cell suspensions of marine bivalve gonad: stimulation of gonial DNA synthesis. Gen Comp Endocrinol. 1999;113:112–120. doi: 10.1006/gcen.1998.7186. [DOI] [PubMed] [Google Scholar]
- Pérez-Fernández BA, Fernandez-de-Cossio-Diaz J, Boggiano T, et al. In-silico media optimization for continuous cultures using genome scale metabolic networks: the case of CHO-K1. Biotechnol Bioeng. 2021;118:1884–1897. doi: 10.1002/bit.27704. [DOI] [PubMed] [Google Scholar]
- Perlman D. Some guidelines for selection of antibiotics for elimination of unwanted microbial contaminants in tissue cultures. Tissue Cult Assoc Man. 1976;2:383–386. doi: 10.1007/BF00918289. [DOI] [Google Scholar]
- Peters DM, Dowd N, Brandt C, Compton T. Human papilloma virus E6/E7 genes can expand the lifespan of human corneal fibroblasts. Vitr Cell Dev Biol. 1996;32:279–284. doi: 10.1007/BF02723060. [DOI] [PubMed] [Google Scholar]
- Pichon D, Cudennec B, Huchette S, et al. Characterization of abalone Haliotis tuberculata-Vibrio harveyi interactions in gill primary cultures. Cytotechnology. 2013;65:759–772. doi: 10.1007/s10616-013-9583-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potts RWA, Gutierrez AP, Cortés-araya Y, et al. Developments in marine invertebrate primary culture reveal novel cell morphologies in the model bivalve Crassostrea gigas. PeerJ. 2020;8:e9180. doi: 10.7717/peerj.9180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raghavan VS, Jayasankar V, Suja CP, Laxmilatha P. Cell culture and gene expression studies in relation to biomineralization in the black-lip pearl oyster, Pinctada margaritifera. J Coast Res. 2019;86:156–163. doi: 10.2112/SI86-024.1. [DOI] [Google Scholar]
- Reich G. A new peptide of the oxytocin/vasopressin family isolated from nerves of the cephalopod Octopus vulgaris. Neurosci Lett. 1992;134:191–194. doi: 10.1016/0304-3940(92)90514-8. [DOI] [PubMed] [Google Scholar]
- Renault T, Flaujac G, Le Deuff RM. Isolation and culture of heart cells from the European flat oyster, Ostrea edulis. Methods Cell Sci. 1995;17:199–205. doi: 10.1007/BF00996127. [DOI] [Google Scholar]
- Renault T, Novoa B. Viruses infecting bivalve molluscs. Aquat Living Resour. 2004;17:397–409. doi: 10.1051/alr:2004049. [DOI] [Google Scholar]
- Richards M, Xu W, Mallozzi A, et al. Production of calcium-binding proteins in Crassostrea virginica in response to increased environmental CO2 concentration. Front Mar Sci. 2018;5:203. doi: 10.3389/fmars.2018.00203. [DOI] [Google Scholar]
- Ridgway RL, Syed N, Lukowiak K, Bulloch AGM. Nerve growth factor (NGF) induces sprouting of specific neurons of the snail, Lymnaea stagnalis. J Neurobiol. 1991;22:377–390. doi: 10.1002/neu.480220406. [DOI] [PubMed] [Google Scholar]
- Rinaldi G, Yan H, Nacif-Pimenta R, et al. Cytometric analysis, genetic manipulation and antibiotic selection of the snail embryonic cell line Bge from Biomphalaria glabrata, the intermediate host of Schistosoma mansoni. Int J Parasitol. 2015;45:527–535. doi: 10.1016/j.ijpara.2015.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rinkevich B. Cell cultures from marine invertebrates: obstacles, new approaches and recent improvements. Prog Ind Microbiol. 1999;35:133–153. doi: 10.1016/S0079-6352(99)80107-6. [DOI] [Google Scholar]
- Rinkevich B. Marine invertebrate cell cultures: new millennium trends. Mar Biotechnol. 2005;7:429–439. doi: 10.1007/s10126-004-0108-y. [DOI] [PubMed] [Google Scholar]
- Rinkevich B. Cell cultures from marine invertebrates: new insights for capturing endless stemness. Mar Biotechnol. 2011;13:345–354. doi: 10.1007/s10126-010-9354-3. [DOI] [PubMed] [Google Scholar]
- Rioult D, Lebel JM, Le Foll F. Cell tracking and velocimetric parameters analysis as an approach to assess activity of mussel (Mytilus edulis) hemocytes in vitro. Cytotechnology. 2013;65:749–758. doi: 10.1007/s10616-013-9558-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robledo Y, Cajaraville MP. Isolation and morphofunctional characterization of mussel digestive gland cells in vitro. Eur J Cell Biol. 1997;72:362–369. [PubMed] [Google Scholar]
- Ryu JH, Nam YK, Gong SP. Factors affecting primary cultures of abalone Haliotis discus hannai ovary-dissociated cells and general culture aspects. Fish Aquat Sci. 2015;18:81–88. doi: 10.5657/FAS.2015.0081. [DOI] [Google Scholar]
- Schell T, Feldmeyer B, Schmidt H, et al. An annotated draft genome for Radix auricularia (Gastropoda, Mollusca) Genome Biol Evol. 2017;9:585–592. doi: 10.1093/gbe/evx032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Segarra A, Pépin JF, Arzul I, et al. Detection and description of a particular Ostreid herpesvirus 1 genotype associated with massive mortality outbreaks of Pacific oysters, Crassostrea gigas, in France in 2008. Virus Res. 2010;153:92–99. doi: 10.1016/j.virusres.2010.07.011. [DOI] [PubMed] [Google Scholar]
- Sigwart JD, Lindberg DR, Chen C, Sun J. Molluscan phylogenomics requires strategically selected genomes. Philos Trans R Soc B Biol Sci. 2021;376:20200161. doi: 10.1098/rstb.2020.0161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stephens EB, Hetrick FM (1979) Decontamination of the american oyster tissues for cell and organ culture. In: Tissue culture association manual. Tissue Culture Association, Rockville, vol 5, pp 1029–1031
- Stout EP, La Clair JJ, Snell TW, et al. Conservation of progesterone hormone function in invertebrate reproduction. Proc Natl Acad Sci U S A. 2010;107:11859–11864. doi: 10.1073/pnas.1006074107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suja CP, Dharmaraj S. In vitro culture of mantle tissue of the abalone Haliotis varia Linnaeus. Tissue Cell. 2005;37:1–10. doi: 10.1016/j.tice.2004.08.002. [DOI] [PubMed] [Google Scholar]
- Suja CP, Raghavan VS, Jayasankar V, et al. Evaluation of different media for cell proliferation in mantle tissue culture of the green mussel, Perna viridis (Linnaeus, 1758) Indian J Fish. 2017;64:224–228. doi: 10.21077/ijf.2017.64.special-issue.76285-34. [DOI] [Google Scholar]
- Suja CP, Sukumaran N, Dharmaraj S. Effect of culture media and tissue extracts in the mantle explant culture of abalone, Haliotis varia Linnaeus. Aquaculture. 2007;271:516–522. doi: 10.1016/j.aquaculture.2007.04.086. [DOI] [Google Scholar]
- Suja CP, Sukumaran N, Dharmaraj S, Meena A. Effect of depuration of animals and use of antimicrobial agents on proliferation of cells and microbial contamination in in-vitro mantle explant culture of the abalone Haliotis varia Linnaeus. Indian J Fish. 2014;61:93–98. [Google Scholar]
- Suwattana D, Jirasupphachok J, Jarayabhand P, Koykul W. In vitro culture of gill and heart tissues of the abalone Haliotis asinina. J Shellfish Res. 2010;29:651–653. doi: 10.2983/035.029.0314. [DOI] [Google Scholar]
- Syed NI, Ridgway RL, Lukowiak K, Bulloch AGM. Transplantation and functional integration of an identified respiratory interneuron in Lymnaea stagnalis. Neuron. 1992;8:767–774. doi: 10.1016/0896-6273(92)90097-w. [DOI] [PubMed] [Google Scholar]
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- Tettamanti G, Malagoli D, Benelli R, et al. Growth factors and chemokines: a comparative functional approach between invertebrates and vertebrates. Curr Med Chem. 2006;13:2737–2750. doi: 10.2174/092986706778521986. [DOI] [PubMed] [Google Scholar]
- Troncone L, De Lisa E, Bertapelle C, et al. Morphofunctional characterization and antibacterial activity of haemocytes from Octopus vulgaris. J Nat Hist. 2015;49:1457–1475. doi: 10.1080/00222933.2013.826830. [DOI] [Google Scholar]
- Van Beneden RJ. Molecular analysis of bivalve tumors: models for environmental/genetic interactions. Environ Health Perspect. 1994;102:81–83. doi: 10.1289/ehp.94102s1281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Beneden RJ, Gardner GR, Blake NJ, Blair DG. Implications for the presence of transforming genes in gonadal tumors in two bivalve mollusk species. Cancer Res. 1993;53:2976–2979. [PubMed] [Google Scholar]
- van der Merwe M, Auzoux-Bordenave S, Niesler C, Roodt-Wilding R. Investigating the establishment of primary cell culture from different abalone (Haliotis midae) tissues. Cytotechnology. 2010;62:265–277. doi: 10.1007/s10616-010-9293-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Kesteren RE, Smit AB, Dirks RW, et al. Evolution of the vasopressin/oxytocin superfamily: characterization of a cDNA encoding a vasopressin-related precursor, preproconopressin, from the mollusc Lymnaea stagnalis. Proc Natl Acad Sci USA. 1992;89:4593–4597. doi: 10.1073/pnas.89.10.4593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaziri H, Benchimol S. Reconstitution of telomerase activity in normal human cells leads to elongation of telomeres and extended replicative life span. Curr Biol. 1998;8:279–282. doi: 10.1016/s0960-9822(98)70109-5. [DOI] [PubMed] [Google Scholar]
- Walker C, Böttger SA, Mulkern J, et al. Mass culture and characterization of tumor cells from a naturally occurring invertebrate cancer model: applications for human and animal disease and environmental health. Biol Bull. 2009;216:23–39. doi: 10.1086/BBLv216n1p23. [DOI] [PubMed] [Google Scholar]
- Wang G, Li N, Zhang L, et al. IGFBP7 promotes hemocyte proliferation in small abalone Haliotis diversicolor, proved by dsRNA and cap mRNA exposure. Gene. 2015;571:65–70. doi: 10.1016/j.gene.2015.06.051. [DOI] [PubMed] [Google Scholar]
- Wen CM, Kou G-H, Chen S. Establishment of cell lines from the pacific oyster. Vitr Cell Dev Biol. 1993;29A:901–903. doi: 10.1007/BF02634224. [DOI] [PubMed] [Google Scholar]
- Wen CM, Kou GH, Chen SN. Cultivation of cells from the heart of the hard clam, Meretrix lusoria (Röding) J Tissue Cult Methods. 1993;15:123–130. doi: 10.1007/BF02388265. [DOI] [Google Scholar]
- Wheeler NJ, Dinguirard N, Marquez J, et al. Sequence and structural variation in the genome of the Biomphalaria glabrata embryonic (Bge) cell line. Parasit Vectors. 2018;11:496–507. doi: 10.1186/s13071-018-3059-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wijsman JWM, Troost K, Fang J, Roncarati A. Global production of marine bivalves. Trends and challenges. In: Smaal AC, Ferreira JG, Grant J, Petersen JK, Strand Ø, editors. Goods and services of marine bivalves. Cham: Springer; 2019. pp. 7–26. [Google Scholar]
- Wildering WC, Lodder JC, Kits KS, Bulloch AGM. Nerve growth factor ( NGF ) acutely enhances high-voltage- activated calcium currents in molluscan neurons. J Neurophysiol. 1995;74:2778–2781. doi: 10.1152/jn.1995.74.6.2778. [DOI] [PubMed] [Google Scholar]
- Xiang L, Kong W, Su J, et al. Amorphous calcium carbonate precipitation by cellular biomineralization in mantle cell cultures of Pinctada fucata. PLoS ONE. 2014;9:e113150. doi: 10.1371/journal.pone.0113150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu W, Vebrosky EN, Richards ML, Armbrust KL. Evaluation of dicloran phototoxicity using primary cardiomyocyte culture from Crassostrea virginica. Sci Total Environ. 2018;628–629:1–10. doi: 10.1016/j.scitotenv.2018.02.022. [DOI] [PubMed] [Google Scholar]
- Yamanaka S. Induction of pluripotent stem cells from mouse fibroblasts by four transcription factors. Cell Prolif. 2008;41:51–56. doi: 10.1111/j.1365-2184.2008.00493.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshino TP, Bickham U, Bayne CJ. Molluscan cells in culture: primary cell cultures and cell lines. Can J Zool. 2013;91:391–404. doi: 10.1139/cjz-2012-0258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshino TP, Laursen JR. Production of Schistosoma mansoni daughter sporocysts from mother sporocysts maintained in synxenic culture with Biomphalaria glabrata embryonic (Bge) cells. J Parasitol. 1995;81:714–722. doi: 10.2307/3283960. [DOI] [PubMed] [Google Scholar]
- Yoshino TP, Wu XJ, Di LH. Transfection and heat-inducible expression of molluscan promoter-luciferase reporter gene constructs in the Biomphalaria glabrata embryonic snail cell line. Am J Trop Med Hyg. 1998;59:414–420. doi: 10.4269/ajtmh.1998.59.414. [DOI] [PubMed] [Google Scholar]
- You Y, Huan P, Liu B. RNAi assay in primary cells: a new method for gene function analysis in marine bivalve. Mol Biol Rep. 2012;39:8209–8216. doi: 10.1007/s11033-012-1668-y. [DOI] [PubMed] [Google Scholar]
- You Y, Huan P, Wang X, Liu B. The potential roles of a laminin receptor in adhesion and apoptosis of cells of the marine bivalve Meretrix meretrix. PLoS ONE. 2012;7:e47104. doi: 10.1371/journal.pone.0047104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yue C, Yu H, Li H, et al. The endogenous EF-1α promoter is highly active in driving gene overexpression in developing embryos of the Pacific oyster Crassostrea gigas. Aquaculture. 2020;522:735134. doi: 10.1016/j.aquaculture.2020.735134. [DOI] [Google Scholar]
- Zweibaum J. Analyse histophysiologique de l’epithelium vibratile en état de survie in vitro. Compt Rend Soc Biol. 1925;93:785–787. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data generated or analyzed during this study are included in this published article.