

Abstract

Current research is focused on cancer treatments other than chemotherapy medications, particularly those derived from natural sources. The goal of this work was to look at the anticancer and biomarker properties of a methanolic extract of Annona squamosa leaves and their extract-loaded noisome. A. squamosa leaves extract and their leaves extract-loaded noisome were prepared. Transmission electron microscopy was used to screen the size of the niosomes loaded with the A. squamosa L. leaves extract. The tumor size, blood picture (hemoglobin, red blood cells, white blood cells), liver functions, kidney function, oxidative stress, and inflammatory markers were evaluated to assess the potential anticancer activity of the A. squamosa leaves extract and A. squamosa leaves extract-loaded noisome in Ehrlich ascites carcinoma. A. squamosa L. leaves extract was found to be an effective anticancer treatment. The protective effect of the loaded extract showed more significant results. All treated groups showed a lower tumor volume compared to the positive control. Liver and kidney functions were improved, and inflammatory markers were decreased. Oxidative stress was improved in tumor, liver, and kidney tissues. A. squamosa leaves contain major anticancer compounds that in general help most enzymes of the liver and kidney and other injured organs to return to their normal levels.
Introduction
Cancer is a significant global public health issue and has the second highest mortality rate in the United States, accounting for about 610.000 deaths in 2022.1 The use of natural plant elements as an alternative to chemotherapy or chemopreventive drugs is an effective strategy in the fight against cancer, especially with the possibility of targeting cancerous cells without healthy cells.2
Anonna squamosa is a midsize tree belonging to the Annonaceae family. In many places of the world, it grows abundantly: for example, in Africa (Egypt), Asia, Australia, and America. It is known by its various names, such as sugar apple and sweetsop in English, Sirafal in Hindi, and Matomoko in Kenya.3A. squamosa’s anticancer activity is selective for distinct cancer cells, as it has antiproliferative4 and antioxidant activity.3 Phytochemicals obtained from the plant, such as terpenoids, flavonoids, alkaloids, glycosides, and phenols, show subtle effects on body physiology and have the desired effects. They have an antioxidant ability that quenches the free radicals and therefore they act as cancer chemo-preventive agents. In comparison to synthetic chemotherapeutic agents, plant-based medicines are found to be safer due to their less-toxic side effects.5
Nanoparticles account for nanoscale applications, providing a high surface area and efficient interaction, and working as carriers for molecules.6,7 Nanoparticles can be selectively used to target the delivery of therapeutic drugs8 and decrease drug resistance by cancer cells.9 Niosomes are distinctive drug nanocarriers developed by non-ionic surfactants and cholesterol self-association in an aqueous solution.10 They can encapsulate hydrophilic and hydrophobic drugs with adequate efficiency. Niosomes are biocompatible, low-cost, biodegradable, and stable.11,12
Previous studies have tried to encapsulate the A. squamosa extract for cancer treatment. Fadholly et al., 201913 prepared chitosan nanoparticles-loaded A. squamosa leaves extract against human colon cancer (WiDr) cell lines. They showed potent cytotoxicity, increasing caspase-3 expression significantly and cell cycle arrest in the G2/M phase. According to Al-Nemari et al., 2022,14 the free A. squamosa leaf extract increased the expression of proliferative and apoptotic markers, which in turn enhanced the extract’s anticancer effect on breast cancer cell lines. Additionally, they conducted an in vivo investigation, and it was found that the treatment of rats with the extracts had an impact on the tumor size, proliferative, and apoptotic indicators.
The current study differs from earlier ones in that it first prepares an extract of A. squamosa leaves, loads it onto niosomes with active ingredients, evaluates the success of the loading and release of the extract, and then evaluates the effectiveness on tumor-bearing mice. This study investigated the antitumoral activity and biomarker properties of the methyl extract of A. squamosa leaves in the Ehrlich ascites carcinoma (EAC) model. Efficiency comparison between the A. squamosa leaves extract (ASLE) and A. squamosa leaves extract-loaded noisome (ASLELN) was done by determining the liver and kidney functions, inflammatory markers, blood picture, oxidative stress, and tumor size.
Results
Loading Efficiency
In the current work, niosomes exhibit a high loading efficiency for the ASL extract (93 ± 3%).
UV Absorption Spectrum
The ultraviolet (UV) absorption spectrum for the ASLE shows an absorption band at wavelength 200–300 nm, one at 400–500 nm, and a small peak at 700 nm, while the loaded niosomes show an enhancement for the peak at 700 nm and the band at 400–500 nm with a slight shift in the position due to the presence of the extract loaded in the niosome membrane (Figure 1).
Figure 1.

UV absorption spectrum of ASLE and ASLELN.
Transmission Electron Microscopy
Under a zooming of 200 nm, the transmission electron microscopy (TEM) image presented the indication that the nanoparticles were highly monodispersed without aggregation. The prepared nanoparticles were well isolated in a spherical shape with an average nanoparticle diameter of 220 nm (Figure 2).
Figure 2.

TEM images of the niosome loaded with the ASL extract.
In Vitro Extract Release
The burst discharge of free ASLE from the dialysis bag took 5 h to attain equilibrium. In the case of the ASLE loaded into the niosomes, the nano loading, besides its ability to save the extract from expected biodegradation in the body, promoted the sustained release of ASLE over 10 h (Figure 3). Niosomes are stable carriers for ASLE, and the slow release reinforces the therapeutic effect of the extract.
Figure 3.
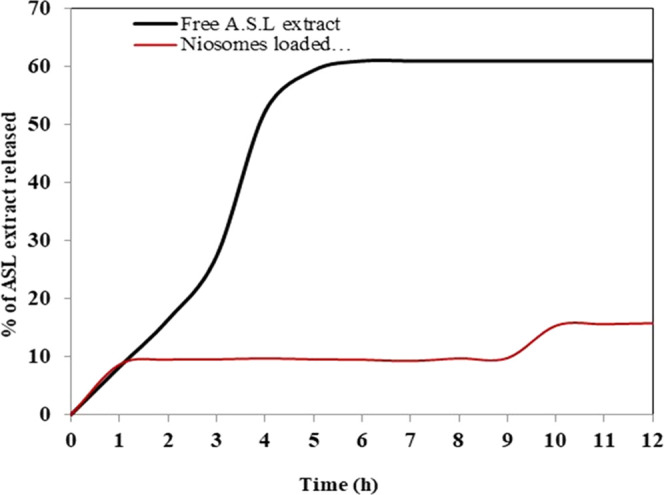
In vitro release curves of the free ASL extract and loaded ASL extract from the cellulose acetate dialysis bag.
Effect of Tested Extracts on the Ehrlich Tumor Size
Figure 4 shows the effect of ASLE and ASLELN on the development of Ehrlich’s volume compared to the Ehrlich solid tumor group (EST) not treated with any formulation. A significant decrease in tumor volume was detected in group 4, which was administered ASLELN before EST formation (Nano form + EST) by 59% (0.132 ± 0.011), and group 6, which was administered ASLELN after EST formation (EST + Nano form) by 63% (0.120 ± 0.008), compared to the EST control group (0.325 ± 0.016).
Figure 4.

Effect of ASLE and ASLELN on the development of Ehrlich’s volume. Each value indicates the mean ± SEM. #indicates significantly dissimilar from the EST control group at p < 0.05. Statistical analysis was performed by one-way ANOVA followed by Tukey multiple comparison test.
Hematological Parameters
The Ehrlich solid tumor (EST) group shows a considerable significant drop in Hb %, lymphocyte, and red blood cell (RBC) count by 29% (10.53 ± 0.29), 22% (53.00 ± 1.73), and 36% (3.30 ± 0.17), respectively, as compared with the normal group (14.90 ± 0.30, 68.17 ± 1.01, and 5.14 ± 0.07, respectively), accompanied by a significant increase in white blood cell (WBC) count and percentage of neutrophils and monocytes by 71% (10.87 ± 0.06), 31% (30.83 ± 0.41), and 144% (2.16 ± 0.14), respectively, as compared with the normal group (6.36 ± 0.18, 23.50 ± 0.86, and 0.89 ± 0.01, respectively). On the other hand, pre-administration of ASLELN in the Ehrlich inoculation group (Gp 4) and post-administration of ASLELN in the Ehrlich inoculation group (Gp 6) exhibited a significant increase in the percentage of Hb, lymphocyte, and RBC count by 28% (13.47 ± 0.29), 22% (64.53 ± 0.90), and 44% (4.74 ± 0.17), and by 20% (12.67 ± 0.16), 29% (68.37 ± 0.18), and 36% (4.49 ± 0.14), respectively, as compared with the Ehrlich group, accompanied by a significant decrease in WBC count and percentage of neutrophils and monocytes by 39% (6.60 ± 0.05), 15% (26.33 ± 0.72), and 35% (1.41 ± 0.06), respectively, and by 22% (8.50 ± 0.34), 27% (22.37 ± 1.38), and 55% (0.96 ± 0.03), respectively, as compared with the EST group (P < 0.05, Table 1).
Table 1. Effects of ASLE and ASLELN on the Hematological Parameters in EST-Bearing Micea.
| groups | Hb content (g%) | RBC (×106 (mm3)) | WBC (×106 (mm3)) | lymphocytes (%) | neutrophils (%) | monocytes (%) |
|---|---|---|---|---|---|---|
| normal | 14.40 ± 0.30 | 5.14 ± 0.07 | 6.36 ± 0.18 | 68.17 ± 1.01 | 23.50 ± 0.86 | 0.89 ± 0.01 |
| EST | 10.53b ± 0.29 | 3.30b ± 0.17 | 10.87b ± 0.06 | 53.00b ± 1.73 | 30.83b ± 0.41 | 2.16b ± 0.14 |
| ASLE pre-EST | 12.20b,c ± 0.15 | 4.08b,c ± 0.08 | 8.50b,c ± 0.28 | 59.67b,c ± 0.33 | 29.08b ± 1.08 | 1.80b,c ± 0.11 |
| ASLELN pre-EST | 13.47b,c ± 0.29 | 4.74c ± 0.17 | 6.60c ± 0.05 | 64.53c ± 0.90 | 26.33c ± 0.72 | 1.41c ± 0.06 |
| EST with ASLE | 12.33b,c ± 0.16 | 4.21b,c ± 0.11 | 8.50b,c ± 0.28 | 66.67c ± 1.33 | 28.90 ± 0.49 | 0.96b,c ± 0.03 |
| EST with ASLELN | 12.67b,c ± 0.16 | 4.49b,c ± 0.14 | 8.50b,c ± 0.34 | 68.37c ± 0.18 | 22.37c ± 1.38 | 0.94b,c ± 0.03 |
Each value indicates the mean ±SEM.
Significantly different from the normal group at p < 0.05.
Significantly different from the control EST group at p < 0.05. Statistical analysis was performed by one-way ANOVA followed by Tukey multiple comparison test.
Inflammatory Markers in Serum
Figure 5 shows a significant increase in the serum levels of COX-2, TNF-α, and NF-κB in the EST group by 4-fold (544.50 ± 23.40), 4.8-fold (315.80 ± 15.46), and 4.6-fold (3.16 ± 0.20), respectively, as compared to the normal group (132.00 ± 2.88, 65.87 ± 4.14, 0.69 ± 0.05). On the contrary, pre-administration of ASLELN in the Ehrlich inoculation group (Gp 4) and post-administration of ASLELN in the Ehrlich inoculation group (Gp 6) showed a significant decrease in the serum levels of COX-2, TNF-α, and NF-κB by 72% (151.70 ± 8.51), 72% (87.80 ± 3.15), respectively, and 72% (0.88 ± 0.07), and by 60% (217.00 ± 15.59), 53% (147.60 ± 11.66), and 49% (1.60 ± 0.11), respectively, as compared with the EST group.
Figure 5.
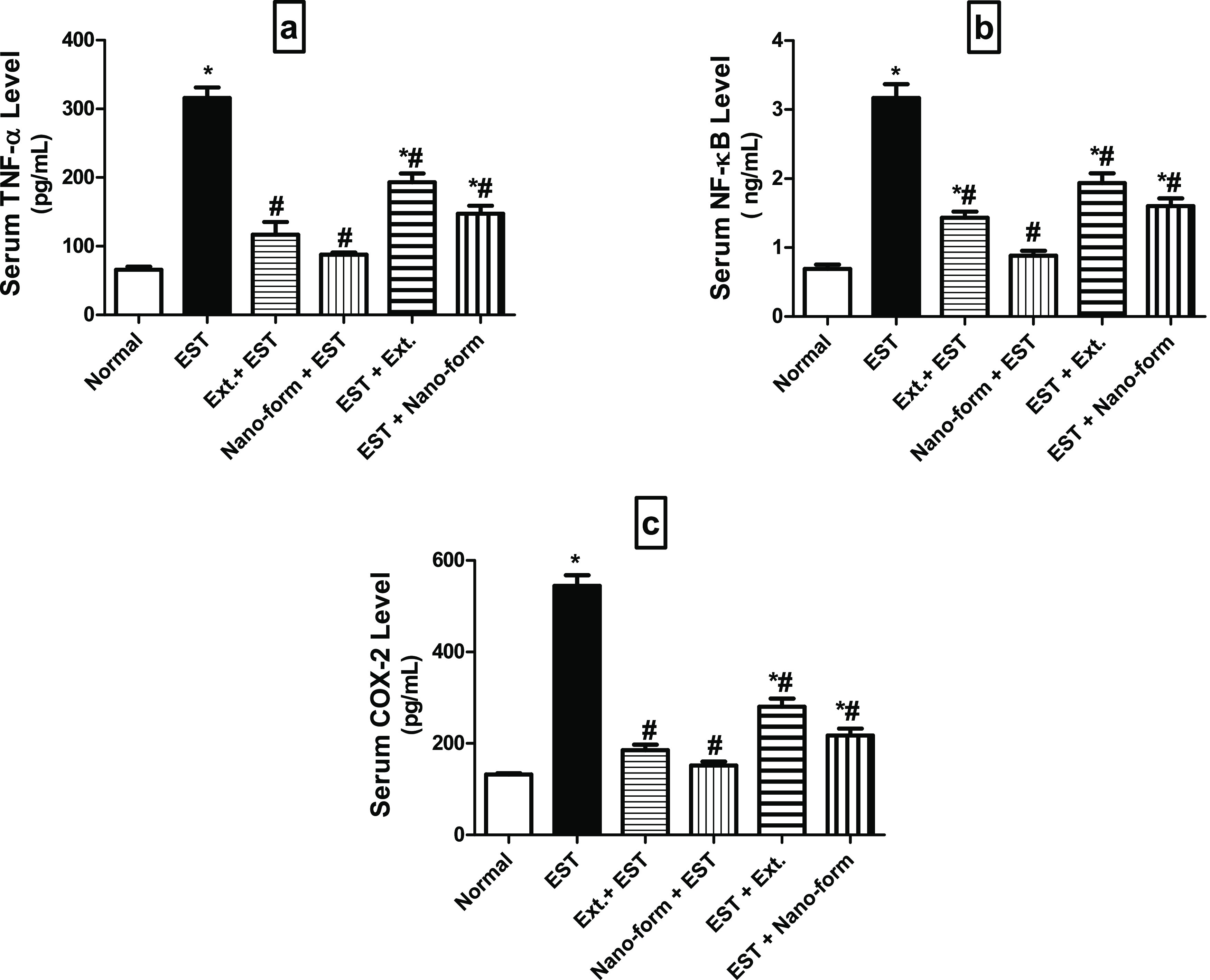
Levels of inflammatory markers (a) TNF-α, (b) NF-kB, and (c) COX-2 in the serum of different studied groups. Each value indicates the mean ± SEM. *Significantly different from the normal group at p < 0.05. #Significantly different from the control EST group at p < 0.05. Statistical analysis was performed by one-way ANOVA followed by Tukey multiple comparison test.
Serum Liver and Renal Functions
Renal Functions
Table 2 shows a considerable rise in the serum creatinine levels and urea in the EST group by 193 and 33%, respectively, in comparison to the normal group. The protected (Gp 4) and treated (Gp 6) groups with ASLELN displayed significantly decreased levels of creatinine and urea by 57 and 51%, respectively, and by 62 and 21%, respectively, as compared to the EST group.
Table 2. Effects of ASLE and ASLELN on Renal Markers in Mice Serum Bearing the Ehrlich Solid Tumor (EST)a.
| groups | creatinine (mg/dL) | urea (mg/dL) |
|---|---|---|
| normal | 0.56 ± 0.02 | 61.87 ± 1.74 |
| EST | 1.64b ± 0.06 | 82.15b ± 5.13 |
| ASLE pre-EST | 0.70c ± 0.06 | 49.46c ± 0.67 |
| ASLELN pre-EST | 0.69c ± 0.05 | 40.50b,c ± 3.50 |
| EST with ASLE | 0.76c ± 0.03 | 56.67c ± 2.71 |
| EST with ASLELN | 0.63c ± 0.03 | 64.49c ± 3.41 |
Each value indicates the mean ± SEM.
Significantly different from the normal group at p < 0.05.
Significantly different from the control EST group at p < 0.05. Statistical analysis was performed by one-way ANOVA followed by Tukey multiple comparison test.
Liver Functions
Table 3 shows a considerable rise in the serum levels of ALP, ALT, AST, and total bilirubin in the Ehrlich group by 33, 21, 86, and 27%, respectively, with a significant decrease in albumin by 26% in comparison to the normal group. The protected (Gp 4) and treated (Gp 6) groups with ASLELN displayed significantly decreased levels of ALT, AST, ALP, and total bilirubin by 20, 16, 36, and 17%, respectively, and by 20, 14, 33, and 18%, respectively, as compared to the EST group and, notably, an increase in albumin level by 28% in the protected group with ASLELN (Gp 4) only.
Table 3. Effects of Tested ASLE and ASLELN on Liver Functions in Mice Serum Bearing the Ehrlich Solid Tumor (EST)a.
| Groups | ALT (U/L) | AST (U/L) | ALP (U/L) | ALb (g/dL) | TP (g/dL) | total bilirubin (mg/dL) |
|---|---|---|---|---|---|---|
| normal | 17.75 ± 0.66 | 15.30 ± 0.10 | 48.23 ± 3.95 | 3.02 ± 0.05 | 6.80 ± 0.41 | 1.03 ± 0.01 |
| EST | 23.63b ± 0.57 | 18.50b ± 0.32 | 89.60b ± 3.98 | 2.24b ± 0.02 | 5.36 ± 0.18 | 1.31b ± 0.01 |
| ASLE pre-EST | 21.88c ± 0.33 | 16.53c ± 0.16 | 73.50b ± 4.60 | 2.59 ± 0.17 | 6.09 ± 0.19 | 1.19b,c ± 0.01 |
| ASLELN pre-EST | 18.88c ± 0.25 | 15.53c ± 0.20 | 57.40c ± 2.30 | 2.87c ± 0.18 | 6.78 ± 0.43 | 1.09c ± 0.02 |
| EST with ASLE | 19.25c ± 0.45 | 16.27c ± 0.53 | 65.27c ± 5.28 | 2.79 ± 0.15 | 6.72 ± 0.14 | 1.07c ± 0.03 |
| EST with ASLELN | 19.00c ± 0.33 | 15.93c ± 0.14 | 59.70c ± 4.60 | 2.78 ± 0.11 | 6.58 ± 0.35 | 1.07c ± 0.03 |
Each value indicates the mean ± SEM.
Significantly different from the normal group at p < 0.05.
Significantly different from the control EST group at p < 0.05. Statistical analysis was performed by one-way ANOVA followed by Tukey multiple comparison test.
Oxidative and Inflammatory Markers
Ehrlich Tumor Homogenate
As illustrated in Figure 6, the tumor reduced glutathione (GSH) content (a) and superoxide dismutase (SOD) activity (c) of the normal group was (13.39 ± 0.12 and 110.60 ± 2.78) and significantly decreased in the Ehrlich group by 27% (9.80 ± 0.18) and 26% (81.44 ± 2.60) respectively as compared with normal rats. Nevertheless, the protected (Gp 4) and treated (Gp 6) groups with ASLELN displayed significantly boost GSH, and SOD by 25% (12.29 ± 0.64), 30% (12.76 ± 0.33), and 33% (108.20 ± 3.05), 26% (102.40 ± 3.35) respectively as compared to EST group.
Figure 6.
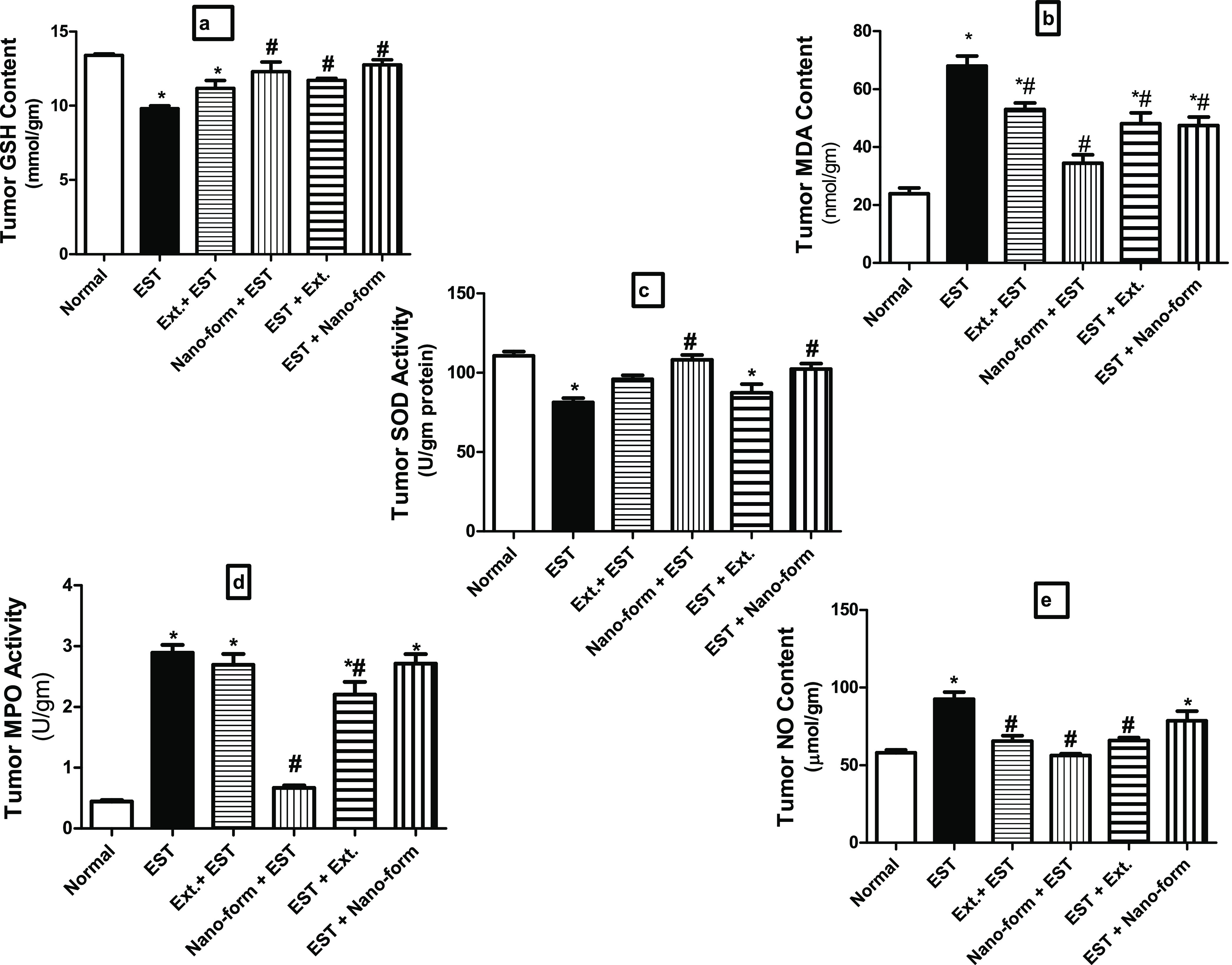
Levels of oxidant and antioxidant markers (a) GSH content, (b) MDA, (c) SOD activity, (d) MPO activity, and (e) NO content in tumor homogenate of different studied groups. Each value indicates the mean ± SEM. *Different significantly from the normal group at p < 0.05. #Different Significantly from the control EST group at p < 0.05. One way of exhibiting statistical analysis was performed by ANOVA followed by Tukey Multiple Comparison Test.
The tumor MDA (b), NO (e) contents, and MPO activity (d) of the normal group were (23.90 ± 1.96, 58.10 ± 1.67, 0.45 ± 0.02) respectively. They significantly increased in the EST group by 184% (67.98 ± 3.47), 59% (92.59 ± 4.63), and 542% (2.89 ± 0.13) respectively as compared with normal rats.
As compared with the EST group, the tumor MDA, NO, and MPO significantly decreased in the protected group with ASLELN (Gp 4) by 50% (2.60 ± 0.12), 39% (56.25 ± 1.20), 77% (0.67 ± 0.04) respectively but in the treated group with ASLELN (Gp 6), only MDA significantly decrease by 30% (47.43 ± 2.91) with no significant change in both NO and MPO.
Hepatic Homogenate
Table 4 showed a significant decrease in the hepatic GSH content and SOD activity in the Ehrlich group by 43 and 36% respectively with significant raise in hepatic MDA, NO content in companies with MPO activity by 150, 88, and 98% respectively in comparison to the normal group. while the protected (Gp 4) and treated (Gp 6) groups with ASLELN displayed significantly decreased MDA, NO, and MPO by 50, 30, 48% respectively, and by 47, 22, 43% respectively as compared to the untreated EST group plus notably increase in GSH and SOD by 57 and 42% respectivelyin the protected group with ASLELN (Gp 4) only.
Table 4. Effects of Tested ASLE and ASLELN on Hepatic Oxidative and Inflammatory Markers in Ehrlich Solid Tumor (EST)-Bearing Micea.
| groups | GSH (mmol/g tissue) | SOD (U/g protein) | MDA (nmol/g tissue) | NO (μmol/g tissue) | MPO (U/g tissue) |
|---|---|---|---|---|---|
| normal | 30.00 ± 1.92 | 138.80 ± 3.61 | 31.11 ± 2.03 | 24.08 ± 1.40 | 28.32 ± 1.76 |
| EST | 17.22b ± 0.55 | 89.19b ± 3.24 | 77.55b ± 2.31 | 45.37b ± 1.80 | 56.19b ± 1.59 |
| ASLE pre-EST | 23.70b,c ± 0.74 | 108.90b,c ± 2.30 | 44.69b,c ± 2.70 | 41.20b ± 0.61 | 48.96b ± 2.12 |
| ASLELN pre-EST | 27.04c ± 0.22 | 126.80c ± 2.09 | 38.84c ± 2.46 | 31.71b,c ± 1.61 | 29.10c ± 1.41 |
| EST with ASLE | 19.61b ± 0.45 | 104.50b ± 2.65 | 51.70b,c ± 3.47 | 36.11b,c ± 0.69 | 34.33c ± 1.87 |
| EST with ASLELN | 21.66b ± 0.96 | 112.60b ± 4.28 | 40.76c ± 3.38 | 35.41b,c ± 1.44 | 32.21c ± 0.93 |
Each value indicates the mean ± SEM.
Different significantly from the normal group at p < 0.05.
Different Significantly from the control EST group at p < 0.05. One way of exhibiting statistical analysis was performed by ANOVA followed by Tukey Multiple Comparison Test.
Renal Homogenate
Table 5 showed a significant decrease in the renal GSH content and SOD activity in the EST group by 34 and 34% respectively with significant raise in renal MDA, NO content in companies with MPO activity by 200, 227, and 137% respectively in comparison to the normal group. while the protected (Gp 4) and treated (Gp 6) groups with ASLELN displayed significantly decreased MDA, NO, and MPO by 53, 20, 49% respectively, and by 46, 22, and 39% respectively as compared to the untreated EST group plus notably increase in GSH and SOD by 42 and 30% respectively in the protected group with ASLELN (Gp 4) only.
Table 5. Effects of Tested ASLE and ASLELN on Renal Oxidative and Inflammatory Markers in Ehrlich Solid Tumor (EST)-Bearing Micea.
| groups | GSH (mmol/g tissue) | SOD (U/g protein) | MDA (nmol/g tissue) | NO (μmol/g tissue) | MPO (U/g tissue) |
|---|---|---|---|---|---|
| normal | 21.11 ± 0.55 | 128.40 ± 4.52 | 24.10 ± 1.34 | 18.15 ± 0.77 | 48.84 ± 3.21 |
| EST | 13.89b ± 0.55 | 84.50b ± 4.31 | 72.51b ± 1.88 | 59.32b ± 2.19 | 115.60b ± 5.51 |
| ASLE pre-EST | 15.89b ± 0.48 | 99.54b ± 1.99 | 41.79b,c ± 2.28 | 51.85b ± 2.13 | 110.80b ± 6.92 |
| ASLELN pre-EST | 19.75c ± 1.30 | 110.20c ± 7.56 | 34.17c ± 2.35 | 47.53b,c ± 1.63 | 58.40c ± 4.59 |
| EST with ASLE | 16.79b ± 0.99 | 96.85b ± 1.81 | 42.59b,c ± 3.44 | 50.00b ± 2.13 | 109.70b ± 5.40 |
| EST with ASLELN | 17.58 ± 0.88 | 103.00b ± 4.76 | 39.12b,c ± 1.47 | 46.30b,c ± 2.82 | 68.13c ± 3.85 |
Each value indicates the mean ± SEM.
Different significantly from the normal group at p < 0.05.
Different Significantly from the control EST group at p < 0.05. One way of exhibiting statistical analysis was performed by ANOVA followed by Tukey Multiple Comparison Test.
Histopathology
The histopathology of the liver, kidney, and tumor tissues in the mice is studied to reveal the treatment effect of free and ASLELN. As shown in Figure 7 Gp 1 has liver tissue with the normal architecture of hepatocytes around the central vein, kidney tissue with normal renal tissue, and skeletal muscle tissue with normal muscle fibers striation. Figure 7 Gp 2 showed liver tissue with congested central vein with infiltration of activated Kupffer′s cells, a glomerular tuft of the kidney with slightly hyperplastic glomerular cells, and skeletal muscles tissue with the invasion of the neoplastic cells to the surrounding skeletal muscles with marked necrosis (H&E, 50 μm). Figure 7 Gp 3 has hepatic tissue of central vein with infiltration of activated Kupffer′s cells, the obliteration of Bowman’s space in the kidney tissue with the hyperplastic glomerular cells with dilatation and congestion of renal blood vessel and muscle bundles with the invading neoplastic cells diffusing all around its adjacent tissue with marked necrosis. Gp 4 has hepatic tissue with infiltration of activated Kupffer′s cells between hepatocytes, kidney tissue with the obliteration of Bowman’s space with the hyperplastic glomerular cells with dilatation and congestion of renal blood vessel, and the neoplastic cells diffusing all around its adjacent tissue with necrosis of skeletal muscles. Gp 5 showed liver with congested central veins with an activated Kupffer′s cells infiltrating in between, kidney with the obliteration of Bowman’s space with the hyperplastic glomerular cells with dilatation and congestion of renal blood vessel and muscle bundles with the invading neoplastic cells diffusing all around its adjacent tissue with marked necrosis. Gp 6 showed the liver with the normal architecture of the hepatic lobules, normal renal tissue architecture, and muscle bundles showing the invading neoplastic cells diffusing all around its adjacent tissue with marked necrosis.
Figure 7.
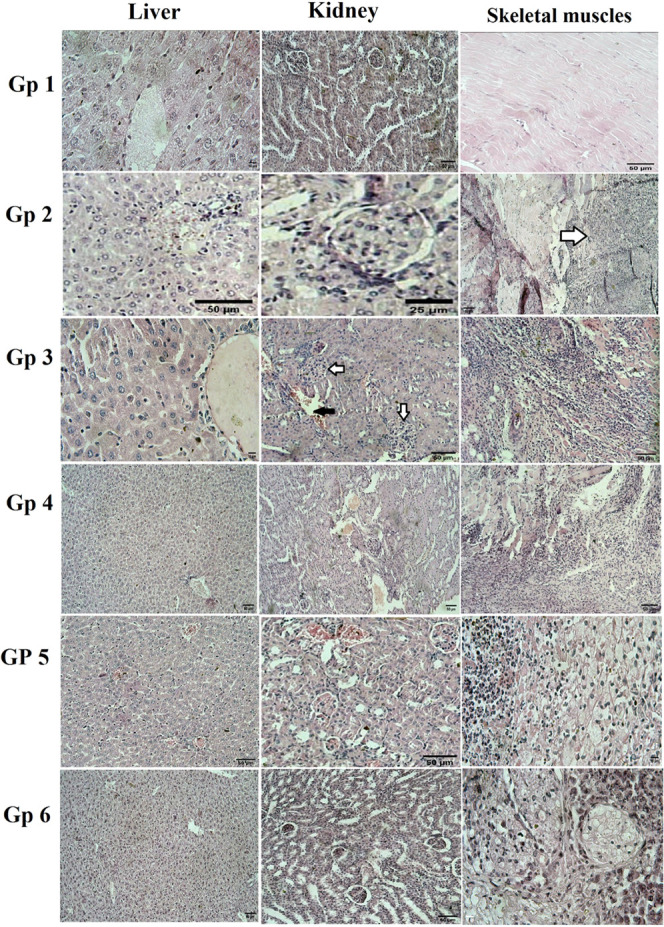
Histopathology photomicrographs for mic (liver, kidney, and skeletal muscles) tissues. Group 1 showed: liver tissue with the normal architecture of hepatocytes around the central vein (H&E, 10 μm), kidney tissue with normal renal tissue (H&E, 50 μm), skeletal muscles tissue with normal muscle fibers striation (H&E, 50 μm). Group 2 showed: liver tissue congested central vein with infiltration of activated Kupffer′s cells (H&E, 50 μm), a glomerular tuft of the kidney with slightly hyperplastic glomerular cells (H&E, 25 μm), skeletal muscles tissue with the invasion of the neoplastic cells to the surrounding skeletal muscles with marked necrosis (H&E, 50 μm). Group 3 showed: hepatic tissue of the central vein with infiltration of activated Kupffer′s cells (H&E, 10 μm), the obliteration of Bowman’s space in the kidney tissue with the hyperplastic glomerular cells (white arrows) with dilatation and congestion of renal blood vessel (black arrow) (H&E, 50 μm), muscle bundles with the invading neoplastic cells diffusing all around its adjacent tissue with marked necrosis (H&E, 50 μm). Group 4 showed: hepatic tissue with infiltration of activated Kupffer′s cells between hepatocytes (H&E, 50 μm), kidney have the obliteration of Bowman’s space with the hyperplastic glomerular cells with dilatation and congestion of renal blood vessel (H&E, 50 μm), the neoplastic cells diffusing all around its adjacent tissue with necrosis of skeletal muscles (H&E, 50 μm). Group 5 showed: that the liver has congested central veins with an activated Kupffer′s cells infiltrating in between (H&E, 50 μm), kidney with the obliteration of Bowman’s space with the hyperplastic glomerular cells with dilatation and congestion of renal blood vessel (H&E, 50 μm), muscle bundles with the invading neoplastic cells diffusing all around its adjacent tissue with marked necrosis (H&E, 10 μm). Group 6 showed: Image of the liver with the normal architecture of the hepatic lobules (H&E, 50 μm), Normal renal tissue architecture (H&E, 50 μm), and muscle bundles showing the invading neoplastic cells diffusing all around its adjacent tissue with marked necrosis (H&E, 10 μm).
Discussion
The challenging goal of cancer research continuously is discovering safe and effective new natural and synthetic products with anti-cancer effects.15 Plant-derived natural products have incredible potential to control cancer cell growth.16 Previous studies confirmed that the ASLE contained flavonoids, alkaloids, saponins, tannins, glycosides, steroids, phenols, and various acetogenin compounds and demonstrated a various array of pharmacological activities involving antioxidant, anti-cancer, and anti-inflammatory properties.17,18 Nowadays, plants are performing a remarkable role in nanoparticle synthesis.19 To ameliorate cancer therapeutic protocols, the current study was implemented to determine the antitumor efficacy and hepato-renal protective effect of ASLELN against Erhlich Ascites Carcinoma (EAC) in mice.
Figure 3 showed the release profile for niosomes loaded ASLE could be described as a two-step release process, involving an initial burst release (first hour), which could be due to diffusion of the compounds adsorbed on the niosomes surface, followed by subsequent slower release (after 10 h) and this could be attributed to the diffusion of A. squamosa leaves extract dispersed into the niosomes inner layers.20,21
Ehrlich tumor is a type of cancer that grows progressively quickly with a very destructive effect that can evolve in almost all strains of mice. Nevertheless, it is commonly used as an experimental animal model for a tumor because of its moderate cost, easy induction, effective reproducibility, and availability to assess the safety and efficacy of anticancer therapies.22
Reactive oxygen species (ROS) and other free radicals are the most potent mediators of cellular toxicity, particularly in cancer.23 Several studies have shown that tumor growth results in the disturbance of the antioxidant system and the production of lipid peroxidation in pivotal organs of the tumor-host.24 In external stress and carcinogenesis the reduction in the endogenous antioxidant system as SOD activity, GSH content is associated with the increase in MDA level which is supposed to be an inhibitor of protective enzymes,25 consequently, it could have both mutagenic and carcinogenic consequences.26 Our results were in harmony with previous studies and confirmed that EST–bearing mice had higher levels of oxidative stress than normal mice, as indicated by increased lipid peroxidation products (MDA), decreased GSH level, and SOD activity in target tissues.27 The formation of reactive oxygen species in cancer tissues causes lipid peroxidation, which leads to an increase in MDA levels, accompanied by the decline in the quantity of GSH content due to its use by the excessive number of free radicals created in the pathogenic state.28 In addition, loss of Mn-SOD activity or neutralization of superoxide in tumor cells and the decrease of mitochondrial number results in the decline in total SOD activity in the liver tissue of tumor-bearing mice.29
Naturally, occurring antioxidant supplementation can be effectively restored redox homeostasis by neutralizing and removing excess oxidants.30 So, administration of ASLE or ASLELN reversed the effect of EAC cells, by increasing SOD activity, and GSH content, and attenuating the EAC—mediated increase of MDA levels, demonstrating the ability of the extracts in reducing oxidative stress in tumor-bearing mice by modulating antioxidant levels. The reduction of oxidative stress in treated EST-bearing mice may be due to the antioxidant compounds content of these extracts as polyphenolic compounds and flavonoids as quercetin, that increase and have the free radical scavenging property,31 confirming the antioxidant activity of the examined extracts especially ASLELN in the group (4). These results were in concurrence with previous studies.32,33
Chronic inflammation has been linked to the onset and progression of several types of cancer.34 Inflammatory biomarkers such as TNF-α, COX-2, iNOS, and NF-kB are involved in enhancing all stages of tumor progression.35,36 Upregulation of these inflammatory biomarkers during chronic inflammation creates a proper environment for malignant cells to proliferate violently during tumor growth.37 Oxidative stresses can trigger inflammatory responses through the activation of NF-KB signaling, which is the master organizer for most inflammatory signaling cascades, including TNF-α, COX-2, and transcription of many inflammatory mediators.38,39 TNF-α can entice COX-2, resulting in enhanced PGE2 synthesis which promotes tumor progression through the reactive oxygen species pathway.40 In our investigation, EST–bearing mice significantly showed a marked increase in TNF-α, COX-2, and NF-KB levels as compared to the normal group. These results are consistent with a previous study.41 The increase in serum TNF-α level may be attributed to the increase in macrophage-produced ROS, that increased lipid peroxidation which prompted immune response and induced the transcriptional factor NF-KB.42 Also, inflammation induces degradation of the inhibitory protein (IkB) that connects to the inactivated form of NF-kB in the cytoplasm, resulting in the liberation of NF-KB to the nucleus and increasing inflammatory-related gene production.43 According to our study, treatment with ASLE or ASLELN was able to reduce the inflammation state in EST-bearing mice by decreasing NF-KB, TNF-α, and COX-2 levels, restoring the level of these markers to normal levels in Gp 4. Therefore, the tumor inhibitory activity of these extracts could be ascribed to the anti-inflammatory activity of their phytochemical contents as polyphenols, alkaloids, and flavonoids as quercetin which is involved in delaying cancer progression.44 It was reported that the most important antioxidant mechanism of polyphenols is the regulation of enzyme activity, as inhibition of oxidases such as cyclooxygenase results in a reduction in the production of ROS in vivo.45 In addition, flavonoid derivatives such as quercetin can exert multiple pharmacological activities including anti-inflammatory activity by minimizing the inflammatory factors production as NF-kB, TNF-α, and COX-2 in Human Colon Cancer Caco-2 Cells.46
Nitric oxide and the pro-oxidant enzyme myeloperoxidase (MPO) were playing a fundamental role in controlling cellular stress and constituting a key component of the innate immune defense system.47 Excessive production of NO and MPO–derived oxidants have been associated with tissue damage as well as the onset and the progression of chronic inflammation-related disorders including cancer involved in EAC-induced toxicity, this agrees with our findings that demonstrated a significant increase in NO level and MPO activity in the selected organs. Our results are in line with ELAblack et al. (2020)48 that explained the increase of NO level may be due to the elevation of lipid peroxidation levels and its product MDA that can enhance host cells, fundamentally macrophages/ monocytes, to produce NO by stimulation of iNOS activity causing tissue damage. Also, overproduction of TNF-α may be the reason for the elevation of NO concentration, where TNF-α acts as a mediator of NO synthesis.49 Besides that, the elevation of MPO activity may be attributed to the high levels of ROS that are produced in several human tumors.50
On the other hand, our results showed the anti-inflammatory activity of ASLE and ASLELN by decreasing NO and MPO levels, especially in Gp (4) get better-ameliorated effects. This may be due to the variety of bioactive properties of the polyphenolic compounds and flavonoids as quercetin content of these extracts. One of the most important antioxidant mechanisms of polyphenols is the regulation of enzyme activity, as inhibition of oxidases such as myeloperoxidase results in a reduction in the production of ROS and organic hydroperoxides in vivo.44 Quercetin improves inflammation by suppressing either the production of inflammatory mediators as NO produced after activation of iNOS expression or the expression and release of cytokines such as TNF-α.51 This result was in line with Kalidindi et al. (2015),17 who reported that aqueous and organic leaf extracts of A. squamosa inhibit the effect of NO-induced in vitro which is attributed to the presence of flavonoids and phenols contents. our results agree with Hendawy et al., (2019)52 who showed that administration of ethanolic and aqueous extracts of A. squamosa extract decreased NF-kB, and NO levels in the brain of AlCl3 treated rats.
The best criteria for judging the efficacy of any anticancer agents are the reduction of solid tumor volume and the vanishing of leukemic cells from the blood. Previous studies reported that the perturbation in the inflammatory regulators such as TNF-α and NO resulted in tumor proliferation and growth, which was elucidated by the increase in the solid tumor volume.53 The current study showed that the inoculation of EAC cells caused a significant increase in solid tumor volume due to cell multiplicity and the development of EAC in the EST-bearing group.4,54 Pre or with the administration of ASLE or ASLELN caused a distinct reduction in the solid tumor volume, with a more pronounced decrease in the treated EST-bearing mice with ASLELN. These results elucidated the antitumor efficacy of the extracts, particularly nanoforms that might be credited to the presence of flavonoid and phenolic compounds which may activate macrophages and inhibit vascular permeability.55 It may be pointed out that, the presence of annonaceous acetogenins compounds as a component of A. squamosa leaves extract56 may contribute to the antitumor activity of these extracts.57 Our results are consistent with Wang et al. (2014)58 who reported that ethyl acetate extract of A. squamosa leaves had anticancer activity on colon cancer and human epidermoid carcinoma cell line.
The tumor growth was accompanied by hematological changes which encompass a decrease in RBCs count, hemoglobin content, and an increase in WBC count, neutrophils, and monocytes. The correlation between anemia and cancer progression was reported in epidemiological studies.59 The main reason for this anemia is iron deficiency which causes the reduction of hemoglobin content of RBCs.60 The elevation of WBC count in EST-bearing mice may be attributed to the activation of bone marrow.61 ASLEL or ASLELN supplementation especially group (4) maintained the normal values of WBC and RBC count and replenishes hemoglobin content, which proposes its hematopoietic protecting action. These results are following the previous study.41 Manthou et al. (2017)62 explained that the presence of flavonoids and polyphenolic compounds increased antioxidant capacity in erythrocytes which prevents oxidation of hemoglobin and RBCs destruction by reactive species. The anti-inflammatory and antioxidant activity of the extracts could explain their hematopoietic activity.18
The vital functions of organs can be inversely affected by tumor growth, regardless of the tumor site.63 Cancer causes liver and kidney dysfunction and interferes in hepatic cell metabolism resulting in alterations in serum enzyme activity.64 In our investigation, elevated serum levels of ALP, ALT, AST, total bilirubin plus serum creatinine and urea levels, with a concomitant decrease in albumin elucidated a development of hepatoxicity and renal tissue injury in EST-bearing mice. Hepatocytes are destroyed then the excess AST released into the bloodstream following the growth of the tumor may cause an elevation in liver enzymes.65 Besides that, the decline in the level of albumin produced by the liver’s hepatocytes in EST–bearing mice, valuable markers for liver function,66 refers to the harmful effect of EAC that causes the decrease in the biosynthetic capacity of the liver to synthesize plasma protein. Also, EAC proliferation induced renal tissue injury and raised kidney function.67 Our results are matched with a recent study.68 The changes in liver and kidney functions could be due to the generation of ROS that induces tissue damage by destroying cellular homeostasis.69 Our data indicated that treatment of EST–bearing mice with ASLE or ASLELN ameliorated the changes in the levels of the hepatotoxicity markers and kidney functions indicating the potent hepato-renal protective effect of these extracts through its scavenging and antioxidant characteristics due to its flavonoid and phenolic compounds contents.70 These findings were consistent with previous results.32,71,72
Conclusions
A. squamosa leaf extracts could aid in the creation of new pharmaceuticals as a source of alternative chemotherapy for sarcoma cancer treatments. As niosomes nanocarriers are cheap, non-toxic, biocompatible, and release drugs with proper control, so this study loads A. squamosa leaves extract onto niosomes to raise its therapeutic efficacy in cancer-infected mice. A. squamosa leaves extract-loaded niosomes and free extract reduced tumor size and improved liver and kidney functions, inflammatory markers, oxidative stress, and hematological parameters relative to positive control. There was no discernible difference between the effectiveness of these two tested formulations in terms of the levels of biochemical markers, however, the extract-loaded niosomes were more effective than the free extract at shrinking tumors. Therefore, in addition to cancer patients, Annona squamosa leaf extracts can be recommended to individuals who have high-risk factors for developing cancer. The molecular mechanisms that A. squamosa leaf extracts target cancer cells require further study.
Experimental Section
Chemicals
Fisher Scientific U.K. provided Methyl alcohol. Phosphate buffer tablets pH 7.4 (purity >98%) was purchased from Bio Shop Canada Inc. Absolute Ethanol, Tween 80 (purity ≥99%), and cholesterol (Mw 386.65, purity ≥99.7%) were purchased from Sigma Aldrich.
Plant Materials and Preparations
Plant Material
Fresh plant materials were collected (March 2021) from a local plant farm at Mansoriya, Giza, Egypt. Plant taxonomists identified and authenticated the herb as A. squamosa L. at the Herbarium, Desert Research Center, Cairo, Egypt. Fresh leaves of the plant were washed with running tap water to clean, shade-dried at room temperature for 7days, and were finely ground using an electric blender. The obtained powder was stored in a dry closed container and kept at −20 °C in the dark until used.10
Plant Extraction
About 770 g of A. squamosa leaves (ASL) powder were weighed accurately and soaked in 1500 mL of 70% methanol and then kept in a dark place for 3 days, carbon dioxide was released frequently. The resulting aqueous fraction was filtered through filter paper. The filtrate was concentrated under vacuum by a rotary evaporator at about 40 °C. The concentrated A. squamosa leaves extract (ASLE) was collected and stored to use later.
Niosomes Preparation and Characterization
Preparation
Ethanol was used to dissolve tween, cholesterol, and the extract in a rounded flask, then the ethanol was evaporated at 45 °C to form dried thin layers on the flask inside the wall. Niosomes were formed by hydration of dried thin layers with phosphate-buffered saline (PBS) (pH 7.4), then subjected to sonication (WUC-A03H, Daihan, Korea) for 5 min to produce small niosomes vesicles. A cooling centrifuge (11,000 rpm for 30 min) (vision scientific 18,000 M, Korea) was used to precipitate niosomes.3
Characterization
Loading Efficiency
Different known concentrations of ASL extract were evaluated by UV–visible spectrophotometry (JENWAY Scanning, 6405, United Kingdom) at a wavelength of 270 nm. ASL extract standardization curve was created by plotting absorbance versus concentration of ASL extract. The initial concentration was evaluated for ASL extract before loading in niosomes. The efficiency of the A. squamosa leaves extract-loaded noisome (ASLELN) was achieved by the centrifugation technique.73 The loaded niosomes suspended in PBS were centrifuged (11,000 rpm, 30 min, VS-18000 M, Korea) to isolate the free extract from the niosomes. The isolated free ASL extract concentration was measured spectrophotometrically using the standard curve to get the final concentration. Then the loading efficiency of niosomes for ASL extract was evaluated from the next equation
UV Spectrum
The absorption of the ASLE and ASLELN was determined by a UV spectrophotometer over the range of 200–1000 nm to confirm the loading process.
Transmission Electron Microscope
The transmission electron microscope (JEOL JEM-1230, Japan) was used to illustrate the niosomes structure. The accelerating voltage was 102 kV to perform this evaluation. Niosomes suspension-loaded by the extract was negatively stained by 1% of phosphotungstic acid. Samples were incubated in grids of perforated carbon-coated for 10 min after the dray had been running for 5 min.3
In Vitro Extract Release
The dialysis technique was used to determine the ASL extract release from niosomes in PBS (pH 7.4). Briefly, 3 ml of ASLELN were placed in a dialysis bag (cellulose acetate Spectra/Por, MWCO-12,000, Spectrum, Canada) and were dived in 100 mL phosphate buffer saline containing 1 mL tween 20 to enhance the extract solubility on the magnetic stirrer (50 rpm, TK22-Kartell, Italy). Every 1 h, the absorbance of 2 mL of immersing solution was measured by UV–visible spectrophotometry at 270 nm.
Animals
In total, 72 male albino mice (weighing 25 ± 2 g) were obtained from the National Research Centre, Cairo, Egypt. Mice were kept in mesh steel wire cages with adjustable temperature controls set at (22 ± 3) °C, light (dark/light cycles every 12 h), and (80 ± 4) % relative humidity, as well as free access to food (diet and water). All procedures were carried out according to the research ethics committee for experimental studies at Cairo University Institutional Animal Care and Use Committee (CU-IACUC), based on verification of application number CU/I/F/27/21.
Tumor Inoculation
Ehrlich ascites carcinoma (EAC) cells were obtained from EAC-bearing mice provided by Cairo University’s Egyptian National Cancer Institute (NCI). 0.2 mL of ascetic fluid from EAC mice were aspirated and diluted in physiological buffer saline. EAC cells were counted before being subcutaneously injected into the left thigh of the mouse’s lower limb to induce a solid tumor in the experimental mice.
Experimental Design
One week before the experiments, all mice were kept in a normal laboratory environment for acclimatization, and then they were divided randomly into six equal groups (12 mice per group) as follows:
Group 1: Normal mice without any treatment and served as a negative control group.
Group 2 (EST): Mice were injected subcutaneously with about 2.5 to 3 × 106 EAC cells/mouse diluted in physiological saline.54 Mice were incubated for 24 h, then received saline orally for 24 successive days, and served as a positive control group (EST-bearing group).
Group 3 (Extract + EST): Before the injection of EAC cells, mice received ASLE orally once daily (250 mg/kg) for 1 week. The oral administration of ASLE was subsequently continued for an additional 24 days (ASLE pre-EST).
Group 4 (Nanoform + EST): Before the injection of EAC cells, mice were given ASLELN orally once daily for 1 week (250 mg/kg), following which the oral administration of ASLELN was continued for an additional 24 days (ASLELN pre-EST).
Group 5 (EST + Extract): Mice were injected with EAC cells, incubated for 24 h, and then given ASLE orally once daily (250 mg/kg) for 24 successive days. (EST with ASLE).
Group 6 (EST + Nanoform): Mice were injected with EAC cells, incubated for 24 h, and then given an oral dose of ASLELN once daily (250 mg/kg) for 24 successive days (EST with ASLELN).
At the end of the experiment, 24 h after the last manipulation and overnight fasting, samples of the blood were obtained and collected from mice’s retro-orbital venous plexus into two separate tubes; the first tube contained EDTA to quantify the hematological indices, the second was clean centrifuge tube and the blood left to coagulate for 20 min at 37 °C then centrifuged (for 20 min at 3000 rpm) to separate serum. The serum was stored (at −80 °C) for further analysis. The biochemical markers used in our analysis were albumin, urea, creatinine, total bilirubin, total protein, ALT, ALP, and AST in addition to other inflammatory markers such as (TNF-α, COX-2, and NF-kB). Thereafter, animals were euthanized by decapitation. Solid tumors, the right thigh of the lower limb, liver, and kidney from the control and the treated groups were excised and washed with cold sterile isotonic saline. To create the 10% homogenate that will be utilized for the evaluation of oxidative stress biomarkers (GSH, SOD, MDA) and inflammatory indicators (MPO, NO), solid tumors, liver, and kidneys were weighed and homogenized in ice-cold PBS. Some fragments from the normal muscles, tumor, liver, and kidneys were kept in 10% neutral buffered formalin for histopathological investigation while the other specimens were stored at −80 °C for biochemical analysis.
Determination of Solid Tumor Volume
Tumor growth was regularly measured twice a week for 2 successive weeks from the 11th day after the tumor induction. The length and width of tumors were measured by vernier calipers. The volume of the tumor mass (V) was calculated by using the following equation
74 Where (a) is the length of the major tumor axis, and the height and width are equal to the minor axis (b). The constant value (0.4) represents the linear regression coefficient.
Biochemical Analysis
Hematological Parameters
A fresh blood sample (0.25 mL) was added to 20 mg EDTA to investigate the hematological parameters such as Hb (Hemoglobin) content, RBCs, WBCs, and the percentage values of differential white cell count (Neutrophils, Lymphocytes, and monocytes).
Determination of Serum Kidney and Liver Functions
The separated serum was used to evaluate ALP (alkaline phosphatases), ALT (alanine aminotransferase), AST (aspartate aminotransferase) activities, total bilirubin, TP (total protein), Alb (albumin), urea, and creatinine levels by using commercially available kits obtained from Biodiagnostic company (Giza, Egypt).
Estimation of Inflammatory Biomarkers Markers
Using an in vitro enzyme-linked immunosorbent assay (ELISA) kit, the levels of Tumor necrosis factor-α (TNF- α; Abbexa, U.K.), nuclear factor-kappa (NF-κB; Sunlong Biotech Co., Ltd., Zhejiang, China), and cyclooxygenase-2(COX-2; Bio Vision) were assayed in serum.
Estimation of the Inflammatory and Oxidative Stress Parameters
Malondialdehyde (MDA) was evaluated based on Uchiyama and Mihara’s (1978) method.75 Reduced glutathione (GSH) was measured.76 The activity of superoxide dismutase (SOD) was determined according to the method of Nandi and Chatterjee (1988).77 Determination of myeloperoxidase (MPO) activity was assessed according to the method of Bradley et al. (1982).78 Nitric oxide (NO) level was measured according to the method of Miranda et al. (2001).79 Total protein was determined according to the method of Lowery et al. (1951).80
Histopathological Examination
Following necropsy, the liver, kidneys, and Ehrlich solid tumors (EST) were immediately removed and preserved in 10% buffered neutral formalin for histological analysis. Paraffin-embedded sections of the solid EAC tumor, right thigh muscles of normal control, liver, and kidneys were sliced into a thickness of (4–5 μm) and stained with hematoxylin and eosin (H&E) solution and examined under a light microscope.81
Statistical Analysis
All values in vivo results were presented as mean ± standard error of the mean (SEM). To evaluate the significance of the difference between treatments, one-way ANOVA was used, followed by the Tukey–Kramer multiple comparisons test. Values of P < 0.05 were considered significant. GraphPad PRISM program was used for statistical analysis (version 5, San Diego, CA).
Glossary
Abbreviations
- ASL
A. squamosa leaves
- ASLE
A. squamosa leaves extract
- ASLELN
A. squamosa leaves extract-loaded niosomes
- EAC
Ehrlich ascites carcinoma
- EST
Ehrlich solid tumor
The authors declare no competing financial interest.
References
- Siegel R. L.; Miller K. D.; Fuchs H. E.; Jemal A. Cancer Statistics, 2022. Ca-Cancer J. Clin. 2022, 72, 7–33. 10.3322/caac.21708. [DOI] [PubMed] [Google Scholar]
- Wu J.; Zhou T.; Wang Y.; Jiang Y.; Wang Y. Mechanisms and Advances in Anti-Ovarian Cancer with Natural Plants Component. Molecules 2021, 26, 5949 10.3390/molecules26195949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abd-Elghany A. A.; Mohamad E. A. Ex-vivo transdermal delivery of Annona squamosa entrapped in niosomes by electroporation. J. Radiat. Res. Appl. Sci. 2020, 13, 164–173. 10.1080/16878507.2020.1719329. [DOI] [Google Scholar]
- Pardhasaradhi B.; Reddy M.; Ali A. M.; Kumari A. L.; Khar A. Antitumour activity of Annona squamosa seed extracts is through the generation of free radicals and induction of apoptosis. Indian J. Biochem. Biophys. 2004, 41, 167–172. [PubMed] [Google Scholar]
- Wang Z.; Li Z.; Ye Y.; Xie L.; Li W. Oxidative stress and liver cancer: etiology and therapeutic targets. Oxid. Med. Cell. Longevity 2016, 2016, 7891574 10.1155/2016/7891574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohamad E. A.; Rageh M. M.; Darwish M. M. A sunscreen nanoparticles polymer based on prolonged period of protection. J. Bioact. Compat. Polym. 2022, 37, 17–27. 10.1177/08839115211061741. [DOI] [Google Scholar]
- Li H.; Gao A.; Jiang N.; Liu Q.; Liang B.; Li R.; Zhang E.; Li Z.; Zhu H. Protective effect of curcumin against acute ultraviolet B irradiation induced photo damage. Photochem. Photobiol. 2016, 92 (6), 808–815. 10.1111/php.12628. [DOI] [PubMed] [Google Scholar]
- Fahmy H. M.; Idris A. M. R.; Elsayed A. A.; Mohamad E. A. Electroporation-enhanced entrapment of diclofenac sodium and ascorbic acid into DPPC liposomes. Res. J. Biotechnol. 2021, 16, 19–26. 10.25303/1611rjbt1926. [DOI] [Google Scholar]
- Abd-Elghany A. A.; Mohamad E. A. Antitumor impact of iron oxide nanoparticles in Ehrlich carcinoma-bearing mice. J. Radiat. Res. Appl. Sci. 2021, 14, 314–321. 10.1080/16878507.2021.1957398. [DOI] [Google Scholar]
- Mohamad E. A.; Fahmy H. M. Niosomes and liposomes as promising carriers for dermal delivery of Annona squamosa extract. Braz. J. Pharm. Sci. 2020, 56, 1–8. 10.1590/s2175-97902019000318096. [DOI] [Google Scholar]
- Abd-Elghany A. A.; Mohamad E. A.; El-Sakhawy M. A.; Mansouri S.; Ismail S. H.; Elneklawi M. S. Enhancement of mechanical properties of chitosan film by doping with sage extract-loaded niosomes. Mater. Res. Express 2022, 9, 035006 10.1088/2053-1591/ac600a. [DOI] [Google Scholar]
- Mohamad E. A.; Mohamed Z. N.; Hussein M. A.; Elneklawi M. S. GANE can improve Lung fibrosis by reducing inflammation via promoting p38MAPK/TGF-β1/NF-κB singling pathway down-regulation. ACS Omega 2022, 7, 3109–3312. 10.1021/acsomega.1c06591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fadholly A.; Proboningrat A.; Iskandar R. P. D.; Rantam F. A.; Sudjarwo S. A. In vitro anticancer activity Annona squamosa extract nanoparticle on WiDr cells. J. Adv. Pharm. Technol. Res. 2019, 10, 149–154. 10.4103/japtr.JAPTR_10_19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Nemari R.; Bacha A. B.; Al-Senaidy A.; Almutairi M. H.; Arafah M.; Al-Saran H.; Abutaha N.; Semlali A. Cytotoxic effects of Annona squamosa leaves against breast cancer cells via apoptotic signaling proteins. J. King Saud Univ. - Sci. 2022, 34, 102013 10.1016/j.jksus.2022.102013. [DOI] [Google Scholar]
- Abdo J. A.; Alsharif F. M.; Salah N.; Elkhawaga O. A. Y. Cytotoxic Effect of Propolis Nanoparticles on Ehrlich Ascites Carcinoma Bearing Mice. Adv. Nanopart. 2019, 08, 55–70. 10.4236/anp.2019.84005. [DOI] [Google Scholar]
- Ramos-Silva A.; Tavares-Carreón F.; Figueroa M.; la Torre-Zavala D.; Gastelum-Arellanez A.; Rodríguez-García A.; Galán-Wong L. J.; Avilés-Arnaut H. Anticancer potential of Thevetia peruviana fruit methanolic extract. BMC Complementary Altern. Med. 2017, 17, 241 10.1186/s12906-017-1727-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalidindi N.; Thimmaiah N. V.; Jagadeesh N. V.; Nandeep R.; Swetha S.; Kalidindi B. Antifungal and antioxidant activities of organic and aqueous extracts of Annona squamosa Linn. leaves. J. Food Drug Anal. 2015, 23, 795–802. 10.1016/j.jfda.2015.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Nemari R.; Al-Senaidy A.; Semlali A.; Ismael M.; Badjah-Hadj-Ahmed A. Y.; Ben Bacha A. GC-MS profiling and assessment of antioxidant, antibacterial, and anticancer properties of extracts of Annona squamosa L. leaves. BMC Complementary Med. Ther. 2020, 20, 296 10.1186/s12906-020-03029-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sayed H. M.; Said M. M.; Morcos N. Y.; El Gawish M. A.; Ismail A. F. Antitumor and Radiosensitizing Effects of Zinc Oxide-Caffeic Acid Nanoparticles against Solid Ehrlich Carcinoma in Female Mice. Integr. Cancer Ther. 2021, 20, 15347354211021920 10.1177/15347354211021920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosseini S. F.; Zandi M.; Rezaei M.; Farahmand F. Two-Step Method for Encapsulation of Oregano Essential Oil in Chitosan Nanoparticles: Preparation, Characterization and in Vitro Release Study. Carbohydr. Polym. 2013, 95, 50–56. 10.1016/j.carbpol.2013.02.031. [DOI] [PubMed] [Google Scholar]
- Keawchaoon L.; Yoksan R. Preparation, Characterization and in Vitro Release Study of Carvacrol-Loaded Chitosan Nanoparticles. Colloids Surf., B 2011, 84, 163–171. 10.1016/j.colsurfb.2010.12.031. [DOI] [PubMed] [Google Scholar]
- Abdel-Rahman M. N.; Kabel A. M. Comparative study between the effect of methotrexate and valproic acid on solid Ehrlich tumor. J. Egypt. Natl. Cancer Inst. 2012, 24, 161–167. 10.1016/j.jnci.2012.08.001. [DOI] [PubMed] [Google Scholar]
- Ghosh S.; Mukherjee S.; Choudhury S.; Gupta P.; Adhikary A.; Baral R.; Chattopadhyay S. Reactive oxygen species in the tumor niche trigger altered activation of macrophages and immunosuppression: role of fluoxetine. Cell. Signalling 2015, 27, 1398–1412. 10.1016/j.cellsig.2015.03.013. [DOI] [PubMed] [Google Scholar]
- Mohamed H. R. H.; Amer M.; El Faky A. S. A. Growth retardation and apoptotic death of tumor cells by Artemisia herba-alba oral administration in Ehrlich solid carcinoma bearing mice. Rev. Bras. Farmacogn. 2019, 29, 763–772. 10.1016/j.bjp.2019.06.007. [DOI] [Google Scholar]
- Mohamad E. A.; Elfky A. A.; El-Gebaly R. H.; Afify A. Study the change in the mosquito larvae (Culex pipiens) in water treated with short pulses of electric field. Electromagn. Biol. Med. 2022, 41, 80–92. 10.1080/15368378.2021.2012787. [DOI] [PubMed] [Google Scholar]
- Roy S.; Kumari D.; Chakraborty M.; Haldar P. K. Anti Proliferative Activity Of Calamus Rotang As A Spotlight On Ehrlich’s Ascites Carcinoma Treated Peritoneal As Well As Solid Tumor Model. Int. J. Pharm. Pharm. Sci. 2018, 85–90. 10.22159/ijpps.2018v10i1.21449. [DOI] [Google Scholar]
- El-Far M.; Salah N.; Essam A.; El-Azim A. O. A.; El-Sherbiny I. M. Silymarin nanoformulation as a potential anticancer agent in experimental Ehrlich ascites carcinoma-bearing animals. Nanomedicine 2018, 13, 1865–1858. 10.2217/nnm-2017-0394. [DOI] [PubMed] [Google Scholar]
- Gangopadhyay A.; Chakraborty M.; Haldar P. K.; Chaulya N. C.; Ghosh A. Evaluation of the anticancer activity of ethanolic extract of cyperus kyllingia endl. In ehrlich ascites carcinoma-induced swiss albino mice. Asian J. Pharm. Clin. Res. 2018, 11, 482–485. 10.22159/ajpcr.2018.v11i10.27926. [DOI] [Google Scholar]
- Lee W.-L.; Huang J.-Y.; Shyur L.-F. Phytoagents for cancer management: regulation of nucleic acid oxidation, ROS, and related mechanisms. Oxid. Med. Cell. Longevity 2013, 2013, 1–22. 10.1155/2013/925804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yılmaz S.; Göçmen A. Y.; Karataş E.; Tokpınar A. Cornus Mas L improves Antioxidant Status in the Liver, Lung, Kidney, Testis, and Brain of Ehrlich Ascites Tumor Bearing Mice. Asian Pac. J. Cancer Prev. 2020, 21, 2531–2537. 10.31557/APJCP.2020.21.9.2531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panda S.; Kar A. Antidiabetic and antioxidative effects of Annona squamosa leaves are possibly mediated through quercetin-3-O-glucoside. Biofactors 2007, 31, 201–210. 10.1002/biof.5520310307. [DOI] [PubMed] [Google Scholar]
- Hassan A.; Mohammed L.; Abd ElMoneim M.; Abd ElBaky A. Hepatic and Renal Protective Effects of Annona muricata Leaf and Fruit Extracts on Ehrlich Ascites Carcinoma in Mice. Zagazig Vet. J. 2019, 47, 234–247. 10.21608/zvjz.2019.12593.1040. [DOI] [Google Scholar]
- Rogovskii V. S. The linkage between inflammation and immune tolerance: interfering with inflammation in cancer. Curr. Cancer Drug Targets 2017, 17, 325–332. 10.2174/1568009617666170109110816. [DOI] [PubMed] [Google Scholar]
- DiDonato J. A.; Mercurio F.; Karin M. NF-κB and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. 10.1111/j.1600-065X.2012.01099.x. [DOI] [PubMed] [Google Scholar]
- Mohamed E. T.; Safwat G. M. Evaluation of cardioprotective activity of Lepidium sativum seed powder in albino rats treated with 5-fluorouracil. Beni-Suef Univ. J. Basic Appl. Sci. 2016, 5, 208–215. 10.1016/j.bjbas.2016.05.001. [DOI] [Google Scholar]
- Greten F. R.; Grivennikov S. I. Inflammation, and cancer: triggers, mechanisms, and consequences. Immunity 2019, 51, 27–41. 10.1016/j.immuni.2019.06.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nafees S.; Rashid S.; Ali N.; Hasan S. K.; Sultana S. Rutin ameliorates cyclophosphamide-induced oxidative stress and inflammation in Wistar rats: role of NFκB/MAPK pathway. Chem.-Biol. Interact. 2015, 231, 98–107. 10.1016/j.cbi.2015.02.021. [DOI] [PubMed] [Google Scholar]
- Benzer F.; Kandemir F. M.; Kucukler S.; Comaklı S.; Caglayan C. Chemoprotective effects of curcumin on doxorubicin-induced nephrotoxicity in Wistar rats: by modulating inflammatory cytokines, apoptosis, oxidative stress, and oxidative DNA damage. Arch. Physiol. Biochem. 2018, 124, 448–457. 10.1080/13813455.2017.1422766. [DOI] [PubMed] [Google Scholar]
- Liu B.; Qu L.; Yan S. Cyclooxygenase-2 promotes tumor growth and suppresses tumor immunity. Cancer Cell Int. 2015, 15, 106 10.1186/s12935-015-0260-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salem A. Z.; Medhat D.; Fathy S. A.; Mohamed M. R.; El-Khayat Z.; El-Daly S. M. Indole glucosinolates exhibit anti-inflammatory effects on Ehrlich ascites carcinoma cells through modulation of inflammatory markers and miRNAs. Mol. Biol. Rep. 2021, 48, 6845–6855. 10.1007/s11033-021-06683-5. [DOI] [PubMed] [Google Scholar]
- Khater S. I.; Ezz-Eldin R.; Saad S.; Gamal F.; El-Abed E.; Arisha A. Dox capsulated chitosan nanoparticles effect as anti-carcinogenic therapeutic agent in mice with Ehrlich carcinoma. J. Anim. Health Prod. 2020, 9, 110–120. 10.17582/journal.jahp/2020/9.s1.110.120. [DOI] [Google Scholar]
- Hayden M. S.; Ghosh S. NF-κB in immunobiology. Cell Res. 2011, 21, 223–244. 10.1038/cr.2011.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibrahim R. Y.; Hassan A. I.; Al-Adham E. K. The anti-ulcerative colitis effects of Annona squamosa Linn. leaf aqueous extract in experimental animal model. Int. J. Clin. Exp. Med. 2015, 8, 21861–21870. [PMC free article] [PubMed] [Google Scholar]
- Ibrahim A. Y.; El-Newry S. A.; El-Raey M. A. Evaluation of the antioxidant, anti-inflammatory, and antitumor properties of Sabal grown in Egypt. Egypt. Pharm. J. 2017, 16, 168–183. 10.4103/epj.epj_23_17. [DOI] [Google Scholar]
- Han M.; Song Y.; Zhang X. Quercetin suppresses the migration and invasion in human colon cancer Caco-2 cells through regulating toll-like receptor 4/nuclear factor-kappa B pathway. Pharmacogn. Mag. 2016, 12, 237–244. 10.4103/0973-1296.182154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devi D. G.; Cibin T.; Ramaiah D.; Abraham A. Bis (3, 5-diiodo-2, 4, 6-trihydroxyphenyl) squaraine: A novel candidate in photodynamic therapy for skin cancer models in vivo. J. Photochem. Photobiol., B 2008, 92, 153–159. 10.1016/j.jphotobiol.2008.06.002. [DOI] [PubMed] [Google Scholar]
- Giridasappa A.; Rangappa D.; Maheswarappa G.; Marilingaiah N. R.; Thammaiah C.; Shareef I.; Subbegowda R. K.; Shivaramu P. D. Phytofabrication of cupric oxide nanoparticles using Simarouba glauca and Celastrus paniculatus extracts and their enhanced apoptotic inducing and anticancer effects. Appl. Nanosci. 2021, 11, 1393–1409. 10.1007/s13204-021-01753-3. [DOI] [Google Scholar]
- ELAblack F. Z.; Elgazzar U.; Hashem A. Antitumor activity of novel azole compound against Ehrlich ascites carcinoma in Swiss albino mice. Insight Chem. Biochem. 2020, 1, 1–8. 10.33552/ICBC.2020.01.000510. [DOI] [Google Scholar]
- Azab K. S.; El Fatih N. M.; El Tawill G.; El Bakary N. M. Pro-apoptotic and anti-neoplastic impact of luteolin on solid Ehrlich carcinoma. bearing mice exposed to gamma radiation. J. Cancer Res. Ther. 2020, 16, 1506–1516. 10.4103/jcrt.JCRT_1116_19. [DOI] [PubMed] [Google Scholar]
- Reuter S.; Gupta S. C.; Chaturvedi M. M.; Aggarwal B. B. Oxidative stress, inflammation, and cancer: how are they linked?. Free Radical Biol. Med. 2010, 49, 1603–1616. 10.1016/j.freeradbiomed.2010.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Begum R.; Sharma M.; Pillai K.; Aeri V.; Sheliya M. A. Inhibitory effect of Careya Arborea on inflammatory biomarkers in carrageenan-induced inflammation. Pharm. Biol. 2015, 53, 437–445. 10.3109/13880209.2014.923005. [DOI] [PubMed] [Google Scholar]
- Hendawy O. M.; ELBana M. A.; Abdelmawlla H. A.; Maliyakkal N.; Hedeab G. M. Effect of Annona squamosa Ethanolic and Aqueous Leave Extracts on Aluminum Chloride-Induced Neuroinflammation in Albino Rats. Biomed. Pharmacol. J. 2019, 12, 1723–1730. 10.13005/bpj/1801. [DOI] [Google Scholar]
- El Bakary N. M.; Alsharkawy A. Z.; Shouaib Z. A.; Barakat E. M. Role of bee venom and melittin on restraining angiogenesis and metastasis in γ-irradiated solid Ehrlich carcinoma-bearing mice. Integr. Cancer Ther. 2020, 19, 1534735420944476 10.1177/1534735420944476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aldubayan M. A.; Elgharabawy R. M.; Ahmed A. S.; Tousson E. Antineoplastic activity and curative role of avenanthramides against the growth of Ehrlich solid tumors in mice. Oxid. Med. Cell. Longevity 2019, 2019, 1–12. 10.1155/2019/5162687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samudrala P. K.; Augustine B. B.; Kasala E. R.; Bodduluru L. N.; Barua C.; Lahkar M. Evaluation of antitumor activity and antioxidant status of Alternanthera brasiliana against Ehrlich ascites carcinoma in Swiss albino mice. Pharmacogn. Res. 2015, 7, 66–73. 10.4103/0974-8490.147211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma C.; Chen Y.; Chen J.; Li X.; Chen Y. A review on Annona squamosa L.: phytochemicals and biological activities. Am. J. Chin. Med. 2017, 45, 933–964. 10.1142/S0192415X17500501. [DOI] [PubMed] [Google Scholar]
- Chen Y.; Chen J.-w.; Zhai J.-h.; Wang Y.; Wang S.-l.; Li X. Antitumor activity and toxicity relationship of annonaceous acetogenins. Food Chem. Toxicol. 2013, 58, 394–400. 10.1016/j.fct.2013.05.028. [DOI] [PubMed] [Google Scholar]
- Wang D.-S.; Rizwani G. H.; Guo H.; Ahmed M.; Ahmed M.; Hassan S. Z.; Hassan A.; Chen Z.-S.; Xu R.-H. Annona squamosa Linn: cytotoxic activity found in leaf extract against human tumor cell lines. Pak. J. Pharm. Sci. 2014, 27, 1559–1563. [PubMed] [Google Scholar]
- Muñoz M.; Gómez-Ramírez S.; Martín-Montañez E.; Auerbach M. Perioperative anemia management in colorectal cancer patients: a pragmatic approach. World J. Gastroenterol. 2014, 20, 1972–1985. 10.3748/wjg.v20.i8.1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharya S.; Prasanna A.; Majumdar P.; Kumar R. S.; Haldar P. K. Antitumor efficacy and amelioration of oxidative stress by Trichosanthes dioica root against Ehrlich ascites carcinoma in mice. Pharm. Biol. 2011, 49, 927–935. 10.3109/13880209.2011.557080. [DOI] [PubMed] [Google Scholar]
- Kim S.; Kim B.; Song Y. S. Ascites modulates cancer cell behavior, contributing to tumor heterogeneity in ovarian cancer. Cancer Sci. 2016, 107, 1173–1178. 10.1111/cas.12987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manthou E.; Georgakouli K.; Deli C. K.; Sotiropoulos A.; Fatouros I. G.; Kouretas D.; Haroutounian S.; Matthaiou C.; Koutedakis Y.; Jamurtas A. Z. Effect of pomegranate juice consumption on biochemical parameters and complete blood count. Exp. Ther. Med. 2017, 14, 1756–1762. 10.3892/etm.2017.4690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta N.; Goswami B.; Mittal P. Effect of standard anthracycline-based neoadjuvant chemotherapy on circulating levels of serum IL-6 in patients of locally advanced carcinoma breast–a prospective study. Int. J. Surg. 2012, 10, 638–640. 10.1016/j.ijsu.2012.11.007. [DOI] [PubMed] [Google Scholar]
- Dolai N.; Karmakar I.; Kumar R. S.; Kar B.; Bala A.; Haldar P. K. Evaluation of antitumor activity and in vivo antioxidant status of Anthocephalus cadamba on Ehrlich ascites carcinoma treated mice. J. Ethnopharmacol. 2012, 142, 865–870. 10.1016/j.jep.2012.05.050. [DOI] [PubMed] [Google Scholar]
- Gressner O. A.; Weiskirchen R.; Gressner A. M. Biomarkers of liver fibrosis: clinical translation of molecular pathogenesis or based on liver-dependent malfunction tests. Clin. Chim. Acta 2007, 381, 107–113. 10.1016/j.cca.2007.02.038. [DOI] [PubMed] [Google Scholar]
- Saravanan R.; Viswanathan P.; Pugalendi K. V. Protective effect of ursolic acid on ethanol-mediated experimental liver damage in rats. Life Sci. 2006, 78, 713–718. 10.1016/j.lfs.2005.05.060. [DOI] [PubMed] [Google Scholar]
- Badr O. M.; Abd-Eltawab H. M.; Sakr S. A. Ameliorative effect of ginger extract against pathological alterations induced in mice bearing solid tumors. J. Biosci. Appl. Res. 2016, 2, 185–196. 10.21608/jbaar.2016.106941. [DOI] [Google Scholar]
- Abd Eldaim M. A.; Tousson E.; Soliman M. M.; El Sayed I. E. T.; Aleem A. A. H. A.; Elsharkawy H. N. Grape seed extract ameliorated Ehrlich solid tumor-induced hepatic tissue and DNA damage with reduction of PCNA and P53 protein expression in mice. Environ. Sci. Pollut. Res. 2021, 28, 44226–44238. 10.1007/s11356-021-13904-8. [DOI] [PubMed] [Google Scholar]
- Medhat D.; Hussein J.; El-Naggar M. E.; Attia M. F.; Anwar M.; Latif Y. A.; Booles H. F.; Morsy S.; Farrag A. R.; Khalil W. K.; El-Khayat Z. Effect of Au-dextran NPs as anti-tumor agent against EAC and solid tumor in mice by biochemical evaluations and histopathological investigations. Biomed. Pharmacother. 2017, 91, 1006–1016. 10.1016/j.biopha.2017.05.043. [DOI] [PubMed] [Google Scholar]
- Ibrahim F.; Jaber A.; Ibrahim G.; Cheble E. Antioxidant activity and total phenol content of different plant parts of Lebanese Annona squamosa Linn. Int. J. Pharm. Pharm. Sci. 2020, 100–105. 10.22159/ijpps.2020v12i8.36992. [DOI] [Google Scholar]
- Rajeshkumar S.; Tamilarasan B.; Sivakumar V. Phytochemical screening and hepatoprotective efficacy of leaves extracts of Annona squamosa against paracetamol-induced liver Toxicity in rats. Int. J. Pharmacogn. 2015, 22, 178–185. [Google Scholar]
- Amokeoja F.; Sani E.; Giwa F.; Garuba T.; Abdulraheem A. O.; Sulaiman F. Assessment of the effects of ethanolic extracts of Annona squamosa leaves and stem on selected biochemical parameters. Biosci. Res. J. 2021, 32, 105–122. [Google Scholar]
- Bendas E. R.; Tadros M. I. Enhanced transdermal delivery of salbutamol sulfate via ethosomes. AAPS PharmsciTech 2007, 8, 213–220. 10.1208/pt0804107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Attia M. A.; Weiss D. W. Immunology of spontaneous mammary carcinomas in mice: V. Acquired tumor resistance and enhancement in strain A mice infected with mammary tumor virus. Cancer Res. 1966, 26, 1787–1800. [PubMed] [Google Scholar]
- Uchiyama M.; Mihara M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. 10.1016/0003-2697(78)90342-1. [DOI] [PubMed] [Google Scholar]
- Bolkent S.; Yanardag R.; Karabulut-Bulan O.; Yesilyaprak B. Protective role of Melissa officinalis L. extract on liver of hyperlipidemic rats: a morphological and biochemical study. J. Ethnopharmacol. 2005, 99, 391–398. 10.1016/j.jep.2005.02.038. [DOI] [PubMed] [Google Scholar]
- Nandi A.; Chatterjee I. Assay of superoxide dismutase activity in animal tissues. J. Biosci. 1988, 13, 305–315. 10.1007/BF02712155. [DOI] [Google Scholar]
- Bradley P. P.; Priebat D. A.; Christensen R. D.; Rothstein G. Measurement of cutaneous inflammation: estimation of neutrophil content with an enzyme marker. J. Invest. Dermatol. 1982, 78, 206–209. 10.1111/1523-1747.ep12506462. [DOI] [PubMed] [Google Scholar]
- Miranda K. M.; Espey M. G.; Wink D. A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric oxide 2001, 5, 62–71. 10.1006/niox.2000.0319. [DOI] [PubMed] [Google Scholar]
- Lowry O.; Rosebrough N.; Farr A. L.; Randall R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. 10.1016/S0021-9258(19)52451-6. [DOI] [PubMed] [Google Scholar]
- Rageh M. M.; El-Garhy M. R.; Mohamad E. A. Magnetic fields enhance the anti-tumor efficacy of low dose cisplatin and reduce the nephrotoxicity. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1475–1485. 10.1007/s00210-020-01855-9. [DOI] [PubMed] [Google Scholar]