

Abbreviations
- CHKα
choline kinase α
- CSC
cancer stem‐like cell
- G‐6‐P
glucose‐6‐phosphate
- GBM
glioblastoma
- HK
hexokinase
- KHKA
ketohexokinase isoform A
- mRNA
messenger RNA
- NF‐κB
Nuclear factor kappa‐light‐chain‐enhancer of activated B cells
- Oct4
octamer‐binding transcription factor 4
- PCK1
phosphoenolpyruvate carboxykinase 1
- PD‐1
programmed cell death 1
- PD‐L1
programmed cell death ligand 1
- PGK1
phosphoglycerate kinase 1
- PKM2
pyruvate kinase M2
- SCLC
small cell lung cancer
- USP11
ubiquitin‐specific protease 11
- VDAC
voltage‐dependent anion channel
1.
The Warburg effect is a key metabolic feature of cancer cells and manifests as substantially increased glycolysis regardless of the presence of oxygen [1]. The initial step of glycolysis is catalyzed by hexokinase (HK), which converts glucose to the metabolic intermediate glucose‐6‐phosphate (G‐6‐P). Four kinds of HK isozymes, HK1, HK2, HK3 and HK4, have been found in mammals. HK1 and HK2 bind to the mitochondrial outer membrane through interactions with voltage‐dependent anion channels (VDACs), and both of these molecules have a high affinity for glucose [2]. HK2 overexpression is frequently detected in cancer cells, leading to enhanced aerobic glycolysis and tumorigenesis that renders HK2 an attractive target for cancer treatment [3]. Important advances in research on cancer metabolism in recent decades have revealed that metabolic enzymes have non‐metabolic functions, which play pivotal roles in tumor development and progression [4, 5, 6]. Recent studies uncovered that HK2 directly promoted tumor immune evasion through the activity of a previously unidentified protein kinase and tumor cell stemness via upregulation of CD133 expression.
A study by Guo et al. [7] published in Cell Metabolism delineated a moonlighting function of HK2 as a glucose sensor intrinsically connecting the Warburg effect to tumor immune evasion. The tumor cell membrane protein programmed cell death ligand 1 (PD‐L1) specifically binds to programmed cell death 1 (PD‐1) on T cells, thereby inhibiting the functions of CD8+ T cells and promoting tumor cell immune evasion. Tumor cells under high glucose conditions exhibited substantially increased CD274 mRNA expression (encoding PD‐L1), which was abolished by HK2 depletion, indicating the indispensable role of HK2 in PD‐L1 expression. Treatment of glioblastoma (GBM) cells with G‐6‐P, deletion of the HK2 N‐terminal 15 amino acids, which are critical for the association of HK2 with the mitochondria, or treatment of GBM cells with a peptide containing the HK2 N‐terminal 15 amino acids promoted the translocation of HK2 from the mitochondria to the cytosol and resulted in an interaction between HK2 and cytosolic IκBα. Importantly, a protein phosphorylation assay demonstrated that HK2 acted as a protein kinase and phosphorylated IκBα at T291 independent of the HK2 ability to bind glucose.
HK2‐phosphorylated IκBα binds to μ‐calpain protease, leading to IκBα degradation and release of the inhibition of IκBα on nuclear factor kappa‐light‐chain‐enhancer of activated B cells (NF‐κB). Activated NF‐κB translocates into the nucleus and induces CD274 transcription and PD‐L1 expression. Knock‐in expression of IκBα T291A by CRISPR/Cas9 genome‐editing technology blocked high glucose‐induced nuclear translocation of NF‐κB and expression of CD274 messenger RNA (mRNA) and PD‐L1. HK2‐N15 peptide treatment of GBM cells reduced the production of IFNγ from cocultured primary CD8+ T cells. This inhibition was largely alleviated by the knock‐in expression of IκBα T291A in GBM cells. Mouse studies showed that IκBα T291A expression strongly reduced brain tumor growth accompanied by reduced PD‐L1 expression and increased CD8+ T‐cell infiltration into the tumor tissue and granzyme B expression. Importantly, combined treatment with the HK2 inhibitor lonidamine and an anti‐PD‐1 antibody achieved a stronger and additive effect on tumor growth inhibition and mouse survival time prolongation with substantially enhanced CD8+ T‐cell infiltration and granzyme B expression in tumors. Immunohistochemistry analyses showed that the levels of HK2 expression, IκBα T291 phosphorylation and PD‐L1 expression are positively correlated with each other in human GBM specimens and predict GBM patient prognosis [7]. These findings demonstrated that HK2 acts as a glucose sensor, sensing intracellular glucose levels, to intrinsically integrate energy metabolism and tumor immunity regulation and underscore the pivotal role of the Warburg effect in tumor immune evasion.
HK2 also has a noncanonical function directly promoting tumor cell stemness [8]. Wang et al. [8] demonstrated that HK2 expression is much higher in CD133+ cancer stem‐like cells (CSCs) of small cell lung cancer (SCLC) than in differentiated CD133− cells and that HK2 expression correlates with poor prognosis of SCLC patients. HK2 depletion inhibited CSC stemness and promoted CSC differentiation, accompanied by reduced CD133 expression. Cell fractionation and coimmunoprecipitation analyses showed that HK2 interacted with CD133 in non‐mitochondrial fractions but not in mitochondrial fractions. Consistently, deletion of the HK2 N‐terminal mitochondrial binding domain increased CD133 expression. Mass spectrometry analyses showed that the deubiquitinase ubiquitin‐specific protease 11 (USP11) is a CD133‐associated protein and that the interaction between USP11 and CD133 was increased in spheroid CSCs compared to differentiated monolayer cells. Importantly, HK2 expression in CSCs promoted the binding of USP11 to CD133, thereby reducing CD133 polyubiquitylation and enhancing its expression.
CD133 expression was shown to promote cancer cell stemness, and CD133+ CSCs of SCLC are highly tumorigenic [9]. HK2 overexpression enhanced the expression of the pluripotency transcription factor octamer‐binding transcription factor 4 (Oct4) and the translation reprogramming factor Lin28, as well as tumor growth, and this increase was largely abrogated by depletion of USP11. However, this abrogation was rescued by the ectopic expression of CD133. In contrast, HK2 depletion inhibited tumor growth, and this inhibition was alleviated by CD133 overexpression [8]. These findings indicated that HK2 promotes USP11‐mediated CD133 expression and tumor growth and first revealed that enhanced glycolysis promotes CSC stemness via a noncanonical function of HK2.
Similar to the metabolic enzymes pyruvate kinase M2 (PKM2) [10], phosphoglycerate kinase 1 (PGK1), ketohexokinase isoform A (KHKA), phosphoenolpyruvate carboxykinase 1 (PCK1) and choline kinase α (CHKα) [4], which exhibit protein kinase activity, non‐mitochondrial HK2 can also act as a protein kinase to promote NF‐κB‐mediated PD‐L1 expression under aerobic glycolysis conditions. In CSCs, non‐mitochondrial HK2 maintains cell stemness by stabilizing CD133 expression (Figure 1).
FIGURE 1.
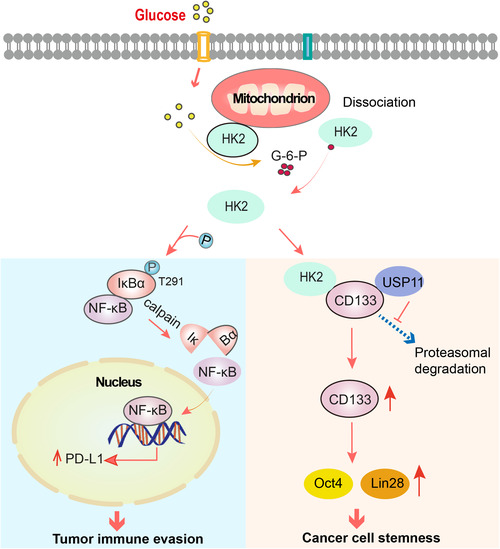
The moonlighting functions of HK2 in tumor immunity and tumor cell stemness. Under aerobic glycolysis conditions, non‐mitochondrial HK2 dissociates from mitochondria and acts as a protein kinase to promote NF‐κB‐mediated PD‐L1 expression and tumor immune evasion. In addition, non‐mitochondrial HK2 maintains cell stemness by USP11‐mediated stabilization of CD133 and subsequent upregulation of Oct4 and Lin28 expression. Abbreviations: HK, hexokinase; NF‐Κb, nuclear factor kappa‐light‐chain‐enhancer of activated B cells; PD‐L1, programmed cell death ligand 1; USP11, ubiquitin‐specific protease 11.
Thus, the Warburg effect results in the multifaceted roles of highly expressed HK2 in tumor cells not only in glycolysis but also in tumor immunity and tumor cell stemness. Altogether, these findings underscore the critical moonlighting functions of HK2 in tumor progression and highlight the potential to target these functions for cancer treatment.
AUTHOR CONTRIBUTIONS
ZML conceptualized the article. HYH, LWX, JHW, DG and ZML wrote and revised the manuscript.
COMPETING INTERESTS
The authors declare no competing interests.
AVAILABILITY OF DATA AND MATERIAL
Not applicable.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
Not applicable.
DECLARATIONS
CONSENT FOR PUBLICATION
Not applicable.
ACKNOWLEDGMENTS
Not applicable.
He H, Xiao L, Wang J, Guo D, Lu Z. Aerobic glycolysis promotes tumor immune evasion and tumor cell stemness through the noncanonical function of hexokinase 2. Cancer Communications. 2023;43:387–390. 10.1002/cac2.12404
Contributor Information
Dong Guo, Email: guodaye@zju.edu.cn.
Zhimin Lu, Email: zhiminlu@zju.edu.cn.
REFERENCES
- 1. Wang Y, Xia Y, Lu Z. Metabolic features of cancer cells. Cancer Commun (Lond). 2018;38(1):65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Robey RB, Hay N. Mitochondrial hexokinases, novel mediators of the antiapoptotic effects of growth factors and Akt. Oncogene. 2006;25(34):4683‐96. [DOI] [PubMed] [Google Scholar]
- 3. Garcia SN, Guedes RC, Marques MM. Unlocking the Potential of HK2 in Cancer Metabolism and Therapeutics. Curr Med Chem. 2019;26(41):7285‐322. [DOI] [PubMed] [Google Scholar]
- 4. Xu D, Shao F, Bian X, Meng Y, Liang T, Lu Z. The Evolving Landscape of Noncanonical Functions of Metabolic Enzymes in Cancer and Other Pathologies. Cell Metab. 2021;33(1):33‐50. [DOI] [PubMed] [Google Scholar]
- 5. Li X, Egervari G, Wang Y, Berger SL, Lu Z. Regulation of chromatin and gene expression by metabolic enzymes and metabolites. Nat Rev Mol Cell Biol. 2018;19(9):563‐78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Bian X, Liu R, Meng Y, Xing D, Xu D, Lu Z. Lipid metabolism and cancer. J Exp Med. 2021;218(1):e20201606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Guo D, Tong Y, Jiang X, Meng Y, Jiang H, Du L, et al. Aerobic glycolysis promotes tumor immune evasion by hexokinase2‐mediated phosphorylation of IκBα. Cell Metabolism. 2022;34(9):1312‐24.e6. [DOI] [PubMed] [Google Scholar]
- 8. Wang J, Shao F, Yang Y, Wang W, Yang X, Li R, et al. A non‐metabolic function of hexokinase 2 in small cell lung cancer: promotes cancer cell stemness by increasing USP11‐mediated CD133 stability. Cancer Commun (Lond). 2022;42(10):1008‐1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Sarvi S, Mackinnon AC, Avlonitis N, Bradley M, Rintoul RC, Rassl DM, et al. CD133+ cancer stem‐like cells in small cell lung cancer are highly tumorigenic and chemoresistant but sensitive to a novel neuropeptide antagonist. Cancer Res. 2014;74(5):1554‐65. [DOI] [PubMed] [Google Scholar]
- 10. Yang W, Xia Y, Hawke D, Li X, Liang J, Xing D, et al. PKM2 phosphorylates histone H3 and promotes gene transcription and tumorigenesis. Cell. 2012;150(4):685‐96. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.