

Abstract
The plant signaling molecule auxin is present in multiple kingdoms of life. Since its discovery, a century of research has been focused on its action as a phytohormone. In land plants, auxin regulates growth and development through transcriptional and non‐transcriptional programs. Some of the molecular mechanisms underlying these responses are well understood, mainly in Arabidopsis. Recently, the availability of genomic and transcriptomic data of green lineages, together with phylogenetic inference, has provided the basis to reconstruct the evolutionary history of some components involved in auxin biology. In this review, we follow the evolutionary trajectory that allowed auxin to become the “giant” of plant biology by focusing on bryophytes and streptophyte algae. We consider auxin biosynthesis, transport, physiological, and molecular responses, as well as evidence supporting the role of auxin as a chemical messenger for communication within ecosystems. Finally, we emphasize that functional validation of predicted orthologs will shed light on the conserved properties of auxin biology among streptophytes.
Keywords: auxin, ecology, evolution, hormone response, plant biology
Subject Categories: Plant Biology
This review gives us an evolutionary perspective on auxin, a key regulator of plant growth and development.
Introduction
In search of the mysterious endogenous plant compound underlying differential growth, auxin was the first plant hormone to be identified (Darwin & Darwin, 1881; Went & Thimann, 1937). Early experiments showed that auxin acts as a chemical messenger by transporting a “growth stimulus” from shoots to roots upon the perception of light and gravity (Darwin & Darwin, 1881; Boysen‐Jensen, 1910). The chemical was identified as indole‐3‐acetic acid (IAA) and represents plants' most abundant natural auxin (Kögl et al, 1934; Went & Thimann, 1937). What is remarkable about auxin is that it influences almost every aspect of plant growth and development. The ability to act locally is conditioned by the ability of plants to control local synthesis, inactivation, and cell–cell transport (Friml, 2021). Auxin not only acts as a coordinating signal between cells, tissues, and organs, but also mediates interaction with the environment, both as a trigger of environmentally controlled development and as a messenger for interkingdom communication (Weijers & Wagner, 2016; Kunkel & Harper, 2018).
Most studies have focused on the role and influence of auxin in the growth and development of flowering plants (angiosperms). However, a similar range of activity has been reported in bryophytes (Reviewed in Kato et al, 2018), suggesting a conserved role across land plants. In addition, some extant green algae also show responses to exogenous IAA (Wood & Berliner, 1979; Ohtaka et al, 2017). Growing phylogenetic and molecular data indicate that the ancestor of modern land plants made their transition to land around 480 million years ago and evolved from streptophyte algae most closely related to the Zygnematophyceae; a group of filamentous and unicellular algae that live in terrestrial and freshwater environments (Wodniok et al, 2011; Timme et al, 2012; Hess et al, 2022).
Therefore, there is an increasing effort among the scientific community to understand which innovations allowed these photosynthetic organisms to move from water to land and become the dominant macroscopic flora we see around us today (de Vries & Archibald, 2018). Among such innovations, signaling pathways regulated by phytohormones have been proposed to be a main driver for the adaptation of plants to new niches and the evolution of complexity (Rensing, 2018; Blázquez et al, 2020). In this review, we address the evolutionary origins of auxin biology in plants (Fig 1). Given that the origins and early diversification of auxin functions predate and coincide with plant terrestrialization (Bowman et al, 2017; Mutte et al, 2018), we place particular emphasis on bryophytes and streptophyte algae. We consider endogenous auxin—IAA biosynthesis, transport, physiological and molecular responses, and the role of auxin in communication within different ecosystems. In doing so, we can now start to understand the evolutionary trajectory that has allowed auxin to become a signaling molecule that regulates so many biological processes in the plant kingdom—the regulatory “giant” of plant biology.
Figure 1. Illustration of a land plant cell with the main components of auxin biology.
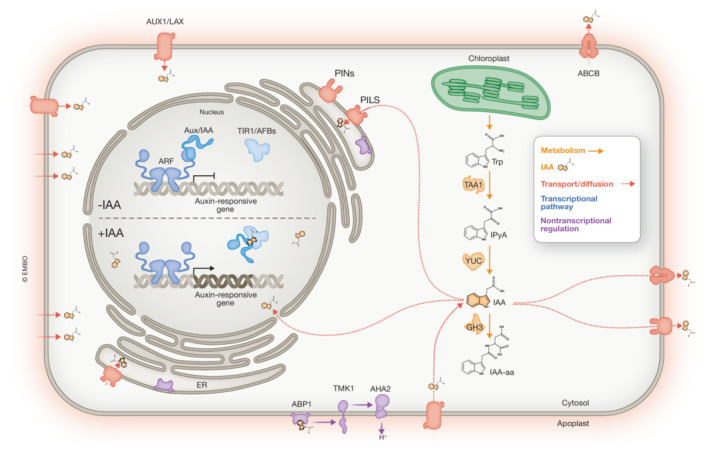
The most common precursor of Indole 3‐acetic acid (IAA; indicated in orange structures) biosynthesis is tryptophan synthesized in the chloroplasts. In plants, indole‐3‐pyruvic acid (IPyA) is the major intermediate and this involves the function of TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS (TAA) followed by decarboxylation catalyzed by members of the YUCCA (YUC) family. IAA can then be stored in the cells by forming amino acid conjugates catalyzed by GRETCHEN HAGEN3 (GH3) auxin‐amido synthetases. IAA is transported inside the cells via importers (AUX/LAX) or by diffusion (protonated form IAAH). IAA is an anionic form inside the cells and is exported via PIN proteins localized at the plasma membrane (PM). PILS is a family of proteins mainly localized to the endoplasmic reticulum (ER) and have an auxin‐transport function and presumably contribute to intracellular auxin homeostasis. A second family of PM transporters, ABCBs, act mainly as IAA efflux carriers, working together with PINs in transporting IAA outside the cells. Inside the nucleus, IAA regulates the NAP. When auxin is abundant in the nuclear environment (orange structures) it binds to members of the TIR1/AFB receptor family (light blue) part of a ubiquitin ligase complex and to AUX/IAA repressors (light blue), targeting the latter for degradation. Thus, the auxin response factors (ARFs) can activate the transcription of auxin‐responsive genes. AUXIN‐BINDING PROTEIN1 (ABP1) functions as auxin receptor in the apoplast, and it is known to mediate transmembrane kinase (TMK1) proteins that mediate rapid responses. TMK1 mediates the phosphorylation of AHA‐plasma membrane H+‐ATPases in the presence of auxin, which is translated into acidification of the apoplast.
Where the giants are…
Auxin is found in multiple kingdoms of life. IAA has been hypothesized to be synthesized in more than 80% of prokaryotes, including several phyla of archaea and bacteria (Zhang et al, 2019 †). Many of them are directly associated with plants with members that include pathogens, nitrogen‐fixing symbionts, plant growth‐promoting rhizobacteria, as well as non‐symbiotic bacteria and marine microorganisms (Amin et al, 2015; Cox et al, 2018; Kunkel & Harper, 2018). Auxins have also been found in several eukaryotes, such as fungi that form symbiotic or non‐symbiotic associations with green lineages (Cohen et al, 2002; Rao et al, 2010; Chanclud & Morel, 2016; Leontovyčová et al, 2020). Other eukaryotes, such as brown and red algae, as well as many chlorophytes and streptophytes, show endogenous accumulation of IAA (Basu et al, 2002; le Bail et al, 2010; Mikami et al, 2016; Bogaert et al, 2019). Given this extremely broad distribution, IAA accumulation in any organism should be interpreted cautiously, unless the presence of associated microbes can be excluded.
That there is no universal pathway for the biosynthesis of IAA (Morffy et al, 2020) is perhaps coupled to its structural simplicity. However, in most cases, tryptophan (Trp) acts as a precursor. The production of IAA flows through several enzymatic pathways named after their intermediate compounds, such as indoleacetamide (IAM) and indole‐3‐pyruvic acid (IPyA) (Reviewed in Morffy & Strader, 2020). Fungi and bacteria generally use the IAM and IPyA pathways for auxin production (Chanclud & Morel, 2016; Morffy et al, 2020). The non‐plant IPyA pathway is based first on the formation of indole‐3‐acetaldehyde (IAAld) by decarboxylases, followed by the conversion of IAAld to IAA by a dehydrogenase (Leontovyčová et al, 2020; Dong et al, 2022). In plants, IPyA is the major biosynthetic intermediate of IAA synthesis (Mashiguchi et al, 2011; Morffy et al, 2020). Here, the deamination of Trp to IpyA is catalyzed by the TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS (TAA) family of aminotransferases (Tao et al, 2008; Mashiguchi et al, 2011). Subsequently, IPyA is decarboxylated to IAA by the YUCCA (YUC) family of flavin‐containing monooxygenases (FMO; Mashiguchi et al, 2011; Stepanova et al, 2011; Dai et al, 2013).
Phylogenetic analyses have shown that both TAA and YUC gene families are deeply conserved across genomes of land plants, being represented both in tracheophytes (lycophytes, ferns, gymnosperms and angiosperms) and bryophytes (liverworts, hornworts, and mosses; Fig 2; Yue et al, 2014; Bowman et al, 2017). The conserved role of the IAA biosynthetic pathway in land plants has been established through genetic and pharmacological analyses in bryophytes (Landberg et al, 2013; Eklund et al, 2015). For example, in the liverwort Marchantia polymorpha, disruption of this pathway causes dramatic developmental defects (Eklund et al, 2015).
Figure 2. Schematic summary of presence or absence of ancestral copies of the main genetic components of auxin biology in plants.
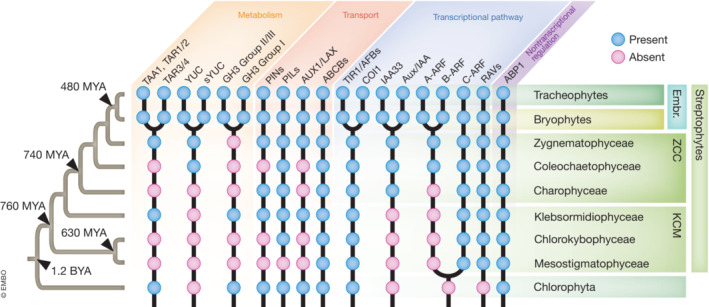
Biosynthesis and metabolism (orange), transport (pink), transcriptional pathway (light blue), and non‐transcriptional regulation (purple). Orthologs present (blue circles) or absent (pink circles). Land plants also referred to as embryophytes (Embr.) are divided into tracheophytes and bryophytes and belong to the Viridiplantae (plants) clade of eukaryotic organisms. Plants are found in almost every habitat on Earth and are divided into two main clades, streptophytes and chlorophytes. The Streptophytes comprise a group of green algae, the streptophyte algae, and the land plants (One Thousand Plant Transcriptomes Initiative, 2019). Streptophyte algae are a paraphyletic group of extant algae that encompass the lower‐branching KCM‐grade classes, Klebsormidiophyceae, Chlorokybophyceae, and Mesostigmatophyceae and the higher‐branching ZCC‐grade classes, Zygnematophyceae, Coleochaetophyceae, and Charophyceae (de Vries & Archibald, 2018).
The TAA gene family is divided into two clades that encode alliinase domain‐containing proteins (Figs 2 and EV1; Wang et al, 2014; Bowman et al, 2017). The TAA clade in Arabidopsis includes three homologs that contain an Alliinase‐C domain (IPR006948): TAA1, TAR1, and TAR2; the Alliinase clade includes two homologs, TAR3 and TAR4, which contain both an Alliinase‐EGF (Epidermal growth factor, IPR006947) domain at their N‐terminus and an Alliinase‐C domain at their C‐terminus. In embryophytes, the TAA subclade contains the orthologs that are involved in the biosynthesis of IAA, whereas the functions of orthologs that contain both Alliinase domains are yet to be described (Tao et al, 2008; Eklund et al, 2015). In this review, we extend previous analyses on the evolutionary origin of TAA homologs to the green algae. We have identified only one ortholog in each algal species (Fig EV1). Most of the chlorophyte orthologs have only the Alliinase‐C domain. In contrast, most of the orthologs found in streptophyte algae contain both the Alliinase‐EGF and the Alliinase‐C domains. These data suggest that the TAA/Alliinase homologs that are present in streptophyte algae are reminiscent of an ancestral gene with unknown substrate specificity present in the last common land plant/bryophyte ancestor. This gene is likely to have duplicated and neofunctionalized in land plants during evolution (Bowman et al, 2021).
Figure EV1. Phylogenetic tree of the TAA gene family with green algae and land plant homologs.
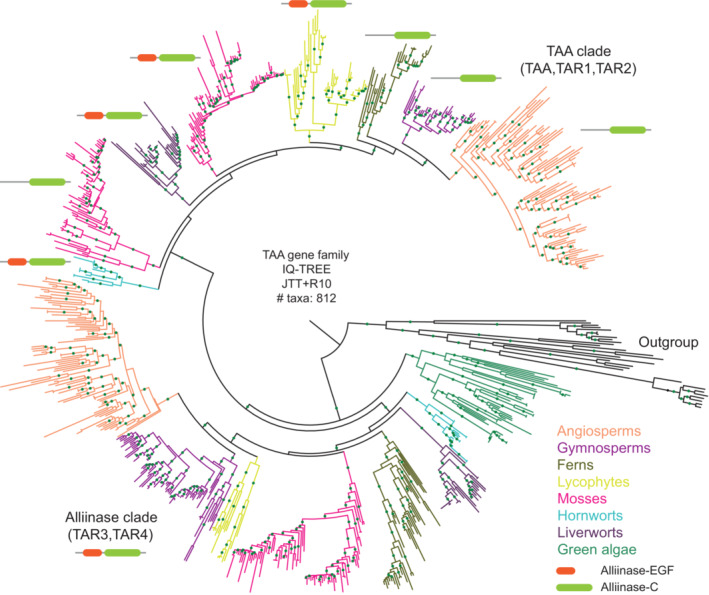
Protein domains, Alliinase‐EGF and Alliinase‐C are indicated with “red” and “green” representations, respectively. TAR3/TAR4 clade shows the consistent presence of both domains in all lineages, whereas in TAA/TAR1/TAR2 clade, some phyla lack Alliinase‐EGF domain. Aminotransferases from land plants, other than TAA members, were used as outgroup sequences to root the tree. Branches that are well supported (bootstrap > 75) are indicated by green dots. Orthologs from each phylum are represented in different colors, as indicated in the right bottom right legend. Basic information about the tree construction: “software,” “model of evolution,” and the “number of taxa” used for phylogenetic tree construction are indicated at the center. The complete tree can be found at the interactive Tree of Life (iTOL) repository: https://itol.embl.de/shared/dolfweijers.
The YUC gene family in land plants is divided into two clades (Figs 2 and EV2; Wang et al, 2014). The YUC clade contains 11 members in Arabidopsis. Some other YUCs from embryophytes are placed in a sister clade to the YUC (sYUC, Fig EV2), but this clade seems to have been independently lost in seed plants and mosses. Some green algae sequences form a sister clade basal to both YUC and sYUC. Further phylogenetic analysis including other FMO family proteins such as flavin monooxygenase 1 (FMO1) and FMO glucosinolate S‐oxygenase (GS‐OX) showed that YUC orthologs in land plants and in streptophyte algae are different from other FMO orthologs (Fig EV2). Both clades of YUCs are conserved among land plants but are not common to all members of the green algae ZCC clade (Zygnematophyceae, Coleochaetophyceae, and Charophyceae; Fig 2). Other phylogenetic studies hypothesize that, even though YUC homologs are identified in algal lineages, these FMOs enzymes are likely an innovation of land plants acquired by horizontal gene transfer from bacteria (Yue et al, 2014; Bowman et al, 2017). Hence, the origin of this gene family in streptophyte algae remains unsolved. Cross‐species complementation, biochemical, and genetic studies in model streptophyte algae are needed to validate the conservation of an ancestral function.
Figure EV2. Phylogenetic tree of the YUC gene family with algae and land plant homologs.
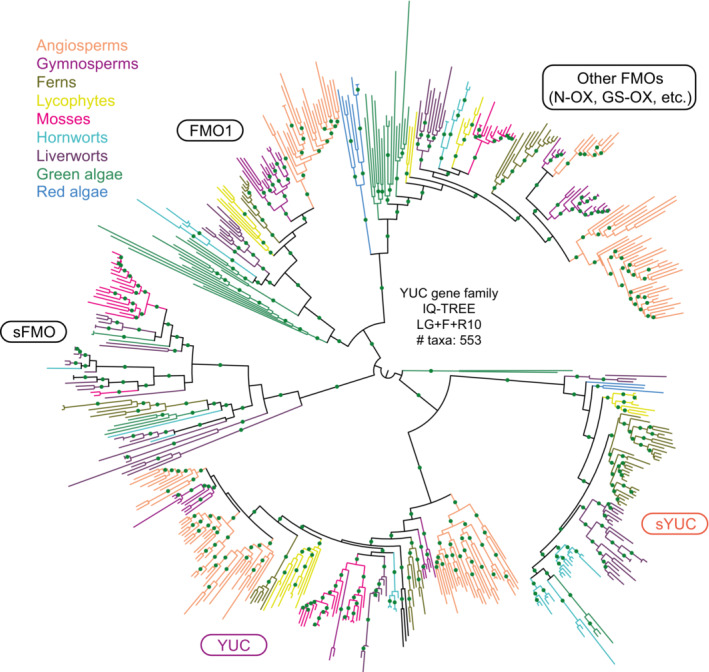
The different FMO clades are indicated by representative Arabidopsis family member names (FMO, flavin monooxyenase 1; GS‐OX, FMO glucosinolate S‐oxygenase; N‐OX, FMO N‐oxygenases; YUC, Yucca). Branches that are well supported (bootstrap > 75) are indicated by green dots. Orthologs from each phylum are represented in different colors, as indicated in the bottom right legend. Basic information about the tree construction: “software,” “model of evolution,” and the “number of taxa” used for phylogenetic tree construction are indicated at the center. The complete tree can be found at the interactive Tree of Life (iTOL) repository: https://itol.embl.de/shared/dolfweijers.
Besides synthesis, other mechanisms control the amount of IAA available during plant growth. These include transport (see next section), conjugation, glycosylation, and oxidation (Reviewed in Casanova‐Sáez et al, 2021). One of the most common inactivation forms of IAA is the formation of amino acid conjugates catalyzed by GRETCHEN HAGEN3 (GH3) auxin‐amido synthetases (Staswick et al, 2005; Ludwig‐Müller et al, 2009). Some conjugates such as IAA‐Ala or IAA‐Leu are reversibly converted to free IAA by amidohydrolases and are considered auxin storage forms (LeClere et al, 2002; Rampey et al, 2004). While others such as IAA‐Asp and IAA‐Glu are irreversibly converted to free IAA and are considered auxin catabolites (Rampey et al, 2004; Mellor et al, 2016; Casanova‐Sáez et al, 2021). Phylogenetic and metabolomic analyses indicate that members of the GH3 family and amino acid‐IAA conjugates are common to land plants (Staswick et al, 2005; Terol et al, 2006; Bowman et al, 2021). The GH3 family is divided into three phylogenetic subgroups with different substrate preferences (Figs 2 and EV3; Bowman et al, 2021). In angiosperms, members of group II function in auxin conjugation, while those of group I and III use jasmonic acid (JA) or benzoate as substrates, respectively (Staswick et al, 2005; Okrent et al, 2009). In the moss Physcomitrium patens, two GH3 homologs of group I have been shown to convert IAA and JA to amino acid conjugates, suggesting a conserved function of this gene family in the land plants (Ludwig‐Müller et al, 2009). All the groups seem to have emerged from a single streptophyte algae ancestral GH3 gene with unknown substrate specificity as observed in a known GH3 homolog of Klebsormidium nitens (KCM clade, Klebsormidiophyceae, Chlorokybophyceae, and Mesostigmatophyceae). However, as this homolog cannot be assigned to any specific subgroup, it has been hypothesized that K. nitens GH3 could have been obtained from bacteria by horizontal gene transfer (Bowman et al, 2021). In this scenario, the molecular machinery by which IAA‐amino acid conjugation maintains auxin homeostasis would be a land plant innovation. Thus genomic, and biochemical data are ambivalent with regard to the capacity of streptophyte algae to synthesize or inactivate auxin in a similar way to land plants.
Figure EV3. Phylogenetic tree of the GH3 gene family with green algae and land plant homologs.
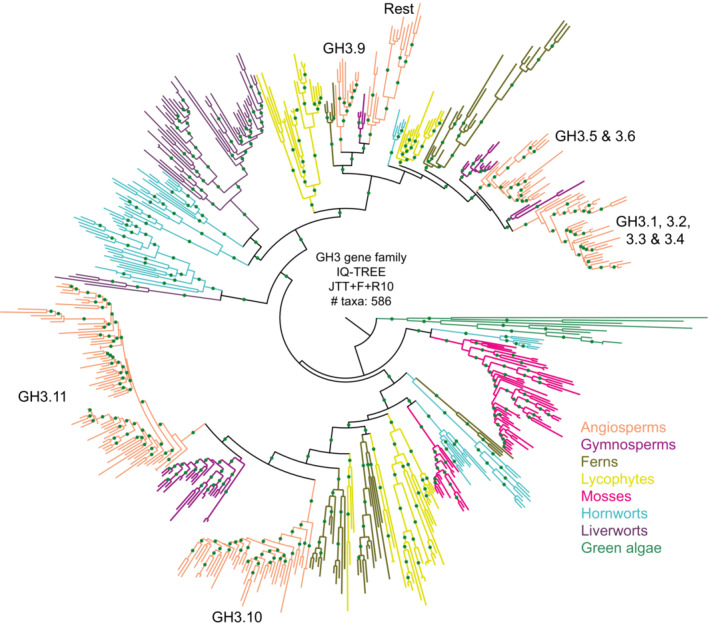
Respective Arabidopsis orthologs that are present in the specific clade are mentioned with the corresponding Arabidopsis family member names. “Rest” includes GH3.7, 3.8, 3.12 until 3.19. Branches that are well supported (bootstrap > 75) are indicated by green dots. Orthologs from each phylum are represented in different colors, as indicated in the bottom right legend. Basic information about the tree construction: “software,” “model of evolution,” and the “number of taxa” used for phylogenetic tree construction are indicated at the center. The complete tree can be found at the interactive Tree of Life (iTOL) repository: https://itol.embl.de/shared/dolfweijers.
Moving giants…
Studies conducted in vascular plants show that once auxin is synthesized in source organs, such as young leaves, it is transported over both short and long distances, partly by diffusion and partly through carriers that offer directionality. The “polarity” of auxin distribution is mainly attributed to the asymmetric localization of members of the PIN‐FORMED (PIN) family of auxin efflux carriers at the plasma membrane (Bennett et al, 1996; Chen et al, 1998; Gälweiler et al, 1998). Therefore, the localization of PIN proteins establishes the directionality of auxin flow, known as “polar auxin transport,” as reviewed in Michniewicz et al (2007). Phylogenetic analyses show that PIN proteins are conserved along the green lineages (Fig 2) and have been classified into canonical and non‐canonical based on the length of their central hydrophilic loop and their cellular localization (Billou et al, 2005; Reviewed in Bennett, 2015; Hammes et al, 2022). Genetic disruption of canonical PINs in P. patens affects auxin transport and tropisms in gametophores, similar to the effects of auxin transport disruption in Arabidopsis shoot and root tropisms (Bennett et al, 2014). In streptophyte algae, homologs of PIN proteins form a single clade (Bogaert et al, 2022). Functional studies of a K. flaccidum PIN show that this protein acts as an efflux carrier in an angiosperm heterologous system (Skokan et al, 2019), suggesting a conserved origin of the biochemical properties of PINs in streptophyte algae (Boot et al, 2012; Skokan et al, 2019). However, genetic dissection of PIN function in model green algae is necessary to validate the transport and biological functions of these proteins.
Another protein family of auxin transporters, the PILS (PIN‐LIKES), has been identified based on homology to the PINs. These proteins are localized to the endoplasmic reticulum, and genetic and pharmacological analyses indicate that they regulate intracellular auxin homeostasis in Arabidopsis (Barbez et al, 2012). Interestingly, phylogenetic analyses of the PILS family show that its members are present in almost all eukaryotic clades, including all the green lineages (Bogaert et al, 2022). These data suggest that if the PILS‐mediated transport function is auxin‐specific, intracellular auxin regulation may have predated the intercellular directional auxin transport (Bogaert et al, 2022). A key question is whether such an ancient function in intracellular partitioning served a role in physiological regulation and signaling, or rather in detoxification or metabolism.
Another family of proteins that acts as auxin efflux carriers in angiosperms is the ATP‐binding cassette subfamily B/multidrug resistance/phosphoglycoprotein (ABCB/MDR/PGP; Noh et al, 2001; Verrier et al, 2008), with some homologs showing auxin influx properties (Kamimoto et al, 2012; Reviewed in Hammes et al, 2022). However, the major contribution to active IAA uptake in plants is made by members of the AUXIN RESISTANT 1/LIKE AUXIN RESISTANT 1 (AUX1/LAX) influx carriers (Bennett et al, 1996). This family of plasma membrane proton co‐transporters was first identified in Arabidopsis auxin‐resistant aux mutants. Members of the ABCB and AUX1/LAX carrier families are deeply conserved throughout green lineages (Fig 2; Vosolsobě et al, 2020; Bowman et al, 2021). Taken together, while there are either well‐defined or more distantly related homologs of all auxin transporters across the green kingdom, functional studies for most streptophytes, that would confirm the conservation of auxin transport driven by PIN, PILS, ABCB, and AUX1/LAX carriers, are lacking.
A picture emerges in which auxin occurs broadly, with land plants (and possibly some algae) actively controlling its accumulation. But what responses occur? What mechanisms underlie these? And how did these originate and evolve?
How to respond to giants…
Auxin controls almost every process in the growth and development of plants, and there is a considerable insight into the physiological, cellular, biochemical, and genetic mechanisms triggering these responses (Weijers & Wagner, 2016; Leyser, 2018). Both endogenous and externally applied auxin triggers changes in growth (Reviewed in Cooke et al, 2002). For example, auxin causes responses such as inducing cell elongation in Arabidopsis hypocotyls that translate to overall hypocotyl growth (Lin et al, 2021). The same auxin stimulus, however, leads to the growth inhibition of roots, a process underlying gravitropic bending (Evans et al, 1994; Fendrych et al, 2018). These growth differences between organs suggest specific mechanisms for auxin‐dependent growth regulation, which has triggered the curiosity of researchers for many decades (See next section).
In the green alga Chara, auxin has a stimulatory effect on the development of rhizoids, similar to its effects on bryophytes, and the induction of root hairs in angiosperms (Sandan & Ogura, 1957; Klambt et al, 1992; Jones & Dolan, 2012). In land plants, the development of rooting systems is hypothesized to be mediated by a conserved auxin—RSL (ROOT HAIR DEFECTIVE 6‐LIKE) genetic network (Jones & Dolan, 2012). Other examples of auxin responses in streptophyte algae have been observed with exogenous IAA treatments of Micrasterias thomasiana and K. nitens. Here, auxin either promotes or inhibits cell division, which could be partly explained by the large differences in the IAA concentrations used for the treatments (Wood & Berliner, 1979; Ohtaka et al, 2017). Thus, while there are physiological effects, it is unclear whether there is a generic algal auxin response mechanism.
In addition to effects on growth, several reports document auxin responses at a cellular level that include plasma membrane depolarization, cytosolic calcium influx, reorganization of the actin network, acidification/alkalinization of the apoplast, and stimulation of cytoplasmic streaming, sometimes within seconds or minutes of auxin application (Thimann & Sweeney, 1937; Arieti & Staiger, 2020; Friml et al, 2022; Li et al, 2022). In Nitella, a charophycean alga, auxin has been shown to stimulate cytoplasmic streaming of internodal cells in a similar way to that seen in Arabidopsis root epidermal cells (Thimann & Sweeney, 1937; Friml et al, 2022). A key question is what biological functions of auxin are mediated by this array of fast and slow cellular responses across the plant kingdom.
Giant steps…
Essentially, all molecular mechanisms that are known to mediate auxin responses were identified in angiosperms, chiefly in Arabidopsis. The auxin response involves a relatively well‐understood transcriptional branch—the nuclear auxin pathway (NAP)—(Reviewed in Weijers & Wagner, 2016; Leyser, 2018). There is also a branch of rapid auxin responses that may mediate such fast cellular responses that cannot be mediated by transcriptional changes (See below; Reviewed in Dubey et al, 2021).
The NAP has been a subject of intense investigation for the last three decades (Abel et al, 1994; Ulmasov et al, 1997a, 1997b, 1999; Boer et al, 2014; Kato et al, 2020), revealing a transcriptional module controlled by auxin that is likely only to be present in land plants (Martin‐Arevalillo et al, 2019; Bowman et al, 2021). Forward genetic screenings using Arabidopsis root growth inhibition have been invaluable in identifying the key components in auxin response (Abel et al, 1995). The NAP involves the perception of auxin by transport inhibitor response 1/auxin F‐box (TIR1/AFB) proteins (Kepinski & Leyser, 2005), substrate‐binding subunits of ubiquitin ligases (Tan et al, 2007). When cellular IAA levels rise, the hormone acts as a molecular glue that increases the affinity of TIR1/AFB to its target substrates, the Aux/IAA transcriptional inhibitor proteins (Tan et al, 2007). This interaction triggers subsequent ubiquitin‐dependent degradation of the repressors (Gray et al, 2001) and presumably enhances the production of cAMP catalyzed by an adenylate cyclase embedded in the auxin pocket of TIR1/AFBs (Qi et al, 2022). At low auxin concentrations, Aux/IAAs recruit TOPLESS co‐repressors (Szemenyei et al, 2008), bind to, and inhibit the activity of DNA‐binding ARF transcription factors (Korasick et al, 2014; Nanao et al, 2014). Thus, auxin regulates this system by promoting proteolysis of Aux/IAAs and releasing ARFs from inhibition, leading to changes in the expression of hundreds to thousands of genes (Weijers & Wagner, 2016). Although the model is simple, in most land plants, each gene family has multiple paralogs resulting in an extensive combinatorial network with multiple outputs (Finet et al, 2013; Freire‐Rios et al, 2020).
Auxin responses regulated by the NAP are deeply conserved across land plants, as shown by different phylogenetic, transcriptomic, functional, and biochemical analyses (Flores‐Sandoval et al, 2015; Kato et al, 2015, 2020; Mutte et al, 2018; Fig 2). Recent genetic studies show that the NAP controls the development of bryophytes such as the liverwort M. polymorpha and the moss P. patens in a manner similar to its action in angiosperms (Fig 2; Lavy et al, 2016; Reviewed in Kato et al, 2018). Following work in Physcomitrium (Prigge et al, 2010; Lavy et al, 2016), genetic and structural analyses in Marchantia revealed a deeply conserved function of a minimal set of auxin signaling components (Reviewed in Kato et al, 2018; Kato et al, 2020). This bryophyte contains one gene copy of each subclass within the ARF family including the canonical A, B, and C types, a non‐canonical ARF (subclass lacking DNA‐binding domain), and a single Aux/IAA and TIR1 proteins (Flores‐Sandoval et al, 2015; Kato et al, 2015; Mutte et al, 2018). Genetic disruption of the A‐class ARF (MpARF1) causes severe developmental phenotypes and a lack of auxin responses that cannot be complemented with a different class of ARF (MpARF2 or MpARF3; Kato et al, 2020). These data suggest dedicated functions of each ARF class. Domain swapping complementation experiments using the Mparf1 mutant and the domains of all the ARF classes showed that class‐A and ‐B compete for the same DNA‐binding sites while class‐C ARF is not involved in auxin responses (Kato et al, 2020). Class‐A ARF can recruit MpIAA and activate auxin‐dependent gene expression, while Class‐B ARF is not auxin‐dependent (Kato et al, 2015, 2020). In the current model, the expression patterns and stoichiometries of both proteins determine the responsiveness of cells to auxin (Kato et al, 2020). This simple model may explain the molecular basis of the transcriptional auxin response network in land plants. It does not, however, tell us how this transcriptional machinery evolved. Of the three NAP protein families, streptophyte algae only contain well‐defined ARF‐like proteins (proto‐ARFs) with predicted similar architecture and perhaps biochemical function to land plant ARFs (Fig 2; Mutte et al, 2018; Martin‐Arevalillo et al, 2019). Phylogenetic analysis and prediction of Proto‐ARF structures show that they fall into two clades: A/B and C, which likely represent the ancestral precursors of land plant ARFs and that already diverged in an ancient clade of streptophyte algae (Fig 2). Accordingly, recent biochemical analysis shows that a Proto‐C‐ARF from Chlorokybus atmophyticus has a DNA‐binding specificity that differs from land plants A and B, and that has been maintained in class‐C ARFs along evolution (Martin‐Arevalillo et al, 2019). Presence of Proto‐C ARFs is common along the streptophyte algae with no specific pattern of appearance or retention, while Proto A/B‐ARFs have been found mainly in the ZCC clade (Martin‐Arevalillo et al, 2019; Mutte, 2020). These observations indicate that an ancestral Proto‐C ARF could have duplicated and possibly acquired new functions in the common ancestor of the ZCC algal clade and the land plants (Fig 2; Martin‐Arevalillo et al, 2019). Curiously, most of the reported species in this algal clade contain only one class of Proto‐ARF, which may reflect independent losses or limited depth of sequencing of the sampled species. The genomes of streptophyte algae encode predicted TIR1/AFB‐like and AUX/IAA‐like proteins (Bowman et al, 2017; Mutte et al, 2018; Fig 2). However, in both cases, they lack critical residues to interact with auxin. Phylogenetic analysis of Aux/IAA homologs shows that this gene family is deeply conserved in the embryophytes, divided into two clades, canonical and non‐canonical (Fig 2; Mutte et al, 2018). In streptophyte algae, there are predicted Aux/IAA orthologs only in the ZCC clade, suggesting that this Proto‐AUX/IAA represents the ancestral precursor of the neofunctionalized land plant AUX/IAAs (Bowman et al, 2017; Mutte et al, 2018). The overall predicted structures of TIR1‐like proteins are similar to land plant auxin (and jasmonate) receptors, but amino acids for auxin binding are not conserved. Taken together, the available functional and phylogenetic analysis allows a plausible ancestral land plant evolutionary scenario to be proposed, in which a diversified Proto‐ARF network was coopted for regulation by auxin upon the evolution of an increased auxin‐binding affinity in proto‐TIR1 and proto‐Aux/IAA proteins (Fig 3). Thus, phylogenomic data suggest that the transcriptional auxin signaling pathway is most likely a land plant innovation. This begs the question of what the ancestral function of the proto‐ARF factors is, and what mutational trajectory ushered the birth of the auxin response system. Functional studies in emerging genetic models of extant streptophyte algae should help answer these questions (Zhou et al, 2020). Likewise, reconstructing ancestral proteins could help to understand the evolutionary trajectory that caused the auxin responses, protein structure, and function to diverge in the green lineages (Scossa & Fernie, 2021).
Figure 3. Evolutionary scenario of ancestral auxin biology in streptophytes.
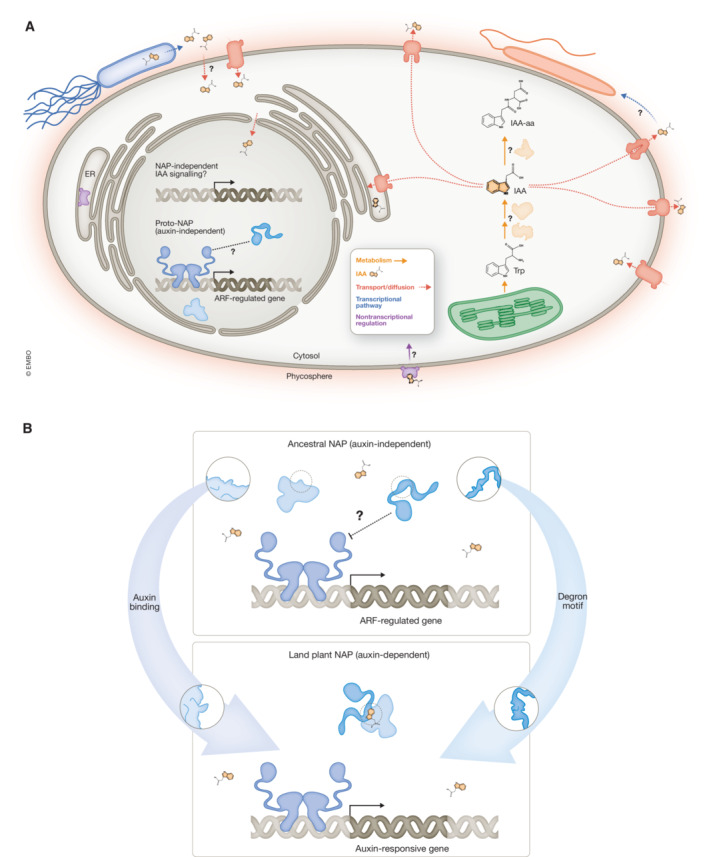
(A) Auxin (IAA) has been found in extant streptophyte algae and likely in the last common ancestor of streptophytes. However, the biosynthetic pathway for its production remains unknown. In streptophyte algae, there are predicted orthologs of gene families that in plants catalyze the conversion of Trp to IAA via IPyA, TAA, and YUC. Among the mechanisms that contribute to regulating IAA levels in plants, storage mediated by members of the GH3 gene family remains controversial in algae as only a few members of this family have been reported. Regarding transport (red), predicted orthologs of PIN, PILS, ABCB, and AUX/LAX are present in members of the green algae, suggesting that the transport function was present in an ancestral streptophyte. Several responses to auxin have been observed in streptophyte algae, but the mechanisms underlying these responses are unknown. In land plants, fast responses to auxin are mediated by auxin perception via the ABP1‐TMK1 module. In extant algae, the ABP1 gene family is well conserved, suggesting a plausible scenario in which an ancestral streptophyte might have been responding to local auxin produced in the phycosphere (red). (B) Homologs of the three main components of the NAP are present in extant algae. However, the TIR1‐like receptor and Aux/IAA‐like predicted orthologs lack the components to interact with auxin. Therefore, there is a plausible land plant scenario in which a diversified Proto‐ARF network came to be regulated by auxin upon innovations in TIR1 and Aux/IAA protein families.
Some auxin responses, such as the rapid root growth inhibition in Arabidopsis, happen too rapidly to involve transcription, yet are also dependent on AFB1 function (Dindas et al, 2018; Fendrych et al, 2018; Serre et al, 2021). Components of a non‐transcriptional AFB1 pathway remain unknown, and it is unclear whether such functions are shared by other species and other TIR1/AFB receptors (Prigge et al, 2020). Recent studies show that auxin triggers very rapid (< 2 min) phosphorylation of about a thousand proteins in Arabidopsis roots, and that this requires both the auxin‐binding protein 1 (ABP1) and its potential partner protein transmembrane kinase 1 (TMK1; Friml et al, 2022). Outcomes of this rapid signaling appear to include the activation of H+‐ATPases that acidify the apoplast and the regulation of Myosin XI that may influence cytoplasmic streaming (preprint: Han et al, 2021; Lin et al, 2021; Friml et al, 2022). Interestingly, disruption of ABP1 and TMK leads to defects in both regeneration from callus and regeneration of vascular strands around a stem wound (preprint: Han et al, 2021; Friml et al, 2022). However, it remains unknown how the ultrafast auxin phospho‐response is connected to slower, developmental responses. Important questions are whether these fast responses are conserved across species. If so, are they regulated by the ABP1‐TMK module in all the land plants (and perhaps algae)? ABP1 is a well‐conserved gene family that is maintained as a single copy in all the green lineages including all the streptophyte algae lineages (Fig 2). Clearly, there are new twists and turns ahead in the adventurous life of the auxin giant…
Communicating through giants…
Auxin produced by microbes has been shown to trigger responses in plants, including changes in growth patterns or developmental programs (Sukumar et al, 2012). In some cases, these changes can be linked to the establishment of symbiosis, such as rhizobia‐associated nodule formation (Hussain & Hasnain, 2011); in other cases, pathogens induce growth changes—for example, rhizogenesis during Agrobacterium rhizogenes infections (Falasca et al, 2000). Auxin can act as an immunity suppressor, mainly through antagonizing the salicylic acid response (Kunkel & Harper, 2018). Other effects seem to be a direct outcome of the established partnership. For example, root architecture changes or overall plant growth can be linked to endogenous auxin signaling modulation. These effects have been commonly reported in the last decades and have been thoroughly reviewed elsewhere for either pathogenic or mutualistic symbioses (Boivin et al, 2016; Kunkel & Harper, 2018).
While microbes widely use auxins to modulate plant development, they themselves can also respond to the presence of auxin. Some studies have reported changes in microbe pathogenicity or colonization habits in response to auxins. For example, just as mutant strains of Fusarium oxysporum with enhanced IAA production resulting from overexpression of auxin biosynthetic genes (tryptophan‐2‐monooxygenase and indole‐3‐acetamide hydrolase) show enhanced virulence of the fungi to Orobanche spp. (broomrapes; Cohen et al, 2002), mutant strains of the fungi Hebeloma cylindrosporum show an enhanced ability to establish mycorrhizal associations with pines (Tranvan et al, 2000). This has also been reported for bacteria such as Rhizobium etli (Spaepen, 2015). In the case of the phytopathogenic bacteria Pseudomonas syringae, auxin production leads not only to increased bacterial growth itself but also to defense inhibition and changes in plant development (Djami‐Tchatchou et al, 2020). These and other studies support the idea of auxin acting as a communication signal between plants and microbes in the rhizosphere (Spaepen, 2015). However, many of these effects can be directly attributed to microbe‐produced IAA; much less attention has been paid to the role of plant‐derived auxins in the rhizosphere. Nonetheless, there are some indications that microbes sense and use plant‐derived IAA during symbiosis initiation. For example, Lotus japonicum root hairs and epidermal cells show increased IAA production prior to mycorrhiza formation (Nadzieja et al, 2018), which is important for the association with the fungal symbiont, suggesting that plant IAA is part of the chemical dialog involved in this symbiosis (van Noorden et al, 2006; Nadzieja et al, 2018).
While numerous studies have shown auxin being used for interkingdom communication between plants and microbes, there are also pieces of evidence pointing to auxin performing an endogenous role in microbes unrelated to plant interactions. Several studies indicate that prokaryotes manifest clear responses to IAA. For example, transcriptional profiling after IAA treatment in Escherichia coli, Agrobacterium tumefaciens, or Bradyrhizobium japonicum shows a general shift towards stress response activation (Bianco et al, 2006; Yuan et al, 2008; Donati et al, 2013). Some of these responses may derive from co‐evolution with plant hosts: for example, Azospirillum brasilense shows extensive transcriptional changes after IAA addition, of which some can be linked to the establishment of bacterium–plant interactions (van Puyvelde et al, 2011). Yet, these responses may also be part of a more general metabolic homeostasis system, which would agree with the idea of IAA being used as a carbon source for bacteria (Jensen et al, 1995; Leveau & Lindow, 2005; Donoso et al, 2016; Conway et al, 2022). In any case, both possibilities are not mutually exclusive, and the widespread presence of IAA‐producing bacteria that are able to respond to its presence suggests that auxin is used as an interkingdom communication signal and may be a derived trait coopted from a commonly used endogenous metabolic signal.
Auxin action is not limited to unicellular or prokaryotic microbes, as it also triggers endogenous responses in fungi, including stress tolerance promotion (Fu et al, 2015; Hsiung et al, 2022). Other effects are the induction of pseudohyphal and hyphal growth in Saccharomyces cerevisiae and Candida albicans, respectively, both capable of IAA production but also of exogenous IAA active incorporation (Prusty et al, 2004; Rao et al, 2010). Morphogenic changes in dimorphic fungi, as well as the induction of stress tolerance mechanisms in yeasts, could be linked to competitive, defensive, or invasive behaviors, suggesting that IAA may be used as a neighboring cell detection signal in these organisms. Furthermore, fungi have been proposed to use IAA as a communication signal not only in plant‐fungal symbioses but also in fungal–fungal and fungi microalgae associations (Fu et al, 2015).
A key question is whether interkingdom auxin signaling is a shared property within the plant kingdom. Green algae–microbe interactions are well‐known in different ecological niches. For example, lichens, the alga–microbe symbioses, are known to produce auxins, mostly by the mycobiont (Epstein & Miles, 1967; Pichler et al, 2020). Auxins also seem positively to affect the growth of both fungi and algae, but the physiological meaning of these responses is unknown (Wang et al, 2010; Pichler et al, 2020). Free‐living chlorophyte algae can also perceive and respond to environmental auxins produced by nearby microbes, as has been shown for the unicellular algae Chlorella vulgaris (Trebouxiophyceae) and Scenodesmus sp. (Chlorophyceae; Bagwell et al, 2014).
While the molecular basis of free‐living green algae interactions with microbes has not been studied in depth, other eukaryotic algae have been shown to either use or respond to IAA present in their phycospheres (Amin et al, 2015; Labeeuw et al, 2016). This is the case of the cosmopolitan diatom Pseudo‐nitzschia multiseries, which associates with bacteria from the Sulfitobacter genus (Amin et al, 2015). The symbiosis formed involves bacterial IAA being uptaken by the alga, and Trp being released by it, and taken by the bacteria, forming an interspecies positive feedback loop dedicated to maintaining the association. Cells of P. multiseries are incapable of producing IAA but are able to respond to it, enhancing their growth and Trp production. Other nitrogenous compounds are also part of the metabolic exchange, but IAA and Trp seem, at least in part, to underlie the chemical communication necessary for the establishment and maintenance of the association. This example indicates that IAA can be used as a chemical signal in open water environments. Moreover, it pinpoints the importance of the phycosphere in enabling the exchange of molecules between neighboring cells, given that local concentrations created in this environment need to be significantly higher than bulk concentrations in water columns to be sensed and recognized by neighboring cells (Jonsson et al, 2009). Green algal microbiomes have recently been reported (Krohn‐Molt et al, 2017; Durán et al, 2022). While these microbes may be different from those of diatoms, their phycosphere does contain auxin‐producing bacteria. Moreover, green algae and land plants share at least a core of their microbiota (Durán et al, 2022), suggesting that a conserved core microbiome exists in the Viridiplantae. Among these, there are several auxin‐producing orders of bacteria. In summary, auxins seem to take part in the interkingdom communication occurring in very different ecosystems, including either land or aquatic environments.
Conclusions and future perspectives
In recent years, transcriptomic and genome‐based phylogenetic inference has opened the door to predict and possibly reconstruct the evolutionary history of the ancestral components of auxin biology (Bowman et al, 2017; Mutte et al, 2018; Fig 2). However, more functional validations of the predicted extant orthologs are required to confirm the conservation of the biochemical properties of all the components and the conservation of the biological role of the components using emerging streptophyte genetic models (Delaux et al, 2019; Martin‐Arevalillo et al, 2019; Skokan et al, 2019; Zhang et al, 2020; Zhou et al, 2020; Kawai et al, 2022).
The evolution of the auxin signaling pathway in land plants results from a co‐adaptation of a pre‐existing transcriptional machinery to respond to a chemical signal, auxin (Fig 3B). Functional studies in streptophyte algae are required to understand which features were regulated by the ancestral algae transcriptional network that gave rise to the land plants. Comparative studies will also be instrumental to unravel the contribution of transcriptional responses to auxin on the diversity of extant green lineages.
Given that auxin is a molecule widespread among many kingdoms of life, several aspects of auxin biology must be considered in the light of evolution and ecology. Here we reviewed evidence suggesting that auxin acts as a cue to support symbiotic partnerships between green lineages, fungi, and bacteria. These data support a plausible evolutionary scenario in which external auxin (in the phycosphere) may have long acted as a chemical messenger for local cellular responses (Fig 3), followed by a later consolidation of endogenous auxin responses that control plant growth and development. Further dissection of auxin responses in algal models, deepening of the understanding of fast, non‐transcriptional auxin responses, and focus on ecological interactions among plants (in the broadest sense) and microbes will help illuminate the natural history of the auxin giant.
Author contributions
Vanessa Polet Carrillo Carrasco: Conceptualization; writing – original draft; writing – review and editing. Jorge Hernandez‐Garcia: Funding acquisition; visualization; writing – review and editing. Sumanth K Mutte: Resources; formal analysis; writing – original draft; writing – review and editing. Dolf Weijers: Conceptualization; supervision; funding acquisition; project administration; writing – review and editing.
Disclosure and competing interests statement
The authors have no competing interests to disclose.
Supporting information
Expanded View Figures PDF
PDF+
Acknowledgements
This work was supported by a Netherlands Organization for Scientific Research (NWO) Open. Competition Domain Science—KLEIN grant (OCENW.KLEIN.02) to D.W., and by a Marie Skłodowska‐Curie Individual Fellowship (H2020‐MSCA‐IF‐2020) to J.H.‐G.
The EMBO Journal (2023) 42: e113018
Footnotes
Correction added on 15 March 2023, after first online publication: This reference callout has been corrected.
Correction added on 15 March 2023, after first online publication: This reference has been corrected.
References
- Abel S, Oeller PW, Theologis A (1994) Early auxin‐induced genes encode short‐lived nuclear proteins. Proc Natl Acad Sci USA 91: 326–330 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abel S, Nguyen MD, Theologis A (1995) The PS‐IAA4/5‐like family of early auxin‐inducible mRNAs in Arabidopsis thaliana . J Mol Biol 251: 533–549 [DOI] [PubMed] [Google Scholar]
- Amin SA, Hmelo LR, van Tol HM, Durham BP, Carlson LT, Heal KR, Morales RL, Berthiaume CT, Parker MS, Djunaedi B et al (2015) Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature 522: 98–101 [DOI] [PubMed] [Google Scholar]
- Arieti RS, Staiger CJ (2020) Auxin‐induced Actin cytoskeleton rearrangements require AUX1. New Phytol 226: 441–459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bagwell CE, Piskorska M, Soule T, Petelos A, Yeager CM (2014) A diverse assemblage of indole‐3‐acetic acid producing bacteria associate with unicellular green algae. Appl Biochem Biotechnol 173: 1977–1984 [DOI] [PubMed] [Google Scholar]
- le Bail A, Billoud B, Kowalczyk N, Kowalczyk M, Gicquel M, le Panse S, Stewart S, Scornet D, Cock JM, Ljung K et al (2010) Auxin metabolism and function in the multicellular brown alga Ectocarpus siliculosus . Plant Physiol 153: 128–144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbez E, Kubeš M, Rolčík J, Béziat C, Pěnčík A, Wang B, Rosquete MR, Zhu J, Dobrev PI, Lee Y et al (2012) A novel putative auxin carrier family regulates intracellular auxin homeostasis in plants. Nature 485: 119–122 [DOI] [PubMed] [Google Scholar]
- Basu S, Sun H, Brian L, Quatrano RL, Muday GK (2002) Early embryo development in Fucus distichus is auxin sensitive. Plant Physiol 130: 292–302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett T (2015) PIN proteins and the evolution of plant development. Trends Plant Sci 20: 498–507 [DOI] [PubMed] [Google Scholar]
- Bennett MJ, Marchant A, Green HG, May ST, Ward SP, Millner PA, Walker AR, Schulz B, Feldmann KA (1996) Arabidopsis AUX1 gene: a permease‐like regulator of root gravitropism. Science 273: 948–950 [DOI] [PubMed] [Google Scholar]
- Bennett T, Liu MM, Aoyama T, Bierfreund NM, Braun M, Coudert Y, Dennis RJ, O'Connor D, Wang XY, White CD et al (2014) Plasma membrane‐targeted PIN proteins drive shoot development in a moss. Curr Biol 24: 2776–2785 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bianco C, Imperlini E, Calogero R, Senatore B, Amoresano A, Carpentieri A, Pucci P, Defez R (2006) Indole‐3‐acetic acid improves Escherichia coli's defences to stress. Arch Microbiol 185: 373–382 [DOI] [PubMed] [Google Scholar]
- Billou I, Xu J, Wildwater M, Willemsen V, Paponov I, Frimi J, Heldstra R, Aida M, Palme K, Scheres B (2005) The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 433: 39–44 [DOI] [PubMed] [Google Scholar]
- Blázquez MA, Nelson DC, Weijers D (2020) Evolution of plant hormone response pathways. Annu Rev Plant Biol 71: 4–5 [DOI] [PubMed] [Google Scholar]
- Boer DR, Freire‐Rios A, van den Berg WAM, Saaki T, Manfield IW, Kepinski S, López‐Vidrieo I, Franco‐Zorrilla JM, de Vries SC, Solano R et al (2014) Structural basis for DNA binding specificity by the auxin‐dependent ARF transcription factors. Cell 156: 577–589 [DOI] [PubMed] [Google Scholar]
- Bogaert KA, Blommaert L, Ljung K, Beeckman T, de Clerck O (2019) Auxin function in the Brown alga Dictyota dichotoma . Plant Physiol 179: 280–299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bogaert KA, Blomme J, Beeckman T, de Clerck O (2022) Auxin's origin: do PILS hold the key? Trends Plant Sci 27: 227–236 [DOI] [PubMed] [Google Scholar]
- Boivin S, Fonouni‐Farde C, Frugier F (2016) How auxin and cytokinin phytohormones modulate root microbe interactions. Front Plant Sci 7: 1240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boot KJM, Libbenga KR, Hille SC, Offringa R, van Duijn B (2012) Polar auxin transport: an early invention. J Exp Bot 63: 4213–4218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowman JL, Kohchi T, Yamato KT, Jenkins J, Shu S, Ishizaki K, Yamaoka S, Nishihama R, Nakamura Y, Berger F et al (2017) Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell 171: 287–304.e15 [DOI] [PubMed] [Google Scholar]
- Bowman JL, Sandoval EF, Kato H (2021) On the evolutionary origins of land plant auxin biology. Cold Spring Harb Perspect Biol 13: a040048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boysen‐Jensen P (1910) Über die Leitung des phototropischen Reizes in Avena–Keimpflanzen. Ber Dtsch Bot Ges 28: 118–120 [Google Scholar]
- Casanova‐Sáez R, Mateo‐Bonmatí E, Ljung K (2021) Auxin metabolism in plants. Cold Spring Harb Perspect Biol 13: a039867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chanclud E, Morel J‐B (2016) Plant hormones: a fungal point of view. Mol Plant Pathol 17: 1289–1297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen R, Hilson P, Sedbrook J, Rosen E, Caspar T, Masson PH (1998) The Arabidopsis thaliana AGRAVITROPIC 1 gene encodes a component of the polar‐auxin‐transport efflux carrier. Proc Natl Acad Sci USA 95: 15112–15117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen BA, Amsellem Z, Maor R, Sharon A, Gressel J (2002) Transgenically enhanced expression of indole‐3‐acetic acid confers hypervirulence to plant pathogens. Phytopathology 92: 590–596 [DOI] [PubMed] [Google Scholar]
- Conway JM, Walton WG, Salas‐González I, Law TF, Lindberg CA, Crook LE, Kosina SM, Fitzpatrick CR, Lietzan AD, Northen TR et al (2022) Diverse MarR bacterial regulators of auxin catabolism in the plant microbiome. Nat Microbiol 7: 1817–1833 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooke TJ, Poli D, Sztein AE, Cohen JD (2002) Evolutionary patterns in auxin action. Plant Mol Biol 49: 319–338 [PubMed] [Google Scholar]
- Cox CE, Brandl MT, de Moraes MH, Gunasekera S, Teplitski M (2018) Production of the plant hormone auxin by salmonella and its role in the interactions with plants and animals. Front Microbiol 8: 2668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dai X, Mashiguchi K, Chen Q, Kasahara H, Kamiya Y, Ojha S, DuBois J, Ballou D, Zhao Y (2013) The biochemical mechanism of auxin biosynthesis by an Arabidopsis YUCCA flavin‐containing monooxygenase. J Biol Chem 288: 1448–1457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darwin C, Darwin F (1881) Das Bewegungsvermögen der Pflanzen, 1st edn. London: John Murray; [Google Scholar]
- Delaux PM, Hetherington AJ, Coudert Y, Delwiche C, Dunand C, Gould S, Kenrick P, Li FW, Philippe H, Rensing SA et al (2019) Reconstructing trait evolution in plant evo–devo studies. Curr Biol 29: R1110–R1118 [DOI] [PubMed] [Google Scholar]
- Dindas J, Scherzer S, Roelfsema MRG, von Meyer K, Müller HM, Al‐Rasheid KAS, Palme K, Dietrich P, Becker D, Bennett MJ et al (2018) AUX1‐mediated root hair auxin influx governs SCFTIR1/AFB‐type Ca2+ signaling. Nat Comm 9: 1174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Djami‐Tchatchou AT, Harrison GA, Harper CP, Wang R, Prigge MJ, Estelle M, Kunkel BN (2020) Dual role of auxin in regulating plant defense and bacterial virulence gene expression during pseudomonas syringae PtoDC3000 pathogenesis. Mol Plant Microbe Interact 33: 1059–1071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donati AJ, Lee HI, Leveau JHJ, Chang WS (2013) Effects of Indole‐3‐acetic acid on the transcriptional activities and stress tolerance of Bradyrhizobium japonicum . PLoS One 8: e76559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong L, Ma Y, Chen CY, Shen L, Sun W, Cui G, Naqvi NI, Deng YZ (2022) Identification and characterization of auxin/IAA biosynthesis pathway in the Rice blast fungus Magnaporthe oryzae . J Fungi 8: 208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donoso R, Leiva‐Novoa P, Zúñiga A, Timmermann T, Recabarren‐Gajardo G, González B (2016) Biochemical and genetic bases of Indole‐3‐acetic acid (auxin phytohormone) degradation by the plant‐growth‐promoting rhizobacterium Paraburkholderia phytofirmans PsJN. Appl Environ Microbiol 83: e01991‐16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dubey SM, Serre NBC, Oulehlová D, Vittal P, Fendrych M (2021) No time for transcription—rapid auxin responses in plants. Cold Spring Harb Perspect Biol 11: a039891 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durán P, Flores‐Uribe J, Wippel K, Zhang P, Guan R, Melkonian B, Melkonian M, Garrido‐Oter R (2022) Shared features and reciprocal complementation of the Chlamydomonas and Arabidopsis microbiota. Nat Comm 13: 1–14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eklund DM, Ishizaki K, Flores‐Sandoval E, Kikuchi S, Takebayashi Y, Tsukamoto S, Hirakawa Y, Nonomura M, Kato H, Kouno M et al (2015) Auxin produced by the indole‐3pyruvic acid pathway regulates development and gemmae dormancy in the liverwort Marchantia polymorpha . Plant Cell 27: 1650–1669 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epstein E, Miles PG (1967) Identification of Indole‐3‐acetic acid in the basidiomycete Schizophyllum commune on JSTOR. Plant Physiol 42: 911–914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans ML, Ishikawa H, Estelle MA (1994) Responses of Arabidopsis roots to auxin studied with high temporal resolution: comparison of wild type and auxin‐response mutants. Planta 194: 215–222 [Google Scholar]
- Falasca G, Reverberi M, Lauri P, Caboni E, de Stradis A, Altamura MM (2000) How agrobacterium rhizogenes triggers de novo root formation in a recalcitrant woody plant: an integrated histological, ultrastructural and molecular analysis. New Phytol 145: 77–93 [Google Scholar]
- Fendrych M, Akhmanova M, Merrin J, Glanc M, Hagihara S, Takahashi K, Uchida N, Torii KU, Friml J (2018) Rapid and reversible root growth inhibition by TIR1 auxin signalling. Nat Plants 4: 453–459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finet C, Berne‐Dedieu A, Scutt CP, Marlétaz F (2013) Evolution of the ARF gene family in land plants: old domains, new tricks. Mol Biol Evol 30: 45–56 [DOI] [PubMed] [Google Scholar]
- Flores‐Sandoval E, Eklund DM, Bowman JL (2015) A simple auxin transcriptional response system regulates multiple morphogenetic processes in the liverwort Marchantia polymorpha . PLoS Genet 11: e1005207 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freire‐Rios A, Tanaka K, Crespo I, van der Wijk E, Sizentsova Y, Levitsky V, Lindhoud S, Fontana M, Hohlbein J, Roeland Boer D et al (2020) Architecture of DNA elements mediating ARF transcription factor binding and auxin‐responsive gene expression in Arabidopsis. Proc Natl Acad Sci USA 117: 24557–24566 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friml J (2021) Fourteen stations of auxin. Cold Spring Harb Perspect Biol 14: a039859 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friml J, Gallei M, Gelová Z, Johnson A, Mazur E, Monzer A, Rodriguez L, Roosjen M, Verstraeten I, Živanović BD et al (2022) ABP1–TMK auxin perception for global phosphorylation and auxin canalization. Nature 609: 575–581 [DOI] [PubMed] [Google Scholar]
- Fu SF, Wei JY, Chen HW, Liu YY, Lu HY, Chou JY (2015) Indole‐3‐acetic acid: a widespread physiological code in interactions of fungi with other organisms. Plant Signal Behav 10: e1048052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gälweiler L, Guan C, Müller A, Wisman E, Mendgen K, Yephremov A, Palme K (1998) Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 282: 2226–2230 [DOI] [PubMed] [Google Scholar]
- Gray WM, Kepinski S, Rouse D, Leyser O, Estelle M (2001) Auxin regulates SCFTIR1dependent degradation of AUX/IAA proteins. Nature 414: 271–276 [DOI] [PubMed] [Google Scholar]
- Hammes UZ, Murphy AS, Schwechheimer C (2022) Auxin transporters—a biochemical view. Cold Spring Harb Perspect Biol 14: a039875 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han H, Verstraeten I, Roosjen M, Mazur E, Rýdza N, Hajný J, Ötvös K, Weijers D, Friml J (2021) Rapid auxin‐mediated phosphorylation of myosin regulates trafficking and polarity in Arabidopsis. bioRxiv 10.1101/2021.04.13.439603 [PREPRINT] [DOI] [Google Scholar]
- Hess S, Williams SK, Busch A, Irisarri I, Delwiche CF, de Vries S, Darienko T, Roger AJ, Archibald JM, Buschmann H et al (2022) A phylogenomically informed five‐order system for the closest relatives of land plants. Curr Biol 32: 4473–4482 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsiung RT, Chiu MC, Chou JY (2022) Exogenous Indole‐3‐acetic acid induced ethanol tolerance in phylogenetically diverse Saccharomycetales yeasts. Microbes Environ 37: ME21053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hussain A, Hasnain S (2011) Interactions of bacterial cytokinins and IAA in the rhizosphere may alter phytostimulatory efficiency of rhizobacteria. World J Microbiol Biotechnol 27: 2645–2654 [Google Scholar]
- Jensen JB, Egsgaard H, van Onckelen H, Jochimsen BU (1995) Catabolism of indole‐3acetic acid and 4‐ and 5‐chloroindole‐3‐acetic acid in Bradyrhizobium japonicum . J Bacteriol 177: 5762–5766 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jonsson PR, Pavia H, Toth G (2009) Formation of harmful algal blooms cannot be explained by allelopathic interactions. Proc Natl Acad Sci USA 106: 11177–11182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones VAS, Dolan L (2012) The evolution of root hairs and rhizoids. Ann Bot 110: 205–212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamimoto Y, Terasaka K, Hamamoto M, Takanashi K, Fukuda S, Shitan N, Sugiyama A, Suzuki H, Shibata D, Wang B et al (2012) Arabidopsis ABCB21 is a facultative auxin importer/exporter regulated by cytoplasmic auxin concentration. Plant Cell Physiol 53: 2090–2100 [DOI] [PubMed] [Google Scholar]
- Kato H, Ishizaki K, Kouno M, Shirakawa M, Bowman JL, Nishihama R, Kohchi T (2015) Auxin‐mediated transcriptional system with a minimal set of components is critical for morphogenesis through the life cycle in Marchantia polymorpha . PLoS Genet 11: e1005084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kato H, Nishihama R, Weijers D, Kohchi T (2018) Evolution of nuclear auxin signaling: lessons from genetic studies with basal land plants. J Exp Bot 69: 291–301 [DOI] [PubMed] [Google Scholar]
- Kato H, Mutte SK, Suzuki H, Crespo I, Das S, Radoeva T, Fontana M, Yoshitake Y, Hainiwa E, van den Berg W et al (2020) Design principles of a minimal auxin response system. Nat Plants 6: 473–482 [DOI] [PubMed] [Google Scholar]
- Kawai J, Kanazawa M, Suzuki R, Kikuchi N, Hayakawa Y, Sekimoto H (2022) Highly efficient transformation of the model zygnematophycean alga Closterium peracerosum‐strigosum‐littorale complex by square‐pulse electroporation. New Phytol 233: 569–578 [DOI] [PubMed] [Google Scholar]
- Kepinski S, Leyser O (2005) The Arabidopsis F‐box protein TIR1 is an auxin receptor. Nature 435: 446–451 [DOI] [PubMed] [Google Scholar]
- Klambt D, Knauth B, Dittmann I (1992) Auxin dependent growth of rhizoids of Chara globularis . Physiol Plant 85: 537–540 [Google Scholar]
- Kögl F, Kostermans DGFR (1934) Hetero‐auxin als Stoffwechselprodukt niederer pflanzlicher Organismen. Isolierung aus Hefe. 13. Mitteilung über pflanzliche Wachstumsstoffe. Hoppe‐Seyler's Z Physiol Chem 228: 113–121 [Google Scholar]
- Korasick DA, Westfall CS, Lee SG, Nanao MH, Dumas R, Hagen G, Guilfoyle TJ, Jez JM, Strader LC (2014) Molecular basis for AUXIN RESPONSE FACTOR protein interaction and the control of auxin response repression. Proc Natl Acad Sci USA 111: 5427–5432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krohn‐Molt I, Alawi M, Förstner KU, Wiegandt A, Burkhardt L, Indenbirken D, Thieß M, Grundhoff A, Kehr J, Tholey A et al (2017) Insights into microalga and bacteria interactions of selected phycosphere biofilms using metagenomic, transcriptomic, and proteomic approaches. Front Microbiol 8: 1941 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunkel BN, Harper CP (2018) The roles of auxin during interactions between bacterial plant pathogens and their hosts. J Exp Bot 69: 245–254 [DOI] [PubMed] [Google Scholar]
- Labeeuw L, Khey J, Bramucci AR, Atwal H, de La Mata AP, Harynuk J, Case RJ (2016) Indole‐3‐acetic acid is produced by Emiliania huxleyi coccolith‐bearing cells and triggers a physiological response in bald cells. Front Microbiol 7: 828 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landberg K, Pederson ERA, Viaene T, Bozorg B, Friml J, Jönsson H, Thelander M, Sundberg E (2013) The Moss Physcomitrella patens reproductive organ development is highly organized, affected by the two SHI/STY genes and by the level of active auxin in the SHI/STY expression domain. Plant Physiol 162: 1406–1419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavy M, Prigge MJ, Tao S, Shain S, Kuo A, Kirchsteiger K, Estelle M (2016) Constitutive auxin response in Physcomitrella reveals complex interactions between aux/IAA and ARF proteins. Elife 5: e13325 [DOI] [PMC free article] [PubMed] [Google Scholar]
- LeClere S, Tellez R, Rampey RA, Matsuda SPT, Bartel B (2002) Characterization of a family of IAA‐amino acid conjugate hydrolases from Arabidopsis . J Biol Chem 277: 20446–20452 [DOI] [PubMed] [Google Scholar]
- Leontovyčová H, Trdá L, Dobrev PI, Šašek V, Gay E, Balesdent MH, Burketová L (2020) Auxin biosynthesis in the phytopathogenic fungus Leptosphaeria maculans is associated with enhanced transcription of indole‐3‐pyruvate decarboxylase LmIPDC2 and tryptophan aminotransferase LmTAM1. Res Microbiol 171: 174–184 [DOI] [PubMed] [Google Scholar]
- Leveau JHJ, Lindow SE (2005) Utilization of the plant hormone indole‐3‐acetic acid for growth by pseudomonas putida strain 1290. Appl Environ Microbiol 71: 2365–2371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leyser O (2018) Auxin Signaling. Plant Physiol 176: 465–479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Gallei M, Friml J (2022) Bending to auxin: fast acid growth for tropisms. Trends Plant Sci 27: 440–449 [DOI] [PubMed] [Google Scholar]
- Lin W, Zhou X, Tang W, Takahashi K, Pan X, Dai J, Ren H, Zhu X, Pan S, Zheng H et al (2021) TMK‐based cell‐surface auxin signalling activates cell‐wall acidification. Nature 599: 278–282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludwig‐Müller J, Jülke S, Bierfreund NM, Decker EL, Reski R (2009) Moss (Physcomitrella patens) GH3 proteins act in auxin homeostasis. New Phytol 181: 323–338 [DOI] [PubMed] [Google Scholar]
- Martin‐Arevalillo R, Thévenon E, Jégu F, Vinos‐Poyo T, Vernoux T, Parcy F, Dumas R (2019) Evolution of the auxin response factors from charophyte ancestors. PLoS Genet 15: e1008400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mashiguchi K, Tanaka K, Sakai T, Sugawara S, Kawaide H, Natsume M, Hanada A, Yaeno T, Shirasu K, Yao H et al (2011) The main auxin biosynthesis pathway in Arabidopsis. Proc Natl Acad Sci USA 108: 18512–18517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mellor N, Band LR, Pěňík A, Novák O, Rashed A, Holman T, Wilson MH, Vo U, Bishopp A, King JR et al (2016) Dynamic regulation of auxin oxidase and conjugating enzymes AtDAO1 and GH3 modulates auxin homeostasis. Proc Natl Acad Sci USA 113: 11022–11027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michniewicz M, Brewer PB, Friml J (2007) Polar auxin transport and asymmetric auxin distribution. Arabidopsis Book 5: e0108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mikami K, Mori IC, Matsuura T, Ikeda Y, Kojima M, Sakakibara H, Hirayama T (2016) Comprehensive quantification and genome survey reveal the presence of novel phytohormone action modes in red seaweeds. J Appl Phycol 28: 2539–2548 [Google Scholar]
- Morffy N, Strader LC (2020) Old town roads: routes of auxin biosynthesis across kingdoms. Curr Opin Plant Biol 55: 21–27 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morffy N, Strader LC, Fernie AR, Wen W (2020) Old town roads: routes of auxin biosynthesis across kingdoms this review comes from a themed issue on physiology and metabolism. Curr Opin Plant Biol 2020: 21–27 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutte SK (2020) Evolutionary analysis of a billion years of auxin biology. Dissertation/Doctoral Thesis, Wageningen University, Wageningen, The Netherlands
- Mutte S, Kato H, Rothfels C, Melkonian M, Wong GK‐S, Weijers D (2018) Origin and evolution of the nuclear auxin response system. Elife 7: e33399 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadzieja M, Kelly S, Stougaard J, Reid D (2018) Epidermal auxin biosynthesis facilitates rhizobial infection in Lotus japonicus . Plant J 95: 101–111 [DOI] [PubMed] [Google Scholar]
- Nanao MH, Vinos‐Poyo T, Brunoud G, Thévenon E, Mazzoleni M, Mast D, Lainé S, Wang S, Hagen G, Li H et al (2014) Structural basis for oligomerization of auxin transcriptional regulators. Nat Commun 5: 3617 [DOI] [PubMed] [Google Scholar]
- Noh B, Murphy AS, Spalding EP (2001) Multidrug resistance–like genes of arabidopsis required for auxin transport and auxin‐mediated development. Plant Cell 13: 2441–2454 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Noorden GE, Ross JJ, Reid JB, Rolfe BG, Mathesius U (2006) Defective Long‐distance auxin transport regulation in the Medicago truncatula super numeric nodules mutant. Plant Physiol 140: 1494–1506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohtaka K, Hori K, Kanno Y, Seo M, Ohta H (2017) Primitive auxin response without TIR1 and aux/IAA in the charophyte alga Klebsormidium nitens . Plant Physiol 174: 1621–1632 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okrent RA, Brooks MD, Wildermuth MC (2009) Arabidopsis GH3.12 (PBS3) conjugates amino acids to 4‐substituted benzoates and is inhibited by salicylate. J Biol Chem 284: 9742–9754 [DOI] [PMC free article] [PubMed] [Google Scholar]
- One Thousand Plant Transcriptomes Initiative (2019) One thousand plant transcriptomes and the phylogenomics of green plants. Nature 574: 679–685 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pichler G, Stöggl W, Candotto Carniel F, Muggia L, Ametrano CG, Holzinger A, Tretiach M, Kranner I (2020) Abundance and extracellular release of phytohormones in aeroterrestrial microalgae (Trebouxiophyceae, Chlorophyta) as a potential chemical signaling source. J Phycol 56: 1295–1307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prigge MJ, Lavy M, Ashton NW, Estelle M (2010) Physcomitrella patens auxin‐resistant mutants affect conserved elements of an auxin‐signaling pathway. Curr Biol 20: 1907–1912 [DOI] [PubMed] [Google Scholar]
- Prigge MJ, Platre M, Kadakia N, Zhang Y, Greenham K, Szutu W, Pandey BK, Bhosale RA, Bennett MJ, Busch W et al (2020) Genetic analysis of the Arabidopsis TIR1/AFB auxin receptors reveals both overlapping and specialized functions. Elife 9: e54740 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prusty R, Grisafi P, Fink GR (2004) The plant hormone indoleacetic acid induces invasive growth in Saccharomyces cerevisiae . Proc Natl Acad Sci USA 101: 4153–4157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Puyvelde S, Cloots L, Engelen K, Das F, Marchal K, Vanderleyden J, Spaepen S (2011) Transcriptome analysis of the rhizosphere bacterium Azospirillum brasilense reveals an extensive auxin response. Microb Ecol 61: 723–728 [DOI] [PubMed] [Google Scholar]
- Qi L, Kwiatkowski M, Chen H, Hoermayer L, Sinclair S, Zou M, del Genio CI, Kubeš MF, Napier R, Jaworski K et al (2022) Adenylate cyclase activity of TIR1/AFB auxin receptors in plants. Nature 611: 133–138 [DOI] [PubMed] [Google Scholar]
- Rampey RA, LeClere S, Kowalczyk M, Ljung K, Sandberg G, Bartel B (2004) A family of auxin‐conjugate hydrolases that contributes to free Indole‐3‐acetic acid levels during Arabidopsis germination. Plant Physiol 135: 978–988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao RP, Hunter A, Kashpur O, Normanly J (2010) Aberrant synthesis of indole‐3‐acetic acid in Saccharomyces cerevisiae triggers morphogenic transition, a virulence trait of pathogenic fungi. Genetics 185: 211–220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rensing SA (2018) Great moments in evolution: the conquest of land by plants. Curr Opin Plant Biol 42: 49–54 [DOI] [PubMed] [Google Scholar]
- Sandan T, Ogura T (1957) Physiological studies on growth and morphogenesis of the isolated plant cell cultured in vitro III. The effects of pH, auxin and metabolic inhibitors. Bot Mag Tokyo 70: 125–130 [Google Scholar]
- Scossa F, Fernie AR (2021) Ancestral sequence reconstruction ‐ an underused approach to understand the evolution of gene function in plants? Comput Struct Biotechnol J 19: 1579–1594 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serre NBC, Kralík D, Yun P, Slouka Z, Shabala S, Fendrych M (2021) AFB1 controls rapid auxin signalling through membrane depolarization in Arabidopsis thaliana root. Nat Plants 7: 1229–1238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skokan R, Medvecká E, Viaene T, Vosolsobě S, Zwiewka M, Müller K, Skůpa P, Karady M, Zhang Y, Janacek DP et al (2019) PIN‐driven auxin transport emerged early in streptophyte evolution. Nat Plants 5: 1114–1119 [DOI] [PubMed] [Google Scholar]
- Spaepen S (2015) Plant hormones produced by microbes. In Principles of plant‐microbe interactions: microbes for sustainable agriculture, Lugtenberg B (ed), pp 247–256. Cham: Springer International Publishing; [Google Scholar]
- Staswick PE, Serban B, Rowe M, Tiryaki I, Maldonado MT, Maldonado MC, Suza W (2005) Characterization of an Arabidopsis enzyme family that conjugates amino acids to Indole‐3‐acetic acid. Plant Cell 17: 616–627 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stepanova AN, Yun J, Robles LM, Novak O, He W, Guo H, Ljung K, Alonso JM (2011) The Arabidopsis YUCCA1 flavin monooxygenase functions in the Indole‐3‐pyruvic acid branch of auxin biosynthesis. Plant Cell 23: 3961–3973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sukumar P, Legué V, Vayssières A, Martin F, Tuskan GA, Kalluri UC (2012) Involvement of auxin pathways in modulating root architecture during beneficial plant‐microorganism interactions. Plant Cell Environ 36: 909–919 [DOI] [PubMed] [Google Scholar]
- Szemenyei H, Hannon M, Long JA (2008) TOPLESS mediates auxin‐dependent transcriptional repression during Arabidopsis embryogenesis. Science 319: 1384–1386 [DOI] [PubMed] [Google Scholar]
- Tan X, Irina Calderon‐Villalobos LA, Sharon M, Zheng C, Robinson CV, Estelle M, Zheng N (2007) Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature 446: 640–645 [DOI] [PubMed] [Google Scholar]
- Tao Y, Ferrer JL, Ljung K, Pojer F, Hong F, Long JA, Li L, Moreno JE, Bowman ME, Ivans LJ et al (2008) Rapid synthesis of auxin via a new tryptophan‐dependent pathway is required for shade avoidance in plants. Cell 133: 164–176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terol J, Domingo C, Talón M (2006) The GH3 family in plants: genome wide analysis in rice and evolutionary history based on EST analysis. Gene 371: 279–290 [DOI] [PubMed] [Google Scholar]
- Thimann KV, Sweeney BM (1937) The effect of auxins upon protoplasmic streaming. J Gen Physiol 21: 123–135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Timme RE, Bachvaroff TR, Delwiche CF (2012) Broad phylogenomic sampling and the sister lineage of land plants. PLoS One 7: e29696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tranvan H, Habricot Y, Jeannette E, Gay G, Sotta B (2000) Dynamics of symbiotic establishment between an IAA‐overproducing mutant of the ectomycorrhizal fungus Hebeloma cylindrosporum and Pinus pinaster . Tree Physiol 20: 123–129 [DOI] [PubMed] [Google Scholar]
- Ulmasov T, Hagen G, Guilfoyle TJ (1997a) ARF1, a transcription factor that binds to auxin response elements. Science 276: 1865–1868 [DOI] [PubMed] [Google Scholar]
- Ulmasov T, Murfett J, Hagen G, Guilfoyle TJ (1997b) Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 9: 1963–1971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulmasov T, Hagen G, Guilfoyle TJ (1999) Activation and repression of transcription by auxin‐response factors. Proc Natl Acad Sci USA 96: 5844–5849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verrier PJ, Bird D, Burla B, Dassa E, Forestier C, Geisler M, Klein M, Kolukisaoglu Ü, Lee Y, Martinoia E et al (2008) Plant ABC proteins – a unified nomenclature and updated inventory. Trends Plant Sci 13: 151–159 [DOI] [PubMed] [Google Scholar]
- Vosolsobě S, Vosolsobě S, Skokan R, Skokan R, Petrášek J, Petrášek J (2020) The evolutionary origins of auxin transport: what we know and what we need to know. J Exp Bot 71: 3287–3295 [DOI] [PubMed] [Google Scholar]
- de Vries J, Archibald JM (2018) Plant evolution: landmarks on the path to terrestrial life. New Phytol 217: 1428–1434 [DOI] [PubMed] [Google Scholar]
- Wang XY, Wei XL, Luo H, Kim JA, Jeon HS, Koh YJ, Hur J‐S (2010) Plant hormones promote growth in lichen‐forming fungi. Mycobiology 38: 176–179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang C, Liu Y, Li SS, Han GZ (2014) Origin of plant auxin biosynthesis in charophyte algae. Trends Plant Sci 19: 741–743 [DOI] [PubMed] [Google Scholar]
- Weijers D, Wagner D (2016) Transcriptional responses to the auxin hormone. Annu Rev Plant Biol 67: 539–574 [DOI] [PubMed] [Google Scholar]
- Went FW, Thimann KV (1937) Phytohormones. New York, NY: Macmillan; [Google Scholar]
- Wodniok S, Brinkmann H, Glöckner G, Heidel AJ, Philippe H, Melkonian M, Becker B (2011) Origin of land plants: do conjugating green algae hold the key? BMC Evol Biol 11: 104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood NL, Berliner MD (1979) Effects of indoleacetic acid on the desmid Micrasterias thomasiana . Plant Sci Lett 16: 285–289 [Google Scholar]
- Yuan ZC, Haudecoeur E, Faure D, Kerr KF, Nester EW (2008) Comparative transcriptome analysis of Agrobacterium tumefaciens in response to plant signal salicylic acid, indole‐3‐acetic acid and γ‐amino butyric acid reveals signalling cross‐talk and Agrobacterium–plant co‐evolution. Cell Microbiol 10: 2339–2354 [DOI] [PubMed] [Google Scholar]
- Yue J, Hu X, Huang J (2014) Origin of plant auxin biosynthesis. Trends Plant Sci 19: 764–770 [DOI] [PubMed] [Google Scholar]
- Zhang P, Jin T, Sahu SK, Xu J, Shi Q, Liu H, Wang Y (2019) The distribution of tryptophan‐dependent Indole‐3‐Acetic Acid synthesis pathways in bacteria unraveled by large‐scale genomic analysis. Molecules 24: 19824–19829 ‡ Correction added on 15 March 2023, after first online publication: This reference has been corrected. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Rodriguez L, Li L, Zhang X, Friml J (2020) Functional innovations of PIN auxin transporters mark crucial evolutionary transitions during rise of flowering plants. Sci Adv 6: eabc8895 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou H, von Schwartzenberg K, Buschmann H (2020) Zygnematophyceae: from living algae collections to the establishment of future models. J Exp Bot 71: 3296–3304 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Expanded View Figures PDF
PDF+