

Abstract
Background
Potassium channels (KCa3.1; Kv1.3; Kir2.1) are necessary for microglial activation, a pivotal requirement for the development of Perioperative Neurocognitive Disorders (PNDs). We previously reported on the role of microglial Kv1.3 for PNDs; the present study sought to determine whether inhibiting KCa3.1 channel activity affects neuroinflammation and prevents development of PND.
Methods
Mice (wild-type [WT] and KCa3.1−/−) underwent aseptic tibial fracture trauma under isoflurane anesthesia or received anesthesia alone. WT mice received either TRAM34 (a specific KCa3.1 channel inhibitor) dissolved in its vehicle (miglyol) or miglyol alone. Spatial memory was assessed in the Y-maze paradigm 6 h post-surgery/anesthesia. Circulating interleukin-6 (IL-6) and high mobility group box-1 protein (HMGB1) were assessed by ELISA, and microglial activitation Iba-1 staining.
Results
In WT mice surgery induced significant cognitive decline in the Y-maze test, p = 0.019), microgliosis (p = 0.001), and increases in plasma IL-6 (p = 0.002) and HMGB1 (p = 0.001) when compared to anesthesia alone. TRAM34 administration attenuated the surgery-induced changes in cognition, microglial activation, and HMGB1 but not circulating IL-6 levels. In KCa3.1−/− mice surgery neither affected cognition nor microgliosis, although circulating IL-6 levels did increase (p < 0.001).
Conclusion
Similar to our earlier report with Kv1.3, perioperative microglial KCa3.1 blockade decreases immediate perioperative cognitive changes, microgliosis as well as the peripheral trauma marker HMGB1 although surgery-induced IL-6 elevation was unchanged. Future research should address whether a synergistic interaction exists between blockade of Kv1.3 and KCa3.1 for preventing PNDs.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12871-023-02030-2.
Keywords: Neuroinflammation, Surgery, Post-operative cognitive decline, Perioperative neurocognitive disorders, Inflammation, Cognition, Anesthesia, Microglia
Background
Perioperative neurocognitive disorder (PND), first described in 1887, [1] is a frequent misdiagnosed complication [2–4]. While several etiological factors have been investigated [5–7], the most plausible explanation implicates trauma-induced inflammation for the development of PND [8–10].
During surgical incision, traumatized tissues release high mobility group box protein1 (HMGB1) into the bloodstream [8]. This damage-associated molecular pattern binds to pattern recognition receptors on circulating, CCR2+ bone marrow-derived monocytes (BM-DMs), triggering the nuclear translocation of the transcription factor NF-kB which activates gene expression and release of pro-inflammatory cytokines including IL-6 and IL-1β [8] and leads to blood-brain barrier disruption [9]. Within the brain parenchyma, the chemokine MCP-1 (also referred to as CCL2) is upregulated and, by signaling through its receptor, attracts CCR2+ BM-DMs [10]; the resulting influx of BM-DMs into brain activates quiescent microglia. Together, BM-DMs and activated microglia release HMGB1, IL-6, and IL-1β, thereby disrupting long-term potentiation and the synaptic plasticity involved in the cognitive functions of learning and memory [11–13]. Exaggerated or unresolved inflammation promotes the development of PND [4, 14–16].
Among strategies that have been investigated to mitigate the development of PND, preventing the activation of microglia may be one of the most promising. Microglia surveil the milieu and are activated by changes in its environment. When differentiated into the pro-inflammatory (“M1”) phenotype, the activated microglia synthesize and release cytokines that propagate the inflammatory process; conversely, when transformed into the anti-inflammatory phenotype (“M2”) microglia support regeneration. M1 microglia play an active role in the development of PNDs following peripheral surgery [10].
Microglia express multiple potassium (K+) channels (KCa3.1; Kv1.3; Kir2.1), which are up-regulated during activation and participate in microglial calcium signaling and neuroinflammation [17]. We previously reported that Kv1.3 channel blockade mitigated PND development [18]. KCa3.1 is a voltage-independent, homo-tetrameric potassium channel which is constitutively associated with its calcium sensor calmodulin and therefore opens in response to increased intracellular calcium (above ~ 150 nM). In turn, a negative membrane potential is maintained through K+ efflux [17, 19]. The channel is expressed on proliferating fibroblasts, on dedifferentiated vascular smooth muscle cells, and on immune cells including microglia and macrophages, activated CCR7+ T cells and IgD+ B cells [17, 19].
In microglia, KCa3.1 has been shown to be involved in respiratory bursting, migration, proliferation, and nitric oxide production, as well neuroapoptosis in organotypic hippocampal slices, suggesting that KCa3.1 suppression may be useful for in neurological diseases featuring microglial activation [19–21]. TRAM34, a small molecule inhibitor, blocks the KCa3.1 channel with an IC50 of 20 nmol/L and exhibits 200- to 1,500-fold selectivity over other K+ channels [19].
The present study investigated whether blockade of microglial KCa3.1 channel activity impacts the trauma-induced inflammatory cascade and protect against the development of PND.
Methods
Animal care
Experimental procedures involving animals were approved by the Animal Care Committee of the Université Libre de Bruxelles. (CEBEA-IBMM 2019-24-105).
Twelve weeks old C57BL/6J mice (Charles River Laboratories, France) and KCa3.1−/− mice (genetic background: C57BL/6J; Mouse Biology Program UC Davis) [20] were separately group-housed with 12 h light/dark cycles in a temperature-controlled environment with ad libitum access to standard rodent chow and water in our animal research facilities during one week prior to experiments.
Experimental groups
12-week-old wild type mice were randomly assigned to surgery (anesthesia followed by surgery) or anesthesia alone groups that either received vehicle or TRAM34 (T) treatment:
Group 1: Anesthesia (+ vehicle).
Group 2: Surgery (+ vehicle).
Group 3: Anesthesia + T(TRAM34).
Group 4: Surgery + T(TRAM34).
12 weeks-old KCa3.1−/− mice were randomized to surgery (anesthesia followed by surgery) or anesthesia alone groups:
Group 1: Anesthesia (without vehicle or TRAM34).
Group 2: Surgery (without vehicle or TRAM34).
In all experiments, 8 to 10 mice were included per group.
Experiments were conducted in the same animal surgery room in the morning. (Fig. 1)
Fig. 1.

Timeline of experiments performed in WT mice. A. Timeline of the experiments to measure cognitive outcome. B. Timeline of the experiments to measure peripheral and hippocampal parameters. Tibia fracture was applied after anesthesia to the surgery groups. C. Illustration of the 3-armed Y-maze device. Each arm had the following dimensions: 35 × 6 cm; wall height: 15 cm. Angles between arms were 120 °. Each mouse was placed in the middle of the Y-maze (as indicated on the image)
Anesthesia
Mice randomized to ‘anesthesia groups’ were subjected to inhalation anesthesia (3% isoflurane in 30% FiO2) as well as subcutaneous buprenorphine (0.1 mg/kg).
Surgical procedure
Under general anesthesia with 3% isoflurane in 30% FiO2, rodents underwent an open tibial fracture of the left hind paw with intramedullary fixation under aseptic conditions. Briefly, buprenorphine (0.1 mg/kg) was injected subcutaneously to provide analgesia after the induction of anesthesia and before skin incision. A 20 G pin was then inserted in the intramedullary canal of the left tibia, and osteotomy was performed after the periosteum was stripped. During the surgical procedure that lasted approximately 10 min, temperature was monitored and maintained at 37 °C with the aid of warming pads [8, 13].
Intra-peritoneal injections
TRAM34 and its vehicle miglyol were provided by the Department of Pharmacology, University of California Davis, Davis, CA, USA (Prof. Heike Wulff).
TRAM34 was synthesized as previously described [19] and was dissolved at a concentration of 8 mg/ml in miglyol 812 neutral oil (Neobee M5®; Spectrum Chemicals), a low viscosity vehicle that is used as a pharmaceutical excipient and well tolerated following i.p., s.c. or oral administration.
TRAM34, 40 mg/kg or miglyol alone were injected intraperitoneally approximately 1 min before surgery.
Other reagents are detailed in the respective methodological subsections.
Assessment of cognitive score by the Y-maze test
“Y-maze test evaluates the mice’s willingness to explore new environments. Rodents typically prefer to investigate a new arm of the maze rather than returning to one that was previously visited. Many parts of the brain, including the hippocampus are involved in this spatial memory test [22].
Mice were not trained before experiments. Six hours after surgery, mice were individually placed in the middle of a Y designed maze (each arm designated A/B/C) (Fig. 1C) and their movement was recorded for 5 min with a camera (Sony DSC-HX50). After 5 min, mice were taken out of the Y-maze and sacrificed. The Y maze was devoid of food and was thoroughly cleaned with ethanol after each recording. The following parameters from the recording were manually obtained and blindly analyzed by an independent researcher: total distance traveled in the maze, total number of arm entries, number of entries made into each arm, and the sequential list of arms entered to assess number of alternations made.
In this study, cognitive score refers to the number of alternations in the Y-maze test.
A significant decrease in alternations is a sign of a cognitive deficit [22].”
Hippocampal microglial presence
After the behavioral assessment was completed, mice were sacrificed and tissue samples collected as previously described [17].
To analyze microglial proliferation, hippocampus was harvested and fixed in 4% formaldehyde (Klinpath, Duiven, Netherlands) and embedded in paraffin. Three µm-thick biopsy slices were cut with a microtome Leica RM2255 (Wetzlar, Germany) and mounted on microscope slides. Deparaffinised tissue sections were pretreated with hot citrate buffer pH6 and endoperoxydases were inactivated by a methanol/H2O2 treatment. After blocking (ScyTek Laboratories, UT, USA), hippocampal sections were incubated for 30 min at room temperature with a specific primary antibody (Anti-Iba1 antibody [EPR16588], Abcam, Cambridge, UK) at a 1:500 dilution. Signal amplification was performed by incubation with the complex UltrasenseStreptavidinePeroxydase RTU (ScyTek Laboratories, UT, USA). Revelation was performed with diaminobenzidine (DAB) (Biogenex, CA, USA). Slides were counterstained with hematoxylin (Cell Signaling, MA, USA). Five fields of each stained slide were acquired with 20x objective. The area percentage of Iba1 staining was obtained by colour deconvolution with Image J (NIH, Md, USA).
Systemic inflammatory response
At the time of sacrifice, blood was harvested in heparin-coated syringes from the inferior vena cava of wild-type mice under terminal isoflurane anesthesia. Plasma was collected after centrifugation of the blood at 10,000 x g for 10 min at room temperature and stored at -80 °C for later analysis. Plasma IL-6 and HMGB1 concentrations were measured using the IL-6 ELISA kit from Millipore corporation (MO, USA) and mouse HMGB-1 Elisa kit (Novus Biologicals (Biotechne, MN, USA), respectively. Plasma samples were diluted twice with the Sample Diluent Buffer of the kit. The standard curve and samples were performed in duplicates. The absorbance was read at 450 nm with an Emax Plus Microplate Reader (Molecular Devices, CA, USA). The standard curve and the samples were performed in duplicates and the absorbance was read at 450 nm.
Statistics
Analysis for normality revealed a non-parametric distribution and the data are displayed graphically with boxplots based on the calculation of the median (P50) and the interquartile range (IQR: P25–P75). Anesthesia and surgery experimental groups were compared by the Kruskal-Wallis test. The p-value was obtained by the non-parametric Wilcoxon Rank Sum Test. The results were considered as significant below the 5% critical level (p < 0.05). Calculations were carried out by Sigma Plot version 12.0 (Systat Software, Chicago, IL).
Results
In wild-type (WT) mice, in the absence of TRAM34 administration, surgery was associated with a decrease in cognitive score (alternations in Y-maze), suggesting the presence of cognitive decline 6 h post-operatively (P = 0.019).
In parallel, surgery also induced an increase in plasma IL-6 (P = 0.002), HMGB1 (P = 0.001), and microgliosis (P = 0.001) when compared to anesthesia alone (Fig. 2 and S1; Fig. 3).
Fig. 2.

Anesthesia/ surgery-dependent changes in cognitive decline, peripheral inflammation and central proliferation of microglia in wild type mice (10 animals in each experimental group) treated with miglyol or miglyol + TRAM34 (T). Experimental groups included 10 animals. NS: not significant; T: TRAM34.
Fig. 3.
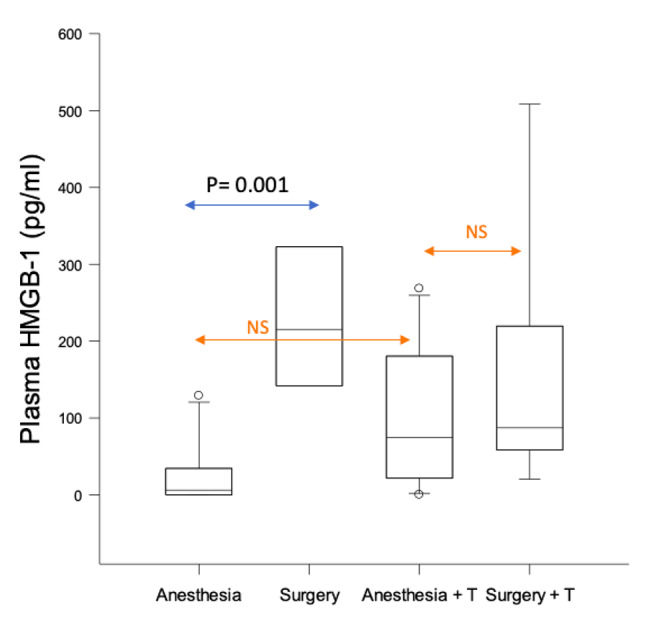
Modulation of surgery-induced rise in peripheral HMGB1 by TRAM34. Comparison of post-surgical changes (surgery vs. anesthesia) in peripheral HMGB1 levels between miglyol (reference conditions) and miglyol + TRAM34 (T) (challenge conditions) in WT mice. Experimental groups included 10 animals. NS: non-significant
Taken together, these results confirm the surgery-induced deleterious cognitive and neuroinflammatory effects observed in previous experimental settings [13, 17].
Based on these results, we then analyzed whether pharmacological inhibition of KCa3.1 channel by TRAM34 would influence the surgery-induced effects. As shown in Fig. 2 and S2, neither cognitive decline nor microglial proliferation was induced by surgery in the presence of TRAM34. Surgery was associated with an increase in peripheral IL-6, regardless of TRAM34 administration (P = 0.049). Surgery-induced peripheral HMGB1 increase was attenuated by TRAM34 (Fig. 3).
Parenthetically, comparison of anesthesia groups (anesthesia + vehicle vs. anesthesia + TRAM34) revealed a decrease in spatial memory cognitive score (P = 0.003) and an increase in microglial proliferation (P = 0.013) (Fig. 2).
In KCa3.1−/− KO mice, surgery neither induced cognitive decline nor microglial proliferation (Fig. 4) although plasma IL-6 was increased (p < 0.001).
Fig. 4.

Surgery-dependent changes in cognitive decline, peripheral inflammation and central microglial presence in KCa3.1−/− KO mice. Experimental groups included 10 animals. NS: non-significant
Discussion
Summary of findings
The ‘surgical phenotype,’ comprising of memory impairment, microgliosis, (proliferation and amoeboid appearance), peripheral inflammation (IL-6) and upregulation of the trauma marker (HMGB1) was present in WT mice after aseptic trauma in the presence of miglyol only (Figs. 2 and 3). Blocking KCa3.1 channel pharmacologically (through TRAM34 administration) (Figs. 2 and 3) or genetically (through KCa3.1 gene inactivation in the KCa3.1−/− mice) (Fig. 4) mitigated the surgical phenotype (apart from circulating IL-6; see below), highlighting a previously unreported potential role of blockade of the KCa3.1 channel for prevention of PND [18].
KCa3.1 blockade and peripheral inflammation
Postoperative IL-6 increase persisted after TRAM34 administration in WT mice (p = 0.049) as well as in KCa3.1−/− mice; similarly, blockade of Kv1.3 channels by PAP-1 did not affect postoperative elevation of circulating IL-6 levels [16]. IL-6 is a potent propagator of the innate immune response to trauma [11] which can both transform into neuroinflammation and facilitate wound healing depending on the involved signaling mechanism [23–25]. Interestingly, the umitigated rise in postoperative IL-6 in the presence of PAP-1 was associated with no deterioration in wound healing suggesting that this peripheral cytokine is regenerative, likely through classical signaling [26]. The influence of TRAM34 adminstration on wound healing remains to be investigated.
KCa3.1 blocking and HMGB1 release
Previous studies supported a putative causal role of surgical-trauma induced HMGB1 in PNDs [8, 27]; TRAM34 administration decreased postoperative elevation in peripheral HMGB1 levels.
As neutralizing HMGB1 prevented the development of PND [8], peripheral HMGB1 levels by themselves might play a more important role in PND development than previously considered.
KCa3.1 blocking and cognition
Previous research indicated the role of TRAM34 in protecting memory performance in murine Alzheimer’s disease [28–30]. TRAM34 also reduced infarction and improved neurological scoring in murine stroke models [21].
Similarly, our data illustrate that KCa3.1 blocking mitigated immediate PND development.
Limitations
Inflammatory markers
While only two markers of inflammation and trauma (IL-6; HMGB1) were consistently measured in these experiments, other markers of inflammation also play a key role in PND development and should be studied [11, 13]. Previous research has, however, shown the importance of these markers in PND development [8, 12].
TRAM34 administration in anesthesia control groups
The KCa3.1 inhibitor TRAM34 had a negative effect on cognition and microgliosis in the anesthesia alone WT mice. (Fig. 3); this interesting finding requires further exploration of the possible interaction between TRAM34 and volatile anesthetic agents.
Blank control group
In this study, ‘anesthesia + vehicle’ was considered to be the blank/ baseline reference for the intervention e.g. Surgery/ TRAM34 injection. An ‘entirely’ blank/ baseline group was not included in the experiments.
Which Kca3.1 channel blocking property is paramount?
As previously mentioned, KCa3.1 channels are not only present on microglia, but also on cell types such as astrocytes, macrophages, T cells and erythrocytes [16].
In this study, experiments show that TRAM34 administration decreased post-operative microgliosis. The effect of TRAM34 administration on astrogliosis was, however, not studied.
TRAM34 administration also had a peripheral effect: decreasing post-operative plasma HMGB1 levels. As monocytes play an important role in the inflammatory cascade leading to PND development [8], TRAM34 administration may also block KCa3.1 channels present in these cells.
Kv1.3 and KCa3.1 channels
As this report and previously published data [21] have established the roles of KCa3.1 and Kv1.3 in PND development, simultaneous administration of TRAM34 and PAP-1 should be investigated in order to determine the specific role of each channel and whether an interaction, including synergism, might take place between these channel blockers.
Conclusion
Previously published data established the role of microglial Kv1.3 in perioperative neurocognitive disorders. The data presented here indicate that perioperative KCa3.1 blockade decreases immediate perioperative cognitive changes, microglial proliferation as well as the peripheral trauma marker HMGB1. It did not influence surgery-induced IL-6 production.
Future research should investigate the specific role of each channel and whether a synergistic interaction occurs between the channel blockers.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgements
The authors acknowledge the University of California, Davis Mouse Biology Program for re-deriving the KCa3.1 –/– mice.
Abbreviations
- PND
Perioperative neurocognitive disorders
- HMGB1
High mobility group box 1 protein
- BM-DMs
Bone marrow-derived monocytes
- CNS
Central nervous system
Author contributions
Study conception/design: SS; JB; HW; MM; VK; JV. Provision of reagents: HW. Data acquisition/analysis: VN; CR; KJ; AA; LS; MCM; MM; VK; JV. Drafting figures: SS; VN; VK; JV. Drafting of manuscript: SS; VK; JV; MM. All authors read and approved the final manuscript.
Funding
SS received a grant from ‘Le Fonds de Recherche Médicale dans le Hainaut’ (FMRH) for this study.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declarations
Ethics approval
Experimental procedures involving animals were approved by the Animal Care Committee of the Université Libre de Bruxelles. (CEBEA-IBMM 2019-24-105). All experiments were conducted in accordance with regulations by the Animal Care Committee of the Université Libre de Bruxelles and in accordance with ARRIVE guidelines [31].
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
senior co-authorship.
References
- 1.Savage GH. Insanity following the use of anaesthetics in operations. BMJ1887; 3:1199–1200.
- 2.Evered L, Silbert B, Knopman DS, Scott DA, DeKosky ST, Oh ES, et al. Recommendations for the nomenclature of Cognitive Change Associated with Anaesthesia and Surgery-2018. Anesthesiology. 2018;129:872–9. doi: 10.1097/ALN.0000000000002334. [DOI] [PubMed] [Google Scholar]
- 3.Steinmetz J, Christensen KB, Lund T, Lohse N, Rasmussen LS, ISPOCD group Long-term consequences of postoperative cognitive dysfunction. Anesthesiology. 2009;110:548–55. doi: 10.1097/ALN.0b013e318195b569. [DOI] [PubMed] [Google Scholar]
- 4.Saxena S, Rodts C, Nuyens V, Lazaron J, Sosnowski V, Verdonk F et al. Preoperative sedentary behavior is neither a risk factor for perioperative neurocognitive disorders nor associated with an increase in peripheral inflammation, a prospective observational cohort study. BMC Anesthesiol. 2020; 284. (PMID: 33187477) [DOI] [PMC free article] [PubMed]
- 5.Patel V, Champaneria R, Dretzke J, Yeung J. Effect of regional versus general anaesthesia on postoperative delirium in elderly patients undergoing surgery for hip fracture: a systematic review. BMJ Open. 2018;8:e020757. doi: 10.1136/bmjopen-2017-020757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Moller JT, Cluitmans P, Rasmussen LS, Houx P, Rasmussen H, Canet J, et al. Long-term postoperative cognitive dysfunction in the elderly ISPOCD1 study. ISPOCD investigators. International Study of Post-Operative Cognitive Dysfunction. Lancet. 1998;351:857–61. doi: 10.1016/S0140-6736(97)07382-0. [DOI] [PubMed] [Google Scholar]
- 7.Sieber FE, Neufeld KJ, GottscalkA,Bigelow GE, Oh ES, Rosenberg PB, et al. Effect of depth of Sedation in older patients undergoing hip fracture repair on postoperative delirium: the STRIDE Randomized Clinical Trial. JAMA Surg. 2018;153:987–95. doi: 10.1001/jamasurg.2018.2602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vacas S, Degos V, Tracey KJ, Maze M. High-mobility group box 1 protein initiates postoperative cognitive decline by engaging bone marrow-derived macrophages. Anesthesiology. 2014;120:1160–7. doi: 10.1097/ALN.0000000000000045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Terrando N, Eriksson LI, Ryu JK, Yang T, Monaco C, Feldmann M, et al. Resolving postoperative neuroinflammation and cognitive decline. Ann Neurol. 2011;70:986–95. doi: 10.1002/ana.22664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Feng X, Valdearcos M, Uchida Y, Lutrin D, Maze M, Koliwad SK. Microglia mediate postoperative hippocampal inflammation and cognitive decline in mice. JCI Insight. 2017;2:e91229. doi: 10.1172/jci.insight.91229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cibelli M, Fidalgo AR, Terrando N, Ma D, Monaco C, Feldmann M, et al. Role of interleukin-1beta in postoperative cognitive dysfunction. Ann Neur. 2010;68:360–8. doi: 10.1002/ana.22082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hu J, Feng X, Valdearcos M, Lutrin D, Uchida Y, Koliwad SK, et al. Interleukin-6 is both necessary and sufficient to produce perioperative neurocognitive disorder in mice. Br J Anaesth. 2018;120:537–45. doi: 10.1016/j.bja.2017.11.096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Feng X, Degos V, Koch LG, Britton SL, Zhu Y, Vacas S, et al. Surgery results in exaggerated and persistent cognitive decline in a rat model of the metabolic syndrome. Anesthesiology. 2013;118:1098–105. doi: 10.1097/ALN.0b013e318286d0c9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Terrando N, Gomez-Galan M, Yang T, Carlstrom M, Gustavsson D, Harding RE, et al. Aspirin-triggered resolvin D1 prevents Surgery-Induced Cognitive decline. FASEBJ. 2013;27:3564–71. doi: 10.1096/fj.13-230276. [DOI] [PubMed] [Google Scholar]
- 15.Yang T, Xu G, Newton PT, Chagin AS, Mkrtchian S, Carlstrom M, et al. Maresin 1 attenuates Neuroinflammation in a mouse model of Perioperative Neurocognitive Disorders. BJA. 2019;122:350–60. doi: 10.1016/j.bja.2018.10.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Saxena S, Lai IK, Li R, Maze M. Neuroinflammation is a putative target for the prevention and treatment of perioperative neurocognitive disorders. Br Med Bull. 2019;130:125–35. doi: 10.1093/bmb/ldz010. [DOI] [PubMed] [Google Scholar]
- 17.Lai IK, Valdearcos M, Morioka K, Saxena S, Feng X, Li R, et al. Blocking the Kv1.3 potassium channel prevents postoperative Neuroinflammation and Cognitive decline without impairing Wound Healing. Br J Anaesth. 2020;125:298–307. doi: 10.1016/j.bja.2020.05.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nguyen HM, Grossinger EM, Horiuchi M, Davis KW, Jin L, Maezawa I, et al. Differential Kv1.3, KCa3.1, and Kir2.1 expression in “Classically” and “Alternatively. Activated Microglia Glia. 2017;65:106–21. doi: 10.1002/glia.23078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wullf H, Miller MJ, Hansel W, Grissmer S, Cahalan MD, Chandy KG. Design of a potent and selective inhibitor of the intermediate-conductance Ca2+-activated K+ channel, IKCa1: a potential immunosuppressant. Proc Natl Acad Sci USA. 2000;97:8151–6. doi: 10.1073/pnas.97.14.8151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kaushal V, Koeberle PD, Wang Y, Schlichter LC. The Ca2+-activated K+ channel KCNN4/KCa3.1 contributes to microglia activation and nitric oxide-dependent neurodegeneration. J Neurosci. 2007;27:234–44. doi: 10.1523/JNEUROSCI.3593-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chen YJ, Raman G, Bodendiek S, O’Donnell ME, Wulff H. The KCa3.1 blocker TRAM34 reduces infarction and neurological deficit in a rat model of ischemia/reperfusion stroke. J Cereb Blood Flow Metab. 2011;31:2363–74. doi: 10.1038/jcbfm.2011.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Miedel JC, Patton JM, Miedel AN, Miedel ES, Levenson JM. Assessment of spontaneous alternation, Novel object recognition and limb clasping in transgenic mouse models of Amyloid-β and tau neuropathology. J Vis Exp. 2017;123:55523. doi: 10.3791/55523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hu J, Zhang Y, Huang C, Feng X, He S, Zhang Y, Maze M. Interleukin-6 trans-signalling in hippocampal CA1 neurones mediates perioperative neurocognitive disorders in mice. BJA. 2022;129:923–36. doi: 10.1016/j.bja.2022.08.019. [DOI] [PubMed] [Google Scholar]
- 24.De Benedetti F, Rucci N, Del Fattore A, Peruzzi B, Paro R, Longo M, Vivarelli M, Muratori F, Berni S, Ballanti P, Ferrari S, Teti A. Impaired skeletal development in interleukin-6-transgenic mice: a model for the impact of chronic inflammation on the growing skeletal system. Arthritis Rheum. 2006 Nov;54(11):3551–63. 10.1002/art.22175. (PMID: 17075861). [DOI] [PubMed]
- 25.Nakahara H, Song J, Sugimoto M, Hagihara K, Kishimoto T, Yoshizaki K, Nishimoto N. Anti-interleukin-6 receptor antibody therapy reduces vascular endothelial growth factor production in rheumatoid arthritis. Arthritis Rheum. 2003 Jun;48(6):1521–9. 10.1002/art.11143. (PMID: 12794819). [DOI] [PubMed]
- 26.Uciechowski P, Dempke WCM. Interleukin-6: a masterplayer in the cytokine network. Oncology. 2020;98:131–7. doi: 10.1159/000505099. [DOI] [PubMed] [Google Scholar]
- 27.Saxena S, Kruys V, De Jongh R, Vamecq J, Maze M. High-mobility Group Box-1 and its potential role in Perioperative Neurocognitive Disorders. Cells. 2021;10:2582. doi: 10.3390/cells10102582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Maezawa I, Zimin PI, Wulff H, Jin LW. Amyloid-beta protein oligomer at low nanomolar concentrations activates microglia and induces microglial neurotoxicity. J Biol Chem. 2011;286:3693–706. doi: 10.1074/jbc.M110.135244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Yi M, Yu P, Lu Q, Geller HM, Yu Z, Chen H. KCa3.1 constitutes a pharmacological target for astrogliosis associated with Alzheimer’s disease. Mol Cell Neurosci. 2016;76:21–32. doi: 10.1016/j.mcn.2016.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yu Z, Dou F, Wang Y, Hou L, Chen H. Ca2+-dependent endoplasmic reticulum stress correlation with astrogliosis involves upregulation of KCa3.1 and inhibition of AKT/mTOR signaling. J Neuroinflammation. 2018;15:316. doi: 10.1186/s12974-018-1351-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Percie du Sert N, Hurst V, Ahluwalia A, Alam S, Avey MT, Baker M, et al. The ARRIVE guidelines 2.0: updated guidelines for reporting animal research. PloS Biol. 2020;18:e3000410. doi: 10.1371/journal.pbio.3000410. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.