

Abstract
Inducible epithelial human β-defensins (hBD) play an important role in intestinal barrier function. In vitro studies showed that clinically effective probiotics induce antimicrobial hBD-2. Here, we aimed to assess the in vivo effect in healthy volunteers and also addressed how defensins affect probiotic survival. Symbioflor 2 containing one strain of several viable genotypes of Escherichia coli was administered to 23 healthy individuals. After 3 weeks, fecal hBD-2 peptide was increased in 78% (mean 3.7-fold; P<0.0001). Interestingly, the fecal hBD-2 peptide was still elevated 9 weeks after treatment (P=0.008). In vitro studies revealed that this effect was mediated by only one out of three tested E. coli genotypes and comparable to probiotic E. coli Nissle 1917 (10- to 15-fold). Functional assays showed that all tested bacteria were similarly killed by defensins allowing to speculate about a suicidal character of this effect. Defensin induction seems to be a common and important mechanism of probiotic treatment.
Introduction
Probiotics are commonly defined as live nonpathogenic microorganisms that confer health benefits on the host, when administered in adequate amounts.1 General formulated expectations include that they should resist gastric, bile, and pancreatic secretions, attach to epithelial cells, and colonize the human intestine. Probiotic microorganisms that are currently in use include most commonly lactic acid bacteria of the genera Lactobacillus and Bifidobacterium but also Escherichia coli or yeasts, such as the Saccharomyces boulardii strain.2 Probiotics are either used as dietary supplements or as pharmaceutical products for therapeutic purposes.
Most recent clinical studies substantiate reliable evidence for the therapeutic efficacy of some, yet not all, probiotic organisms. The probiotics Bifidobacterium bifidum, Streptococcus thermophilus, or E. coli Nissle 1917 (Mutaflor) administered to toddlers highly significantly reduced the incidence of acute diarrhea and rotavirus shedding.3, 4 In adults, the efficacy of probiotics was demonstrated by shorter durations of traveler's diarrhea and other self-limited gastrointestinal infections.5 In recent years, several studies substantiate that the intestinal flora plays a key role in the development of inflammatory bowel disease.6 This chronic inflammation of the intestine is often grouped into two major entities, Crohn's disease and ulcerative colitis, based on clinical features and histopathology. Although inflammation in ulcerative colitis is typically restricted to the colon, that of Crohn's disease occurs at many sites, most commonly in the small intestine and in the colon. In addition to genetic factors in inflammatory bowel disease, numerous studies have implicated a key role of the intestinal microbiota in the pathogenesis of this disease.7, 8 Especially for ulcerative colitis patients, probiotics offer a safe alternative to current therapy. Three independent studies have found an equal efficacy on the remission maintenance of ulcerative colitis after administration of the probiotic E. coli Nissle 1917 compared to the standard treatment with 5-aminosalicylate.9, 10, 11 The advantage of probiotic treatment was a prolongation of remission without any adverse effects in contrast to conventional treatment. The probiotic mixture VSL no. 3, composed of eight different bacterial species, also diminished symptoms in patients suffering from inflammatory bowel diseases.12
We have recently hypothesized that the beneficial effect of E. coli Nissle 1917 and other probiotic bacteria in the intestinal tract might be at least partly because of the induction of protective host antimicrobials. An upregulation of a diminished antimicrobial defense as seen in Crohn's disease13 could be one important mechanism by which some probiotics exert their beneficial effect in inflammatory bowel disease patients and thus inhibit the invasion of the mucosal surface by commensals and other microorganisms. In vitro experiments showed that incubation of intestinal epithelial cells with these bacteria provoked a strong induction of inducible human β-defensin-2 (hBD-2) expression.14 Defensins are components of the armory of endogenous antimicrobials, which are part of the innate immune system.15 They are small, cationic (positively charged) peptides of which most are classified into α- or β-defensins. The human β-defensins provide a first line of defense against potentially pathogenic microbes at the body's mucosal frontiers as they are produced by epithelial cells of the lung, skin, and intestinal tract.16 Defensins, produced by the mucosal epithelium, have a broad antibiotic spectrum against Gram-negative (E. coli, Salmonella) and -positive (Staphylococcus aureus) bacteria as well as fungi (Candida albicans), and viruses (human immunodeficiency virus). Despite the known induction of antimicrobial peptides in cell culture, nothing is known about the in vivo influence and especially if the real treatment under normal doses is sufficient to induce antimicrobial peptide secretion in the intestinal lumen. Thus, we investigated whether the administration of a known probiotic E. coli viable bacterial culture (Symbioflor 2) might impact on the hBD-2 peptide secretion in the feces. To exclude other possible influences caused by disease, we used healthy individuals. In a second step, after we observed in vivo secretion upon treatment with Symbioflor, we tested which viable E. coli genotype induced defensin expression in cell culture. As it is known that different clinically used probiotic E. coli strains do not survive in the normal flora and have to be administered daily, we also tested if these bacteria are resistant to antimicrobial defensin killing.
Results
Symbioflor 2 induces hBD-2 fecal peptide secretion in man
The administration of Symbioflor 2 was well tolerated by all study participants and no adverse events were observed. Five individuals with placebo treatment did not show any significant changes in hBD-2 peptide levels (Figure 1a ). Taken together, the overall increase of hBD-2 protein fecal level after 3 weeks of probiotic treatment was 3.7-fold as compared to before treatment (P<0.0001; Figure 1b). To assess whether the probiotic uptake results in increased hBD-2 secretion, we also determined the percentage of study participants who expressed a higher hBD-2 secretion into the feces after the intake of Symbioflor 2. After 3 weeks, 78% showed an increase of more than 20%—and 74% more than 30%—of hBD-2 secretion as compared to before treatment. At week 12, 9 weeks after the end of the probiotic administration, the hBD-2 fecal levels were determined in a smaller subset and compared with the baseline values of the same individuals (Figure 1c). Even though the levels were lower than at week 3, fecal hBD-2 protein was still significantly higher than at the beginning of the study (P=0.008; Figure 1c).
Figure 1.
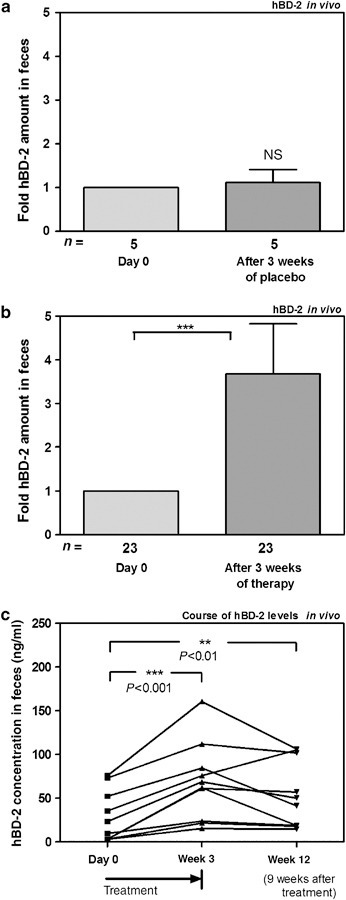
Probiotics stimulate hBD-2 protein secretion in vivo. (a) hBD-2 peptide secretion after 3 weeks of placebo treatment in five individuals. (b) In total, 23 individuals received preparations of Symbioflor 2 for 3 weeks. HBD-2 peptide was determined by ELISA in stool samples. Data were normalized to hBD-2 levels before treatment that were set as 1 and represent the mean±s.e.m. (c) Probiotics exert a post-treatment effect on hBD-2 secretion. HBD-2 peptide levels are shown in a subset at day 0, after 3 weeks of probiotic treatment and 9 weeks after stop of treatment. Paired t-tests were used for the comparison of values before and after treatment. The data represent the means±s.e.m. **P<0.01; ***P<0.001.
hBD-2 induction by probiotics in colonic epithelial cells
To confirm the in vivo studies and to identify the inducing bacterium, we performed cell culture experiments. We tested the hBD-2 stimulating effect of different probiotics including different genotypes of the Symbioflor E. coli DSM 17252 (G1, G2, and G3) as well as a positive (E. coli Nissle 1917) and negative control (E. coli K12). In previous studies, we have shown that different probiotic strains including E. coli Nissle 1917, Lactobacillus fermentum, Pediococcus Pentosaceus, and Lactobacillus acidophilus PZ 1129 consistently induce hBD-2 mRNA and protein secretion in vitro.14, 17 In contrast, more than 35 E. coli clinical isolates as well as E. coli K12 showed no effect.14 Herein, only one of the three Symbioflor 2 genotype strains (G2) showed a strong induction of hBD-2 in Caco-2 cells after 6 h of incubation that was comparable to E. coli Nissle 1917 (Figure 2a ). When we tested the hBD-2 induction of G2 at different time points, we observed the highest value after 12 h of incubation (Figure 2b). However, the induction of hBD-2 reached a significant extent at all tested time points demonstrating the consistency of the induction capacity. The housekeeping gene expression (glyceraldehyde-3-phosphate dehydrogenase) remained stable at all time points. The induction of hBD-2 was also shown to be dose-dependent with a maximum at an optical density of 0.3 (Figure 2c). In contrast, the other E. coli G1 and G3 did not induce hBD-2 in Caco-2 cells at any time (Figure 2b) or any tested bacterial concentration (data not shown).
Figure 2.
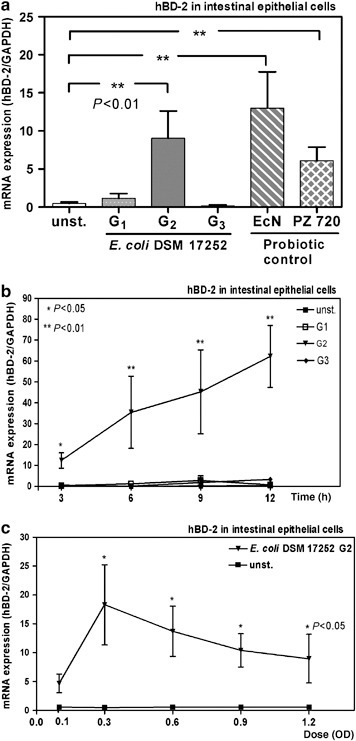
Symbioflor 2 induces hBD-2 in intestinal epithelial cells in vitro. (a) Incubation of Caco-2 cells with three genotypes of Symbioflor 2 E. coli strain (G1, G2, G3). Caco-2 cells were incubated for 6 h with the heat-killed test bacteria and two probiotic strains as controls (optical density of 0.3). RNA was isolated, reverse-transcribed into cDNA, and the amount of hBD-2 copies was determined by real-time PCR. (b) Time course of hBD-2 induction by Symbioflor 2. Caco-2 cells were treated with the three genotypes (G1, G2, G3) of Symbioflor 2 E. coli strain for 3, 6, 9, and 12 h. (c) Dose dependence, Caco-2 cells were treated for 6 h with elevating doses of the genotype G2 of the Symbioflor 2 E. coli strain. The data represent the mean±s.e.m. of three independent experiments performed in duplicates. Unpaired t-tests were used to evaluate the statistical significance for comparisons between the groups. The data represent the means±s.e.m. *P<0.05; **P<0.01.
Probiotic E. coli bacteria strains are not resistant to antimicrobial peptides
Next it was studied whether the same strains are sensitive to antimicrobial killing by natural defensins. In plate diffusion assays, hBD-2 showed a strong potency to kill E. coli bacteria that is consistent with previous reports. Interestingly there were no differences between any of the tested strains. The two probiotic E. coli bacteria that strongly induced hBD-2 (E. coli Nissle, DSM 17252 G2) were killed in an almost equal extent as compared to E. coli K12 as well as Symbioflor 2 bacterial genotypes with negative defensin induction (DSM 17252: G1 and G3; Figure 3a ). We also tested if the same bacteria were different toward resistance to other antimicrobial peptides. Similar to hBD-2 killing, all tested bacteria were potently eliminated by hBD-3 with no differences between any of the strains (Figure 3b). In addition, lysozyme, which is another known antimicrobial host molecule, equally showed antibiotic activity with no difference between probiotic and other tested E. coli bacteria (Figure 3c).
Figure 3.
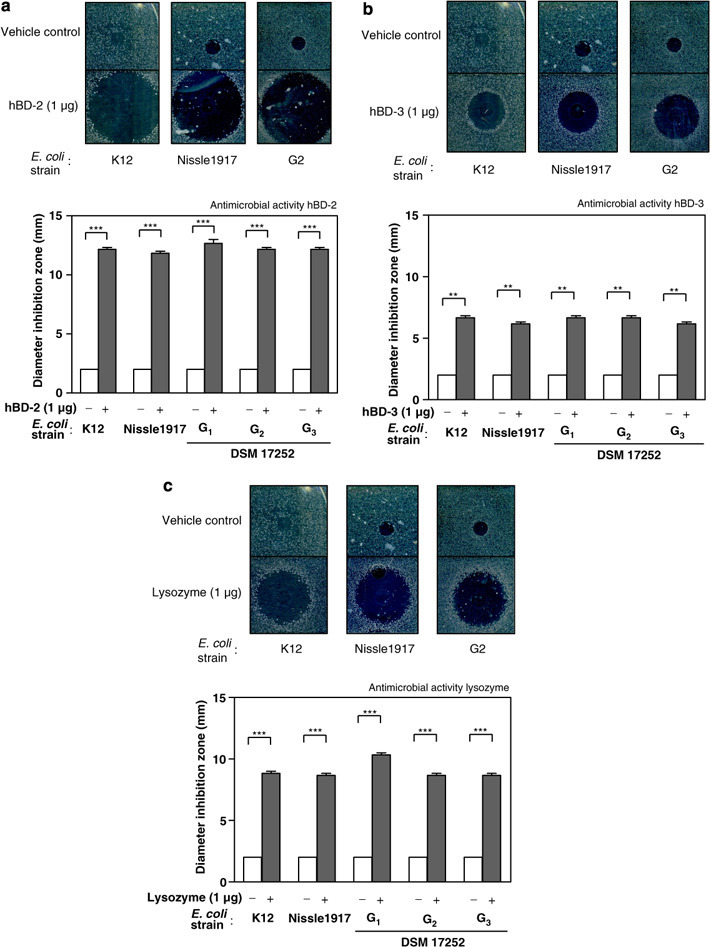
Antimicrobial resistance of probiotic and control E. coli toward antimicrobial peptides. (a) hBD-2, (b) hBD-3, (c) lysozyme. Representative antimicrobial plate assays are shown for each antimicrobial (upper part). The data represent the mean±s.e.m. and were analyzed for statistical significance using one sample t-test. **P<0.01; ***P<0.001.
Discussion
Herein, we show for the first time that oral administration of regular doses of a probiotic E. coli preparation (Symbioflor 2) induces mucosal human β-defensin peptide secretion into the feces of healthy individuals. Consistent with these in vivo studies, we identified one E. coli strain that is part of the commercial mixture as a stimulatory agent in vitro. As the tested probiotic bacteria are not resistant to antimicrobial host factors such as hBD-2, these functional studies presented here allow to speculate about a possible suicidal character at the place of defensin secretion.
It was recently demonstrated that E. coli Nissle 1917, as well as L. acidophilus, L. fermentum, and P. pentosaceus as well as VSL no. 3 and other probiotic strains highly specifically activate colonic cells to synthesize defensins in vitro. 14, 17 In contrast 40 other clinical E. coli isolates lacked this effect.14 This specific effect is consistent with the observation that inducible β-defensins are normally—despite the presence of numerous E. coli species in the gut microbiota—absent in the healthy intestine. However, the major limitation of all these studies is the exclusive focus on in vitro experiments as performed in intestinal (tumor) epithelial cells. Unfortunately there are many examples of experimental effects that cannot be confirmed in vivo. For this reason we studied if a regular dose of a commercial probiotic preparation (Symbioflor 2) can induce active antimicrobial peptide secretion in healthy individuals and how this in vivo effect translates back to experiments using cell culture. Interestingly, hBD-2 was still increased even 9 weeks after the administration of Symbioflor 2 was stopped, suggesting additional stimulatory mechanisms that cannot be clearly explained by the presented data. Compared to the controls E. coli Nissle 1917 and PZ720, only one of the Symbioflor 2 genotypes had a similarly strong capacity to induce hBD-2 in cell culture. This induction of hBD-2 was further enhanced by an increased contact time of the bacteria with the colonic epithelial cells that differed from the induction pattern of E. coli Nissle 1917.14 On the basis of recent observations with E. coli Nissle 1917, we also expected an increase of the effect by increasing the bacterial concentration. However, all doses except the smallest reached the same level in inducing hBD-2 expression with no dose dependence. Together these time and dose data suggest that the probiotic stimulatory strength is similar to other probiotic strains (e.g., Nissle) but the mechanisms of induction may be different. In case of E. coli Nissle 1917, the hBD-2 induction mechanism is mediated by a specific flagellin.18 In these studies genetically manipulated E. coli Nissle 1917 deficient in flagellin protein failed to induce defensin expression. Moreover, isolated flagellin from E. coli Nissle 1917 strain was effective in contrast to similar doses of flagellin isolated by another apathogenic E. coli strain. Thus, it is likely that the flagellin structure of the E. coli strain Nissle 1917 exhibits some important modifications compared to other E. coli strains that also express flagella but lack the ability of hBD-2 induction. Interestingly, the E. coli Symbioflor 2 strains including G2 do not express Flagella protein18 despite similar hBD-2 induction. Similarly other defensin stimulating probiotic strains such as the bacterial mix VSL no. 3 or other Lactobacilli also lack flagella. Thus, even if the result of hBD-2 induction can be found by all tested probiotic strains, the relevant factors and mechanisms mediating this immune stimulatory effect appear to differ from strain to strain. These factors that are likely to be different between different E. coli, Lactobacilli, and other probiotic species still remain to be investigated.
Another important question concerns the biological relevant concentrations of hBD-2. In this study, the hBD-2 protein concentration was measured in overall stool samples. Of note, the main biological locus of action of antimicrobial defensins is probably the mucus that is attached to the mucosal layer.19 The defensin concentration as measured here likely reflects the luminal rather then the mucus concentration. Thus, the amounts in the mucus would be much higher as the obtained values of hBD-2 are a dilution of the original amount as secreted by mucosal epithelial cells.
To study the possible functional consequence of defensin induction, we tested if Symbioflor 2 bacterial cultures are able to resist antimicrobial killing. We found that there were no differences between the genotypes and probiotic and normal E. coli were similarly killed by hBD-2 in vitro. The same observation was found for other antimicrobial peptides such as hBD-3 and lysozyme. Thus, even though the probiotic strains are very specific in provoking an antimicrobial host response, they do not seem to be provided with protective factors to resist an antimicrobial attack by the host. Possibly, this could at least partly explain why both, E. coli Nissle as well as Symbioflor 2 have to be continuously administered. Moreover, it is quite possible that E. coli strains that do not induce hBD-2 also will not stably colonize the gut. Thus, there may or may not be a cause and effect with hBD-2 induction. Another functional question is the effect of probiotic hBD-2 induction, especially that of G2 on the luminal intestinal microbial flora. Unfortunately there are no direct data available. Our own recent studies have shown that very small changes in antimicrobial defensins (human defensin 5) significantly alter the composition of the downstream luminal microbiota.20 To mechanistically address the question we used an HD-5 transgenic mouse model in which we compared heterozygotic (+/−) with homozygotic (+/+) mice (twofold difference in small intestinal HD5 expression).20 It would be attractive to create a human β-defensin 2 transgenic mouse model and also analyze the composition of microbiota in these mice. In addition, future studies should systematically analyze the composition of the microbiota after treatment with G2 and the other probiotic components in rodent models as well as in humans. The elucidation of probiotic actions is still on an initial stage, despite increasing numbers of studies confirming their beneficial effects on animals and humans. More insights into the probiotic-induced immune regulation are relevant to fortify the application of this promising treatment strategy.
Taken together, this study suggests that the stimulation of protective host factors such as defensins is an important and broad mechanism of probiotic action in vivo even though the precise mechanisms seem to vary between individual probiotic strains.
Methods
Human study and sample collection In total, 31 healthy individuals who gave informed consent were included in the study. Of those, 5 individuals were treated with placebo and 26 individuals received Symbioflor 2 (containing viable commensal E. coli bacteria). From the initial verum group of 26, 3 individuals were excluded from the study, 1 because of irregular drug intake, and 2 because of study discontinuation because of compliance. Thus, the final verum group contained 23 individuals for a total of 21 days of therapy. Symbioflor 2 was administered in standard dosage starting with two times five drops daily and a daily increase by two times one drop until 15 drops daily were reached. Fourteen drops are 1 ml containing 1.5–4.5×107 viable bacterial counts. The total duration of treatment was 21 days. Stool samples were taken at baseline and after 3 weeks (stop of treatment). From some subjects, a third sample was collected after 12 weeks (9 weeks after stop of treatment). Owing to lack of compliance, the latter sample was not obtained in all cases (n=10 out of the total of 23). The study was approved by the local ethical committee and all volunteers gave written informed consent.
hBD-2 protein concentration (hBD-2 ELISA) The protein assessment of hBD-2 secretion was performed as described.21 After collection, stool samples were frozen at −20 °C. Before analysis, 100 mg of stool were prepared using the commercially available stool preparation kit (Roche Diagnostics GmbH, Mannheim, Germany; cat no. 745804). hBD-2 protein concentration was assessed in stool samples at different time points in all participants of the study. The protocol was used according to the supplier's recommendation (ALPCO, Windham, Germany). In brief, the β-defensin-2 in standard and samples is bound to an available excess of polyclonal antibodies against β-defensin-2, which are immobilized on the surface of the microtiter plate. After a washing step, to remove all interfering substances, the quantification of bound β-defensin 2 is carried out by adding a biotinylated polyclonal anti-β-defensin-2 antibody. This antibody is detected with horseraddish peroxidase-labeled streptavidin. The amount of converted substrate by peroxidase is directly proportional to the amount of bound β-defensin-2 and can be determined photometrically at 450 nm.
Bacterial strains and growth conditions Bacterial strains used in this study are shown in Table 1 . The probiotic strains were grown overnight at 37 °C under gentle agitation at 200 r.p.m. in trypticase soy broth. To obtain bacteria in a linear growth phase, 100 μl of the bacterial suspension were added to 10 ml fresh trypticase soy broth medium and grown under permanent shaking for 5 h. Heat inactivation of the bacteria was carried out in a water bath at 65 °C for 1 h. Bacteria were concentrated by centrifugation, washed with phosphate-buffered saline and adjusted to a density of 3×108 cells/ml with fetal calf serum- and antibiotic-free cell culture medium. For testing of dose dependency, bacteria were adjusted to the optical densities 0.1, 0.3, 0.6, 0.9, and 1.2.
Table 1.
Bacterial strains used in this study
| Strain | Strain designation/serotype | Type of isolate | Source |
|---|---|---|---|
| E. coli Nissle 1917 (EcN) | O6:K5:H1 | Pharmaceutical | 1 |
| E. coli PZ 720 | O83:K24:H31 | Probiotic strain | 1 |
| E. coli DSM 17252 | 2 | ||
| S2 G1: E. coli Genotype 1/2 | Osp.:H- | 2 | |
| S2 G2: E. coli Genotype 3/10 | O13.:H- | 2 | |
| S2 G3: E. coli Genotype 4/9 | Osp.:H- | 2 | |
| Enterococcus fecalis | |||
| Klebsiella spec. | |||
Osp, spontaneously agglutinating i.e., the classical rough E. coli type.
1 Ardeypharm collection of strains, Pharma-Zentrale GmbH, Herdecke, Germany.
2 SymbioPharm GmbH, Germany.
Cell culture Caco-2 cells (German Collection of Microorganisms and Cell cultures (DSMZ) ACC 169) were cultivated in Dulbecco's modified Eagle's medium containing 25 mM HEPES, 2 mM glutamine, 10% fetal calf serum, 50 μg/ml gentamicin, and 1% nonessential amino acids as described.14 Cells were grown as monolayers in 75-cm2 flasks (Greiner, Frickenhausen, Germany) at 37 °C in a 5% CO2—95% air atmosphere with 90% humidity. For stimulation experiments, cells were seeded at a density of 3.2×105 cells/well in 12-well culture plates (Becton Dickinson GmbH, Heidelberg, Germany). Cells grown to ∼70% confluence in culture wells were incubated overnight with serum- and antibiotic-free medium to eliminate serum-induced hBD-2 expression and prevent any influence of antibiotics on the immune response. To determine hBD-2 mRNA expression, Caco-2 cells were incubated with the bacteria for different time points including 3, 6, and 12 h.
RNA isolation and cDNA synthesis After stimulation, cells were washed with phosphate-buffered saline and harvested with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the supplier's protocol.14 Subsequently 1 μg of total RNA was reverse transcribed into cDNA with oligo (dT) primers and 15 U/μg AMV Reverse Transcriptase (Promega, Madison, USA) according to standard procedures.
Real-time RT-PCR Real-time reverse transcription PCR (RT-PCR) analyses were performed in a fluorescence temperature cycler (LightCycler; Roche Diagnostics GmbH) according to manufacturer's instructions.14 As a template served cDNA corresponding to 10 ng of RNA in a 10 μl reaction mixture containing 3 mM MgCl2, 0.5 μM of each primer and 1×LightCycler-FastStart DNA Master SYBR Green I mix (Roche Diagnostics GmbH). Initial denaturation at 95 °C for 10 min was followed by 45 cycles, each cycle consisting of 95 °C for 15 s, the primer-specific annealing temperature for 5 s and elongation at 72 °C for 15 s. For hBD-2 (sense 5′-ATCAGCCATGAGGGTCTTGT-3′; antisense 5′-GAGACCACAGGTGCCAATTT-3′), the annealing temperature was set to 60 °C. Amplification using these primers resulted in a 172-bp fragment. As an internal control gene, we used the common housekeeping gene glyceraldehyde-3-phosphate dehydrogenase. For glyceraldehyde-3-phosphate dehydrogenase (sense 5′-CCAGCCGAGCCACATCGCTC-3′; antisense 5′-ATGAGCCCCAGCCTTCTCCAT-3′), we used a touchdown protocol with a primary temperature of 66 °C and a target temperature of 60 °C. At the end of each run, melting curve profiles were achieved by cooling the sample to 65 °C for 15 s and then heating slowly at 0.20 °C/s up to 95 °C with continuous measurement of fluorescence to confirm amplification of specific transcripts. Cycle-to-cycle fluorescence emission readings were monitored and analyzed using LightCycler software (Roche Diagnostics GmbH). Melting curves were generated after each run to confirm amplification of specific transcripts. The specificity of the amplification products was verified by subjecting the amplification products to electrophoresis on a 2% agarose gel and visualization by ethidium bromide staining. Relative mRNA expression is given as a ratio between target gene and glyceraldehyde-3-phosphate dehydrogenase gene expression.
Antimicrobial assay To test if different E. coli strains including probiotic bacteria are susceptible to antibacterial host peptides, we used human β-defensin-2 and -3 as well as lysozyme. Antimicrobial activity was determined by using a radial diffusion assay as described by Lehrer et al. 22 with minor modifications. Briefly, 1×107 bacterial CFU were inoculated into 10 ml of warm sterile underlay agar (0.03% w/v trypticase soy broth (trypticase soy broth, Becton Dickinson GmbH), 10 mM sodium phosphate, pH 7.4, 1% low-electroendosmosis-type agarose (Sigma-Aldrich, Steinheim, Germany) and 0.02% v/v Tween 20 (Merck, Darmstadt, Germany)). After strong vortexing, the liquid agar was poured into square Petri dishes (Becton Dickinson GmbH) and small wells were punched with a biopsy puncher. Test substances lysozyme (1 μg), hBD-2 (1 μg) and hBD-3 (1 μg) were diluted in 5 μl of 0.01% acetic acid, which also served as vehicle control, and added into the wells. After incubation for 3 h at 37 °C, the gel was overlayed with 10 ml of full-nutrition overlay agar (6% w/v trypticase soy broth, 1% low-EEO agarose) and incubated for 16 h at 37 °C. The gel was stained as described by Lehrer et al. and diameter of inhibition zone was measured; indicated values were obtained from triplicates.
Statistics Data were analyzed using GraphPad Prism (Version 3.1 for Windows, GraphPad Software, San Diego, CA, USA). For the description of random samples, the mean±standard error of the mean (s.e.m.) was used. Paired t-tests were used to evaluate differences in hBD-2 levels before and after treatment in stool samples (Figure 1). Data from hBD-2 induction experiments in colonic epithelial cells were analyzed using unpaired t-tests (Figure 2). For statistical analysis of the antimicrobial plate assays the one sample t-test was used. P<0.05 was considered as statistically significant.
Acknowledgments
We thank Jutta Bader and Kathleen Siegel for excellent technical support. This work was supported by the Robert Bosch Foundation. Jan Wehkamp is an Emmy Noether scholar of Deutsche Forschungsgemeinschaft (DFG) that also supported this work (WE 436/1-1). The study was also financially supported by Symbio Herborn Group GmbH, Herborn, Germany.
Footnotes
Supplementary information The online version of this article (doi:10.1038/mi.2008.77) contains supplementary material, which is available to authorized users.
Acknowledgments
PowerPoint slide for Fig. 1
PowerPoint slide for Fig. 2
PowerPoint slide for Fig. 3
PowerPoint slide for Table 1
References
- 1.Ouwehand AC, Salminen S, Isolauri E. Probiotics: an overview of beneficial effects. Antonie Van Leeuwenhoek. 2002;82,:279–289. doi: 10.1023/A:1020620607611. [DOI] [PubMed] [Google Scholar]
- 2.Saavedra JM. Clinical applications of probiotic agents. Am. J. Clin. Nutr. 2001;73,:1147S–1151S. doi: 10.1093/ajcn/73.6.1147S. [DOI] [PubMed] [Google Scholar]
- 3.Saavedra JM, Bauman NA, Oung I, Perman JA, Yolken RH. Feeding of Bifidobacterium bifidumStreptococcus thermophilus to infants in hospital for prevention of diarrhoea and shedding of rotavirus. Lancet. 1994;344,:1046–1049. doi: 10.1016/S0140-6736(94)91708-6. [DOI] [PubMed] [Google Scholar]
- 4.Henker J. The probiotic Escherichia coli strain Nissle 1917 (EcN) stops acute diarrhoea in infants and toddlers. Eur. J. Pediatr. 2007;166,:311–318. doi: 10.1007/s00431-007-0419-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Marteau PR, de VM, Cellier CJ, Schrezenmeir J. Protection from gastrointestinal diseases with the use of probiotics. Am. J. Clin. Nutr. 2001;73,:430S–436S. doi: 10.1093/ajcn/73.2.430s. [DOI] [PubMed] [Google Scholar]
- 6.Sartor RB. Intestinal microflora in human and experimental inflammatory bowel disease. Curr. Opin. Gastroenterol. 2001;17,:324–330. doi: 10.1097/00001574-200107000-00005. [DOI] [PubMed] [Google Scholar]
- 7.Rutgeerts P. Effect of fecal stream diversion on recurrence of Crohn's disease in the neoterminal ileum. Lancet. 1991;338,:771–774. doi: 10.1016/0140-6736(91)90663-A. [DOI] [PubMed] [Google Scholar]
- 8.Podolsky DK. Inflammatory bowel disease. N. Engl. J. Med. 2002;347,:417–429. doi: 10.1056/NEJMra020831. [DOI] [PubMed] [Google Scholar]
- 9.Kruis W, Schutz E, Fric P. Double-blind comparison of an oral Escherichia coli preparation and mesalazine in maintaining remission of ulcerative colitis. Aliment Pharmacol. Ther. 1997;11,:853–858. doi: 10.1046/j.1365-2036.1997.00225.x. [DOI] [PubMed] [Google Scholar]
- 10.Kruis W. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut. 2004;53,:1617–1623. doi: 10.1136/gut.2003.037747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rembacken BJ, Snelling AM, Hawkey PM, Chalmers DM, Axon AT. Non-pathogenic Escherichia coli vs. mesalazine for the treatment of ulcerative colitis: a randomised trial. Lancet. 1999;354,:635–639. doi: 10.1016/S0140-6736(98)06343-0. [DOI] [PubMed] [Google Scholar]
- 12.Gionchetti P. Probiotics—role in inflammatory bowel disease. Dig. Liver Dis. 2002;34(Suppl 2):S58–S62. doi: 10.1016/S1590-8658(02)80166-9. 12408442. [DOI] [PubMed] [Google Scholar]
- 13.Wehkamp J, Schmid M, Stange EF. Defensins and other antimicrobial peptides in inflammatory bowel disease. Curr. Opin. Gastroenterol. 2007;23,:370–378. doi: 10.1097/MOG.0b013e328136c580. [DOI] [PubMed] [Google Scholar]
- 14.Wehkamp J. NF-kB and AP-1 mediated induction of human beta defensin-2 in intestinal epithelial cells by E. coli Nissle 1917 a novel effect of a probiotic bacterium. Infect. Immun. 2004;72,:5750–5758. doi: 10.1128/IAI.72.10.5750-5758.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Schroder JM. Epithelial antimicrobial peptides: innate local host response elements. Cell. Mol. Life Sci. 1999;56,:32–46. doi: 10.1007/s000180050004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Huttner KM, Bevins CL. Antimicrobial peptides as mediators of epithelial host defense. Pediatr. Res. 1999;45,:785–794. doi: 10.1203/00006450-199906000-00001. [DOI] [PubMed] [Google Scholar]
- 17.Schlee M, Harder J, Koten B, Stange EF, Wehkamp J, Fellermann K. Probiotic lactobacilli and VSL#3 induce enterocyte beta-defensin 2. Clin. Exp. Immunol. 2008;151,:528–535. doi: 10.1111/j.1365-2249.2007.03587.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schlee M, Wehkamp J, Altenhoefer A, Oelschlaeger TA, Stange EF, Fellermann K. The induction of human beta-defensin-2 by the probiotic Escherichia coli Nissle 1917 is mediated through flagellin. Infect. Immun. 2007;75,:2399–2407. doi: 10.1128/IAI.01563-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Meyer-Hoffert U. Secreted enteric antimicrobial activity localises to the mucus surface layer. Gut. 2008;57,:764–771. doi: 10.1136/gut.2007.141481. [DOI] [PubMed] [Google Scholar]
- 20.Wehkamp J. Reduced Paneth cell α-defensins in ileal Crohn's disease. Proc. Natl. Acad. Sci. USA. 2005;102,:18129–18134. doi: 10.1073/pnas.0505256102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Langhorst J, Wieder A, Michalsen A, Musial F, Dobos GJ, Rueffer A. Activated innate immune system in irritable bowel syndrome? Gut. 2007;56,:1325–1326. doi: 10.1136/gut.2007.125005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lehrer RI, Rosenman M, Harwig SS, Jackson R, Eisenhauer P. Ultrasensitive assays for endogenous antimicrobial polypeptides. J. Immunol. Methods. 1991;137,:167–173. doi: 10.1016/0022-1759(91)90021-7. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
PowerPoint slide for Fig. 1
PowerPoint slide for Fig. 2
PowerPoint slide for Fig. 3
PowerPoint slide for Table 1