

Abstract
Fruit shape and size are important appearance and yield traits in cucumber, but the underlying genes and their regulatory mechanisms remain poorly understood. Here we identified a mutant with spherical fruits from an Ethyl Methane Sulfonate (EMS)-mutagenized library, named the qiu mutant. Compared with the cylindrical fruit shape in 32X (wild type), the fruit shape in qiu was round due to reduced fruit length and increased fruit diameter. MutMap analysis narrowed the candidate gene in the 6.47 MB range on Chr2, harboring the FS2.1 locus reported previously. A single-nucleotide polymorphism (SNP) (11359603) causing a truncated protein of CsaV3_2G013800, the homolog of tomato fruit shape gene SlTRM5, may underlie the fruit shape variation in the qiu mutant. Knockout of CsTRM5 by the CRISPR-Cas9 system confirmed that CsaV3_2G013800/CsTRM5 was the causal gene responsible for qiu. Sectioning analysis showed that the spherical fruit in qiu resulted mainly from increased and reduced cell division along the transverse and longitudinal directions, respectively. Meanwhile, the repressed cell expansion contributed to the decreased fruit length in qiu. Transcriptome profiling showed that the expression levels of cell-wall-related genes and abscisic acid (ABA) pathway genes were significantly upregulated in qiu. Hormone measurements indicated that ABA content was greatly increased in the qiu mutant. Exogenous ABA application reduced fruit elongation by inhibiting cell expansion in cucumber. Taken together, these data suggest that CsTRM5 regulates fruit shape by affecting cell division direction and cell expansion, and that ABA participates in the CsTRM5-mediated cell expansion during fruit elongation in cucumber.
Introduction
Cucumber (Cucumis sativus L.) is a vegetable crop with important economic and biological value. The cucumber fruit is a kind of pepo fruit with three fused carpels and parietal placentation [1]. Unlike most fruits harvested at the mature stage, cucumber is usually harvested at an immature stage for fresh consumption or production of processed pickles 1–2 weeks after floral anthesis [2]. In cucumber, fruit length varies from 5 to 60 cm, and fruit shape varies from round to cylindrical [3, 4]. Fruit size and shape have been under intensive selection during domestication due to their large effects on fruit appearance and yield in cucumber [5].
Fruits originate from the carpel primordium and subsequently undergo ovary development and fertilization, which is required for fruit development in most species [3, 6, 7]. Cell division occurs mainly in early fruit development, whereas cell expansion continues until final fruit size and shape are reached [8–10]. Subsequently, fruit morphological development ceases and the fruit enters the ripening stage [11]. Among fruit morphogenesis processes, the rate, duration and direction of cell division, as well as isotropic and anisotropic cell expansion, contribute greatly to the final fruit shape and size [11]. In horticultural crops, using tomato as the model plant for studying fruit morphogenesis, some regulators have been identified for fruit shape and size. Among these, the WUS (WUSCHEL)–CLV3 (CLAVATA3) pathway plays essential roles in floral meristem development and locule number determination in tomato [12–15]. Tomato varieties with eight or more locules nearly always carry both the fascinated (fas) and the locule number (lc) mutation. SlCLV3 and SlWUS underlie fas and lc, respectively [14, 15]. OVATE regulates gynoecium shape by controlling the cell division pattern in both the proximal–distal and the medial–lateral direction [16]. The OVATE family proteins (OFPs) interact with TONNEAU1 recruiting motif proteins (TRMs) to regulate fruit shape via antagonistic effects on fruit elongation in tomato [17]. SUN, which encodes a calmodulin binding protein, functions in fruit shape specification by increasing cell division in the longitudinal direction and decreasing cell division in the transverse direction of fruit [18]. In addition to cell division, dramatic enlargement in cell size led to increased fruit size [4, 10]. Cell Size Regulator (CSR), encoding an unknown protein, affects fruit size/weight by positively regulating cell enlargement in tomato [19].
In cucumber, fruit morphogenesis goes through three sequential phases: the fruit set phase, which involves ovary development before anthesis and fertilization; the cell division phase; and the cell expansion phase, which exhibits a typical sigmoidal pattern [3, 4]. Fruit shape is often described by fruit length (FL), fruit diameter (FD) and fruit shape index (FSI, the ratio of FL to FD) in cucumber [3]. Ovary shape and final fruit shape are generally highly correlated, indicating that cucumber fruit shape is determined before anthesis [20–22]. FL and FD largely describe fruit growth along proximal–distal and medial–lateral axes, respectively. A total of 42 quantitative trait loci (QTLs) were found associated with fruit shape (FS), FSI and fruit weight (FW), while only two QTLs were fine-mapped [3]. Specifically, FS1.2 and FS2.1 underlie the round fruit shape in WI7239 cucumber [22]. CsSUN, a homolog of the tomato fruit shape gene SUN, underlies FS1.2 [22]. CsTRM5, an ortholog of SlTRM5, was speculated to be the candidate gene for FS2.1 [17], but the specific functions of CsSUN and CsTRM5 remained elusive in cucumber. Additionally, three genes were identified as controlling fruit length in cucumber. SF1 (Short Fruit 1), encoding a RING-type E3 ligase, regulates fruit elongation by orchestrating the ethylene synthesis effect on cell division in developing fruit [23]. SF2 (Short Fruit 2) encodes a histone deacetylase complex1 (HDC1) homolog that acts on fruit length by regulating cell proliferation [24]. A gain-of-function allele of MADS-box transcription factor CsFUL1, CsFUL1A, has a negative effect on fruit length by inhibiting cell division and expansion in Asian long cucumber [25]. So far, the genes and their regulatory mechanisms underlying fruit shape variation remain poorly understood in cucumber.
In this study, a novel cucumber mutant with spherical fruits was identified and designated as qiu. Mapping data showed that CsTRM5 may underlie the fruit shape variation in qiu. Further analysis with the CRISPR-Cas9 knockout system confirmed that CsTRM5 was the causal gene responsible for the spherical fruit phenotype in qiu. Cytological, transcriptomic, and hormone treatment analyses showed that CsTRM5 regulated fruit shape by mediating cell division direction and cell expansion in cucumber.
Results
The cucumber qiu mutant produced shorter and wider fruits
To identify genes regulating fruit shape in cucumber, one mutant bearing ball-like fruits, named the qiu mutant, was chosen for further characterization from M3 lines among an EMS-mutagenized library (Supplementary Data Fig. S1A and B). Fruit development dynamics, including FL, FD, and FSI at 0, 10, and 30 DAA, were observed in wild-type (32X) and qiu mutant (Fig. 1A–F). Compared with 32X, the qiu mutant showed decreased fruit length and increased fruit diameter during different developmental stages (Fig. 1A–F). The FSI of qiu mutant fruit was close to 1 at 0, 10, and 30 DAA, while that of 32X was between 2.28 ± 0.29 and 2.83 ± 0.12 (Fig. 1F), and the resulting fruit shape was nearly spherical in qiu and cylindrical in 32X. Therefore, fruit morphogenesis is determined as early as anthesis in cucumber.
Figure 1.
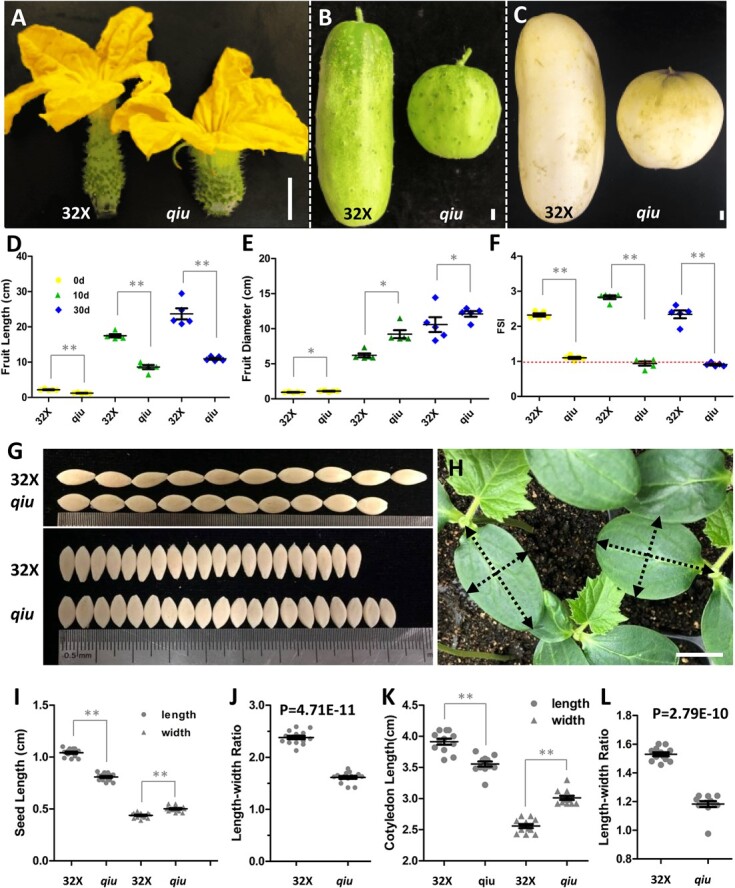
Phenotypic characterization of 32X and qiu mutant. A–C Fruits of 32X and qiu mutant at 0, 10 and 30 DAA. D–F Statistical data analysis of fruit length, diameter and FSI in 32X and qiu mutant. G, H Seed and cotyledon phenotypes in 32X and qiu. I, J Quantification data on seed length, width, and length/width ratio in 32X and qiu. K, L Quantification data on cotyledon length, width and length/width ratio. Scale bars: 1 cm in A–C and H.
In addition to fruit phenotype, the seeds in qiu mutant plants became shorter and wider compared with 32X (Fig. 1G). The length and width of seeds in qiu were 0.81 ± 0.04 and 0.50 ± 0.03 cm, a decrease of 22% and an increase of 28% compared with 32X, respectively (Fig. 1I and J). Similarly, cotyledons displayed the same changes as fruits and seeds in qiu compared with 32X (Fig. 1H, K, and L). Hypocotyl length, plant height, and male/female flower size in qiu were smaller compared with 32X (Supplementary Data Fig. S1C–F).
Inheritance analysis and mapping of the candidate gene for qiu mutant
To perform genetic analysis of qiu, an F2 population was constructed using 32X as the pistillate parent and qiu as the pollen parent. All F1 individuals by reciprocal crossing (32X × qiu and qiu × 32X) displayed a cylindrical fruit shape like that of 32X, indicating the dominance of cylindrical fruit shape over spherical fruit phenotype. Among 163 F2 individuals there was a total of 125 plants with cylindrical fruits and 38 plants with spherical fruits, representing a segregation ratio of 3:1 (χ2 = 0.247, χ20.05 = 3.841) (Table 1). The results verified that the spherical fruit phenotype of qiu was controlled by a single recessive gene.
Table 1. Inheritance analysis of fruit shape in qiu mutant in cucumber.
| Population | Cylinder-shaped | Spherical-shaped | Segregation ratio | χ 2 | χ 2 0.05 |
|---|---|---|---|---|---|
| F 1(32X × qiu) | 15 | 0 | |||
| F 1(qiu×32X) | 14 | 0 | |||
| F 2(32X × qiu) | 125 | 38 | 3:1 | 0.247 | 3.841 |
The fruit shape index of cylindrical fruit (like 32X) is between 2.0 and 3.0, and that of spherical fruit (like qiu) is between 0.9 and 1.2.
To identify the candidate region contributing to the spherical fruit phenotype in qiu, a modified MutMap was performed. After resequencing, a total of 30 277 Mb (99.27% coverage), 15 443 Mb (99.31% coverage), 17.617 Mb (99.23% coverage), and 22 844 Mb (99.22% coverage) clean reads for WT pool (32X), mutant pool (qiu), S-pool (lines with spherical fruit in F2), and C-pool (lines with cylindrical fruit in F2) plants were obtained, respectively (Supplementary Data Table S3). We detected 32 463 SNPs (homozygous between two parents) detected between the wild-type and mutant pools and used for SNP-index analysis. To validate correlation accuracy, we restricted the SNPs as follows: (i) those with SNP index value <0.3 and SNP depth <7 in both the C-pool and the S-pool were filtered out; (ii) those with a missing SNP index in the C-pool or S-pool were filtered out. After filtering, 8345 SNVs (single-nucleotide variants) were used for ΔSNP-index analysis and plotting. A total of 90 SNVs harboring a high ΔSNP index (out of 95% confidence values; SNP index >0.7 in S-pool and <0.3 in C-pool; SNP index >0.7 in C-pool and <0.3 in S-pool) were identified within the whole genome (Supplementary Data Table S4), and resulted in three peak regions (Fig. 2A). Among them, one peak harbored multiple SNVs located at 10.71–17.18 Mb on Chr2 and had high quality; the other two peaks each harbored only one SNV in Chr6 or Chr7 and had low confidence (Fig. 2A). Therefore, the peak in Chr2 was named as the qiu locus, which harbored the reported FS2.1 [22]. SNP annotation results showed that there were 11 SNPs in the exon region causing amino acid change within the qiu locus (Supplementary Data Table S5). Among them, only two SNPs (11359603 and 14075205) were located in the coding regions of cucumber functional proteins, and the remaining nine SNPs, encoding proteins of fungi or bacteria, were not mapped to the cucumber genome and thus were not considered as the candidate sites for qiu locus. The SNP from G to A (11359603) caused a premature stop codon and a severely truncated protein (from 776 to 373 amino acids) of CsaV3_2G013800, which encoded the homolog of SlTRM5 associated with fruit shape in tomato (Fig. 2C and D). The other SNP (14075205), a C-to-T transition from 32X to the qiu line, causing an alanine-to-threonine change in CsaV3_2G016800, encoded a WD40 repeat-like superfamily protein (Supplementary Data Table S5). Further protein sequence analysis showed that this amino acid is not conserved between species (Supplementary Data Fig. S2A). Expression analysis showed that the expression level of both CsaV3_2G013800 and CsaV3_2G016800 was unchanged in ovaries of 32X and qiu (Supplementary Data Fig. S2B and C). In conclusion, we assumed that CsaV3_2G013800 may be the candidate gene for qiu and the FS2.1 locus, and named it CsTRM5.
Figure 2.

Identification of the candidate gene in qiu mutant. A MutMap analysis identified the qiu locus (red box) in the cucumber genome. B MutMap analysis identified the qiu locus in Chr2. C Phylogenetic analysis of TRMs in cucumber, Arabidopsis, rice, and tomato. D Protein sequence information on CsTRM5 in 32X and qiu lines. The red box indicates the changed amino acid between the 32X and qiu lines.
Temporal and spatial expression pattern of CsTRM5
To explore the expression pattern of CsTRM5, qRT–PCR analysis was performed in different organs, including tip, stem, leaf, tendril, flower buds, flowers, and fruits at different stages in cucumber. CsTRM5 displayed a universal expression pattern (Fig. 3A). In situ hybridization showed that CsTRM5 was expressed in shoot apical meristem, floral meristem, leaf, and young flower buds (Fig. 3B–H). CsTRM5 transcripts were detected in sepal, petal, and stamen primordia in young flower buds (Fig. 3C–E). Among male flower buds, CsTRM5 was expressed in petal and stamen (Fig. 3E). CsTRM5 was expressed in whole fruit and highly expressed in placenta at stage 9 of female flower buds (Fig. 3F). Subsequently CsTRM5 signals were mainly enriched in ovule integuments (Fig. 3G). No signal was detected upon hybridization with the sense CsTRM5 probe (Fig. 3I).
Figure 3.
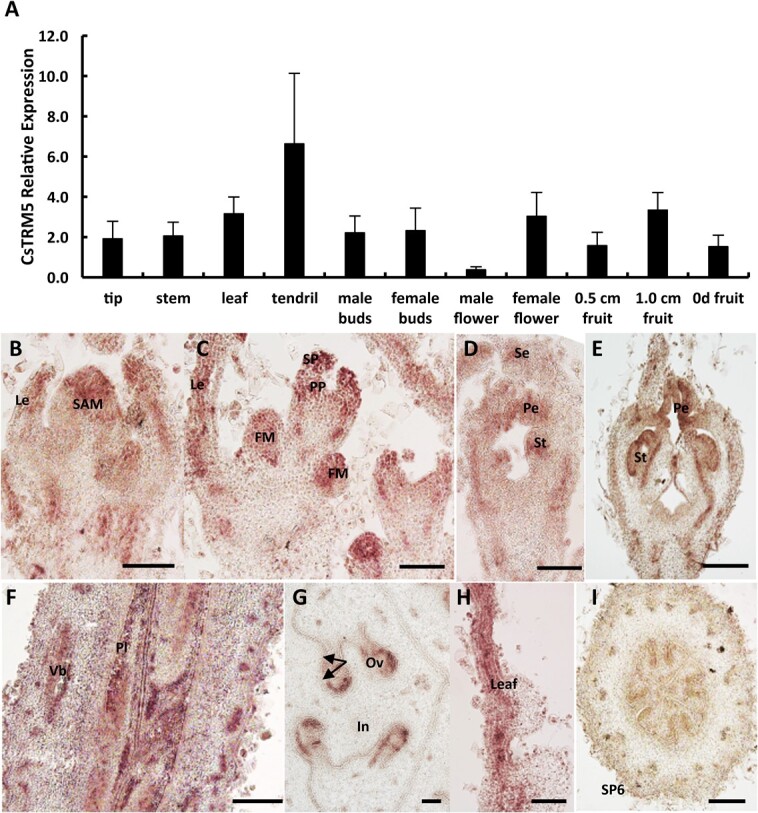
Expression pattern analysis of CsTRM5 in cucumber.A Expression levels of CsTRM5 in different organs of 32X line detected by qRT–PCR. B–HIn situ hybridization of CsTRM5 in cucumber shoot tip (B), leaf (H), and floral organs (C–G). I, Negative control of CsTRM5 sense probe in fruit cross-section. Le, leaf; SAM, shoot apical meristem; FM, floral meristem; SP, sepal primordium; PP, petal primordium; Se, sepal; Pe, petal; St, stamen; Vb, vascular bundle; Pl, placenta; Ov, ovule; In, integument. Scale bars: 100 μm.
Knockout of CsTRM5 resulted in decreased fruit shape index in cucumber
In order to confirm the function of CsTRM5 in cucumber, we constructed CsTRM5 knockout lines in XTMC using the CRISPR-Cas9 gene editing system. The two targets were located on the second and third exons, respectively (Supplementary Data Fig. S3A). Two homozygous loss-of-function mutant lines (Cstrm5-cr #1 and #2) were obtained for further characterization (Fig. 4A). Both Cstrm5-cr #1 and Cstrm5-cr #2 produced fruits with decreased length and increased diameter, resulting in a change in FSI from 4.9–5.6 in control to 2.3–2.9 in mutant lines (Fig. 4B–F), consistent with the phenotype of qiu and 32X (Fig. 1A–E). Similarly, the length/width ratio of seeds was also decreased, by 17 and 19% in Cstrm5-cr #1 and Cstrm5-cr #2, respectively, compared with XTMC (Fig. 4G–J). These data suggested that CsTRM regulates fruit and seed shape in cucumber.
Figure 4.

Mutation forms and phenotypes of Cstrm5-cr lines. A Mutation forms of two homozygous T1 transgenic Cstrm5-cr lines obtained using the CRISPR-Cas9 system. B, C Fruits at 10 and 30 DAA in XTMC and Cstrm5-cr lines. D, E Fruit length and diameter of XTMC and Cstrm5-cr lines. F FSI in XTMC and Cstrm5-cr lines. G Seed phenotype in XTMC and Cstrm5-cr lines. H–J Seed length, width, and length/width ratio in XTMC and Cstrm5-cr lines. Scale bars: 1 cm in B,C and G.
Mutation in CsTRM5 led to change in cell division direction in fruits
In both qiu and Cstrm5-cr lines, fruit length and diameter changed (Figs 1A–E and 4B–E). Further characterization of fruit internal morphogenesis during fruit development indicated that qiu fruit exhibited larger pericarp thickness (1.77 ± 0.21 cm in qiu, 1.42 ± 0.31 cm in 32X) and ventricle diameter (4.16 ± 0.27 cm in qiu, 3.65 ± 0.34 cm in 32X) than wild-type fruit (Fig. 5A–E), indicating that both pericarp and ventricle had a positive effect on the increase in fruit diameter. Locule number generally influences fruit diameter and size [1]. We checked the locule number of 32X and qiu and found that both possessed three locules (Fig. 5A–C), suggesting that locule number was not the cause of the increased diameter in qiu mutant.
Figure 5.

Cell division direction and cell expansion were changed in the qiu mutant. A–C Cross-sections and longitudinal sections of fruits at 0, 10 and 30 DAA in 32X and qiu mutant. D, E Pericarp thickness and ventricle diameter of fruit at 10 DAA in 32X and qiu. F Longitudinal paraffin section of fruit at 10 DAA in 32X and qiu. Gray double-headed arrow indicates ventricle diameter; black double-headed arrow indicates pericarp thickness. G, H Pericarp cell number per 1000 μm and total cell number in cross direction. I, J, Pericarp cell number per 1000 μm and total cell number in longitudinal direction. K, L PT, FL, cell size, and cell number ratio of qiu to 32X in cross (K) or longitudinal direction (L). Scale bars: 1 cm in A–C, 200 μm in F.
To further explore the cellular basis of the change in FSI, we evaluated pericarp thickness, cell size and cell number of the pericarp in the longitudinal and cross directions. Cucumber fruit can be roughly estimated as a cylinder, so the pericarp has the same length as the fruit. We dissected fruits longitudinally at 10 DAA and investigated the cell number per unit length in the cross and longitudinal directions in the pericarp. The cell number per unit length was increased in both the cross (9.6 ± 1.0 in qiu, 8.4 ± 0.7 in 32X) and the longitudinal (8.6 ± 1.1 in qiu, 5.8 ± 0.8 in 32X) direction, indicating that the cell size became smaller in qiu compared with 32X (Fig. 5G and I). We counted higher cell numbers in the cross direction in qiu (170.1 ± 20.1) than in 32X (119.2 ± 25.4) (Fig. 5D and F–H), indicating that there was more cell proliferation in the cross direction in the pericarp of qiu. Combining fruit length with cell number per unit length along the longitudinal direction, the total cell numbers were decreased in the longitudinal direction in qiu (762.3 ± 108.5) than in 32X (1005.2 ± 53.0) (Figs 3D and 5F, I, and J), suggesting less cell proliferation in the longitudinal direction. In addition, we calculated the proportions of pericarp thickness (PT), FL, cell size and cell number in qiu relative to 32X in both cross and longitudinal directions (Fig. 5K and L). The data showed that the increased PT in qiu is directly caused by the increase in cell number in the cross direction, and the decreased FL in qiu is caused by both reduced cell size and decreased cell number in the longitudinal direction (Fig. 5K and L). To sum up, cell division in the pericarp was enhanced in the cross direction and repressed in the longitudinal direction in qiu compared with 32X, while cell expansion was inhibited in both the cross and the longitudinal direction in qiu.
Likewise, we evaluated pericarp thickness and cell size and number in the longitudinal and cross directions in XTMC and Cstrm5-cr lines (Supplementary Data Fig. S3B–I). The relative data between XTMC and Cstrm5-cr lines displayed similar trends to that in 32X and qiu (Supplementary Data Fig. S3B–I and Fig. 5). Cell size became smaller in Cstrm5-cr lines in both the cross and the longitudinal direction, whereas cell number increased in the cross direction and decreased in the longitudinal direction in Cstrm5-cr lines compared with the wild-type (Supplementary Data Fig. S3B–I). These data suggested that CsTRM5 regulated fruit shape by functioning in the cell division direction and cell expansion in cucumber, in which the change in cell division direction plays a vital role.
The abscisic acid pathway was involved in fruit shape regulation by CsTRM5 in cucumber
To identify putative downstream targets of CsTRM5, RNA-seq of young fruits/ovaries at 0 DAA between 32X and qiu lines was performed and 43 DEGs were obtained, of which 28 were upregulated and 15 were downregulated (Supplementary Data Table S6). Cell elongation in plants requires addition and rearrangements of cell wall components. Among these DEGs, we examined seven genes that were reported to regulate cell wall activities (Fig. 6A). For example, CsaV3_1G030240, encoding a fasciclin-like arabinogalactan protein, displayed a 46.4-fold (log2 fold change = 5.54) increase in the qiu mutant (Supplementary Data Table S6). In Arabidopsis, fasciclin-like arabinogalactans (FLAs) contributed to the cell wall matrix by affecting cellulose deposition [38]. CsaV3_7G026220 displayed a 3.74-fold increase in the mutant and encoded a pectin methylesterase affecting cell elongation by modifying the cell wall [39]. In addition, DEG analysis showed that ABA- and cytokinin-related genes were significantly upregulated in qiu compared with those in 32X (Fig. 6B). ABA is the major regulator of senescence and stress and plays an important role in cell wall modifications, while cytokinin is involved in cell division [40, 41]. CsaV3_4G007760 was increased 3.4-fold in the qiu mutant and encoded 9-cis-epoxycarotenoid dioxygenase, whose homolog in Arabidopsis is a key enzyme in the biosynthesis of ABA; CsaV3_4G007770 was increased 16.27-fold in the qiu mutant, it encoded an ABCG-type transporter participating in ABA transport; CsaV3_3G026410 and CsaV3_3G013180 were increased 3.92- and 3.71-fold in the qiu mutant, respectively, and both function in the ABA signal pathway (Fig. 6B) [42–44]. qRT–PCR results validated the elevated expression of the above four ABA-related genes in young fruits of qiu compared with 32X (Supplementary Data Fig. S4). In addition, CsaV3_1G039330, a homolog of AHP (Arabidopsis thaliana histidine phosphotransfer protein), which positively regulates cytokinin signaling [45], was increased 22.59-fold in qiu compared with wild-type (Fig. 6B).
Figure 6.
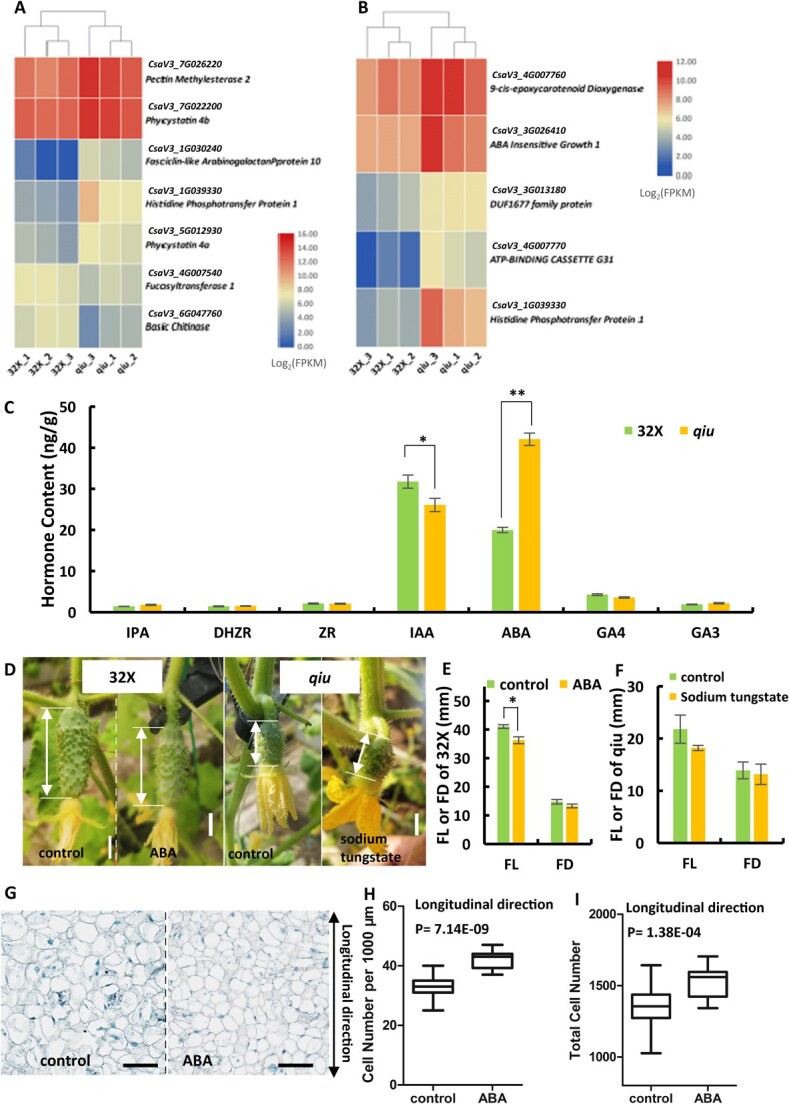
ABA plays an important role in fruit length variation in cucumber. A, B DEGs relative to cell wall development (A) and hormone (B) pathways between qiu and 32X lines. C Hormone content of young fruits in qiu and 32X lines. D Fruit phenotype of 32X and qiu lines after ABA or sodium tungstate treatment. E, F Change in FL and FD after ABA (E) or sodium tungstate (F) treatment. G Longitudinal section of fruit at 3 DAA in 32X after ABA treatment. H, I Pericarp cell number per 1000 μm and total cell number in longitudinal direction. Scale bars: 1 cm in D, 100 μm in G.
To further explore the relationship between CsTRM5 and hormone pathways, ovaries at the green bud stage from qiu and 32X were used for hormone measurements. Our data showed that the IAA level was significantly decreased, while ABA content was dramatically increased in qiu compared with 32X (Fig. 6C). Contents of cytokinin were not changed (Fig. 6C). IAA was shown to promote cell expansion by inducing cell-wall acidification in plants [46, 47]. In cucumber, auxin was reported to mediate fruit length and fruit neck elongation by cell division and cell expansion [25, 48]. In qiu, the reduced IAA content was consistent with smaller cell size and reduced FL in fruit (Fig. 5F–H). ABA is well known for mediating plant responses to abiotic stress by participating in cell expansion [49]. To verify the role of ABA in fruit shape regulation in cucumber, exogenous treatment with ABA or ABA biosynthesis inhibitor (sodium tungstate) was performed in 32X and qiu, respectively. Our data showed that fruit elongation in 32X was significantly reduced upon ABA treatment compared with control fruits (Fig. 6D and E). However, upon treatment with sodium tungstate, both FL and FD of qiu displayed no significant changes (Fig. 6D–F). Sectioning and statistical analysis showed that the cell size was significantly reduced in 32X after ABA treatment (Fig. 6G and H), suggesting that ABA was involved in the CsTRM5-mediated cell expansion in the longitudinal direction. Additionally, total cell number in 32X after ABA treatment was increased in longitudinal direction (Fig. 6I), not like the decreased total cell number in qiu compered to 32X (Fig. 5J), implying that ABA may not participate in CsTRM5-mediated cell division in cucumber.
Discussion
CsTRM5 was responsible for qiu mutant with spherical fruit shape
Recent studies have revealed that TRM family members control the shape of fruit, seed and leaf in tomato, Arabidopsis, and tobacco [17, 50–54]. TONNEAU1 (TON1) proteins share similarity with a human centrosomal protein and TRM1-TON1 recruitment is essential for microtubule organization in the cortex [30]. Plant microtubule arrays are involved in the direction of cell division and expansion [30]. In Arabidopsis, LONGIFOLIA1 (LNG1) and LONGIFOLIA2 (LNG2) promote longitudinal polar cell elongation to regulate leaf morphology [50]. LNG1/2/3/4 acted redundantly on leaf longitudinal growth by changing turgor pressure and activating XTHs (xyloglucan endotransglucosylase/hydrolases) [51]. PIF4 can induce thermomorphogenic growth by promoting LNG1/2 expression levels [55]. SLG7 (Slender Grain on Chromosome 7), encoding a protein homologous to AtLNG1 and AtLNG2, produces slender grains [52], and copy number variation of GL7 (Grain Length on Chromosome 7) contributes to the grain size diversity in rice [54]. In tomato, TRMs interact with OVATE family proteins to regulate cell division patterns in fruit shape by co-locating microtubules [17]. In cucumber, FS2.1 was mapped into 10 candidate genes, including a homolog of AtTRM5/SlTRM5 [17]. All these TRMs belong to the Arabidopsis LONGIFOLIA (TRM1–5) clade, indicating that LONGIFOLIA subclade members participated in organ morphology by changing cell division and cell elongation in different plant species.
In this study, we identified a novel mutant, qiu, with spherical fruits from an EMS-mutagenesis library. Inheritance analysis of qiu showed that the spherical fruit phenotype was controlled by a single recessive gene (Table 1). A modified MutMap method was used for mapping the qiu locus, which was delimited to a 6.47-MB region harboring the FS2.1 locus. In this region, one SNP caused a truncated protein of CsaV3_2G013800 (Supplementary Data Table S5). The phylogenetic analysis showed that CsaV3_2G013800 is an ortholog of AtTRM5, sharing 34.23% identity. In a previous study, CsTRM5 (CsaV3_2G013800/Csa2G227860) was also presumed to be the candidate gene for spherical fruit shape in the FS2.1 locus [17]. In this study, we speculated that CsaV3_2G013800 was the candidate gene for the spherical fruit shape in qiu and FS2.1. Considering the low transgenic efficiency of 32X, we constructed knockout lines of CsaV3_2G013800 using the CRISPR-Cas9 system in the XTMC inbred line. We found that the fruits of Cstrm5-cr lines were shorter and thicker, consistent with the change in the fruits of qiu. But fruit shape in Cstrm5-cr lines was a short cylinder, not round like qiu. This may be due to the fact that XTMC has a larger FSI than 32X. The FSI of Cstrm5-cr lines was half of that of XTMC, consistent with the FSI of qiu being about half of that of 32X (Figs 1F and 4F). Except for fruit shape, the change in seed and cotyledon shape in Cstrm5-cr lines is consistent with those in qiu. The above phenotypic analysis confirmed that CsaV3_2G013800/CsTRM5 was responsible for the spherical fruit shape in qiu.
CsTRM5 affects fruit shape by regulating cell division direction and cell expansion in cucumber
The fruit is a complex organ that develops along three axes: proximal–distal, medial–lateral, and abaxial–adaxial [56, 57]. Growth in the proximal–distal and medial–lateral axes is reflected in fruit length (longitudinal growth) and fruit diameter (cross growth), respectively. In cucumber, Pan et al. [3] summarized the fruit development dynamics of 11 genotypes from 6 days before anthesis to 30 DAA, including four lines with long cylindrical fruits, three lines with short cylindrical fruits, and four lines with spherical fruits. All these lines showed the typical sigmoidal pattern of FL and FD, and FSI of 0 DAA fruit and final mature fruit (30 DAA fruit) exhibited a high correlation (r = 0.9876), indicating that pre-anthesis factors play important roles in specifying fruit shape in cucumber [3]. The rate, duration, and direction of cell division, as well as isotropic and anisotropic cell expansion, together determined fruit shape and size [11, 17, 58].
The dynamics of fruit development at three points, including 0 DAA (ovary stage), 10 DAA (commercial fruit stage), and 30 DAA (mature fruit stage), were investigated in this study to examine fruit shape change between qiu and 32X. The results showed that fruit shape was determined at the pre-anthesis stage. The smaller cell size in the cross direction and larger fruit diameter in qiu indicated that its fruit growth along the medial–lateral axis is mainly due to enhanced cell proliferation in this direction (Fig. 5). Combining data on cell size along the longitudinal direction and fruit length of qiu and 32X, it can be concluded that both cell division and cell expansion were reduced along the proximal–distal direction in qiu. Similar data were found in Cstrm5-cr knockout lines (Supplementary Data Fig. S3). These findings suggested that CsTRM5 regulates fruit shape by modifying the cell division direction and cell expansion in cucumber. In tomato, Sltrm5 could rescue the fruit elongation phenotype of the double mutant ovate/suppressor of ovate (sov1) by antagonizing the role in cell division pattern of the proximal portion of the fruit in the proximo–distal direction, while the trm5 mutant alone produced fruits with similar shape to that of wild-type [17]. In cucumber, the fruit length variations were much greater than in tomato. Consistently, the effect of CsTRM5 on fruit shape regulation was much more dramatic than that of SlTRM5, suggesting the crucial role of CsTRM5 in fruit shape determination in cucumber.
Abscisic acid may be involved in CsTRM5-mediated fruit elongation in cucumber
Plant hormones are closely related to fruit shape. During the fruit morphogenesis process, cell division and cell expansion contribute greatly to final fruit shape and size [11]. In this study, among 43 DEGs between 32X and qiu, four ABA-related genes, and one cytokinin-related gene were identified (Fig. 6B). One key enzyme (CsaV3_4G007760) in the biosynthesis of ABA and one ABA transporter (CsaV3_4G007770) in qiu were upregulated ~3.4- and ~16.3-fold over 32X (Supplementary Data Table S6). The ABA content was dramatically increased in qiu compared with 32X (Fig. 6C), implying that ABA may act downstream of CsTRM5 during fruit shape determination. ABA is well known as the major player in senescence and stress through cell wall modifications [59]. Cell elongation in plants requires addition and rearrangements of cell wall components [60]. Cellulose and pectin are components of the cell wall. Seven genes regulating cell wall activities were identified among DEGs, including FLA (CsaV3_1G030240) affecting cellulose deposition, and CsaV3_7G026220, modifying the cell wall as a pectin methylesterase [38, 39]. CsTRM5 is a member of the LONGIFOLIA subclade, whose member LNG3/4 could activate cell wall modification-related genes to affect cell elongation [51].
Exogenous ABA treatment resulted in reduced fruit elongation in 32X by inhibiting cell size in the longitudinal direction, but displayed no effect on fruit diameter (Fig. 6D–H). Unlike the decreased total cell number in qiu in the longitudinal direction compared to 32X (Fig. 5J), the total cell number in 32X was increased after ABA treatment (Fig. 6I). These results indicated that ABA is involved in CsTRM5-mediated cell expansion during fruit elongation, but not in the regulation of cell division direction during fruit shape formation. Whether and how IAA or other hormone pathways play roles in the CsTRM5-mediated cell division direction need further studies in cucumber.
Materials and methods
Plant materials and growth conditions
The material of 32X (South China type cucumber) with short cylindrical fruits was a high-generation inbred line. The mutant qiu with spherical fruits was discovered among M3 lines in an Ethyl Methane Sulfonate (EMS)-mutagenized population of the 32X line. The 32X line as female parent was crossed with qiu as male parent to develop 15 F1-1 plants and the reciprocal cross produced 14 F1-2 plants. A total of 163 F2 plants were produced by self-crossing of F1-1 for an inheritance study, genotyping, and gene identification. Also used in the study was XTMC (North China type), with long fruits, also a high-generation inbred line. The materials used in this study were planted according to standard practices in greenhouses in China Agricultural University, Beijing, or in Hebei Normal University of Science and Technology, Qinhuangdao.
Phenotype analysis
Fruit measurements were taken of three fruits between the 15th and 25th nodes of each plant, and five or more plants were measured. To ensure sufficient seed set, open flowers were tagged and hand-pollinated. Seed length and width were measured using 20 seeds. For cotyledon shape analysis, cotyledons were collected from 10 or more seedlings at the one true leaf stage.
Identification of the candidate gene for qiu by MutMap
A modified MutMap method was used to identify the candidate gene for qiu. A total of 20 lines with cylindrical fruits and 20 lines with spherical fruits were selected from the F2 population. DNA was extracted from each plant and mixed in equal amounts to construct the cylindrical fruit bulk (C-pool) and the spherical fruit mutant bulk (S-pool). DNA from the two parental plants (20 individuals) was extracted to construct the wild-type pool (P1, 32X) and the mutant pool (P2, qiu), respectively. DNA from the two parental plants, C-pool and S-pool, were re-sequenced with Illumina HiSeq TMPE150. The raw reads were filtered using the NGSQC toolkit software [26]. BWA was used to align the clean reads to the reference genome [27], and single-nucleotide polymorphisms (SNPs) were detected using the Genome Analysis Toolkit (GATK) [28]. ANNOVAR software was used for functional annotation for SNP detection results [29].
Phylogenetic tree construction
Arabidopsis TRM proteins (TRM1, TRM2, TRM3, TRM4, TRM5, TRM6) and the homologs in tomato, rice, and cucumber were used for phylogenetic tree analysis [30]. Protein sequences of TRMs from cucumber were obtained using BLAST tools in CuGenDB (Chinese Long_V3.0). All protein sequences were aligned and constructed using MEGA6.0 by the neighbor-joining method. Accession numbers of TRM sequences used for phylogenetic analysis are provided in Supplementary Data Table S1.
Quantitative real-time PCR
Plants at blooming and fruit-bearing stage were used for sampling under the long-day condition. The young shoot tip, stem, leaf, tendril, male buds, female buds, male flower, female flower, fruit at anthesis, and fruits of 0.5 and 1.0 cm were sampled for RNA extraction using Trizol reagent, then reverse-transcribed to cDNA using the TianScript II RT Kit (Tiangen, China). Quantitative real-time PCR (qRT–PCR) was performed using TB Green™ Premix Ex Taq™ (Takara, Japan) on the Bio-Rad CFX384 system. UBIQUITIN EXTENSION PROTEIN (UBI-EP, Csa000874) was used as the internal reference gene [31]. Three biological replicates and three technical replicates were performed for each gene. The primer information is listed in Supplementary Data Table S2.
Paraffin sectioning
Fruit mesocarps were sampled along the longitudinal direction with 3.7% formalin–acetic acid–alcohol (FAA) overnight, dehydrated with a grade series of ethanol, infiltrated with xylene, and then embedded in paraffin (Leica, 39 601 095, Germany). The sections (10 μm) were used for cell morphology observation by deparaffinating and rehydration under a D72 light microscope (Olympus).
In situ hybridization
The 20-day-old shoot tips and male and female flower buds were fixed with 3.7% FAA, then embedded in paraffin. Sense and antisense probes were amplified with gene-specific primers (Supplementary Data Table S2) using SP6 and T7 RNA polymerase with the DIG RNA Labeling Kit (SP6/T7) (Roche, USA), respectively. Sample sectioning and hybridization were performed as described previously [32].
Cucumber transformation
Two target sequences (19 bp) located in the second and third exons were used for construction of a CRISPR-Cas9 knockout vector (pKSE402 with GFP fluorescent screening marker pCBC-DT1T2 as an intermediate vector) (Supplementary Data Table S2). The recombinant knockout vector was transformed into Agrobacterium tumefaciens EHA105. The XTMC line, with long fruits, had the same CsTRM5 genotype as the 32X line and a well-established CRISPR system, and was thus used as transgenic background material [33]. Cotyledons at 36 hours after germination as the explants were infected with EHA105 (in infection liquid with OD600 = 0.2–0.3) under negative pressure. After 3 days of co-culture in darkness, explants were transferred to bud differentiation medium with Timentin (200 μg/l) under 16 hours light/8 hours dark at 26°C for 3–4 weeks. Then the buds with GFP were selected and excised from explant to rooting medium. Homozygous T1 mutants without vector (GFP-free) were identified from T0 transgenic lines for further phenotype observation and data statistics. The detailed cucumber transformation protocol was as described previously [34]. Primer information is listed in Supplementary Data Table S2.
Transcriptome analysis
Fruits at 0 days after anthesis (DAA) from 32X and qiu were collected for RNA-seq analysis. Three biological replicates were performed. RNA-seq libraries were constructed using the NEB Next Ultra 488 Directional RNA Library Prep Kit (NEB, USA), then loaded on to an Illumina HiSeq 4000 platform to generate 150-bp paired-end reads. The cut-offs for differentially expressed genes (DEGs) were a change in expression of at least 2.0-fold and a false discovery rate (FDR) of <0.05. The Gene Ontology (GO) terms or categories with a P-value <.05 were identified as significantly enriched. Sequencing data were deposited in the NCBI Gene Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/) under accession number PRJNA804348.
Hormone quantification by enzyme-linked immunosorbent assay
Ovaries at green bud stage of qiu and 32X were harvested to determine endogenous hormone contents, including indoleacetic acid (IAA), abscisic acid (ABA), gibberellic acids (GA3 and GA4), and three types of cytokinin: indolepropionic acid (IPA), trans-zeatin-riboside (ZR), and dihydrozeatinauxin (DHZR). About 0.5 g of fresh samples was collected and homogenized in 2 ml of 80% methanol. Extraction and hormone quantification were performed using the enzyme-linked immunosorbent assay (ELISA) following the protocol described in previous reports [35, 36]. Three biological replicates were performed for each genotype.
Exogenous abscisic acid and abscisic acid inhibitor application
ABA and its synthesis inhibitor sodium tungstate were used as described previously [37]. 32X was treated with 50 ABA or 0 (control) μmol/l ABA (Shanghai Yuanye Bio-Technology Co., Ltd, CAS#14375-45-2), a synthetic ABA. Meanwhile, qiu was treated with 3 or 0 (control) mmol/l sodium tungstate (Tianjin Oubokai Chemical Co. LTD, CAS#10213-10-2). The ovary and corolla were sprayed once at anthesis and fruit measurement was performed 3 days after treatment. The results are the means ± SE of at least three replicates.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31772327 and 32025033), the Key R&D Program of Hebei Province (21326309D), the Scientific Research Foundation of Hebei Normal University of Science and Technology (2019YB015 and 2020JK004), and the Natural Science Foundation of Hebei Province (C2020407015).
Author contributions
X.Zhang, L.Y., and X.Liu designed the research; Y.X., X.Liu, and C.S. performed the experiments; X.Zhang, X.Liu, and Y.X. wrote the paper; X.S., X.Li, H.C., J.G., L.L., A.Y., Z.Z., and X.Zhu provided experimental assistance; all authors revised the manuscript.
Data availability
The datasets have been submitted to the NCBI-SRA database with the BioProject ID PRJNA804348.
Conflict of interest statement
The authors declare no conflict of interests.
Supplementary data
Supplementary data is available at Horticulture Research online.
Supplementary Material
Contributor Information
Yang Xie, Hebei Key Laboratory of Horticultural Germplasm Excavation and Innovative Utilization, College of Horticulture Science and Technology, Hebei Normal University of Science and Technology, Qinhuangdao 066004, China.
Xiaofeng Liu, State Key Laboratories of Agrobiotechnology, Joint International Research Laboratory of Crop Molecular Breeding, Beijing Key Laboratory of Growth and Developmental Regulation for Protected Vegetable Crops, Department of Vegetable Sciences, China Agricultural University, Beijing 100193, China; Engineering Laboratory of Genetic Improvement of Horticultural Crops of Shandong Province, College of Horticulture, Qingdao Agricultural University, Qingdao 266109, China.
Chengzhen Sun, Hebei Key Laboratory of Horticultural Germplasm Excavation and Innovative Utilization, College of Horticulture Science and Technology, Hebei Normal University of Science and Technology, Qinhuangdao 066004, China.
Xiaofei Song, Hebei Key Laboratory of Horticultural Germplasm Excavation and Innovative Utilization, College of Horticulture Science and Technology, Hebei Normal University of Science and Technology, Qinhuangdao 066004, China.
Xiaoli Li, Hebei Key Laboratory of Horticultural Germplasm Excavation and Innovative Utilization, College of Horticulture Science and Technology, Hebei Normal University of Science and Technology, Qinhuangdao 066004, China.
Haonan Cui, Hebei Key Laboratory of Horticultural Germplasm Excavation and Innovative Utilization, College of Horticulture Science and Technology, Hebei Normal University of Science and Technology, Qinhuangdao 066004, China.
Jingyu Guo, State Key Laboratories of Agrobiotechnology, Joint International Research Laboratory of Crop Molecular Breeding, Beijing Key Laboratory of Growth and Developmental Regulation for Protected Vegetable Crops, Department of Vegetable Sciences, China Agricultural University, Beijing 100193, China.
Liu Liu, State Key Laboratories of Agrobiotechnology, Joint International Research Laboratory of Crop Molecular Breeding, Beijing Key Laboratory of Growth and Developmental Regulation for Protected Vegetable Crops, Department of Vegetable Sciences, China Agricultural University, Beijing 100193, China.
Ao Ying, State Key Laboratories of Agrobiotechnology, Joint International Research Laboratory of Crop Molecular Breeding, Beijing Key Laboratory of Growth and Developmental Regulation for Protected Vegetable Crops, Department of Vegetable Sciences, China Agricultural University, Beijing 100193, China.
Zeqin Zhang, State Key Laboratories of Agrobiotechnology, Joint International Research Laboratory of Crop Molecular Breeding, Beijing Key Laboratory of Growth and Developmental Regulation for Protected Vegetable Crops, Department of Vegetable Sciences, China Agricultural University, Beijing 100193, China.
Xueyun Zhu, Hebei Key Laboratory of Horticultural Germplasm Excavation and Innovative Utilization, College of Horticulture Science and Technology, Hebei Normal University of Science and Technology, Qinhuangdao 066004, China.
Liying Yan, Hebei Key Laboratory of Horticultural Germplasm Excavation and Innovative Utilization, College of Horticulture Science and Technology, Hebei Normal University of Science and Technology, Qinhuangdao 066004, China.
Xiaolan Zhang, State Key Laboratories of Agrobiotechnology, Joint International Research Laboratory of Crop Molecular Breeding, Beijing Key Laboratory of Growth and Developmental Regulation for Protected Vegetable Crops, Department of Vegetable Sciences, China Agricultural University, Beijing 100193, China.
References
- 1. Li S, Pan Y, Wen Cet al. . Integrated analysis in bi-parental and natural populations reveals CsCLAVATA3 (CsCLV3) underlying carpel number variations in cucumber. Theor Appl Genet. 2016;129:1007–22. [DOI] [PubMed] [Google Scholar]
- 2. Ando K, Carr KM, Grumet R. Transcriptome analyses of early cucumber fruit growth identifies distinct gene modules associated with phases of development. BMC Genomics. 2012;13:518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Pan Y, Wang Y, McGregor Cet al. . Genetic architecture of fruit size and shape variation in cucurbits: a comparative perspective. Theor Appl Genet. 2020;133:1–21. [DOI] [PubMed] [Google Scholar]
- 4. Gillaspy G, Ben-David H, Gruissem W. Fruits: a developmental perspective. Plant Cell. 1993;5:1439–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Che G, Zhang X. Molecular basis of cucumber fruit domestication. Curr Opin Plant Biol. 2019;47:38–46. [DOI] [PubMed] [Google Scholar]
- 6. Monforte AJ, Diaz A, Caño-Delgado Aet al. . The genetic basis of fruit morphology in horticultural crops: lessons from tomato and melon. J Exp Bot. 2014;65:4625–37. [DOI] [PubMed] [Google Scholar]
- 7. Roeder AH, Yanofsky MF. Fruit development in Arabidopsis. Arabidopsis Book. 2006;4:e0075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Azzi L, Deluche C, Gévaudant Fet al. . Fruit growth-related genes in tomato. J Exp Bot. 2015;66:1075–86. [DOI] [PubMed] [Google Scholar]
- 9. Eldridge T, Łangowski Ł, Stacey Net al. . Fruit shape diversity in the Brassicaceae is generated by varying patterns of anisotropy. Development. 2016;143:3394–406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Xiao H, Radovich C, Welty Net al. . Integration of tomato reproductive developmental landmarks and expression profiles, and the effect of SUN on fruit shape. BMC Plant Biol. 2009;9:49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Knaap E, Ostergaard L. Shaping a fruit: developmental pathways that impact growth patterns. Semin Cell Dev Biol. 2018;79:27–36. [DOI] [PubMed] [Google Scholar]
- 12. Lippman Z, Tanksley SD. Dissecting the genetic pathway to extreme fruit size in tomato using a cross between the small-fruited wild species Lycopersicon pimpinellifolium and L. esculentum var. Giant Heirloom. Genetics. 2001;158:413–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Barrero LS, Tanksley SD. Evaluating the genetic basis of multiple-locule fruit in a broad cross section of tomato cultivars. Theor Appl Genet. 2004;109:669–79. [DOI] [PubMed] [Google Scholar]
- 14. Xu C, Liberatore KL, MacAlister CAet al. . A cascade of arabinosyltransferases controls shoot meristem size in tomato. Nat Genet. 2015;47:784–92. [DOI] [PubMed] [Google Scholar]
- 15. Munos S, Ranc N, Botton Eet al. . Increase in tomato locule number is controlled by two single-nucleotide polymorphisms located near WUSCHEL. Plant Physiol. 2011;156:2244–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Wu S. The roles of OVATE and other elongation genes in regulating proximal-distal patterning of tomato fruit. Ph.D. Thesis, The Ohio State University; 2015. [Google Scholar]
- 17. Wu S, Zhang B, Keyhaninejad Net al. . A common genetic mechanism underlies morphological diversity in fruits and other plant organs. Nat Commun. 2018;9:4734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Wu S, Xiao H, Cabrera Aet al. . SUN regulates vegetative and reproductive organ shape by changing cell division patterns. Plant Physiol. 2011;157:1175–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Mu Q, Huang Z, Chakrabarti Met al. . Fruit weight is controlled by Cell Size Regulator encoding a novel protein that is expressed in maturing tomato fruits. PLoS Genet. 2017;13:e1006930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Sinnott EW. A developmental analysis of inherited shape differences in cucurbit fruits. Am Nat. 1936;70:245–54. [Google Scholar]
- 21. Weng Y, Colle M, Wang Yet al. . QTL mapping in multiple populations and development stages reveals dynamic quantitative trait loci for fruit size in cucumbers of different market classes. Theor Appl Genet. 2015;128:1747–63. [DOI] [PubMed] [Google Scholar]
- 22. Pan Y, Liang X, Gao Met al. . Round fruit shape in WI7239 cucumber is controlled by two interacting quantitative trait loci with one putatively encoding a tomato SUN homolog. Theor Appl Genet. 2017;130:573–86. [DOI] [PubMed] [Google Scholar]
- 23. Xin T, Zhang Z, Li Set al. . Genetic regulation of ethylene dosage for cucumber fruit elongation. Plant Cell. 2019;31:1063–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Zhang Z, Wang B, Wang Set al. . Genome-wide target mapping shows histone deacetylase complex1 regulates cell proliferation in cucumber fruit. Plant Physiol. 2020;182:167–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Zhao J, Jiang L, Che Get al. . A functional allele of CsFUL1 regulates fruit length through repressing CsSUP and inhibiting auxin transport in cucumber. Plant Cell. 2019;31:1289–307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Wang N, Zhang Y, Huang Set al. . Defect in Brnym1, a magnesium-dechelatase protein, causes a stay-green phenotype in an EMS-mutagenized Chinese cabbage (Brassica campestris L. ssp. pekinensis) line. Hortic Res. 2020;7:8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Li H, Durbin R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics. 2010;26:589–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. McKenna A, Hanna M, Banks Eet al. . The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010;20:1297–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Wang K, Li M, Hakonarson H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010;38:e164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Drevensek S, Goussot M, Duroc Yet al. . The Arabidopsis TRM1–TON1 interaction reveals a recruitment network common to plant cortical microtubule arrays and eukaryotic centrosomes. Plant Cell. 2012;24:178–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Wan H, Zhao Z, Qian Cet al. . Selection of appropriate reference genes for gene expression studies by quantitative real-time polymerase chain reaction in cucumber. Anal Biochem. 2010;399:257–61. [DOI] [PubMed] [Google Scholar]
- 32. Zhang X, Zhou Y, Ding Let al. . Transcription repressor HANABA TARANU controls flower development by integrating the actions of multiple hormones, floral organ specification genes, and GATA3 family genes in Arabidopsis. Plant Cell. 2013;25:83–101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Che G, Pan Y, Liu Xet al. . Natural variation in CRABS CLAW contributes to fruit length divergence in cucumber. Plant Cell. 2022;koac335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Hu B, Li D, Liu Xet al. . Engineering non-transgenic gynoecious cucumber using an improved transformation protocol and optimized CRISPR/Cas9 system. Mol Plant. 2017;10:1575–8. [DOI] [PubMed] [Google Scholar]
- 35. Wang Y, Li B, Du Met al. . Mechanism of phytohormone involvement in feedback regulation of cotton leaf senescence induced by potassium deficiency. J Exp Bot. 2012;63:5887–901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Yang J, Zhang J, Wang Zet al. . Hormonal changes in the grains of rice subjected to water stress during grain filling. Plant Physiol. 2001;127:315–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Li DD, XiaoWei Z, FengJiao Let al. . Hydrogen sulfide interacting with abscisic acid counteracts oxidative damages against chilling stress in cucumber seedlings. Acta Hortic Sin. 2018;45:2395–406. [Google Scholar]
- 38. MacMillan CP, Mansfield SD, Stachurski ZHet al. . Fasciclin-like arabinogalactan proteins: specialization for stem biomechanics and cell wall architecture in Arabidopsis and Eucalyptus. Plant J. 2010;62:689–703. [DOI] [PubMed] [Google Scholar]
- 39. Hewezi T, Howe P, Maier TRet al. . Cellulose binding protein from the parasitic nematode Heterodera schachtii interacts with Arabidopsis pectin methylesterase: cooperative cell wall modification during parasitism. Plant Cell. 2008;20:3080–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Forlani S, Masiero S, Mizzotti C. Fruit ripening: the role of hormones, cell wall modifications, and their relationship with pathogens. J Exp Bot. 2019;70:2993–3006. [DOI] [PubMed] [Google Scholar]
- 41. Yang W, Cortijo S, Korsbo Net al. . Molecular mechanism of cytokinin-activated cell division in Arabidopsis. Science. 2021;371:1350–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Nambara E, Marion-Poll A. Abscisic acid biosynthesis and catabolism. Annu Rev Plant Biol. 2005;56:165–85. [DOI] [PubMed] [Google Scholar]
- 43. Kang J, Yim S, Choi Het al. . Abscisic acid transporters cooperate to control seed germination. Nat Commun. 2015;6:8113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Preciado J, Begcy K, Liu T. The Arabidopsis HD ZIP class II transcription factor ABIG1 functions in leaf development. J Exp Bot. 2022;73:1978–91. [DOI] [PubMed] [Google Scholar]
- 45. Liu Z, Yuan L, Song Xet al. . AHP2, AHP3, and AHP5 act downstream of CKI1 in Arabidopsis female gametophyte development. J Exp Bot. 2017;68:3365–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Lin W, Zhou X, Tang Wet al. . TMK-based cell-surface auxin signalling activates cell-wall acidification. Nature. 2021;599:278–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Cao M, Chen R, Li Pet al. . TMK1-mediated auxin signalling regulates differential growth of the apical hook. Nature. 2019;568:240–3. [DOI] [PubMed] [Google Scholar]
- 48. Wang Z, Zhou Z, Wang Let al. . The CsHEC1-CsOVATE module contributes to fruit neck length variation via modulating auxin biosynthesis in cucumber. Proc Natl Acad Sci USA. 2022;119:e2209727229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Chen K, Li GJ, Bressan RAet al. . Abscisic acid dynamics, signaling, and functions in plants. J Integr Plant Biol. 2020;62:25–54. [DOI] [PubMed] [Google Scholar]
- 50. Lee YK, Kim GT, Kim IJet al. . LONGIFOLIA1 and LONGIFOLIA2, two homologous genes, regulate longitudinal cell elongation in Arabidopsis. Development. 2006;133:4305–14. [DOI] [PubMed] [Google Scholar]
- 51. Lee YK, Rhee JY, Lee SHet al. . Functionally redundant LNG3 and LNG4 genes regulate turgor-driven polar cell elongation through activation of XTH17 and XTH24. Plant Mol Biol. 2018;97:23–36. [DOI] [PubMed] [Google Scholar]
- 52. Zhou Y, Miao J, Gu Het al. . Natural variations in SLG7 regulate grain shape in rice. Genetics. 2015;201:1591–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Wang S, Li S, Liu Qet al. . The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nat Genet. 2015;47:949–54. [DOI] [PubMed] [Google Scholar]
- 54. Wang Y, Xiong G, Hu Jet al. . Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat Genet. 2015;47:944–8. [DOI] [PubMed] [Google Scholar]
- 55. Hwang G, Zhu JY, Lee YKet al. . PIF4 promotes expression of LNG1 and LNG2 to induce thermomorphogenic growth in Arabidopsis. Front Plant Sci. 2017;8:1320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Knaap E, Chakrabarti M, Chu YHet al. . What lies beyond the eye: the molecular mechanisms regulating tomato fruit weight and shape. Front Plant Sci. 2014;5:227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Lazzaro MD, Wu S, Snouffer Aet al. . Plant organ shapes are regulated by protein interactions and associations with microtubules. Front Plant Sci. 2018;9:1766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Colle M, Weng Y, Kang Yet al. . Variation in cucumber (Cucumis sativus L.) fruit size and shape results from multiple components acting pre-anthesis and post-pollination. Planta. 2017;246:641–58. [DOI] [PubMed] [Google Scholar]
- 59. Jahan MS, Shu S, Wang Yet al. . Melatonin pretreatment confers heat tolerance and repression of heat-induced senescence in tomato through the modulation of ABA- and GA-mediated pathways. Front Plant Sci. 2021;12:381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Irshad M, Canut H, Borderies Get al. . A new picture of cell wall protein dynamics in elongating cells of Arabidopsis thaliana: confirmed actors and newcomers. BMC Plant Biol. 2008;8:94. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets have been submitted to the NCBI-SRA database with the BioProject ID PRJNA804348.