

Abstract
To synchronize flowering time with spring, many plants undergo vernalization, a floral-promotion process triggered by exposure to long-term winter cold. In Arabidopsis thaliana, this is achieved through cold-mediated epigenetic silencing of the floral repressor, FLOWERING LOCUS C (FLC). COOLAIR, a cold-induced antisense RNA transcribed from the FLC locus, has been proposed to facilitate FLC silencing. Here, we show that C-repeat (CRT)/dehydration-responsive elements (DREs) at the 3′-end of FLC and CRT/DRE-binding factors (CBFs) are required for cold-mediated expression of COOLAIR. CBFs bind to CRT/DREs at the 3′-end of FLC, both in vitro and in vivo, and CBF levels increase gradually during vernalization. Cold-induced COOLAIR expression is severely impaired in cbfs mutants in which all CBF genes are knocked-out. Conversely, CBF-overexpressing plants show increased COOLAIR levels even at warm temperatures. We show that COOLAIR is induced by CBFs during early stages of vernalization but COOLAIR levels decrease in later phases as FLC chromatin transitions to an inactive state to which CBFs can no longer bind. We also demonstrate that cbfs and FLCΔCOOLAIR mutants exhibit a normal vernalization response despite their inability to activate COOLAIR expression during cold, revealing that COOLAIR is not required for the vernalization process.
Research organism: A. thaliana
eLife digest
Long spells of cold winter weather may feel miserable, but they are often necessary for spring to blossom. Indeed, many plants need to face a prolonged period of low temperatures to be able to flower; this process is known as vernalization.
While the molecular mechanisms which underpin vernalization are well-known, it is still unclear exactly how plants can ‘sense’ the difference between short and long periods of cold. Jeon, Jeong et al. set out to explore this question by focusing on COOLAIR, one of the rare genetic sequences identified as potentially being able to trigger vernalization. COOLAIR is a long noncoding RNA, a partial transcript of a gene that will not be ‘read’ by the cell to produce a protein but which instead regulates how and when certain genes are being switched on. COOLAIR emerges from the locus of the FLC gene, which is one of the main repressors of flowering, and it gradually accumulates in the plant when temperatures remain low for a long period. While some evidence suggests that COOLAIR may help to switch off FLC, other studies have raised some doubts about its involvement in vernalization.
In response, Jeon, Jeong et al. examined the FLC gene in a range of plants closely related to A. thaliana, and in which COOLAIR also accumulates upon cold exposure. This helped them identify a class of proteins, known as CBFs, which could bind to sequences near the FLC gene to activate the production of COOLAIR when the plants were kept in cold conditions for a while. CBFs were already known to help plants adapt to short cold snaps, but these experiments confirmed that they could act as both short- and long-term cold sensors.
This work allowed Jeon, Jeong et al. to propose a model in which CBF and therefore COOLAIR levels increase as the cold persists, until changes in the structure of the FLC gene prevent CBF from binding to it and COOLAIR production drops. Unexpectedly, examining the fate of mutants which could not produce COOLAIR revealed that these plants could still undergo vernalization, suggesting that the long noncoding RNA is in fact not necessary for this process.
These results should prompt other scientists to further investigate the role of COOLAIR in vernalization; they also give insight into how coding and noncoding sequences may have evolved together in various members of the A. thaliana family to adapt to the environment.
Introduction
Appropriate timing of flowering provides evolutionary advantage of plant reproductive success. As sessile organisms, plants have evolved mechanisms through which seasonal cues coordinate the transition to flowering. One of the significant environmental factors affecting flowering time of plants adapted to temperate climates is the temperature changes during the seasons, and plants have evolved complex sensory mechanisms to monitor the surrounding temperature to properly control the timing of flowering and tolerate thermal stress (Went, 1953; Penfield, 2008; Ding and Yang, 2022).
Cold acclimation and vernalization are two responses of plants to low temperatures. Cold acclimation is generally initiated by a short period of non-freezing cold exposure and increases the frost tolerance of plants (Weiser, 1970; Gilmour et al., 1988; Guy, 1990; Thomashow, 1999; Chinnusamy et al., 2007). The three C-REPEAT (CRT)/DEHYDRATION-RESPONSIVE ELEMENT (DRE) BINDING FACTORs (CBFs) and their encoding genes serve as signaling hubs for cold acclimation in Arabidopsis thaliana (Stockinger et al., 1997; Medina et al., 1999; Thomashow, 2010; Ding et al., 2019). When exposed to low temperatures, the transcription of CBFs is rapidly promoted by a group of cold-signal transducers, including the Ca2+/calmodulin-binding proteins, CALMODULIN-BINDING TRANSCRIPTION ACTIVATORs (CAMTAs) (Doherty et al., 2009; Kim et al., 2013; Kidokoro et al., 2017); the clock proteins, CIRCADIAN CLOCK-ASSOCIATED 1 (CCA1) and LATE-ELONGATED HYPOCOTYL (LHY) (Dong et al., 2011); and the brassinosteroid-responsive proteins, BRASSINAZOLE-RESISTANT 1 (BZR1) and CESTA (CES) (Eremina et al., 2016; Li et al., 2017). In addition, cold also enhances the stability or activity of CBF proteins. For example, cold facilitates the interaction between CBFs and BASIC TRANSCRIPTION FACTOR 3s (BTF3s), which promotes CBF stability (Ding et al., 2018), and cold triggers degradation of co-repressor, HISTONE DEACETYLASE 2C (HD2C), thereby allowing CBFs to activate their targets (Park et al., 2018b). Furthermore, cold reduces oxidized CBFs, which increases active CBF monomers (Lee et al., 2021). Low-temperature-induced CBFs, in turn, activate the expression of cold-regulated (COR) genes by binding to CRT/DREs in their promoters (Stockinger et al., 1997; Medina et al., 1999). Diverse arrays of cryoprotective proteins encoded by COR genes allow plants to overcome freezing stress (Gilmour et al., 1988; Hajela et al., 1990; Thomashow et al., 1997; Shinozaki et al., 2003; Shi et al., 2018; Ding et al., 2019).
In contrast to cold acclimation, vernalization, a floral-promotion process that occurs during winter, requires an extended cold period (Napp-Zinn, 1955; Chouard, 1960; Michaels and Amasino, 2000). This allows plants to synchronize the timing of flowering with favorable spring conditions. Vernalization in A. thaliana is mainly achieved by silencing the floral repressor gene, FLOWERING LOCUS C (FLC) (Michaels and Amasino, 1999; Sheldon et al., 1999; Sheldon et al., 2000; Michaels and Amasino, 2001). FLC encodes a MADS-box protein that represses the expression of floral activator genes, SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1) and FLOWERING LOCUS T (FT, encoding florigen), by directly binding to their promoter regions (Lee et al., 2000; Michaels et al., 2005; Helliwell et al., 2006). In Arabidopsis winter annuals, such as San Feliu-2, Löv-1, and Sweden (SW) ecotypes, flowering is prevented by the high expression of FLC before exposure to winter cold (Lee et al., 1993; Shindo et al., 2005; Park et al., 2018a). This is caused by the strong transcriptional activation of FLC by the FRIGIDA (FRI) supercomplex, which recruits general transcription factors and several chromatin modifiers (Michaels and Amasino, 1999; Sheldon et al., 1999; Johanson et al., 2000; Choi et al., 2011; Li et al., 2018). Prior to vernalization, FLC chromatin is highly enriched with active histone marks such as histone H3 acetylation and trimethylation of Lys4 or Lys36 at H3 (H3K4me3/H3K36me3) (Bastow et al., 2004; Yang et al., 2014). In contrast, prolonged cold exposure results in gradual deacetylation of FLC chromatin and concomitant removal of H3K4me3 and H3K36me3 from the FLC (Bastow et al., 2004; Yang et al., 2014; Nishio et al., 2016). Additionally, VP1/ABI3-LIKE 1 (VAL1) and VAL2 recruit Polycomb Repressive Complex 2 (PRC2) onto FLC chromatin, thereby accumulating the repressive histone mark, H3 Lys27 trimethylation (H3K27me3), in the nucleation region around the first exon and intron of FLC (Sung and Amasino, 2004b; Wood et al., 2006; De Lucia et al., 2008; Angel et al., 2011; Nishio et al., 2016; Qüesta et al., 2016; Yuan et al., 2016). Subsequently, upon returning to warm temperatures, H3K27me3 marks are spread over the entire FLC chromatin region by LIKE HETEROCHROMATIN PROTEIN 1 (LHP1), which ensures stable FLC suppression and renders plants competent to flower (Mylne et al., 2006; Sung et al., 2006; Yang et al., 2017).
Several long-term cold-induced factors have been shown to play crucial roles in the epigenetic silencing of FLC. VERNALIZATION INSENSITIVE 3 (VIN3) family genes, which are upregulated by prolonged cold, encode plant homeodomain (PHD) proteins that recognize H3K9me2 enriched in FLC chromatin during vernalization (Sung and Amasino, 2004b; Kim and Sung, 2013; Kim and Sung, 2017a). These proteins mediate the recruitment of PRC2 and the subsequent deposition of H3K27me3 at the FLC nucleation region (Sung and Amasino, 2004b; De Lucia et al., 2008; Kim and Sung, 2013). In addition, vernalization-induced long noncoding RNAs (lncRNAs) are involved in such histone modifications. COLDAIR and COLDWRAP, the two lncRNAs transcribed from the first intron and promoter region of FLC, respectively, are required for H3K27me3 deposition in response to long-term cold (Heo and Sung, 2011; Kim and Sung, 2017b). COLDAIR and COLDWRAP are also thought to affect the formation of the intragenic chromatin loop at the FLC, which may be a part of the FLC silencing mechanism (Kim and Sung, 2017b). Unlike COLDAIR and COLDWRAP, which are transcribed in the sense direction of FLC, another lncRNA, COOLAIR, is an antisense transcript expressed from the 3′-end of FLC (Swiezewski et al., 2009). The gradual accumulation of COOLAIR reaches a maximum within a few weeks of cold exposure, whereas COLDAIR and COLDWRAP show peaks at a later phase of vernalization (Csorba et al., 2014; Kim and Sung, 2017b). COOLAIR was reported to remove active histone marks from FLC chromatin (Liu et al., 2007; Csorba et al., 2014; Fang et al., 2020; Xu et al., 2021a; Zhu et al., 2021). Particularly, in summer annuals, phase-separated RNA-processing complexes favor co-transcriptional proximal polyadenylation of COOLAIR (Marquardt et al., 2014; Wang et al., 2014; Fang et al., 2019; Wu et al., 2020; Xu et al., 2021b). The COOLAIR-processing machinery exhibits transient and dynamic interactions with an H3K4 demethylation complex, leading to FLC suppression at warm temperatures (Liu et al., 2007; Fang et al., 2020; Xu et al., 2021a). COOLAIR is also likely to be involved in reducing H3K36me3 at the FLC during vernalization process (Csorba et al., 2014). COOLAIR was reported to promote the sequestration of the FRI complex from the FLC promoter by condensing it into phase-separated nuclear bodies (Zhu et al., 2021). This has been suggested to cause the inactivation of FLC, which is probably accompanied by the silencing of FLC chromatin through the removal of H3K36me3. However, a previous study has raised the issue that COOLAIR appears not to be necessary for vernalization (Helliwell et al., 2011; Luo et al., 2019).
Compared with the signaling pathway of cold acclimation, how a long-term cold signal is transduced to trigger the induction of VIN3 and lncRNAs is less well understood. It is marginally known that some chromatin modifiers, NAC WITH TRANSMEMBRANE MOTIF 1-LIKE 8 (NTL8), and CCA1/LHY act as positive regulators of VIN3 during the vernalization process (Kim et al., 2010; Jean Finnegan et al., 2011; Zhao et al., 2020; Kyung et al., 2022). However, little is known about the upstream regulators of COOLAIR required for cold-induction. Recent reports have shown that an NAC domain-containing protein, NTL8, and the WRKY transcription factor, WRKY63, can bind to the promoter of COOLAIR and activate its expression (Zhao et al., 2021; Hung et al., 2022). However, whether NTL8 and WRKY63 are necessary for the full extent of COOLAIR induction during vernalization has not been thoroughly addressed. In this study, we identified that CRT/DREs at the 3′-end of the FLC are required for the long-term cold response of COOLAIR. Additionally, we show that CBFs, which accumulate during the long-term winter cold, act as upstream regulators of COOLAIR during vernalization.
Results
A CRT/DRE-binding factor, CBF3, directly binds to CRT/DREs at the 3′-end of the FLC
The proximal promoter region of COOLAIR is highly conserved among FLC orthologs from A. thaliana relatives (Castaings et al., 2014). Thus, we assumed that the cis-element conferring a long-term cold response would exist within that block. A comparison of the region near the transcriptional start site (TSS) of COOLAIR revealed the conservation of two CRT/DREs among six FLC orthologs from five related species of the Brassicaceae family (Figure 1A). CRT/DRE, the core sequence of which is CCGAC, is a regulatory element that imparts cold- or drought-responsive gene expression (Baker et al., 1994). CBF1, 2, and 3 encode APETALA 2 (AP2) domain-containing proteins that can bind to CRT/DRE and are usually present in the promoters of cold- and drought-responsive genes (Stockinger et al., 1997; Medina et al., 1999). Consistent with this, the Arabidopsis cistrome database and the genome-wide chromatin immunoprecipitation sequencing (ChIP-seq) results from a previous study suggested that CBFs bind to the 3′-end sequence of FLC containing CRT/DREs (Figure 1—figure supplement 1; O’Malley et al., 2016; Song et al., 2021).
Figure 1. CBF3 directly binds to the CRT/DREs at the 3′-end of FLC.
(A) Comparison of sequences around the 3′-end of the FLC orthologs from Arabidopsis relatives. The upper graphic presents the gene structure of AtFLC. The black bars, black lines, and white bars indicate exons, introns, and untranslated regions (UTRs), respectively. The blue line presents the region used for sequence comparison among six orthologous genes from five plant species. In the sequence alignment below, the gray line indicates the 3′-UTR of FLC orthologs, the blue arrow indicates the transcriptional start site (TSS) of AtCOOLAIR, and the red lines indicate CRT/DREs. At, Arabidopsis thaliana; Cr, Capsella rubella; Al, Arabidopsis lyrata; Aa, Arabis alpina; Th, Thellungiella halophila. (B) EMSA using one of the CRT/DREs located at the AtCOOLAIR promoter, DRE1, as a probe. In the upper graphic showing AtFLC gene structure, CRT/DRE-like sequences are marked as blue arrows and labeled as a, b, c, and d for CRT/DRE-like sequences and as 1 and 2 for CRT/DRE sequences. For the competition assay, these CRT/DRE-like sequences and mutant forms of DRE1 and DRE2 were used as competitors. A 100-fold molar excess of unlabeled competitors was added. No protein (−) or maltose-binding protein (MBP) were used as controls. FP, free probe. (C) ChIP assay result showing the enrichment of CBF3-myc protein on the AtFLC locus. Samples of NV, 10V, 20V, and 40V plants of pSuper:CBF3-myc were collected at zeitgeber time (ZT) 4 in an SD cycle. The CBF3-chromatin complex was immunoprecipitated (IP) with anti-myc antibodies (blue bars), and mouse IgG (black bars) was used as a control. Positions of qPCR amplicons used for ChIP-qPCR analysis are illustrated as P1, P2, and P3 in the upper graphic. The blue arrow in the graphic denotes the position of CRT/DREs on the AtCOOLAIR gene. ChIP-qPCR results have been represented as mean ± SEM of the three biological replicates in the lower panel. Open circles and squares represent each data point. Relative enrichments of the IP/5% input were normalized to that of pTUB2. The blue shadings indicate cold periods. Asterisks indicate a significant difference between IgG and anti-myc ChIP-qPCR results at each vernalization time point (*, p < 0.05; **, p < 0.01; unpaired Student’s t-test).

Figure 1—figure supplement 1. CBF proteins directly bind to the sequence at the 3′-end of FLC in DAP-seq.

Figure 1—figure supplement 2. CBF3 binds to DRE1 and DRE2 on the COOLAIR promoter and DREc on the first exon of FLC.
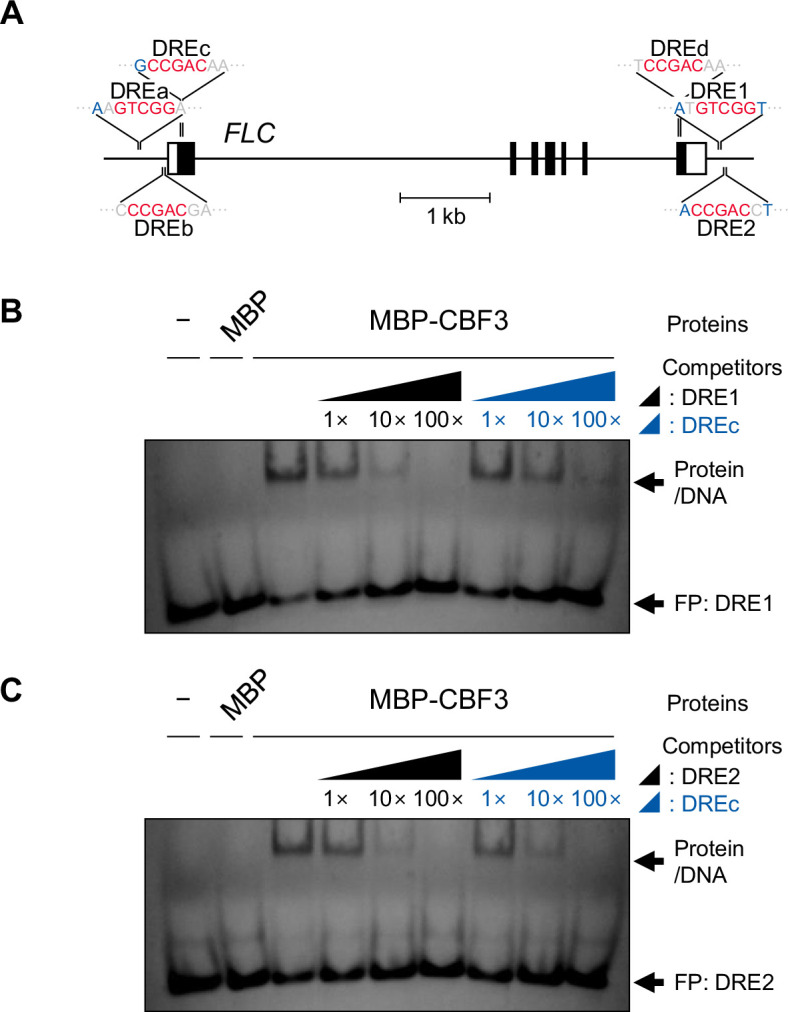
We performed an electrophoretic mobility shift assay (EMSA) to confirm this binding using probes harboring CRT/DREs (named DRE1 or 2) from the COOLAIR promoter. The mobility of these two Cy5-labeled probes was retarded by maltose-binding protein (MBP)-fused CBF3 (Figure 1B, Figure 1—figure supplement 2). The band shift was competed out by adding an excess amount of unlabeled DRE1 or 2 oligonucleotides. In contrast, competitors containing mutant forms of DRE1 or 2 (DRE1m or 2m, respectively) failed to compete (Figure 1B). We also tested the binding of other CRT/DRE-like sequences in the FLC locus. Two (DREa and b) were present in the FLC promoter, while the other two (DREc and d), were present in the first and last exons, respectively (Figure 1—figure supplement 2). Only DREc competed with the band shift caused by the CBF3-DRE1 interaction (Figure 1B, Figure 1—figure supplement 2). The presence or absence of bases that determine the binding affinity between CBFs and CRT/DRE could explain the differences in CBF3-binding patterns among CRT/DRE-like sequences at the FLC locus (Maruyama et al., 2004).
Subsequently, a chromatin immunoprecipitation (ChIP) assay using a CBF3-overexpressing transgenic plant, pSuper:CBF3-myc (Liu et al., 2017), was conducted to determine whether CBF3 is associated with the FLC region containing CRT/DREs in vivo. For ChIP assay to show binding affinity of CBF3 protein to the COOLAIR promoter during vernaliation, we used pSuper:CBF3-myc instead of pCBF3:CBF3-myc because endogenous CBF3 transcript level is changed during vernalization as shown afterward. Consistent with the in vitro results, CBF3-myc protein was enriched at the 3′-end of FLC and the first exon where DREc was located (Figure 1C). Without vernalization (NV), CBF3-myc was highly enriched at both the 5′- and 3′-ends of FLC (P1 and P3), indicating that CBF3 protein can activate FLC under warm conditions, as previously reported (Seo et al., 2009). However, enrichment in the P1 region rapidly disappeared during the vernalization period. In contrast, enrichment on P3 was maintained until 10 days of vernalization (10V) and was subsequently reduced. This is coincident with the expression pattern of COOLAIR, which declines during the late phase of vernalization (Csorba et al., 2014).
COOLAIR is one of the CBF regulons in Arabidopsis
CBF genes are rapidly and transiently induced upon exposure to cold (Medina et al., 1999). CBF regulons, which are CBF-targeted genes, are correspondingly up- or down-regulated after cold intervals of a few hours (Gilmour et al., 1998; Liu et al., 1998; Fowler and Thomashow, 2002). As COOLAIR contains CBF3-binding sites in its promoter (Figure 1), we analyzed whether COOLAIR shows CBF regulon-like expression. The expression of genes assigned to the CBF regulon is up- or down-regulated by CBF overexpression under warm conditions (Park et al., 2015). Similarly, we found that the total transcript level of COOLAIR increased in CBF3-overexpressing transgenic plants (pSuper:CBF3-myc) grown at room temperature (22 °C) compared to that in the wild type, Columbia-0 (Col-0) (Figure 2A). Such upregulation was mainly due to the type I.i or type II.i COOLAIR isoforms. As the levels of both proximal (type I) and distal (type II) variants of COOLAIR were higher in the CBF3 overexpressor than in the wild type, it is likely that COOLAIR transcription, instead of the 3′-processing events, is affected by CBF3 (Figure 2A; Liu et al., 2010; Marquardt et al., 2014). It has been reported that the targets of CBF1 or 2 are not entirely identical to those of CBF3, although CBF genes show high sequence similarity (Novillo et al., 2007). Thus, we compared the expression levels of all COOLAIR isoforms in transgenic plants overexpressing CBF1, 2, or 3 (Seo et al., 2009). Overall, all CBF-overexpressing plants showed increased levels of COOLAIR (Figure 2—figure supplement 1). However, each CBF-overexpressing plant showed a subtly different effect on COOLAIR expression. The CBF2 overexpressor showed the highest level of the proximal COOLAIR isoform, whereas the CBF3 overexpressor showed the highest level of distal COOLAIR. In contrast, the CBF1-overexpressing plants showed a slightly smaller increase in proximal COOLAIR levels and a negligible increase in distal COOLAIR, compared to those in the wild type, Wassilewskija-2 (Ws-2). Such results imply that the three CBFs regulate COOLAIR transcription subtly differently.
Figure 2. COOLAIR shows a similar expression pattern with CBF regulons.
(A) Expression levels of total COOLAIR and COOLAIR isoforms in the wild-type (Col-0) and CBF3-overexpressing plants (pSuper:CBF3-myc [CBF3ox]). The gene structures of FLC and COOLAIR variants are illustrated in the upper panel. The thin black arrows indicate the transcription start sites of FLC and COOLAIR. The thick black arrows indicate the exons of each COOLAIR isoform. The structures of proximal (I.i) and distal (II.i and II.ii) COOLAIR isoforms are shown. The primer sets used for total, proximal (I.i), and distal (II.i, II.ii) COOLAIRs are marked in Figure 2—figure supplement 1. Relative transcript levels of total COOLAIR and COOLAIR variants were normalized to that of UBC and have been represented as mean ± SEM of three biological replicates. Dots and squares represent each data point. Asterisks indicate a significant difference as compared to the wild type (*, p < 0.05; **, p < 0.01; ***, p < 0.001; unpaired Student’s t-test). (B) Expression dynamics of total COOLAIR after short-term (0, 0.5, 1, 1.5, 2, 3, 6, and 24 hr) cold treatment. Wild types (FRI Col) were subjected to 4 °C cold and harvested at each time point. Relative transcript levels of total COOLAIR to UBC were normalized to that of non-cold treated wild type. The values have been represented as mean ± SEM of three biological replicates. The blue shading indicates periods under cold treatment. Significant differences have been marked using different letters (a, b; p < 0.05; one-way ANOVA followed by Tukey’s post-hoc test). (C) Transcript levels of total COOLAIR and COOLAIR isoforms in wild type and cbfs-1 mutant before (−Cold) and after (+Cold) a day of 4 °C cold treatment. Relative levels of total COOLAIR and COOLAIR variants were normalized to that of PP2A. Values have been represented as mean ± SEM of three biological replicates. Dots and squares indicate each data point. Significant differences have been marked using different letters (a, b; p < 0.05; two-way ANOVA followed by Tukey’s post-hoc test).

Figure 2—figure supplement 1. Overexpression of CBF1, 2, or 3 causes an increase in COOLAIR level, but the effects on COOLAIR variants are subtly different.

Another characteristic of CBF regulons is that their expression is activated by a day of cold exposure (Park et al., 2015). Thus, we analyzed if COOLAIR is also induced by short-term cold exposure. We treated wild-type (FRI Col) plants with 0, 0.5, 1, 1.5, 2, 3, 6, and 24 hr of cold (4 °C), then measured the levels of total COOLAIR. The results showed that cold treatment for less than 3 hr failed to induce COOLAIR expression, although CBF transcripts reached a peak at 3 hr of cold exposure (Figures 2B and 3A; Gilmour et al., 1998). However, after 6 hr of cold treatment, when CBF3 protein levels peaked, COOLAIR was strongly induced (Figures 2B and 3D). Thus, short-term cold-triggered COOLAIR expression dynamics were similar to those of other CBF-targeted genes; that is, the expression was highly increased after CBF transcript levels reached a peak upon cold exposure (Gilmour et al., 1998; Fowler and Thomashow, 2002). To confirm whether CBFs are responsible for the rapid cold response of COOLAIR, we examined COOLAIR induction after a day of cold treatment in the wild type and cbfs-1 mutant, in which all three CBFs were knocked out (Figure 2C; Jia et al., 2016). All isoforms, as well as total COOLAIR, failed to be induced by short-term cold exposure in the cbfs mutant, although a drastic increase was observed in the wild type. Thus, these results strongly suggest that CBFs are required for COOLAIR induction, even under a short period of cold exposure.
Figure 3. Protein levels of CBFs increase during the vernalization process.
(A) Transcript levels of CBF1, 2, and 3 under short-term cold exposure. Wild-type plants were subjected to 0, 1, 3, 6, 12, and 24 hr of 4 °C cold treatment. Relative levels of CBF1, 2, and 3 were normalized to that of UBC. Values have been represented as mean ± SEM of three biological replicates. The blue shadings indicate cold periods. Significant differences have been marked using different letters (a–c; p < 0.05; one-way ANOVA followed by Tukey’s post-hoc test). (B) Transcript levels of CBF1, 2, and 3 after 20V and 40V. Wild-type plants were treated with 4 °C vernalization under an SD cycle and collected at ZT4. Relative levels of CBF1, 2, and 3 were normalized to that of UBC. Values have been represented as mean ± SEM of three biological replicates. The blue shadings denote periods under cold. Significant differences have been marked using different letters (a–c; p < 0.05; one-way ANOVA followed by Tukey’s post-hoc test). (C) Daily rhythms of CBF1, 2, and 3 transcript levels in NV or 40V plants. Col-0 plants grown under an SD cycle were collected every 4 hr between ZT0 and ZT24. Relative transcript levels of CBF1, 2, and 3 were normalized to that of PP2A. Values have been represented as mean ± SEM of three biological replicates. The white and black bars represent light and dark periods, respectively. Asterisks indicate a significant difference between NV and 40V (***, p < 0.001; two-way ANOVA). (D) Dynamics of CBF3 protein level under short-term cold exposure. The pCBF3:CBF3-myc transgenic plants were subjected to 0, 1, 3, 6, 12, and 24 hr of 4 °C cold treatment. CBF3 proteins were detected using anti-myc antibodies. Rubisco was considered the loading control. Numbers below each band indicate relative signal intensity compared to 6 hr. The mean values of two biological replicates are presented. (E) Increase of CBF3 protein level during the vernalization process. The pCBF3:CBF3-myc transgenic plants, subjected to 4 °C vernalization, were collected at ZT4 of the indicated time point. CBF3 proteins were detected using anti-myc antibodies. Rubisco was considered the loading control. Numbers below each band indicate relative signal intensity compared to 1V. The mean values of three biological replicates are presented.

Figure 3—figure supplement 1. Vernalization increases CBF3 protein levels with a minor effect on the daily rhythm.

CBFs accumulate during vernalization
We subsequently investigated whether CBFs are also responsible for vernalization-induced COOLAIR activation. It has been reported that COOLAIR is gradually upregulated as the cold period persists and peaks after 2–3 weeks of vernalization (Csorba et al., 2014). However, most studies on CBF expression have been performed within a few days since the function of CBFs has been analyzed only in the context of short-term cold (Gilmour et al., 1998; Liu et al., 1998; Medina et al., 1999; McKhann et al., 2008). Therefore, we investigated the expression patterns of CBFs before and after long-term cold exposure to determine the correlation between the expression of CBFs and COOLAIR during vernalization. As previously shown, the levels of all three CBFs peaked within 3 hr of cold exposure and then decreased rapidly (Figure 3A). However, CBF levels increased again as the cold period was prolonged (> 20V), suggesting that both short-term and vernalizing cold treatments upregulated CBF expression (Figure 3B).
Because CBFs exhibit rhythmic oscillations (Dong et al., 2011), we also investigated whether the rhythmic expression of CBFs is affected by vernalization. Wild-type (Col-0) plants were collected every 4 hr under a short-day photoperiod (SD, 8 hr light/16 hr dark) under both NV and 40V conditions. As shown in Figure 3C, although the rhythmic pattern of each CBF was variable, the overall transcript levels of all three CBFs were much higher at 40V than under NV. Consistent with the transcript level, the level of CBF3 protein in vernalized plants was higher than that in NV plants during all circadian cycles (Figure 3—figure supplement 1).
To verify whether the increased transcription of CBF3 led to protein accumulation, we measured the level of CBF3 at each time point of cold treatment using the pCBF3:CBF3-myc plant (Jia et al., 2016). Following the rapid and transient induction of CBF3 under short-term cold conditions (Figure 3A; Medina et al., 1999), CBF3-myc protein levels peaked within 6 hr of cold exposure and then decreased (Figure 3D). Thus, the peak of the CBF3 transcript and that of the CBF3 protein showed a gap of few hours. Notably, the peak of the CBF3 protein correlated with the time when COOLAIR was rapidly induced (Figures 2B and 3D). During vernalization, the level of CBF3-myc protein gradually declined until 10V, but then increased again (Figure 3E). Thus, the level at 40V is similar to that at 1V. When the plants were transferred to room temperature for 4 d after 40V (40VT4), the protein level rapidly decreased. These results suggest that CBFs are responsible for the progressive upregulation of COOLAIR, at least during the early phase of vernalization (Csorba et al., 2014).
CBFs are involved in vernalization-induced COOLAIR expression
To clarify whether CBFs are responsible for long-term cold-mediated COOLAIR induction, we quantified COOLAIR levels in wild type and cbfs mutant during vernalization. As shown in Figure 2B and C, the COOLAIR level was considerably increased by a day of low-temperature treatment in the wild type but quickly declined afterward (Figure 4A). The COOLAIR level increased again after 4V and reached a secondary peak at approximately 10V to 20V. It then decreased during the remaining vernalization period, as similar to previous results (Csorba et al., 2014). The expression dynamics of COOLAIR were similar to those of two well-known CBF targets, COR15A and RESPONSIVE TO DESICCATION 29A (RD29A), although their levels remained high until the end of the vernalization period (Figure 4—figure supplement 1). In contrast to the wild-type plants, the cbfs mutant showed severely reduced COOLAIR expression, as well as COR15A and RD29A levels, during all the period of vernalization (Figure 4A, Figure 4—figure supplement 1). Analysis of RNA sequencing (RNA-seq) data from previous studies also revealed that the levels of total COOLAIR, and proximal and distal COOLAIR variants were reduced in the cbfs mutant in Col-0 or SW ecotype background when exposed to either short-term (3 and 24 hr) or long-term (14V) cold, which supports our results (Figure 4—figure supplement 2; Park et al., 2018a; Song et al., 2021). All these data support that CBFs are necessary to fully induce COOLAIR in the early phase of vernalization.
Figure 4. CBFs are involved in vernalization-induced COOLAIR expression.
(A) Expression dynamics of total COOLAIR in the wild-type, cbfs-1, and FLCΔCOOLAIR-1 plants during vernalization. Relative transcript levels of total COOLAIR were normalized to that of PP2A. Values have been represented as mean ± SEM of three biological replicates. Asterisks indicate a significant difference compared to the wild-type (***, p < 0.001; two-way ANOVA followed by Tukey’s post-hoc test). (B) Transcript level of total COOLAIR in wild type (Col-0) and CBF3-overexpressing transgenic plant (pSuper:CBF3-myc [CBF3ox]) during vernalization. Relative levels of total COOLAIR were normalized to that of PP2A. Values have been represented as mean ± SEM of three biological replicates. Asterisks indicate a significant difference between the wild-type and CBF3 overexpressor (***, p < 0.001; two-way ANOVA). Blue shading denotes cold periods in (A) and (B). (C) Effect of first frost-mimicking treatment (8 hr of freezing [< 0 °C]) on the level of total COOLAIR in NV, 10V, and 20V wild type and cbfs-1. The upper panel shows a schematic of the experimental procedure. The non-frost treated (−F) wild type and cbfs-1 were collected at ZT22 after an 8 hr of dark treatment at 22 °C (NV) or 4 °C (10V and 20V). For the first frost treatment, wild type and cbfs-1 mutant were treated with an additional 8 hr of −1 °C (+F) under dark, and then the whole seedlings were collected at ZT22 for analysis. All the plants were grown under an SD cycle. The gray shadings denote dark periods. Total COOLAIR levels have been represented as mean ± SEM of three biological replicates in the lower panel. Dots and squares indicate each data point. Relative levels of total COOLAIR were normalized to that of PP2A. Asterisks indicate a significant difference (*, p < 0.05; ***, p < 0.001; two-way ANOVA followed by Tukey’s post-hoc test). ns, not significant (p ≥ 0.05).

Figure 4—figure supplement 1. CBF regulons show strongly reduced expression in cbfs mutants similar to COOLAIR.

Figure 4—figure supplement 2. Cold-triggered COOLAIR expression is impaired in cbfs mutants in Col-0 and Sweden-ecotype background.

Figure 4—figure supplement 3. Freezing temperature increases the levels of CBFs.

Consistent with the reduced COOLAIR levels in cbfs mutants, the CBF3 overexpressor, pSuper:CBF3-myc, showed a much higher COOLAIR level than the wild type (Col-0), throughout the vernalization period (Figure 4B). It is noteworthy that COOLAIR expression in pSuper:CBF3-myc was further upregulated by cold, especially during the early vernalization phase (10V), and then suppressed during the late phase. This result implies that both the transcriptional and post-transcriptional regulations of CBFs are involved in the long-term cold response of COOLAIR.
A recent study showed that the first seasonal frost (< 0 °C) during the winter season strongly induces COOLAIR expression (Zhao et al., 2021). Therefore, we tested whether CBFs are also required for the strong induction of COOLAIR triggered by freezing temperatures. Wild-type and cbfs plants were subjected to NV, 10V, and 20V, with or without an additional 8 hr of −1 °C freezing treatment. Irrespective of the pre-treatment with non-freezing cold, the freezing temperature increased the levels of both COOLAIR and CBF in the wild type (Figure 4C, Figure 4—figure supplement 3). However, the cbfs mutants showed a much smaller increase in the COOLAIR level than the wild type when exposed to 8 hr of sub-zero cold after 10V and 20V (Figure 4C). In particular, COOLAIR levels were not elevated by freezing treatment in NV cbfs. Thus, CBFs seem responsible for both the gradual increase of COOLAIR during vernalization and the strong COOLAIR induction triggered by freezing temperatures.
CRT/DREs at the 3′-end of FLC are necessary for CBFs-mediated induction of COOLAIR during vernalization
Since CBF3 could bind to the CRT/DREs in the first exon (DREc) and the 3′-end of FLC (DRE1 and 2) (Figure 1B and C), we investigated whether CRT/DRE is responsible for the CBF-mediated long-term cold response of COOLAIR. To address this, we performed an A. thaliana protoplast transfection assay using the 35S:CBF3-HA effector construct and the pCOOLAIRDRE:LUC reporter construct, in which the 1 kb wild-type COOLAIR promoter was fused to the luciferase reporter gene. The luciferase activity assay showed that CBF3-HA protein activated the transcription of the COOLAIR promoter (Figure 5A). In contrast, CBF3-HA failed to increase luciferase activity when the mutant version of the COOLAIR promoter with mutations in the DRE1 and 2 sequences (pCOOLAIRDREm:LUC) was used as a reporter. Additionally, co-transfection of the 35S:CBF3-HA effector and pFLC:LUC reporter construct, which harbors the 1 kb sequence with the promoter and first exon of FLC, did not show increased luciferase activity. These results strongly indicate that CBF3 can activate COOLAIR transcription through DRE1 and 2 located in the COOLAIR promoter, but not via the first exon of FLC.
Figure 5. COOLAIR promoter region containing CRT/DREs is necessary for the COOLAIR induction by vernalization.
(A) Arabidopsis protoplast transfection assay showing that CBF3 activates COOLAIR promoter with wild-type CRT/DRE (DRE) but fails to activate the one with mutant CRT/DRE (DREm). A schematic of the reporter constructs used for the luciferase assay is presented in the upper panel. pFLC:LUC contains 1 kb of the promoter, the 5′-UTR, and the first exon of FLC. The blue line in the pFLC:LUC graphic indicates the location of DREc. Wild-type and mutant forms of pCOOLAIR:LUC include 1 kb of the COOLAIR promoter with the 3′-UTR and the last exon of FLC. The blue and red lines mark the positions of the wild-type (DRE) and mutant (DREm) forms of DRE1 and DRE2, respectively. Each reporter construct was co-transfected into Arabidopsis protoplast together with the 35S:CBF3-HA effector construct. In parallel, 35S-NOS plasmid was transfected as a control. The result is shown below. Relative luciferase activities were normalized to that of the 35S-NOS control. Values have been represented as mean ± SEM of three biological replicates. Dots and squares represent each data point. (B) Transcript levels of proximal (I.i) and distal (II.i and II.ii) COOLAIR isoforms in wild-type, FLCΔCOOLAIR-1, FLCΔCOOLAIR-3, and FLCΔCOOLAIR-4 plants before and after 14 days of vernalization. FLCΔCOOLAIR-1 and FLCΔCOOLAIR-3 have a 324- and 685 bp deletion of the COOLAIR promoter region, respectively, where DRE1 and 2 are located. FLCΔCOOLAIR-4 has a deletion in the 301 bp COOLAIR promoter region outside of DRE1 and 2 location. The positions of the deleted region are marked in red, orange, and green lines with reversed triangle in the upper graphic. Blue arrows denote CRT/DRE-like sequences. Black and gray bars denote exons, thin lines denote introns, and white bars denote UTRs of FLC and its neighboring gene (AT5G10130). Results of RT-PCR analysis are shown below. UBC was used as a quantitative control. (C) Transcript levels of proximal (I.i) COOLAIR isoform in wild-type, cbfs-1, and FLCΔCOOLAIR-1 plants during vernalization. The position of the deleted region is marked in red lines with a reversed triangle in the upper graphic. Black bars denote exons, thin lines denote introns, and white bars denote UTRs of FLC. The thin black arrow below the gene structure indicates the transcriptional start site of COOLAIR. The thick black arrow below denotes exons of the type I.i COOLAIR variant. The position of the amplicon used for the qPCR analysis is marked with blue arrows. The result of qPCR analysis is presented in the lower panel. Relative transcript levels were normalized to that of PP2A. Values have been represented as mean ± SEM of three biological replicates. The blue shading indicates periods under cold treatment. Asterisks indicate a significant difference, as compared to NV (*, p < 0.05; **, p < 0.01; ***, p < 0.001; two-way ANOVA followed by Tukey’s post-hoc test).

Figure 5—figure supplement 1. Genomic sequences near the deleted regions in two COOLAIR promoter deletion lines, FLCΔCOOLAIR-3 and FLCΔCOOLAIR-4.

Figure 5—figure supplement 2. Levels of CAS and unspliced FLC are not significantly altered in the FLCΔCOOLAIR mutant lines.

Figure 5—figure supplement 3. Proximal COOLAIR level is not significantly increased by CBF3 overexpression in the FLCΔCOOLAIR mutant lacking CRT/DREs.

To further determine whether the 3′-end sequence of FLC is required for cold-induction of COOLAIR expression, we measured COOLAIR expression before and after vernalization in the FLCΔCOOLAIR mutant (FLCΔCOOLAIR-1), that lacks a 324 bp portion of the COOLAIR promoter region (Luo et al., 2019). Reverse transcription PCR (RT-PCR) analysis showed that neither proximal nor distal COOLAIR variants were detected in this mutant, and the levels did not increase after 14 day cold exposure (Figure 5B). Using the CRISPR-Cas9 system, we generated additional FLCΔCOOLAIR mutant lines, FLCΔCOOLAIR-3, and FLCΔCOOLAIR-4 (Figure 5B, Figure 5—figure supplement 1). FLCΔCOOLAIR-3 has a 685 bp deletion in the COOLAIR promoter region where both CRT/DREs are located. As expected, this mutant failed to express COOLAIR even after 14V, which is similar to the FLCΔCOOLAIR-1 mutant. In contrast, the FLCΔCOOLAIR-4 mutant, which bears a 301 bp deletion in the distal COOLAIR promoter region (upstream of CRT/DREs) and a 13 bp sequence replacement downstream of CRT/DREs, showed a normal vernalization-induced increase in all COOLAIR variants, similar to the wild type. This strongly suggests that CRT/DREs located at the COOLAIR promoter are critical for CBF-mediated COOLAIR induction during vernalization. Notably, the low-abundance convergent antisense transcripts (CAS; Zhao et al., 2021) were similarly reduced by 14V in all genotypes, that is, the wild type and FLCΔCOOLAIR mutants (Figure 5B, Figure 5—figure supplement 2), suggesting their regulation is independent of COOLAIR.
Although the common COOLAIR TSS was eliminated in FLCΔCOOLAIR-1 (Figure 5B), the proximal COOLAIR isoform (type I.i) was still slightly detected by quantitative PCR (qPCR), probably due to alternative TSSs (Figure 5C). However, the expression of the proximal COOLAIR variant in the FLCΔCOOLAIR-1 mutant was not induced throughout the vernalization period, while the wild type showed type I.i COOLAIR peaks at 1V and 20V (Figure 5C). Furthermore, CBF3 overexpression did not significantly increase type I.i COOLAIR levels in the FLCΔCOOLAIR-1 mutant, while it caused a strong increase in proximal COOLAIR in the wild-type background (Col-0) (Figure 5—figure supplement 3). Taken together, our results strongly suggest that the COOLAIR promoter region containing the two CRT/DREs is necessary for the long-term cold response of COOLAIR.
CBFs-mediated COOLAIR induction during vernalization is not required for FLC silencing
Finally, we investigated the role of COOLAIR induced by CBFs in the vernalization-triggered FLC silencing process. COOLAIR was reported to facilitate the removal of H3K36me3 from FLC chromatin during vernalization, whereas the other lncRNAs, COLDAIR and COLDWRAP, are required for H3K27me3 deposition (Csorba et al., 2014; Berry and Dean, 2015; Kim and Sung, 2017b; Zhu et al., 2021). Therefore, we compared the enrichment of H3K4me3, H3K36me3, and H3K27me3 on FLC chromatin in non-vernalized or vernalized wild-type and cbfs plants. As reported (Bastow et al., 2004; Sung and Amasino, 2004b; Yang et al., 2014), the enrichments of H3K4me3 and H3K36me3 were reduced, but that of H3K27me3 was increased in FLC chromatin at 40V in the wild type (Figure 6A–C). However, although COOLAIR levels were significantly reduced in the cbfs mutant (Figure 4A), vernalization-mediated reductions in H3K4me3 and H3K36me3 levels and an increase in H3K27me3 levels in the cbfs mutant were comparable to those in the wild type (Figure 6A–C). This result suggests that CBF-mediated COOLAIR induction is not required for vernalization-induced epigenetic changes on FLC chromatin.
Figure 6. Vernalization-triggered epigenetic silencing of FLC is not affected by CBFs-mediated COOLAIR induction.

(A–C) Enrichments of H3K36me3 (A), H3K4me3 (B), and H3K27me3 (C) on the FLC locus in NV or 40 V wild-type and cbfs-1 plants. The whole seedlings were collected at ZT4 under an SD cycle. Modified histones were immunoprecipitated with anti-H3K36me3, anti-H3K4me3, or anti-H3K27me3 antibodies. H3K36me3, H3K4me3, and H3K27me3 enrichments of the IP/5% input were normalized to those of ACT7 or FUS3. Relative enrichments have been represented as mean ± SEM of three biological replicates. One-way ANOVA and Tukey’s post-hoc test were performed on the ChIP results obtained using two primer pairs corresponding to the FLC nucleation region (Yang et al., 2017). Asterisks indicate a significant difference (*, p < 0.05). ns, not significant (P ≥ 0.05). (D) Transcript levels of FLC mRNA in wild-type, cbfs-1 and FLCΔCOOLAIR-1 plants during and after vernalization. Blue shading denotes cold periods. Relative levels were normalized to that of PP2A. Values have been represented as mean ± SEM of three biological replicates. ns, not significant between wild type and mutants, as assessed using two-way ANOVA followed by Tukey’s post-hoc test (p ≥ 0.05). (E) Transcript levels of FLC mRNA in wild type, FLCΔCOOLAIR-1, FLCΔCOOLAIR-3, and FLCΔCOOLAIR-4 during and after vernalization. Blue shading denotes cold periods. Relative levels were normalized to that of PP2A, and then normalized to NV of each genotype. Values have been represented as mean ± SEM of three biological replicates. ns, not significant between wild type and mutants, as assessed using two-way ANOVA followed by Tukey’s post-hoc test (p ≥ 0.05). (F) Effect of the first frost-mimicking treatment (8 hr freezing) on the FLC mRNA level in wild type and cbfs-1 after vernalization. NV, 10V, 20V, NV +F, 10V+F, and 20V+F plants were subjected to the treatments described in Figure 4C. Relative levels were normalized to that of PP2A. Values have been represented as mean ± SEM of three biological replicates. Dots and squares represent each data point. Asterisks indicate a significant difference (*, p < 0.05; **, p < 0.01; two-way ANOVA followed by Tukey’s post-hoc test). ns, not significant (p ≥ 0.05).
Consistent with the epigenetic changes, the FLC level in the cbfs mutant was gradually suppressed during the vernalization process, and the suppression was maintained after 40VT4 similar to that observed in the wild type (Figure 6D). Such result is consistent with the RNA-seq data previously reported using cbfs mutants in an SW ecotype (Park et al., 2018a). RNA-seq data analysis showed that the cbfs mutant in the SW background exhibited reduced FLC levels after 14V, similar to the wild type (Figure 4—figure supplement 2).
Because our results indicate that COOLAIR induction is not required for FLC silencing and the function of COOLAIR in vernalization-induced FLC silencing is still debated (Helliwell et al., 2011; Csorba et al., 2014; Zhao et al., 2021; Zhu et al., 2021), we analyzed whether the loss-of-function mutations FLCΔCOOLAIR-1 and FLCΔCOOLAIR-3 result in any defect in the vernalization response. In contrast to the transgenic lines used in previous reports, FLCΔCOOLAIR-1 and FLCΔCOOLAIR-3, which have small deletions in the COOLAIR promoter region, are unlikely to cause any unexpected effects on FLC chromatin. FLCΔCOOLAIR-4, which has a deletion outside of the CRT/DRE region, exhibited a gradual decrease and eventual silencing of FLC by long-term cold, similar to the wild type (Figure 6E). Interestingly, both FLCΔCOOLAIR-1 and FLCΔCOOLAIR-3 mutants exhibited similar decreases and silencing of FLC by vernalization (Figures 5B, 6D and E). These results show that FLC is normally silenced by vernalization regardless of COOLAIR induction by cold.
Consistent with this, the cbfs mutant, compared to the wild type, did not show any difference in the reduction of FLC levels upon exposure to a freezing cold for 8 hr after NV, 10V, and 20V (Figure 6F). However, the same freezing cold treatment caused the failure of strong induction of COOLAIR in the cbfs (Figure 4C). These results also support that CBF-mediated COOLAIR induction is not required for FLC silencing by vernalization.
As expected from the FLC levels, vernalization-mediated promotion of flowering was comparable in the wild type and cbfs (Figure 7A and B), indicating that cbfs exhibits a normal vernalization response. Consistently, the flowering time of FLCΔCOOLAIR-1 and FLCΔCOOLAIR-3 was accelerated by vernalization, similar to that of the wild type (Figure 7A, C and D). These results show that FLC silencing occurs regardless of whether COOLAIR is induced during a cold exposure that results in vernalization. In summary, our data strongly support that CBFs, upregulated by long-term cold exposure, induce COOLAIR expression (Figure 8); however, cold-mediated COOLAIR induction is not required for FLC silencing during vernalization.
Figure 7. CBF-mediated COOLAIR induction under long-term cold is not absolutely necessary for vernalization response.

(A) Photographs of NV and 40 V wild type, cbfs-1, and FLCΔCOOLAIR-1. NV plants were grown at 22 °C under LD and 40V plants were grown at 22 °C under LD after being treated for 40V under SD. The photos were taken when the NV plants started to bolt and the 40V plants showed the first open flower. (B) Flowering times of NV and vernalized (10V–40V) wild-type and cbfs-1 plants. Flowering time was measured in terms of the number of rosette leaves produced when bolting. Plants were grown under LD after vernalization treatment. Bars and error bars represent the mean ± standard deviation (SD) of three biological replicates. Each dot and square indicates individual flowering time. Significant differences have been marked using different letters (a–e; two-way ANOVA followed by Tukey’s post-hoc test; p < 0.05). (C) Flowering times of non-vernalized (NV) and vernalized (10V–40V) wild-type and FLCΔCOOLAIR-1 plants. Flowering time was measured in terms of the number of rosette leaves produced when bolting. Plants were grown under LD after vernalization. Bars and error bars indicate the mean ± SD. Each dot and square represents the individual flowering time. Significant differences have been indicated using different letters (a–e; two-way ANOVA followed by Tukey’s post-hoc test; P < 0.05). (D) Flowering times of non-vernalized (NV) and vernalized (14V–42V) wild-type, FLCΔCOOLAIR-1, FLCΔCOOLAIR-3, and FLCΔCOOLAIR-4 plants. Flowering time was measured in terms of the number of rosette leaves produced when bolting. Plants were grown under LD after vernalization. Bars and error bars indicate the mean ± SD. Each dot, square, triangle, and polygon represents the individual flowering time. Significant differences have been indicated using different letters (a–g; two-way ANOVA followed by Tukey’s post-hoc test; p < 0.05).
Figure 8. Schematic model describing the mechanism of CBFs-mediated COOLAIR expression during the early phase of vernalization.

During the early phase of vernalization, increased CBF upregulates COOLAIR expression by binding to CRT/DREs at the 3′-end of FLC. In the late phase of vernalization, owing to the silencing of FLC chromatin, CBF proteins are released from the CRT/DREs in the COOLAIR promoter, which leads to a reduction in COOLAIR levels.
Discussion
Winter is challenging for sessile plants: cold weather makes reproduction difficult, and frost threatens survival. Cold acclimation and vernalization, the two types of low-temperature memories, allow plants to overcome this harsh season by making them more viable during winter or flowering in the subsequent warm spring (Thomashow, 1999; Michaels and Amasino, 2000). Although it is well understood how plants recognize and remember short-term cold exposure to increase frost hardiness, there is still much to know about the long-term cold sensing mechanism that establishes the ability to flower.
FLC orthologs from a range of A. thaliana relatives express the vernalization-induced antisense lncRNA, COOLAIR (Castaings et al., 2014). As the proximal sequence blocks within the COOLAIR promoters are conserved, they are likely targets of cold sensor modules. This study showed that CBF proteins act as cold sensors binding to the CRT/DREs in the conserved promoter region, thereby activating the transcription of COOLAIR. Based on the data presented here, we propose a working model (Figure 8) in which a prolonged low-temperature environment gradually increases the levels of CBF proteins, which activate COOLAIR transcription in the early phase of vernalization by binding to CRT/DREs at the 3′-end of FLC. However, as cold period extended, CBF proteins are excluded from the COOLAIR promoter, probably because silencing of FLC chromatin occurs regardless of CBFs level. This may account for a decrease in COOLAIR levels during the later phase of vernalization. Consistent with this, a previous report showed a continuous increase in the COOLAIR level throughout the vernalization period in the vin3 mutant, which has a defect in the maintenance of FLC silencing (Swiezewski et al., 2009). Thus, VIN3-dependent Polycomb silencing is at least partially involved in the release of CBF proteins from the COOLAIR promoter.
Although CBFs are upregulated by cold to trigger COOLAIR expression, CBF-mediated COOLAIR induction during vernalization is not solely due to increased CBF levels because the CBF3 overexpression lines still showed COOLAIR induction by vernalization (Figure 4B). This may indicate that additional factors participate in the regulation of CBFs function. As aforementioned, it has previously been reported that low temperatures cause functional activation of CBFs through the degradation of HD2C which maintains an epigenetically inactive state of downstream targets at warm temperatures (Park et al., 2018b). Moreover, cold triggers the monomerization or stabilization of CBF proteins, thereby promoting their functions (Ding et al., 2018; Lee et al., 2021). Similarly, such an additional regulatory mechanism of CBF function may be enhanced by long-term winter cold to fine-tune COOLAIR expression.
In addition, other thermosensors are likely to facilitate COOLAIR induction by vernalization. Recently, it was suggested that NTL8, an NAC domain-containing transcription factor, acts as an upstream regulator of COOLAIR (Zhao et al., 2021). NTL8 also binds to the COOLAIR promoter region, which is not far from CBF-binding sites (O’Malley et al., 2016; Zhao et al., 2021). COOLAIR is highly expressed in both dominant mutant alleles, ntl8-Ds, and NTL8-overexpressing transgenic plants at warm temperatures. Thus, it has been proposed that slow accumulation of the NTL8 protein, due to reduced dilution during slower growth at low temperatures, causes COOLAIR induction during vernalization. In addition, a WRKY transcription factor, WRKY63, has been suggested as another inducer of COOLAIR transcription during vernalization (Hung et al., 2022). WRKY63 is enriched in the COOLAIR promoter region, and COOLAIR expression is reduced in the wrky63 mutant, suggesting that WRKY63 functions as a COOLAIR activator. Unlike NTL8, both transcript and protein levels of WRKY63 were upregulated by vernalization, which may lead to an increase in COOLAIR expression. Thus, it would be worthwhile to elucidate whether NTL8, WRKY63, and CBFs synergistically activate COOLAIR transcription during vernalization. The reason why vernalization-triggered COOLAIR induction requires more than one thermosensor is unclear. Presumably, multiple thermosensory modules ensure precise and robust COOLAIR expression during winter, when the temperature fluctuates.
Transcription of COOLAIR has been proposed to govern the FLC chromatin environment in both warm and cold temperatures (Csorba et al., 2014; Fang et al., 2020; Xu et al., 2021b). Previous studies have inferred that COOLAIR is required to remove H3K36me3 from FLC chromatin, especially under low-temperature conditions. This argument is supported by the delayed H3K36me3 removal in vernalized FLC-TEX transgenic plants, where the COOLAIR promoter is substituted by the rbcs3B terminator sequence (Csorba et al., 2014; Marquardt et al., 2014; Wang et al., 2014). In addition, COOLAIR is reported to promote condensation of FRI upon cold exposure (Zhu et al., 2021). Thus, FRI is sequestered from the FLC promoter as a nuclear condensate, which causes the transcriptional repression of FLC. Therefore, it has been suggested that vernalization-induced COOLAIR prevents the activation of FLC by the FRI complex and causes epigenetic silencing of FLC chromatin through a decrease of H3K36me3 during the vernalization process.
However, there is some controversy as to whether COOLAIR is required for vernalization and the associated FLC silencing. In contrast to the FLC-TEX lines, some T-DNA insertion lines which lack COOLAIR induction by long-term cold exposure showed a normal vernalization response (Helliwell et al., 2011). Likewise, the results from this study show that COOLAIR transcription is not required for the epigenetic silencing of FLC. The cbfs mutant, which exhibits severely reduced COOLAIR induction, shows relatively normal vernalization responses such that flowering time is fairly well accelerated and active histone marks, H3K36me3 and H3K4me3, are reduced in FLC chromatin while the repressive histone mark, H3K27me3, is increased in FLC chromatin similar to the wild type (Figures 6 and 7). Consistently, FLCΔCOOLAIR-1, and FLCΔCOOLAIR-3 mutants, which have almost undetectable levels of COOLAIR, exhibit both FLC suppression and flowering acceleration by vernalization similar to that seen in the wild type (Figures 6 and 7). Moreover, the freezing treatment mimicking the first frost did not affect FLC suppression in the cbfs, although the mutant showed defects in CBF-mediated COOLAIR induction (Figures 4C and 6F). It is noteworthy that our finding that vernalization and the associated FLC silencing do not require COOLAIR expression was obtained using cbfs and FLCΔCOOLAIR mutant lines rather than the transgenic lines used in previous studies. Thus, we can exclude the possibility that transgenes somehow provide cryptic promoters or cause unexpected epigenetic effects.
COOLAIR is highly conserved among Arabidopsis relatives (Castaings et al., 2014); thus, our finding that COOLAIR is not required for vernalization-induced FLC silencing is surprising. There are several caveats in the studies of COOLAIR function. First, different experimental schemes may affect the vernalization response differently. A recent study of plants grown in fluctuating temperatures designed to mimic natural environments may indicate a role for COOLAIR in vernalization-mediated FLC repression (Zhao et al., 2021), and perhaps fluctuating temperatures that mimic field conditions may have different effects on the vernalization response of cbfs or FLCΔCOOLAIR mutants as well. However, the experiments in most studies that claimed a role for COOLAIR in vernalization were performed in laboratory conditions with controlled temperatures as in this study. If indeed COOLAIR does have a role in vernalization in specific environmental conditions, the lack of a role for COOLAIR in vernalization in other conditions such as those in this study may be due to functional redundancy between COOLAIR and other FLC repressors such that other repressors, such as COLDAIR, obscure the effect of COOLAIR on FLC silencing, as has been proposed before (Castaings et al., 2014). This issue should be addressed in future studies.
Although short-term and long-term cold memories elicit different developmental responses, both are triggered by the same physical environment. Therefore, it is not surprising that a common signaling network governs both processes. CBF genes have been highlighted for their roles in cold acclimation (Gilmour et al., 1998; Liu et al., 1998; Jia et al., 2016; Zhao et al., 2016). CBFs rapidly accumulate upon short-term cold exposure, thereby activating the transcription of cold-responsive genes, such as COR15A and RD29A (Stockinger et al., 1997; Liu et al., 1998). Interestingly, CBFs are also highly accumulated during vernalization and mediate long-term cold-induced COOLAIR expression. Hence, our results indicate that the cold signaling pathways that establish the short-term and long-term cold responses are not sharply distinguishable, as suggested previously (Sung and Amasino, 2004a; Seo et al., 2009).
Materials and methods
Plant materials and treatments
Arabidopsis thaliana Columbia-0 (Col-0), Wassilewskija-2 (Ws-2), and Col FRISf2 (FRI Col) were used as the wild types in this study. “Wild type” in this paper mainly refers to FRI Col unless otherwise specified. The FRI Col (Lee et al., 1994), cbfs-1, pCBF3:CBF3-myc (Jia et al., 2016), pSuper:CBF3-myc (Liu et al., 2017), FLCΔCOOLAIR-1 (Luo et al., 2019), 35S:CBF1, 35S:CBF2, and 35S:CBF3 Gilmour et al., 2004 have been previously described. FRI cbfs-1 was generated by crossing the FRI Col with cbfs-1 stated above. FRI FLCΔCOOLAIR-1 was generated by crossing FRI Col with FLCΔCOOLAIR-1 in the Col-0 background, as described above. pSuper:CBF3-myc FLCΔCOOLAIR-1 was generated by crossing pSuper:CBF3-myc with FLCΔCOOLAIR-1.
For all experiments using plant materials, seeds were sown on half-strength Murashige and Skoog (MS) medium (Duchefa, Haarlem, Netherlands) containing 1% sucrose and 1% plant agar (Duchefa) and stratified at 4 °C for 3 days. Plants were germinated and grown at 22 °C under a short-day (SD) cycle (8 hr light/16 hr dark) with cool white fluorescent lights (100 μmol·m−2·s−1). For the vernalization treatment, the plants were grown at 22 °C for the prescribed period and then transferred to a 4 °C growth chamber under an SD cycle. To adjust the developmental stage, the plants vernalized for 40 days (40V), 30 days (30V), 20 days (20V), 10 days (10V), 4 days (4V), and 1 day (1V) were transferred to 4 °C after being grown at 22 °C for 10, 12, 14, 15, 15, and 16 days, respectively. Non-vernalized (NV) plants were grown at 22 °C for 16 days. For 0.5, 1, 1.5, 2, 3, 6, and 24 hr of cold treatment, the plants were grown at 22 °C for 16 days before the 4 °C treatment. For the freezing treatment, NV, 10V, and 20V plants were transferred to −1 °C under dark condition for 8 hr (F). To measure flowering time of vernalized plants, plants were transferred to 22 °C under a long-day (LD) cycle (16 hr light/8 hr dark) directly after the vernalization treatment.
Generation of FLCΔCOOLAIR lines by CRISPR-Cas9
The construction of FLCΔCOOLAIR-3 and FLCΔCOOLAIR-4 are as previously described (Luo et al., 2019). Briefly, two pairs of sgRNAs were designed for the deletion of COOLAIR promoter regions (5′-CTTCACAGTGAAGAAGCCTA-3′ and 5′-AAATGCACTCTTACGTAACG-3′ for FLCΔCOOLAIR-3; 5′-TTATCCTAAACGCGTATGGT-3′ and 5′-CGTAGTTCCGTCATCCATGA-3′ for FLCΔCOOLAIR-4), and sgRNA-Cas9 cassettes were introduced into Arabidopsis by floral dipping. Homozygous deletion mutants were screened in T2 generation, and sgRNA-Cas9 free mutants were further isolated in T3 generation.
Plasmid construction
To generate the 35S:CBF3-HA construct, the entire coding sequence of CBF3 was amplified using PCR and cloned into the HBT-HA-NOS plasmid (Yoo et al., 2007). For pCOOLAIRDRE:LUC and pCOOLAIRDREm:LUC construction, the 1 kb sequence containing the COOLAIR promoter, the 3′-untranslated region (UTR), and the last exon of FLC was amplified by PCR and cloned into the LUC-NOS plasmid (Hwang and Sheen, 2001). To generate pCOOLAIRDREm:LUC, two CRT/DREs in the COOLAIR promoter, 5′-AGGTCGGT-3′ and 5′-ACCGACAT-3′, were replaced with 5′-CGAGGTGT-3′ and 5′-TGAACCCA-3′, respectively. For pFLC:LUC construction, the 1 kb sequence containing the promoter, 5′-UTR, and the first exon of FLC was amplified by PCR and was cloned into the LUC-NOS plasmid.
EMSA
Maltose-binding protein (MBP) and MBP-CBF3 recombinant fusion proteins were induced by 500 μM isopropyl β-D-thiogalactoside (IPTG) in the Esecherichia coli BL21 (DE3) strain. Cell extracts were isolated with a buffer containing 20 mM Tris-HCl (pH 7.4), 1 mM EDTA, 200 mM NaCl, 10% glycerol, 1 mM DTT, and 1 mM PMSF. Proteins were purified from cell extracts using MBPTrap HP column (GE Healthcare, Chicago, IL, USA) with ÄKTA FPLC system (Amersham Biosciences, Amersham, UK). The Cy5-labeled probes and unlabeled competitors were generated by annealing 25 bp-length oligonucleotides. Electrophoretic mobility shift assay (EMSA) was performed as previously described with a few modifications (Seo et al., 2012). For each EMSA reaction, 5 μM of protein and 100 nM of Cy5-labeled probe were incubated at room temperature in a binding buffer containing 10 mM Tris-HCl (pH 7.5), 1 mM EDTA, 50 mM NaCl, 5% glycerol, and 5 mM DTT. For the competition assay, the reaction mixtures were incubated in the presence of 100 fold molar excess of each competitor. The reaction mixtures were electrophoresed at 100 V after incubation. The Cy5 signals were detected using WSE-6200H LuminoGraph II (ATTO, Amherst, NY, USA).
ChIP assay
Approximately 4 g of seedlings grown on MS plates was collected and cross-linked using 1% (v/v) formaldehyde. Nuclei were isolated from seedlings using a buffer containing 20 mM PIPES-KOH (pH 7.6), 1 M hexylene glycol, 10 mM MgCl2, 0.1 mM EGTA, 15 mM NaCl, 60 mM KCl, 0.5% Triton X-100, 5 mM β-mercaptoethanol, and 1×cOmplete EDTA-free Protease Inhibitor Cocktail (Roche, Basel, Switzerland) as described previously with a few modifications (Shu et al., 2014). Isolated nuclei were lysed with a buffer containing 50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1% (v/v) Triton X-100, and 1% SDS (w/v), and were subsequently sonicated using a Digital Sonifier (Branson, Danbury, CT, USA). Sheared chromatin was diluted with a buffer containing 50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1% (v/v) Triton X-100, and 1 mM EDTA. Chromatin immunoprecipitation (ChIP) was performed by incubating sheared chromatin with Protein G Sepharose 4 Fast Flow beads (GE Healthcare) and antibodies. Anti-myc (MBL, Woburn, MA, USA; #M192-3) and normal mouse IgG1 (Santa Cruz Biotechnology, Santa Cruz, CA, USA; #sc-3877) were used to detect CBF3 protein enrichment at the FLC locus, and anti-H3K36me3 (Abcam, Cambridge, UK; #Ab9050), anti-H3K4me3 (Millipore, Bedford, MA, USA; #07–473), and anti-H3K27me3 (Millipore; #07–449) were used to detect histone modifications on the FLC chromatin. DNA was extracted with phenol:chloroform:isoamyl alcohol (25:24:1, v/v) or Chelex 100 Resin according to the manufacturer’s instruction.
Gene expression analysis
Total RNA was isolated from –100 mg of seedlings grown on MS agar plate using TRI Reagent (Sigma-Aldrich, St Louis, MO, USA) or TS Plant Mini Kit (Taeshin Bio Science, Gyeonggi-do, Korea). cDNA was generated using 4 μg of total RNA, 5 units of recombinant DNase I (TaKaRa, Kyoto, Japan), 200 units of RevertAid reverse transcriptase (Thermo Scientific, Waltham, MA, USA), and buffer containing 0.25 mM dNTP and 0.1 μg oligo(dT)18. Quantitative PCR analysis was performed using iQ SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) and a CFX96 Real-Time PCR system (Bio-Rad).
To examine COOLAIR expression in the COOLAIR promoter deletion lines, Total RNAs were extracted using the RNeasy Plus Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions, followed by the digestion of residual genomic DNA by the gDNA wiper (Vazyme, Nanjing, China). 1.0 µg RNA from each sample was taken for cDNA synthesis using the HiScript III 1st Strand cDNA Synthesis Kit (Vazyme) with the transcript-specific primers (5′-TGGTTGTTATTTGGTGGTGTGAA-3′ for COOLAIR class I and 5′- GCCCGACGAAGAAAAAGTAG-3′ for class II; Zhao et al., 2021). Semi-quantitative PCR amplifications were performed, followed by agarose gel separation of PCR products.
Immunoblotting
Total protein was isolated from –100 mg of seedlings using a buffer containing 50 mM Tris-HCl (pH 7.5), 100 mM NaCl, 10 mM MgCl2, 1 mM EDTA, 10% glycerol, 1 mM PMSF, 1 mM DTT and 1×cOmplete EDTA-free Protease Inhibitor Cocktail (Roche). Fifty micrograms of total protein were loaded onto SDS-PAGE gels and separated by electrophoresis. The proteins were transferred to PVDF membranes (Amersham Biosciences) and probed with an anti-myc (MBL; #M192-3; 1:10,000 dilution) antibody overnight at 4 °C. The samples were then probed with horseradish peroxidase (HRP)-conjugated anti-mouse IgG (Cell Signaling, Danvers, MA, USA; #7076; 1:10,000 dilution) antibody at room temperature. The signals were detected with ImageQuant LAS 4000 (GE Healthcare) using WesternBright Sirius ECL solution (Advansta, San Jose, CA, USA).
RNA-seq data analysis
We retrieved two sets of RNA sequencing (RNA-seq) data deposited in the National Center for Biotechnology Information (NCBI) under BioProject accession codes PRJNA416120 and PRJNA732005. PRJNA416120 contains raw reads from NV, 1V, and 14V wild-type and cbfs plants in a Sweden-ecotype (SW) background (Park et al., 2018a). PRJNA732005 contains reads from 0, 3, and 24 hr cold-treated wild-type and cbfs plants in a Col-0 background (Song et al., 2021). The reads were aligned to the Arabidopsis TAIR 10 reference genome and annotated in ARAPORT 11 using STAR version 2.7.10 a. Isoform estimation was performed using Salmon version 1.6.0.
Luciferase assay using Arabidopsis protoplast
Protoplast isolation and transfection were performed as previously described with some modifications (Yoo et al., 2007). Protoplasts were isolated from leaves of SD-grown wild-type plant (Col-0) using a buffer containing 150 mg Cellulase Onozuka R-10 (Yakult, Tokyo, Japan), 50 mg Maceroenzyme R-10 (Yakult), 20 mM KCl, 20 mM MES-KOH (pH 5.6), 0.4 M D-mannitol, 10 mM CaCl2, 5 mM β-mercaptoethanol, and 0.1 g bovine serum albumin. For protoplast transfection, 200 μg of plasmid DNA and isolated protoplasts were incubated in a buffer containing 0.1 M D-mannitol, 50 mM CaCl2, and 20% (w/v) PEG. Luciferase activity in the protoplasts was measured using the Luciferase Assay System (Promega, Madison, WI, USA) and MicroLumat Plus LB96V microplate luminometer (Berthold Technologies, Bad Wildbad, Germany).
Acknowledgements
We thank Dr. Shuhua Yang (China Agricultural University, China) for providing the cbfs-1, pCBF3:CBF3-myc, and pSuper:CBF3-myc seeds. This work was supported by the Cooperative Research Program for Agriculture Science and Technology Development (No. PJ01315201 and PJ01315401), Rural Development Administration, Republic of Korea. This work was also supported by National Research Foundation of Korea (NRF) grants funded by the Korean government (MSIT) (No. 2019R1A2C2004313, 2021R1A5A1032428, and 2022R1A2C1091491). Y He’s laboratory was funded by the grants from the National Natural Science Foundation of China (No. 31830049 and 31721001). M Jeon and D Jeong were supported by Brain Korea 21 Plus Project. We thank Editage (http://www.editage.co.kr) for English language editing.
Funding Statement
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Contributor Information
Yuehui He, Email: yhhe@pku.edu.cn.
Ilha Lee, Email: ilhalee@snu.ac.kr.
Richard Amasino, University of Wisconsin Madison, United States.
Jürgen Kleine-Vehn, University of Freiburg, Germany.
Funding Information
This paper was supported by the following grants:
National Research Foundation of Korea 2019R1A2C2004313 to Ilha Lee.
National Research Foundation of Korea 2022R1A2C1091491 to Ilha Lee.
National Natural Science Foundation of China 31830049 to Yuehui He.
National Natural Science Foundation of China 31721001 to Yuehui He.
National Research Foundation of Korea 2021R1A5A1032428 to Ilha Lee.
Additional information
Competing interests
No competing interests declared.
No competing interests declared.
Author contributions
Conceptualization, Data curation, Formal analysis, Validation, Investigation, Visualization, Methodology, Writing - original draft, Writing – review and editing.
Conceptualization, Data curation, Formal analysis, Validation, Investigation, Methodology, Writing – review and editing.
Conceptualization, Data curation, Formal analysis, Validation, Investigation, Visualization, Methodology.
Conceptualization, Data curation, Formal analysis, Validation, Investigation, Visualization, Methodology.
Validation, Investigation, Visualization, Methodology.
Validation, Writing – review and editing.
Validation, Writing – review and editing.
Conceptualization, Resources, Data curation, Supervision, Funding acquisition, Validation, Writing - original draft, Project administration, Writing – review and editing.
Conceptualization, Resources, Data curation, Supervision, Funding acquisition, Validation, Writing - original draft, Project administration, Writing – review and editing.
Additional files
Data availability
No new data have been generated for this manuscript. Previously published datasets used for this study are deposited in NCBI, under BioProject accession codes PRJNA416120 and PRJNA732005.
The following previously published datasets were used:
Park S, Gilmour SJ, Grumet R, Thomashow MF. 2018. Potential role of the CBF Pathway contributing to local adaptation of ecotypes collected from Italy and Sweden (thale cress) NCBI BioProject. PRJNA416120
Song Y, Zhang X, Li M, Yang H, Fu D, Lv J, Ding Y, Gong Z, Shi Y, Yang S. 2021. Genome-wide identification of CBFs targets in Arabidopsis. NCBI BioProject. PRJNA732005
References
- Angel A, Song J, Dean C, Howard M. A polycomb-based switch underlying quantitative epigenetic memory. Nature. 2011;476:105–108. doi: 10.1038/nature10241. [DOI] [PubMed] [Google Scholar]
- Baker SS, Wilhelm KS, Thomashow MF. The 5’-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold-, drought- and ABA-regulated gene expression. Plant Molecular Biology. 1994;24:701–713. doi: 10.1007/BF00029852. [DOI] [PubMed] [Google Scholar]
- Bastow R, Mylne JS, Lister C, Lippman Z, Martienssen RA, Dean C. Vernalization requires epigenetic silencing of FLC by histone methylation. Nature. 2004;427:164–167. doi: 10.1038/nature02269. [DOI] [PubMed] [Google Scholar]
- Berry S, Dean C. Environmental perception and epigenetic memory: mechanistic insight through FLC. The Plant Journal. 2015;83:133–148. doi: 10.1111/tpj.12869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castaings L, Bergonzi S, Albani MC, Kemi U, Savolainen O, Coupland G. Evolutionary conservation of cold-induced antisense rnas of flowering locus C in Arabidopsis thaliana perennial relatives. Nature Communications. 2014;5:4457. doi: 10.1038/ncomms5457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chinnusamy V, Zhu J, Zhu JK. Cold stress regulation of gene expression in plants. Trends in Plant Science. 2007;12:444–451. doi: 10.1016/j.tplants.2007.07.002. [DOI] [PubMed] [Google Scholar]
- Choi K, Kim J, Hwang HJ, Kim S, Park C, Kim SY, Lee I. The FRIGIDA complex activates transcription of FLC, a strong flowering repressor in arabidopsis, by recruiting chromatin modification factors. The Plant Cell. 2011;23:289–303. doi: 10.1105/tpc.110.075911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chouard P. Vernalization and its relations to dormancy. Annual Review of Plant Physiology. 1960;11:191–238. doi: 10.1146/annurev.pp.11.060160.001203. [DOI] [Google Scholar]
- Csorba T, Questa JI, Sun Q, Dean C. Antisense COOLAIR mediates the coordinated switching of chromatin states at FLC during vernalization. PNAS. 2014;111:16160–16165. doi: 10.1073/pnas.1419030111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Lucia F, Crevillen P, Jones AME, Greb T, Dean C. A PHD-polycomb repressive complex 2 triggers the epigenetic silencing of FLC during vernalization. PNAS. 2008;105:16831–16836. doi: 10.1073/pnas.0808687105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding Y, Jia Y, Shi Y, Zhang X, Song C, Gong Z, Yang S. OST1-mediated BTF3L phosphorylation positively regulates cbfs during plant cold responses. The EMBO Journal. 2018;37:e98228. doi: 10.15252/embj.201798228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding Y, Shi Y, Yang S. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. The New Phytologist. 2019;222:1690–1704. doi: 10.1111/nph.15696. [DOI] [PubMed] [Google Scholar]
- Ding Y, Yang S. Surviving and thriving: how plants perceive and respond to temperature stress. Developmental Cell. 2022;57:947–958. doi: 10.1016/j.devcel.2022.03.010. [DOI] [PubMed] [Google Scholar]
- Doherty CJ, Van Buskirk HA, Myers SJ, Thomashow MF. Roles for arabidopsis CAMTA transcription factors in cold-regulated gene expression and freezing tolerance. The Plant Cell. 2009;21:972–984. doi: 10.1105/tpc.108.063958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong MA, Farré EM, Thomashow MF. Circadian CLOCK-ASSOCIATED 1 and late elongated hypocotyl regulate expression of the C-repeat binding factor (CBF) pathway in arabidopsis. PNAS. 2011;108:7241–7246. doi: 10.1073/pnas.1103741108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eremina M, Unterholzner SJ, Rathnayake AI, Castellanos M, Khan M, Kugler KG, May ST, Mayer KFX, Rozhon W, Poppenberger B. Brassinosteroids participate in the control of basal and acquired freezing tolerance of plants. PNAS. 2016;113:E5982–E5991. doi: 10.1073/pnas.1611477113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang X, Wang L, Ishikawa R, Li Y, Fiedler M, Liu F, Calder G, Rowan B, Weigel D, Li P, Dean C. Arabidopsis FLL2 promotes liquid–liquid phase separation of polyadenylation complexes. Nature. 2019;569:265–269. doi: 10.1038/s41586-019-1165-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang X, Wu Z, Raitskin O, Webb K, Voigt P, Lu T, Howard M, Dean C. The 3′ processing of antisense rnas physically links to chromatin-based transcriptional control. PNAS. 2020;117:15316–15321. doi: 10.1073/pnas.2007268117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fowler S, Thomashow MF. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway [ W ] The Plant Cell. 2002;14:1675–1690. doi: 10.1105/tpc.003483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilmour SJ, Hajela RK, Thomashow MF. Cold acclimation in Arabidopsis thaliana. Plant Physiology. 1988;87:745–750. doi: 10.1104/pp.87.3.745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilmour SJ, Zarka DG, Stockinger EJ, Salazar MP, Houghton JM, Thomashow MF. Low temperature regulation of the arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced cor gene expression. The Plant Journal. 1998;16:433–442. doi: 10.1046/j.1365-313x.1998.00310.x. [DOI] [PubMed] [Google Scholar]
- Gilmour SJ, Fowler SG, Thomashow MF. Arabidopsis transcriptional activators CBF1, CBF2, and CBF3 have matching functional activities. Plant Molecular Biology. 2004;54:767–781. doi: 10.1023/B:PLAN.0000040902.06881.d4. [DOI] [PubMed] [Google Scholar]
- Guy CL. Cold acclimation and freezing stress tolerance: role of protein metabolism. Annual Review of Plant Physiology and Plant Molecular Biology. 1990;41:187–223. doi: 10.1146/annurev.pp.41.060190.001155. [DOI] [Google Scholar]
- Hajela RK, Horvath DP, Gilmour SJ, Thomashow MF. Molecular cloning and expression of cor (cold-regulated) genes in Arabidopsis thaliana. Plant Physiology. 1990;93:1246–1252. doi: 10.1104/pp.93.3.1246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helliwell CA, Wood CC, Robertson M, James Peacock W, Dennis ES. The arabidopsis FLC protein interacts directly in vivo with SOC1 and FT chromatin and is part of a high-molecular-weight protein complex. The Plant Journal. 2006;46:183–192. doi: 10.1111/j.1365-313X.2006.02686.x. [DOI] [PubMed] [Google Scholar]
- Helliwell CA, Robertson M, Finnegan EJ, Buzas DM, Dennis ES. Vernalization-repression of arabidopsis FLC requires promoter sequences but not antisense transcripts. PLOS ONE. 2011;6:e21513. doi: 10.1371/journal.pone.0021513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heo JB, Sung S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science. 2011;331:76–79. doi: 10.1126/science.1197349. [DOI] [PubMed] [Google Scholar]
- Hung FY, Shih YH, Lin PY, Feng YR, Li C, Wu K. WRKY63 transcriptional activation of COOLAIR and COLDAIR regulates vernalization-induced flowering. Plant Physiology. 2022;190:532–547. doi: 10.1093/plphys/kiac295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang I, Sheen J. Two-component circuitry in arabidopsis cytokinin signal transduction. Nature. 2001;413:383–389. doi: 10.1038/35096500. [DOI] [PubMed] [Google Scholar]
- Jean Finnegan E, Bond DM, Buzas DM, Goodrich J, Helliwell CA, Tamada Y, Yun JY, Amasino RM, Dennis ES. Polycomb proteins regulate the quantitative induction of vernalization insensitive 3 in response to low temperatures. The Plant Journal. 2011;65:382–391. doi: 10.1111/j.1365-313X.2010.04428.x. [DOI] [PubMed] [Google Scholar]
- Jia Y, Ding Y, Shi Y, Zhang X, Gong Z, Yang S. The cbfs triple mutants reveal the essential functions of cbfs in cold acclimation and allow the definition of CBF regulons in arabidopsis. The New Phytologist. 2016;212:345–353. doi: 10.1111/nph.14088. [DOI] [PubMed] [Google Scholar]
- Johanson U, West J, Lister C, Michaels S, Amasino R, Dean C. Molecular analysis of FRIGIDA, a major determinant of natural variation in arabidopsis flowering time. Science. 2000;290:344–347. doi: 10.1126/science.290.5490.344. [DOI] [PubMed] [Google Scholar]
- Kidokoro S, Yoneda K, Takasaki H, Takahashi F, Shinozaki K, Yamaguchi-Shinozaki K. Different cold-signaling pathways function in the responses to rapid and gradual decreases in temperature. The Plant Cell. 2017;29:760–774. doi: 10.1105/tpc.16.00669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim DH, Zografos BR, Sung S. Mechanisms underlying vernalization-mediated VIN3 induction in arabidopsis. Plant Signaling & Behavior. 2010;5:1457–1459. doi: 10.4161/psb.5.11.13465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim Y, Park S, Gilmour SJ, Thomashow MF. Roles of CAMTA transcription factors and salicylic acid in configuring the low-temperature transcriptome and freezing tolerance of arabidopsis. The Plant Journal. 2013;75:364–376. doi: 10.1111/tpj.12205. [DOI] [PubMed] [Google Scholar]
- Kim DH, Sung S. Coordination of the vernalization response through a VIN3 and FLC gene family regulatory network in arabidopsis. The Plant Cell. 2013;25:454–469. doi: 10.1105/tpc.112.104760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim DH, Sung S. The binding specificity of the PHD-finger domain of VIN3 moderates vernalization response. Plant Physiology. 2017a;173:1258–1268. doi: 10.1104/pp.16.01320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim DH, Sung S. Vernalization-triggered intragenic chromatin loop formation by long noncoding rnas. Developmental Cell. 2017b;40:302–312. doi: 10.1016/j.devcel.2016.12.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyung J, Jeon M, Jeong G, Shin Y, Seo E, Yu J, Kim H, Park CM, Hwang D, Lee I. The two clock proteins CCA1 and LHY activate VIN3 transcription during vernalization through the vernalization-responsive cis-element. The Plant Cell. 2022;34:1020–1037. doi: 10.1093/plcell/koab304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee I, Bleecker A, Amasino R. Analysis of naturally occurring late flowering in Arabidopsis thaliana. Molecular & General Genetics. 1993;237:171–176. doi: 10.1007/BF00282798. [DOI] [PubMed] [Google Scholar]
- Lee I, Michaels SD, Masshardt AS, Amasino RM. The late-flowering phenotype of FRIGIDA and mutations in LUMINIDEPENDENS is suppressed in the landsberg erecta strain of arabidopsis. The Plant Journal. 1994;6:903–909. doi: 10.1046/j.1365-313X.1994.6060903.x. [DOI] [Google Scholar]
- Lee H, Suh SS, Park E, Cho E, Ahn JH, Kim SG, Lee JS, Kwon YM, Lee I. The AGAMOUS-LIKE 20 MADS domain protein integrates floral inductive pathways in arabidopsis. Genes & Development. 2000;14:2366–2376. doi: 10.1101/gad.813600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee ES, Park JH, Wi SD, Kang CH, Chi YH, Chae HB, Paeng SK, Ji MG, Kim WY, Kim MG, Yun DJ, Stacey G, Lee SY. Redox-dependent structural switch and CBF activation confer freezing tolerance in plants. Nature Plants. 2021;7:914–922. doi: 10.1038/s41477-021-00944-8. [DOI] [PubMed] [Google Scholar]
- Li H, Ye K, Shi Y, Cheng J, Zhang X, Yang S. Bzr1 positively regulates freezing tolerance via CBF-dependent and CBF-independent pathways in arabidopsis. Molecular Plant. 2017;10:545–559. doi: 10.1016/j.molp.2017.01.004. [DOI] [PubMed] [Google Scholar]
- Li Z, Jiang D, He Y. Frigida establishes a local chromosomal environment for flowering locus C mrna production. Nature Plants. 2018;4:836–846. doi: 10.1038/s41477-018-0250-6. [DOI] [PubMed] [Google Scholar]
- Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K, Shinozaki K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in arabidopsis. The Plant Cell. 1998;10:1391–1406. doi: 10.1105/tpc.10.8.1391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu F, Quesada V, Crevillén P, Bäurle I, Swiezewski S, Dean C. The arabidopsis RNA-binding protein FCA requires a lysine-specific demethylase 1 homolog to downregulate FLC. Molecular Cell. 2007;28:398–407. doi: 10.1016/j.molcel.2007.10.018. [DOI] [PubMed] [Google Scholar]
- Liu F, Marquardt S, Lister C, Swiezewski S, Dean C. Targeted 3’ processing of antisense transcripts triggers arabidopsis FLC chromatin silencing. Science. 2010;327:94–97. doi: 10.1126/science.1180278. [DOI] [PubMed] [Google Scholar]
- Liu Z, Jia Y, Ding Y, Shi Y, Li Z, Guo Y, Gong Z, Yang S. Plasma membrane CRPK1-mediated phosphorylation of 14-3-3 proteins induces their nuclear import to fine-tune CBF signaling during cold response. Molecular Cell. 2017;66:117–128. doi: 10.1016/j.molcel.2017.02.016. [DOI] [PubMed] [Google Scholar]
- Luo X, Chen T, Zeng X, He D, He Y. Feedback regulation of FLC by flowering locus T (ft) and fd through a 5’ FLC promoter region in arabidopsis. Molecular Plant. 2019;12:285–288. doi: 10.1016/j.molp.2019.01.013. [DOI] [PubMed] [Google Scholar]
- Marquardt S, Raitskin O, Wu Z, Liu F, Sun Q, Dean C. Functional consequences of splicing of the antisense transcript COOLAIR on FLC transcription. Molecular Cell. 2014;54:156–165. doi: 10.1016/j.molcel.2014.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maruyama K, Sakuma Y, Kasuga M, Ito Y, Seki M, Goda H, Shimada Y, Yoshida S, Shinozaki K, Yamaguchi-Shinozaki K. Identification of cold-inducible downstream genes of the arabidopsis DREB1A/CBF3 transcriptional factor using two microarray systems. The Plant Journal. 2004;38:982. doi: 10.1111/j.1365-313X.2004.02100.x. [DOI] [PubMed] [Google Scholar]
- McKhann HI, Gery C, Bérard A, Lévêque S, Zuther E, Hincha DK, De Mita S, Brunel D, Téoulé E. Natural variation in CBF gene sequence, gene expression and freezing tolerance in the versailles core collection of Arabidopsis thaliana. BMC Plant Biology. 2008;8:105. doi: 10.1186/1471-2229-8-105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medina J, Bargues M, Terol J, Pérez-Alonso M, Salinas J. The arabidopsis CBF gene family is composed of three genes encoding AP2 domain-containing proteins whose expression is regulated by low temperature but not by abscisic acid or dehydration. Plant Physiology. 1999;119:463–470. doi: 10.1104/pp.119.2.463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michaels SD, Amasino RM. Flowering locus C encodes a novel MADS domain protein that acts as a repressor of flowering. The Plant Cell. 1999;11:949–956. doi: 10.1105/tpc.11.5.949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michaels SD, Amasino RM. Memories of winter: vernalization and the competence to flower. Plant, Cell and Environment. 2000;23:1145–1153. doi: 10.1046/j.1365-3040.2000.00643.x. [DOI] [Google Scholar]
- Michaels SD, Amasino RM. Loss of flowering locus C activity eliminates the late-flowering phenotype of FRIGIDA and autonomous pathway mutations but not responsiveness to vernalization. The Plant Cell. 2001;13:935–941. doi: 10.1105/tpc.13.4.935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michaels SD, Himelblau E, Kim SY, Schomburg FM, Amasino RM. Integration of flowering signals in winter-annual arabidopsis. Plant Physiology. 2005;137:149–156. doi: 10.1104/pp.104.052811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mylne JS, Barrett L, Tessadori F, Mesnage S, Johnson L, Bernatavichute YV, Jacobsen SE, Fransz P, Dean C. LHP1, the arabidopsis homologue of heterochromatin protein1, is required for epigenetic silencing of FLC. PNAS. 2006;103:5012–5017. doi: 10.1073/pnas.0507427103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Napp-Zinn K. Genetische grundlagen des kltebedrfnisses bei Arabidopsis thaliana (L.)heynh. Die Naturwissenschaften. 1955;42:650. doi: 10.1007/BF00622395. [DOI] [Google Scholar]
- Nishio H, Buzas DM, Nagano AJ, Suzuki Y, Sugano S, Ito M, Morinaga SI, Kudoh H. From the laboratory to the field: assaying histone methylation at flowering locus C in naturally growing arabidopsis halleri. Genes & Genetic Systems. 2016;91:15–26. doi: 10.1266/ggs.15-00071. [DOI] [PubMed] [Google Scholar]
- Novillo F, Medina J, Salinas J. Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulon. PNAS. 2007;104:21002–21007. doi: 10.1073/pnas.0705639105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Malley RC, Huang SSC, Song L, Lewsey MG, Bartlett A, Nery JR, Galli M, Gallavotti A, Ecker JR. Cistrome and epicistrome features shape the regulatory DNA landscape. Cell. 2016;165:1280–1292. doi: 10.1016/j.cell.2016.04.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park S, Lee CM, Doherty CJ, Gilmour SJ, Kim Y, Thomashow MF. Regulation of the arabidopsis CBF regulon by a complex low-temperature regulatory network. The Plant Journal. 2015;82:193–207. doi: 10.1111/tpj.12796. [DOI] [PubMed] [Google Scholar]
- Park S, Gilmour SJ, Grumet R, Thomashow MF. CBF-dependent and CBF-independent regulatory pathways contribute to the differences in freezing tolerance and cold-regulated gene expression of two arabidopsis ecotypes locally adapted to sites in sweden and italy. PLOS ONE. 2018a;13:e0207723. doi: 10.1371/journal.pone.0207723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park J, Lim CJ, Shen M, Park HJ, Cha JY, Iniesto E, Rubio V, Mengiste T, Zhu JK, Bressan RA, Lee SY, Lee BH, Jin JB, Pardo JM, Kim WY, Yun DJ. Epigenetic switch from repressive to permissive chromatin in response to cold stress. PNAS. 2018b;115:E5400–E5409. doi: 10.1073/pnas.1721241115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penfield S. Temperature perception and signal transduction in plants. The New Phytologist. 2008;179:615–628. doi: 10.1111/j.1469-8137.2008.02478.x. [DOI] [PubMed] [Google Scholar]
- Qüesta JI, Song J, Geraldo N, An H, Dean C. Arabidopsis transcriptional repressor val1 triggers polycomb silencing at FLC during vernalization. Science. 2016;353:485–488. doi: 10.1126/science.aaf7354. [DOI] [PubMed] [Google Scholar]
- Seo E, Lee H, Jeon J, Park H, Kim J, Noh YS, Lee I. Crosstalk between cold response and flowering in arabidopsis is mediated through the flowering-time gene SOC1 and its upstream negative regulator FLC. The Plant Cell. 2009;21:3185–3197. doi: 10.1105/tpc.108.063883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seo PJ, Park MJ, Lim MH, Kim SG, Lee M, Baldwin IT, Park CM. A self-regulatory circuit of circadian CLOCK-ASSOCIATED1 underlies the circadian CLOCK regulation of temperature responses in arabidopsis. The Plant Cell. 2012;24:2427–2442. doi: 10.1105/tpc.112.098723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheldon CC, Burn JE, Perez PP, Metzger J, Edwards JA, Peacock WJ, Dennis ES. The FLF MADS box gene: a repressor of flowering in arabidopsis regulated by vernalization and methylation. The Plant Cell. 1999;11:445–458. doi: 10.1105/tpc.11.3.445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheldon CC, Rouse DT, Finnegan EJ, Peacock WJ, Dennis ES. The molecular basis of vernalization: the central role of flowering locus C (FLC) PNAS. 2000;97:3753–3758. doi: 10.1073/pnas.97.7.3753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi Y, Ding Y, Yang S. Molecular regulation of CBF signaling in cold acclimation. Trends in Plant Science. 2018;23:623–637. doi: 10.1016/j.tplants.2018.04.002. [DOI] [PubMed] [Google Scholar]
- Shindo C, Aranzana MJ, Lister C, Baxter C, Nicholls C, Nordborg M, Dean C. Role of FRIGIDA and flowering locus C in determining variation in flowering time of arabidopsis. Plant Physiology. 2005;138:1163–1173. doi: 10.1104/pp.105.061309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinozaki K, Yamaguchi-Shinozaki K, Seki M. Regulatory network of gene expression in the drought and cold stress responses. Current Opinion in Plant Biology. 2003;6:410–417. doi: 10.1016/s1369-5266(03)00092-x. [DOI] [PubMed] [Google Scholar]
- Shu H, Nakamura M, Siretskiy A, Borghi L, Moraes I, Wildhaber T, Gruissem W, Hennig L. Arabidopsis replacement histone variant H3.3 occupies promoters of regulated genes. Genome Biology. 2014;15:R62. doi: 10.1186/gb-2014-15-4-r62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Y, Zhang X, Li M, Yang H, Fu D, Lv J, Ding Y, Gong Z, Shi Y, Yang S. The direct targets of cbfs: in cold stress response and beyond. Journal of Integrative Plant Biology. 2021;63:1874–1887. doi: 10.1111/jipb.13161. [DOI] [PubMed] [Google Scholar]
- Stockinger EJ, Gilmour SJ, Thomashow MF. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. PNAS. 1997;94:1035–1040. doi: 10.1073/pnas.94.3.1035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sung S, Amasino RM. Vernalization and epigenetics: how plants remember winter. Current Opinion in Plant Biology. 2004a;7:4–10. doi: 10.1016/j.pbi.2003.11.010. [DOI] [PubMed] [Google Scholar]
- Sung S, Amasino RM. Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature. 2004b;427:159–164. doi: 10.1038/nature02195. [DOI] [PubMed] [Google Scholar]
- Sung S, He Y, Eshoo TW, Tamada Y, Johnson L, Nakahigashi K, Goto K, Jacobsen SE, Amasino RM. Epigenetic maintenance of the vernalized state in Arabidopsis thaliana requires like heterochromatin protein 1. Nature Genetics. 2006;38:706–710. doi: 10.1038/ng1795. [DOI] [PubMed] [Google Scholar]
- Swiezewski S, Liu F, Magusin A, Dean C. Cold-induced silencing by long antisense transcripts of an arabidopsis polycomb target. Nature. 2009;462:799–802. doi: 10.1038/nature08618. [DOI] [PubMed] [Google Scholar]
- Thomashow MF, Stockinger EJ, Jaglo-Ottosen KR, Gilmour SJ, Zarka DG. Function and regulation of Arabidopsis thaliana cor (cold-regulated) genes. Acta Physiologiae Plantarum. 1997;19:497–504. doi: 10.1007/s11738-997-0046-1. [DOI] [Google Scholar]
- Thomashow MF. Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annual Review of Plant Physiology and Plant Molecular Biology. 1999;50:571–599. doi: 10.1146/annurev.arplant.50.1.571. [DOI] [PubMed] [Google Scholar]
- Thomashow MF. Molecular basis of plant cold acclimation: insights gained from studying the CBF cold response pathway. Plant Physiology. 2010;154:571–577. doi: 10.1104/pp.110.161794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang ZW, Wu Z, Raitskin O, Sun Q, Dean C. Antisense-mediated FLC transcriptional repression requires the P-tefb transcription elongation factor. PNAS. 2014;111:7468–7473. doi: 10.1073/pnas.1406635111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiser CJ. Cold resistance and injury in woody plants: knowledge of hardy plant adaptations to freezing stress may help us to reduce winter damage. Science. 1970;169:1269–1278. doi: 10.1126/science.169.3952.1269. [DOI] [PubMed] [Google Scholar]
- Went FW. The effect of temperature on plant growth. Annual Review of Plant Physiology. 1953;4:347–362. doi: 10.1146/annurev.pp.04.060153.002023. [DOI] [Google Scholar]
- Wood CC, Robertson M, Tanner G, Peacock WJ, Dennis ES, Helliwell CA. The Arabidopsis thaliana vernalization response requires a polycomb-like protein complex that also includes vernalization insensitive 3. PNAS. 2006;103:14631–14636. doi: 10.1073/pnas.0606385103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu Z, Fang X, Zhu D, Dean C. Autonomous pathway: flowering locus C repression through an antisense-mediated chromatin-silencing mechanism. Plant Physiology. 2020;182:27–37. doi: 10.1104/pp.19.01009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C, Fang X, Lu T, Dean C. Antagonistic cotranscriptional regulation through ARGONAUTE1 and the THO/TREX complex orchestrates FLC transcriptional output. PNAS. 2021a;118:e2113757118. doi: 10.1073/pnas.2113757118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C, Wu Z, Duan HC, Fang X, Jia G, Dean C. R-loop resolution promotes co-transcriptional chromatin silencing. Nature Communications. 2021b;12:1790. doi: 10.1038/s41467-021-22083-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang H, Howard M, Dean C. Antagonistic roles for h3k36me3 and h3k27me3 in the cold-induced epigenetic switch at arabidopsis FLC. Current Biology. 2014;24:1793–1797. doi: 10.1016/j.cub.2014.06.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang H, Berry S, Olsson TSG, Hartley M, Howard M, Dean C. Distinct phases of polycomb silencing to hold epigenetic memory of cold in arabidopsis. Science. 2017;357:1142–1145. doi: 10.1126/science.aan1121. [DOI] [PubMed] [Google Scholar]
- Yoo SD, Cho YH, Sheen J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nature Protocols. 2007;2:1565–1572. doi: 10.1038/nprot.2007.199. [DOI] [PubMed] [Google Scholar]
- Yuan W, Luo X, Li Z, Yang W, Wang Y, Liu R, Du J, He Y. A cis cold memory element and A trans epigenome reader mediate polycomb silencing of FLC by vernalization in arabidopsis. Nature Genetics. 2016;48:1527–1534. doi: 10.1038/ng.3712. [DOI] [PubMed] [Google Scholar]
- Zhao C, Zhang Z, Xie S, Si T, Li Y, Zhu JK. Mutational evidence for the critical role of CBF transcription factors in cold acclimation in arabidopsis. Plant Physiology. 2016;171:2744–2759. doi: 10.1104/pp.16.00533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Y, Antoniou-Kourounioti RL, Calder G, Dean C, Howard M. Temperature-dependent growth contributes to long-term cold sensing. Nature. 2020;583:825–829. doi: 10.1038/s41586-020-2485-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Y, Zhu P, Hepworth J, Bloomer R, Antoniou-Kourounioti RL, Doughty J, Heckmann A, Xu C, Yang H, Dean C. Natural temperature fluctuations promote COOLAIR regulation of FLC. Genes & Development. 2021;35:888–898. doi: 10.1101/gad.348362.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu P, Lister C, Dean C. Cold-induced arabidopsis FRIGIDA nuclear condensates for FLC repression. Nature. 2021;599:657–661. doi: 10.1038/s41586-021-04062-5. [DOI] [PMC free article] [PubMed] [Google Scholar]