

Abstract
Neuronal polarization is a complex molecular process regulated by intrinsic and extrinsic mechanisms. Nerve cells integrate multiple extracellular cues to generate intracellular messengers that ultimately control cell morphology, metabolism, and gene expression. Therefore, second messengers' local concentration and temporal regulation are crucial elements for acquiring a polarized morphology in neurons. This review article summarizes the main findings and current understanding of how Ca2+, IP3, cAMP, cGMP, and hydrogen peroxide control different aspects of neuronal polarization, and highlights questions that still need to be resolved to fully understand the fascinating cellular processes involved in axodendritic polarization.
Keywords: calcium, IP3, cAMP/cGMP, hydrogen peroxide, neuronal polarity
Introduction
Neurons are highly polarized cells with one axon and several dendrites. Dendrites are input sites for neurotransmission, while axons trigger action potentials that propagate to distal terminals and induce the release of neurotransmitters to target cells. To perform these distinct functions, dendrites and axons exhibit different morphologic and biochemical features. A fundamental question in neurobiology is how neurons acquire their functional and morphologic polarity. Understanding neuronal polarization mechanisms is essential for understanding neuronal development and synaptic function as well as for designing interventions that overcome nervous system dysfunctions that characterize most neurodegenerative diseases. This review focuses on the extracellular signals and second messengers involved in initial molecular polarization, local signal amplification, and global inhibition during axon specification.
Acquisition of neuronal polarity in vivo and in vitro
The process of neuronal polarization has been studied for decades. Dissociated rodent hippocampal neurons grown in vitro are the most used experimental system for these studies (Kater et al., 1988; Bradke and Dotti, 2000; Arimura and Kaibuchi, 2007; Banker, 2018). Cultured hippocampal neurons become polarized even in the absence of extracellular polarizing cues, indicating that they possess intrinsic molecular programs that underlie polarity.
After plating in culture, hippocampal and cortical neurons transit through a series of stereotyped steps that have been classified into five stages (Dotti et al., 1988; Caceres et al., 2012) (Fig. 1A). After adhesion to the substrate, neurons extend a lamellipodium veil with numerous thin filopodia that project and retract (Stage 1). After the first DIV, neurons have formed several immature neurites (Stage 2). In culture, all these neurites are morphologically indistinguishable, and they continually grow and retract. Eventually, one of these minor neurites extends rapidly and begins to express several molecular markers that distinguish it from sibling neurites; this process becomes the axon (Stage 3). This prospective axon grows and branches, while smaller neurites further develop into mature dendrites (Stage 4). Finally, the typical synaptic specializations of excitatory neurons, such as dendritic spines, develop (Stage 5), allowing synaptic transmission between neurons.
Figure 1.
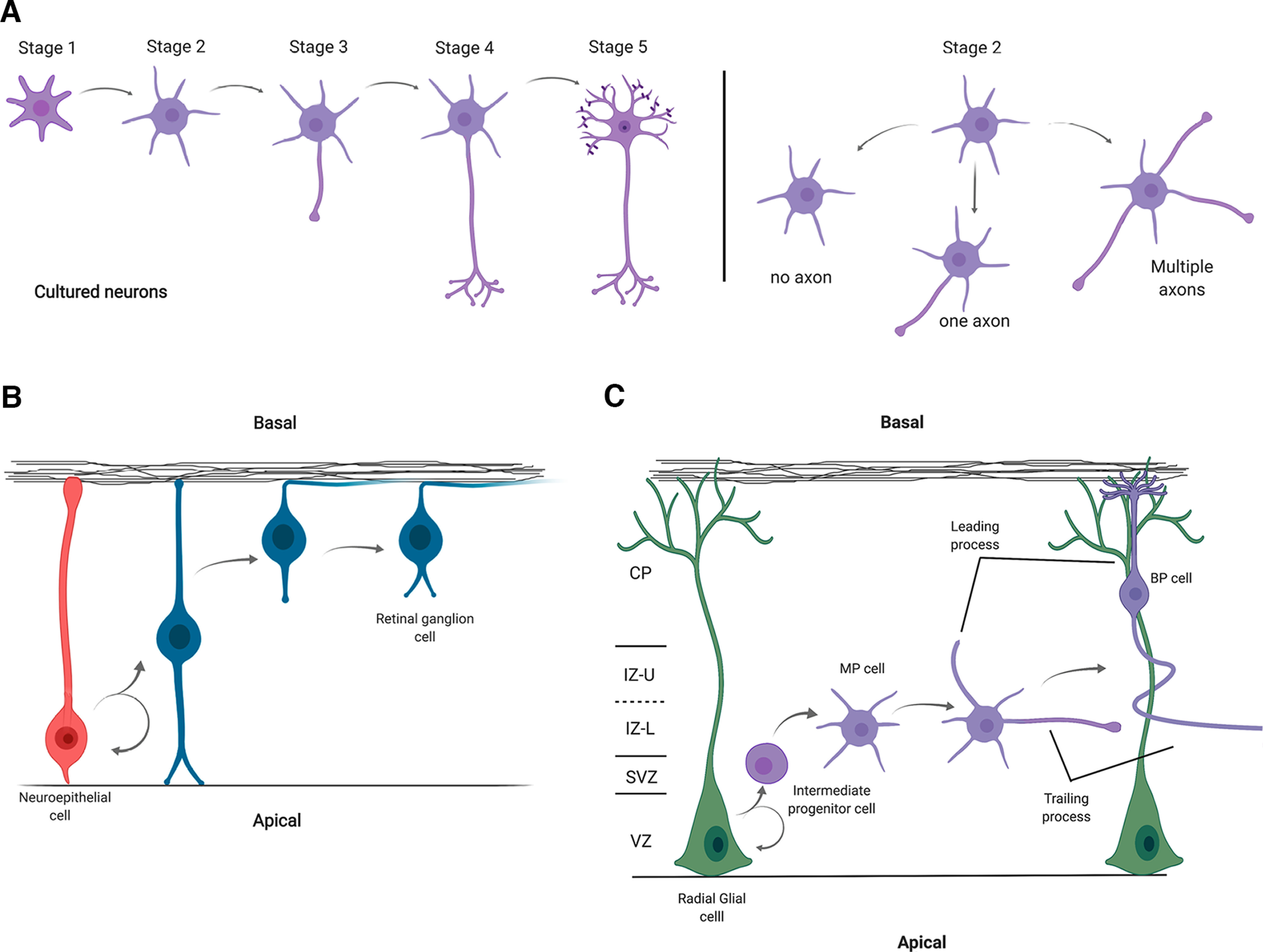
Neuronal polarization in cultured cells and in vivo. A, Left, Stage 1 neurons have several thin filopodia. Stage 2 neurons have multiple minor processes. Stage 3 neurons possess one long neurite and several minor processes. The longest neurite finally develops into an axon. Then the remaining neurites develop into dendrites (Stage 4). Finally, the neurons develop dendritic spines and axonal branches (Stage 5). Right, Experimental manipulations that reduce or increase the levels of several polarity regulators and/or associated signaling pathways result in axonal loss or the generation of multiple axons, respectively. B, Retinal ganglion cells are directly derived from neuroepithelial cells. The apical process of the daughter cell will develop into a dendrite, while the basal process that is in contact with the basal lamina becomes an axon. C, Most pyramidal neurons are generated from the radial glial cells in the ventricular zone (VZ) through the generation of intermediate progenitor cells in the subventricular zone (SVZ). The newly generated neurons in the lower part of the intermediate zone (IZ-L) extend multiple neurites (multipolar cells [MP]). Then most of the MP cells extend the trailing process tangentially (future axon) and generate the leading process (future dendrite). Finally, the multipolar cells transform into bipolar cells (BP) and migrate toward the cortical plate (CP) through radial glial cells.
In contrast to cultured neurons, neuronal polarization in vivo shows different characteristics depending on brain region and developmental stage. The acquisition of neuronal polarity in vivo can be separated into two types: neurons directly generated from neuroepithelial cells and neurons generated from neural stem cells defined as radial glial progenitors (Funahashi et al., 2014; Namba et al., 2015). Retinal ganglion cells and bipolar cells in the developing vertebrate retina are generated directly from neuroepithelial cells, and inherit the apicobasal polarity of these cells. The apical membrane develops into dendrites in these cells, whereas the basal membrane becomes the axon (Fig. 1B) (Morgan et al., 2006; Zolessi et al., 2006; Randlett et al., 2011).
In contrast, other neurons, such as cerebellar granule neurons and cortical and hippocampal pyramidal neurons, are generated directly or indirectly from radial glial cells and undergo extensive morphologic and biochemical changes as they become polarized. Although radial glial cells inherit the apical-basal polarity of the neuroepithelial cells from which they are derived, this polarity is lost during the generation of intermediate progenitors and neuronal progeny (Noctor et al., 2004; Attardo et al., 2008; Kriegstein and Alvarez-Buylla, 2009). Therefore, neurons derived from these cells must reestablish polarity (Fig. 1C) (Gao and Hatten, 1993; Komuro et al., 2001; Barnes and Polleux, 2009). This process involves a series of steps. First, symmetric divisions of the radial glial cells allow expanding their pool.
In contrast, asymmetric divisions yield a postmitotic neuron, an intermediate progenitor, that migrates away from the ventricle to continue its development. In the subventricular zone, this intermediate precursor terminally divides, generating two neurons, which continue migrating to the intermediate zone. During this process, neurons dynamically extend and retract neurites, like Stage 2 of cultured neurons; these cells are called multipolar cells. Then, most of these cells grow a trailing process toward the ventricle, which later becomes the axon, followed by the extension of another neurite basally to form the leading migratory process (Calderon de Anda et al., 2008; Hatanaka and Yamauchi, 2013; Namba et al., 2014; Sakakibara et al., 2014). These cells, now called bipolar cells, migrate to reach its destination, and develop into mature neurons (Fig. 1C). Alternatively, a minor group of pyramidal cells is generated by direct neurogenesis, where neurons proceed directly from radial glial, without intermediate precursor generation, in the deeper cortical layers (Noctor et al., 2001; Hevner, 2006).
Proposed model for axon specification
Despite these advances in understanding the establishment of neuronal polarity, how only a single axon is specified in neurons is a question that remains open.
Axonal specification can be divided into three sequential processes: initial molecular polarization (symmetry breaking), selective axonal growth, and complete molecular polarization (cytoskeletal rearrangement, cargo addressing, axon initial segment [AIS] formation, and polarity maintenance) (Schelski and Bradke, 2017).
Although hippocampal neurons typically form a single axon, a different process can become the axon if the original axon is transected in vitro (Dotti and Banker, 1987), even from mature dendrites of neurons integrated into neuronal circuits (Gomis-Ruth et al., 2008). This indicates that more than one neurite has the potential to become an axon. Therefore, neurons must have a robust mechanism to ensure the generation of only one axon; for example, the developing axon might generate an inhibitory signal preventing another neurite from developing axonal properties. It was proposed that neurite extension and retraction are controlled by positive and negative signaling molecules, which diffuse within neurites to regulate membrane recruitment, protein transport, dynamic rearrangement of the cytoskeleton, and selective regulation of protein synthesis and degradation (Arimura and Kaibuchi, 2007; Cheng and Poo, 2012; Giandomenico et al., 2022). Continuous elongation during axon specification may be supported by a positive feedback loop sustaining signaling pathway activation. As this positive feedback is activated in one neurite, a strong negative feedback signal is generated, preventing other neurites from forming a second axon (Arimura and Kaibuchi, 2007). Such negative regulation could involve membrane elimination, degradation of proteins, a decrease in dynamics of F-actin, and microtubule catastrophe (Arimura and Kaibuchi, 2007; Namba et al., 2015). Mathematical modeling has led to the conclusion that this local-activation–global-inhibition model most likely explains the process of neuronal polarization (Naoki et al., 2011; Naoki and Ishii, 2014).
In the model for axon specification described above, it is proposed that selective accumulation of positive factors, such as CMRP-2 (for microtubule polymerization and receptor trafficking), Par complex (for Rac1 and Cdc42 activation), and PI3K activators, facilitate the development of a neurite into an axon (Namba et al., 2015). This accumulation is regulated by the active anterograde transport, diffusion, and degradation of these positive factors. The local accumulation and activation of these positive factors activate signaling molecules that act as a bistable switch, which depends on the concentration of these molecules. Therefore, we propose that the production of second messengers constitutes a reliable mechanism to regulate both positive feedback loops and negative long-distance signals, underlying axon specification.
Extracellular molecules underlying axon specification
Several secreted factors, including neurotrophins and TGF-beta1 (TGF-β), regulate neuronal polarization in the hippocampus and developing neocortex in vivo (Arimura et al., 2009; Cheng et al., 2011b; Nakamuta et al., 2011). These factors act upstream of various second messengers (Table 1). BDNF and neurotrophin-3 (NT-3) act through tropomyosin receptor kinases (Trks), which activate downstream effectors, including phosphoinositide 3-kinase (PI3K) and phospholipase C (PLC). These effectors act on phosphatidylinositol 4,5-bisphosphate to produce phosphatidylinositol 3,4,5-trisphosphate (PIP3) and inositol 1,4,5-trisphosphate (IP3), respectively (Yano and Chao, 2000; Reichardt, 2006). PIP3 is concentrated at the tip of the nascent axon during Stage 3 in culture, and the inhibition of PI3K prevents axon formation, indicating that it plays a critical role in axon specification (Shi et al., 2003; Menager et al., 2004). PIP3 recruits and allows the activation of phosphoinositide-dependent kinase (PDK), resulting in the phosphorylation and activation of Akt, which in turn phosphorylates and consequently inactivates glycogen synthase kinase-3β (GSK-3β). The inactivation of GSK-3β leads to dephosphorylation and activation of microtubule assembly-promoting proteins, such as collapsin response mediator protein-2 (CRMP-2) and microtubule stabilizing factors, such as Tau, triggering neurite extension and subsequent axonal development (Jiang et al., 2005; Yoshimura et al., 2005; W. Y. Kim et al., 2006; Tan et al., 2013). PIP3 also activates the small guanosine triphosphatase (GTPase) Rac1 through activation of another small GTPase, Cdc42, along with the Par complex and Rac guanine exchange factors (GEFs), such as Tiam or Stef (Shi et al., 2003; Suzuki and Ohno, 2006; McCaffrey and Macara, 2009). The Par protein complex, which consists of atypical PKC (aPKC), Par3, and Par6, accumulates at the tips of axons, mediating signaling from Cdc42 to Rac1, and thus promoting axon specification through actin filament remodeling (Shi et al., 2003; Nishimura et al., 2005; González-Billault et al., 2012). It is proposed that Rac1 can activate PI3K, generating a positive feedback loop that sustains its activity (Namba et al., 2015).
Table 1.
Extracellular signals and their downstream second messengers that regulate neuronal polarization
| Extracellular signals | Regulation | Cell type | Effect | Model | 2° messenger associated | Downstrean effectors | References |
|---|---|---|---|---|---|---|---|
| BDNF | Autocrine or paracrine | Hippocampal and motor neurons | Neurite growth and axon differentiation | In vitro | cAMP calcium | PKA/LKB1 CaMKII/LIMK | Cheng et al., 2011b; Saito et al., 2013; Dombert et al., 2017 |
| NT-3 | Autocrine or paracrine | Hippocampal and cortical neurons | Neurite growth and axon differentiation | In vitro, in vivo | cAMP calcium IP3 | PKA/LKB1 CaMKI/RhoA, CaMKI/MARK2 IP3R | Nakamuta et al., 2011 |
| Wnt5a | Autocrine or paracrine | Cortical neurons | Neurite growth and axon differentiation | In vitro, in vivo | Calcium IP3 | PKC CaMKII/GSK-3 IP3R/TRPC | Zhang et al., 2007; Hutchins and Kalil, 2008; Li et al., 2009, 2010; Song et al., 2010; Hutchins et al., 2011; Horigane et al., 2016 |
| IGF-1 | Undefined | Retinal ganglion cell and hippocampal neurons | Axonal growth | In vitro, in vivo | Calcium | Undefined | Sosa et al., 2006; Nieto Guil et al., 2017; Guo et al., 2018 |
| TGF-β | Autocrine or paracrine | Cortical and hippocampal neurons | Axon differentiation and growth | In vivo | Undefined | Undefined | Yi et al., 2010 |
| Reelin | Paracrine | Cortical neurons | Growth cone motility and axonal growth | In vitro | Undefined | Undefined | Chen et al., 2005; Leemhuis et al., 2010 |
| Glutamate | Autocrine or paracrine | Hippocampal neurons | Impair axonal growth and branching | In vitro | Calcium | Calcineurin/AKT/GSK-3β | Y. Wang et al., 2018 |
| Sema3A | Paracrine | Hippocampal and cortical neurons | Impair axonal development/promotes dendrite growth | In vitro, in vivo | cGMP ROS | PDE4 CRMP-2 | Morinaka et al., 2011; Shelly et al., 2011 |
Other extracellular cues, such as Wnts, insulin-like growth factor 1 (IGF-1), TGF-β, and Reelin, have also been implicated in axon specification or elongation. Neuronal aPKC is directly activated by Dishevelled (Dvl), an immediate downstream effector of Wnt5a signaling, which preferentially accumulates at the tip of the growing axon. Moreover, overexpression of Dvl leads to neurons with supernumerary axons while Dvl downregulation induces axonal loss, which is rescued by overexpression of aPKC (Zhang et al., 2007). Activation of the IGF-1 receptor is essential for establishing hippocampal neuronal polarity and initiating axonal outgrowth through its activation of the PI3K–Cdc42 pathway (Sosa et al., 2006). TGF-β induces rapid growth and differentiation of an axon in vivo and in vitro via activation of the TGF-β receptor type 2 (TβR2). Moreover, genetic enhancement of TβR2 activity promotes the formation of multiple axons by site-specific phosphorylation of Par6 (Yi et al., 2010). Reelin increases growth cone motility and filopodia formation by activating Cdc42 in a mechanism that requires apolipoprotein E receptor 2, Disabled-1, and phosphatidylinositol 3-kinase (Leemhuis et al., 2010).
Despite these advances in understanding the establishment of neuronal polarity, how only a single axon is specified in cultured hippocampal neurons is a question that remains open. Accordingly, it has been proposed that signaling factors can create positive and negative feedback loops that can affect one another (Andersen and Bi, 2000; Arimura and Kaibuchi, 2007).
Second messengers as diffusible mediators regulating axon-dendrite polarization
Second messengers are small intracellular signaling molecules produced enzymatically to converting extracellular cues into intracellular effects. Their concentration can be quickly modified in specific cell compartments, and they can integrate information from multiple independent upstream inputs that influence the rates of their synthesis and degradation. Finally, second messengers can have multiple downstream targets, thereby simultaneously amplifying cell signaling to affect several cellular systems.
Here we will discuss how cAMP/cGMP, calcium (Ca2+), inositol triphosphate (IP3), and reactive oxygen species (ROS) are implicated in the regulation of axon-dendrite polarization.
Calcium
Ca2+ plays an important role in neuronal cells, where it mediates essential physiological processes, including regulation the establishment of polarity (Berridge, 1998). However, the Ca2+ concentration must be highly regulated in specific compartments because an excessively high Ca2+ concentration is cytotoxic (Peters, 1986; Weber, 2012). The regulation of Ca2+ homeostasis in neurons involves several calcium-conducting channels and numerous calcium-dependent proteins, including kinases, phosphatases, transcription factors, and other enzymes. Calcium entry into the cytoplasm from the extracellular space is mediated by voltage-gated Ca2+ channels (VGCCs), calcium-conducting AMPARs, NMDARs, Ca2+ release-activated Ca2+ (CRAC) channels, and canonical transient receptor potential channels (TRPCs). In addition, ryanodine receptors (RyRs) and inositol 1,4,5-triphosphate receptors (IP3Rs) mediate calcium efflux from the endoplasmic reticulum (ER), a major calcium store in cells, to the cytoplasm. Subsequently, the depletion of ER calcium stores induces the activation of calcium-conducting channels to mediate calcium influx and store refilling, a process called store-operated calcium entry (SOCE) (Toescu, 2007). Both TRPC and CRAC channels participate in this process, which culminates with calcium ions being transferred from the cytosol to the ER lumen by Sarco/endoplasmic reticulum Ca2+-ATPase (SERCA) (Fig. 2).
Figure 2.
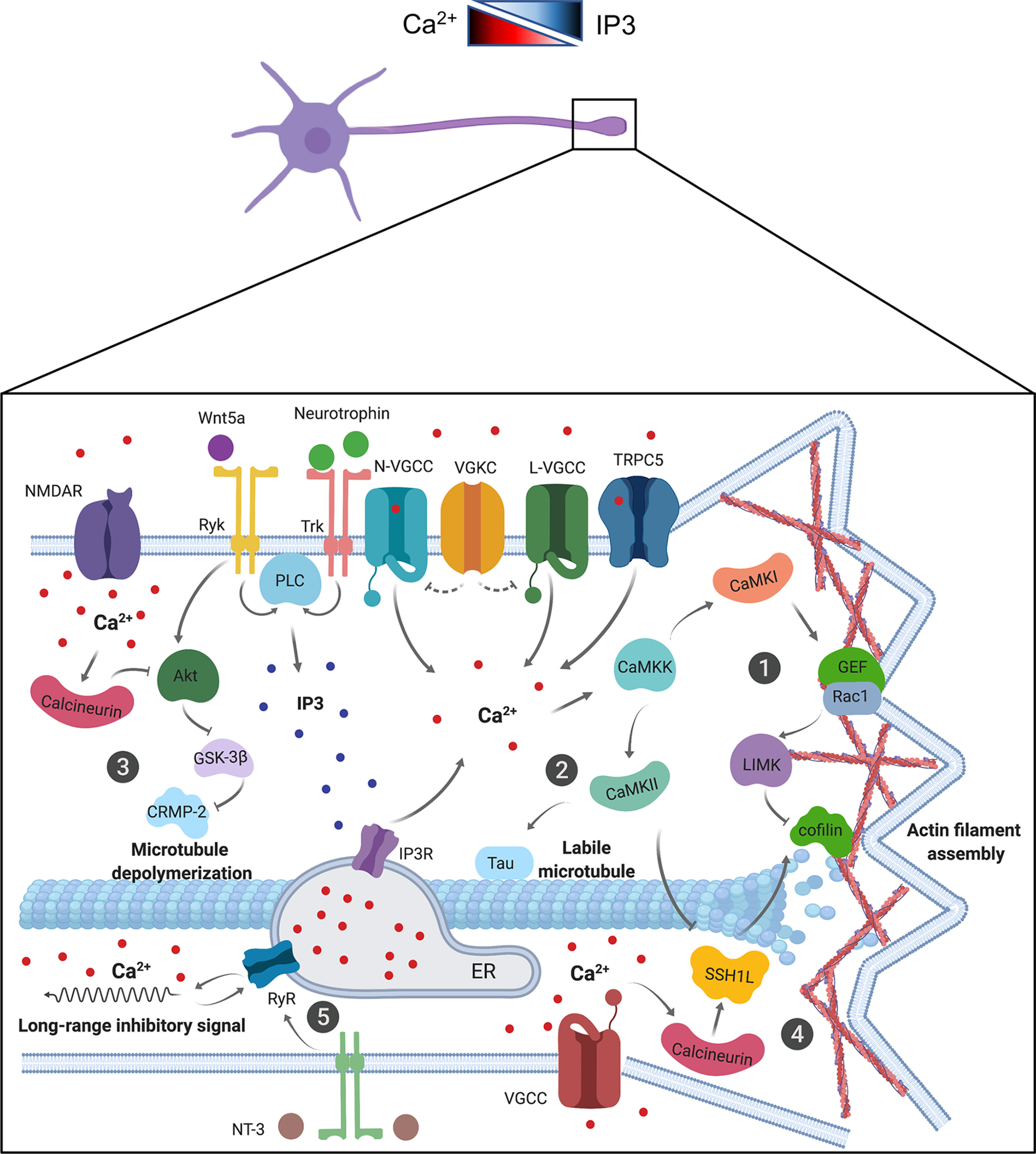
Calcium and IP3 signaling pathways are involved in neuronal polarization. Top, Schematic representation of Ca2+ and IP3 gradients in neurons. Bottom, Diagram showing a growth cone. NMDAR, N-VGCC, L-VGCC, and TRPC5 are responsible for calcium influx from extracellular sources, whereas RyR and IP3R induce calcium release from the endoplasmic reticulum (ER). Alternatively, Ryk and Trk activation by Wnt5a and neurotrophin induce PLC-mediated IP3 production. Calcium activates CaMKK or calcineurin phosphatase, impacting several effectors to regulate axon elongation. ①, CaMKK activates the CaMKI-Rac1-LIMK-cofilin and CaMKII-SSH1L-cofilin signaling pathways allowing actin filament assembly and ② the CaMKII-tau1 signaling pathway increasing the labile MT fraction. ③, Calcineurin represses Akt-GSK-3-CRMP-2 signaling, thus inducing MT depolymerization and activating cofilin phosphatase SSH1L ④. ⑤, NT3-signaling is responsible for the generation of long-range inhibitory calcium waves from the axonal tip to the soma through activation of RyR.
The first evidence that Ca2+ regulates the establishment of neuronal polarity was reported in 1987 (Mattson and Kater, 1987). This study strongly suggested that generating a calcium gradient is essential for axon specification. Axon formation is inhibited in a medium without calcium or by using calcium-permeating agents (ionophores). Moreover, the formation of a Ca2+ gradient through the focused application of a calcium ionophore in neurons induces axon formation at a site distant from the site of ionophore application, indicating that the axon tends to form where intracellular calcium levels are low (Mattson and Kater, 1987; Mattson et al., 1988a, 1990; Holliday et al., 1991). These findings suggest that intracellular Ca2+ levels control axonal differentiation and that gradients of intracellular Ca2+ might play a role in establishing neuronal polarity during development.
Several studies have shown that calcium differentially regulates growth cone motility and neurite elongation. Transient elevation of intracellular calcium levels promotes filopodium formation and inhibits neurite elongation (Kater et al., 1988; al-Mohanna et al., 1992; Rehder and Kater, 1992). However, other studies have demonstrated that elevating intracellular calcium concentration in growth cones promotes neurite outgrowth (Connor, 1986; Ciccolini et al., 2003; Brailoiu et al., 2005), suggesting that there is an optimal range for intracellular calcium that supports maximal outgrowth. Moreover, calcium levels reduction, using low concentration of calcium channel blockers, accelerates neurite elongation and reduces growth cone filopodia and lamellipodia. While high concentration of calcium channel blockers supress neurite elongation and growth cone filopodia and lamellipodia retraction, indicating that calcium differentially regulates elongation and growth cone motility (Mattson and Kater, 1987; Kater et al., 1988; Mattson et al., 1988a, 1988b; Letourneau, 1996).
Calcium regulates multiple components of neurite growth depending on its overall concentration and/or fluctuations in its concentration (Tang et al., 2003; Gomez and Zheng, 2006). Some studies shown that the frequency and amplitude of calcium transients are inversely proportional to rates of axon outgrowth (Tang et al., 2003). Accordingly, its proposed that low-frequency calcium waves enhance axon elongation, whereas high-frequency calcium waves mediate inhibitory signaling, inhibiting axonal growth (Takano et al., 2017).
The activation of specific channels that generate calcium transients with distinct kinetics and spatial spread could underpin the discrepancy observed among some studies, as well as differences in the resting calcium levels in different neuron types (Tang et al., 2003; Hutchins and Kalil, 2008). Numerous calcium channels are known to modulate the frequency and amplitude of calcium transients, therefore regulating axonal outgrowth (Fig. 2). TRPC calcium channels are associated with axon generation, were TRPC channels inactivation markedly blocks axon formation (Davare et al., 2009). However, the main channels implicated in this regulation are L-type and N-type VGCCs, which are expressed in cortical and hippocampal neurons during development (Hell et al., 1993; Dolmetsch et al., 2001; Timmermann et al., 2002; Bouron, 2020). The predominant pore-forming α1 subunits of L-type VGCCs are Cav1.2 and Cav1.3, and their expression is higher in the developing cortex than in the adult cortex. KO of endogenous Cav1.2 reduces total neurite length (Kamijo et al., 2018). In contrast, gain of function of Cav1.2 in mechanosensory neurons of C. elegans, causes the growth of an ectopic process from the cell body, and axons extend beyond their normal termination point (Buddell and Quinn, 2021).
N-type VGCC (Cav2.2) channels promote axonal elongation by generating spontaneous Ca2+ transients mediated by BDNF/TrkB signaling. TrkB receptors and Cav2.2 channels are clustered close together at axon tips in axonal growth cones of motor neurons. BDNF enhances Cav2.2 clustering and calcium influx, leading to phosphorylation of LIM kinases and cofilin, promoting F-actin assembly in growth cones (Dombert et al., 2017). IGF-1 also induces an increase in intracellular Ca2+ by promoting VGCC activation in a PI3K-dependent manner, and this promotes axon growth in RGCs (Blair and Marshall, 1997; Guo et al., 2018).
Importantly, Ca2+ levels must be optimized for normal axonal growth because excess calcium inhibits axonal elongation. For example, high Ca2+ influx through VGCCs suppresses LIM kinase 1 expression, allowing cofilin dephosphorylation and suppressing axonal elongation (Tojima et al., 2003). Voltage-gated K+ (Kv) channels prevent excessive calcium influx through VGCCs. When transiently expressed in the axonal growth cones of many embryonic neurons, Kv3.4 reduces excessive Ca2+ influx through Cav channels, reducing the frequency and amplitude of Ca2+ transients.
The role of calcium in the establishment of neuronal polarity mainly involves the regulation of cytoskeletal dynamics mediated by the activation of several calcium-binding proteins, including calcineurin and calmodulin-dependent protein kinases (CaMKs) (Singh et al., 1996; Yamamoto et al., 2002; Wen et al., 2004; Li et al., 2014). In polarized neurons (Stage 3), overexpression of constitutively active isoforms of CaMKI or CaMKK stimulates growth of the established axon (Wayman et al., 2004; Nakamuta et al., 2011); whereas overexpression of these proteins before Stage 1 induces the generation of neurons with multiple axons (Davare et al., 2009). In contrast, overexpression of dominant-negative isoforms of CaMKI or CaMKK causes growth cone collapse and significantly reduces total axonal length and branching (Wayman et al., 2004). Similar effects have been observed using specific inhibitors: inhibiting CaMKI partially blocks the formation of the axon and inhibiting CaMKK completely and reversibly impairs axon formation and elongation (Davare et al., 2009). The phosphorylation and, therefore activation of CaMKI in this context, depends on TRPC5 channel activity.
Downstream signaling associated with CaMKI activation might involve Rac1 activation. It has been reported that the Rac1-GEFs β-PIX, Tiam, and Stef can be activated by CaMKK/CaMKI signaling in dendrites (Takemoto-Kimura et al., 2007; Saneyoshi et al., 2008). A localized increase of CaMKI activity because of calcium influx via TRPC5 channels might similarly induce GEF-mediated modulation of Rac1 signaling in neurite growth cones (Fig. 2). CaMKI also phosphorylates microtubule affinity-regulating kinase 2 (MARK2) (Uboha et al., 2007). MARK2 negatively regulates axon generation; knockdown of MARK2 induces the formation of multiple axons and promotes axon outgrowth, whereas ectopic expression of MARK2 causes phosphorylation of tau (Ser262) and results in axonal loss (Chen et al., 2005).
CaMKII is implicated in regulating axon growth by multiple ways. Like CaMKI, the pharmacological or genetic inhibition of CaMKII impairs axon outgrowth in peripheral and CNSs. One way CaMKII affects axon growth is by phosphorylating Ser262 in the Tau microtubule-binding site, thus controlling the reorganization of microtubule dynamics in cortical growth cones (Li et al., 2014). In addition, CaMKII inhibition affects the length of the F-actin filaments in DRG neuronal growth cones (Xi et al., 2019). It has been reported that CaMKII participates in Wnt5a signaling (see Fig. 5). Local stimulation with Wnt5a generates axon growth by triggering calcium transients leading to the activation of calcium-sensitive effectors, such as PKC and CaMKII (Hutchins et al., 2011; Horigane et al., 2016). In addition, CaMKII is reported to phosphorylate and inactive GSK-3β in neurons, reinforcing CaMKII role in neuronal polarity (Song et al., 2010). CaMKII is also involved in BDNF-induced neuritogenesis via activation of LIMK1, which regulates actin cytoskeletal reorganization by phosphorylating and inactivating actin-depolymerizing factor and cofilin (Saito et al., 2013).
Figure 5.
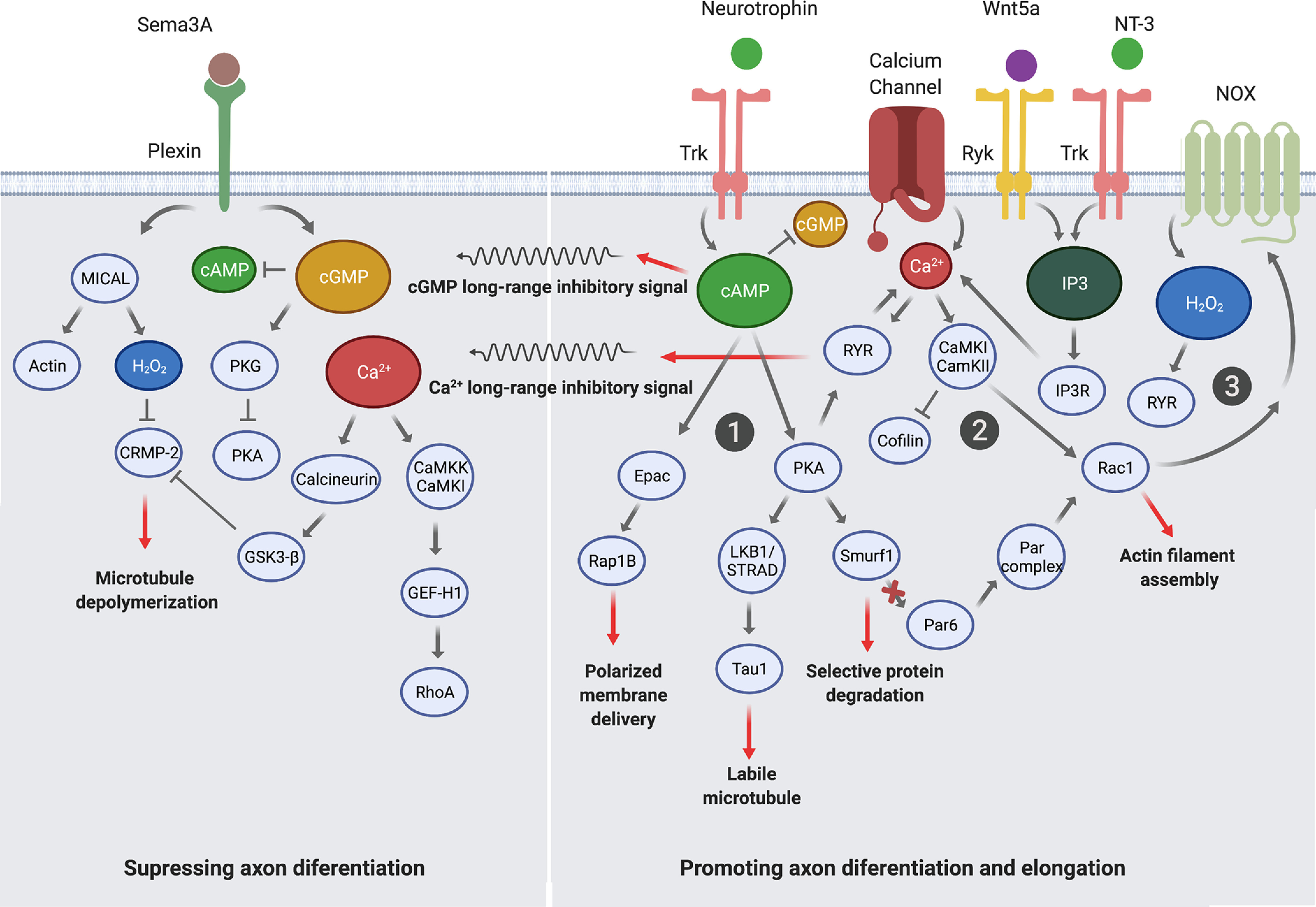
Crosstalk between second messenger signaling pathways is involved in neuronal polarization. Right, Diagram showing a growth cone and signaling pathways promoting axon differentiation and elongation. Left, Diagram showing a minor neurite and signaling pathways suppressing axon differentiation. ①, Trk activation induces the generation of cAMP, which activates the EPAC-Rap1b signaling pathway controlling polarized membrane delivery; the PKA-Par complex-Rac1 signaling pathway, triggering actin filament assembly; RyR activation triggering calcium release; and finally a cGMP long-inhibitory signal, reducing cAMP levels in the soma and other neurites. ②, Calcium influx from calcium channels and release from ER mediated by RyR and IP3R induce CaMKK-mediated cofilin inactivation and Rac1 activation, generating actin filament assembly and NOX activation. Additionally, calcium generates an RyR-mediated calcium long-range inhibitory signal to the other neurites, which activates calcineurin, activating GSK-3-CRMP-2 signaling and inducing MT depolymerization. ③, H2O2 production by the NOX complex leads to RyR and IP3R activation, producing calcium release, which finally activates Rac1, generating a positive feedback loop and enhancing actin filament polymerization. In the other dendrites, Sema3A-dependent MICAL activation induces oxidation of CRMP-2, promoting its GSK-3β-dependent phosphorylation, and thus inducing MT depolymerization.
The RyR also participates in neuronal polarization by generating long-inhibitory Ca2+ waves that induce shortening of minor neurite length to ensure the formation of a single axon (Nakamuta et al., 2011; Takano et al., 2017). Local application of NT-3 to axon terminals generates long-range RyR-dependent Ca2+ waves, which act as inhibitory signals propagated from the axon terminal to the cell body, increasing the phosphorylation of CaMKI in the cell body and the axon, but not in minor neurites (Fig. 2). The minor neurite retraction induced by local application of NT-3 in the axon terminal is mediated by activation of RhoA GTPase in cell bodies, which diffuses only into short minor neurites (but not into the long axons), preventing their outgrowth by regulating its own GEF, named GEF-H1, a CaMKI substrate (Takano et al., 2017).
NMDAR-mediated Ca2+ influx also is involved in axonal outgrowth. NMDAR subunits NR1, NR2A, and NR2B are enriched in axonal tips of cultured hippocampal neurons at Stages 2/3, with lower expression at Stages 4/5 (Herkert et al., 1998; P.Y. Wang et al., 2011; Y. Wang et al., 2018). Activation of NMDARs inhibited axonal outgrowth and branching in immature neurons, by inactivation of Akt and activation of GSK-3β in a calcium/calcineurin-dependent manner (Y. Wang et al., 2018). Furthermore, Ca2+ transients, acting via calcineurin, have been reported to reduce neurite extension by promoting depolymerization of actin filaments by suppressing LIM kinase 1 expression (Tojima et al., 2003) and by activating slingshot phosphatase SSH1L, which induces cofilin dephosphorylation (Lau et al., 1999; Lautermilch and Spitzer, 2000; Figge et al., 2012) (Fig. 2).
Finally, the nAChR has been reported to negatively regulate axonal elongation. In particular, α7 nAChRs are expressed during development and contribute to the CNS's cholinergic mechanisms of growth and plasticity. Calcium signaling mediated by α7 nAChRs at the growth cone activates the cytoskeletal regulatory enzyme calpain, leading to spectrin cleavage and reducing microtubule elongation (King and Kabbani, 2018).
In summary, calcium plays a dual role in axon specification and neurite growth, making it the best candidate to coordinate extracellular signals in the model of local activation and global inhibition during neuronal polarization. Calcium transients of lower frequency promote axon generation by regulating the activity of CMRP-2 through inactivation of GSK-3β, thus regulating MT assembly, and by activating Rac1 and LIMK/cofilin to regulate F-actin dynamics. At the same time, calcium transients of high-frequency act as long-range inhibitory signals through the generation of Ca2+ waves, propagated from the axon terminal to the cell body, increasing RhoA activity in the cell body through CaMKI and ensuring the elongation of only one axon. More studies evaluating the dual role of calcium in neuronal polarity should be performed to understand better the mechanisms that regulate the acquisition of polarity and, more importantly, how it is possible to generate calcium waves of different frequencies and magnitudes.
IP3
Many extracellular stimuli act on plasma membrane receptors coupled to PLC, which hydrolyzes phosphatidylinositol 4,5-bisphosphate to release IP3 into the cytosol. IP3 activates IP3R, a Ca2+ channel localized on the ER membrane, leading to the generation of calcium transients through calcium release from internal stores.
Accordingly, the neurotrophic factor NT-3 induces axon specification by rapidly increasing Ca2+ in growth cones in an IP3-dependent way, leading to activation of a CaMKK (Nakamuta et al., 2011). Moreover, NGF, which contributes to axon elongation and turning during axonal guidance, also induces IP3 changes in the growth cones of growing axons asymmetrically to control the pathfinding and turning responses. Interestingly, locally photolysis-induced IP3 production in one side of the growth was sufficient to induce a growth cone turning, same response triggered by NGF, reinforcing the notion that temporal and local control of this second messenger constitute an additional regulation system in the acquisition of neuronal polarity (Akiyama et al., 2009).
The role of IP3 during axonal elongation also has been studied in relation to Wnt5a signaling. Wnt5a increases outgrowth of cortical axons by activating the atypical tyrosine kinase receptor, which induces calcium transients through activation of IP3 receptors. Moreover, this effect is mimicked using activators of PLC (Li et al., 2009, 2010; Hutchins et al., 2011, 2012).
Notably, it has been reported that IP3-dependent signaling can be regulated by the inositol triphosphate 3 kinase in dendritic spines (Johnson and Schell, 2009). As some of the events that regulate dendritic spine morphology can also operate during polarity acquisition, it remains open as to whether such mechanisms can determine axon specification or elongation.
Previous reports show that IP3R inhibitors inhibit axonal elongation, indicating a role of IP3R in axonal growth (Takei et al., 1998; Iketani et al., 2009). More recently, it was reported that the Type 3 IP3R is fundamental to transduce NGF extracellular signals. Interestingly, growth cones deficient in this receptor become hypersensitive to NGF and lose their response to polarized cues. It was proposed that such abnormal behavior is associated with a global rather than a local modification of calcium levels, supporting the idea that calcium itself must be tightly controlled during polarization events (Chan et al., 2020) (Fig. 2) (see above).
In summary, to date, the role of IP3 in neuronal polarity seems to be as a calcium signal modulator, allowing local activation of polarity mediators by local calcium release. Importantly, there is still fragmented knowledge on the roles of IP3 in neuronal differentiation, and we anticipate that novel molecular mechanisms will be discovered.
cAMP and cGMP
The cAMP signaling pathway is essential for numerous neuronal functions, including neuronal development, cellular excitability, synaptic plasticity, learning, and memory (Pasterkamp, 2012). The cAMP signaling pathway involves GPCRs, which are activated at the plasma membrane on ligand binding. An active G-protein is released and in turn activates a membrane-bound adenylyl cyclase (AC) to generate cAMP from ATP. cAMP binds and activates three main effector proteins: the cyclic-nucleotide-gated ion channels, the protein kinase A (PKA), and exchange proteins activated by cAMP (Epac). The cAMP signal is then terminated by the actions of the cAMP-degrading enzymes phosphodiesterases (PDEs) (Batty et al., 2017).
In 1994, the first evidence suggesting a role for cAMP in axonal development was reported, using Xenopus spinal neurons. Local perfusion with a water-soluble, membrane-permeable analog of cAMP (8-Br-cAMP) increased neurite extension (Zheng et al., 1994). Further studies, using cortical and hippocampal neurons, revealed that the effect of cAMP in polarity is mediated by activation of PKA, which phosphorylates LKB1 at the Ser431 site, promoting axonal specification (Barnes et al., 2007; Shelly et al., 2007) (Fig. 3).
Figure 3.
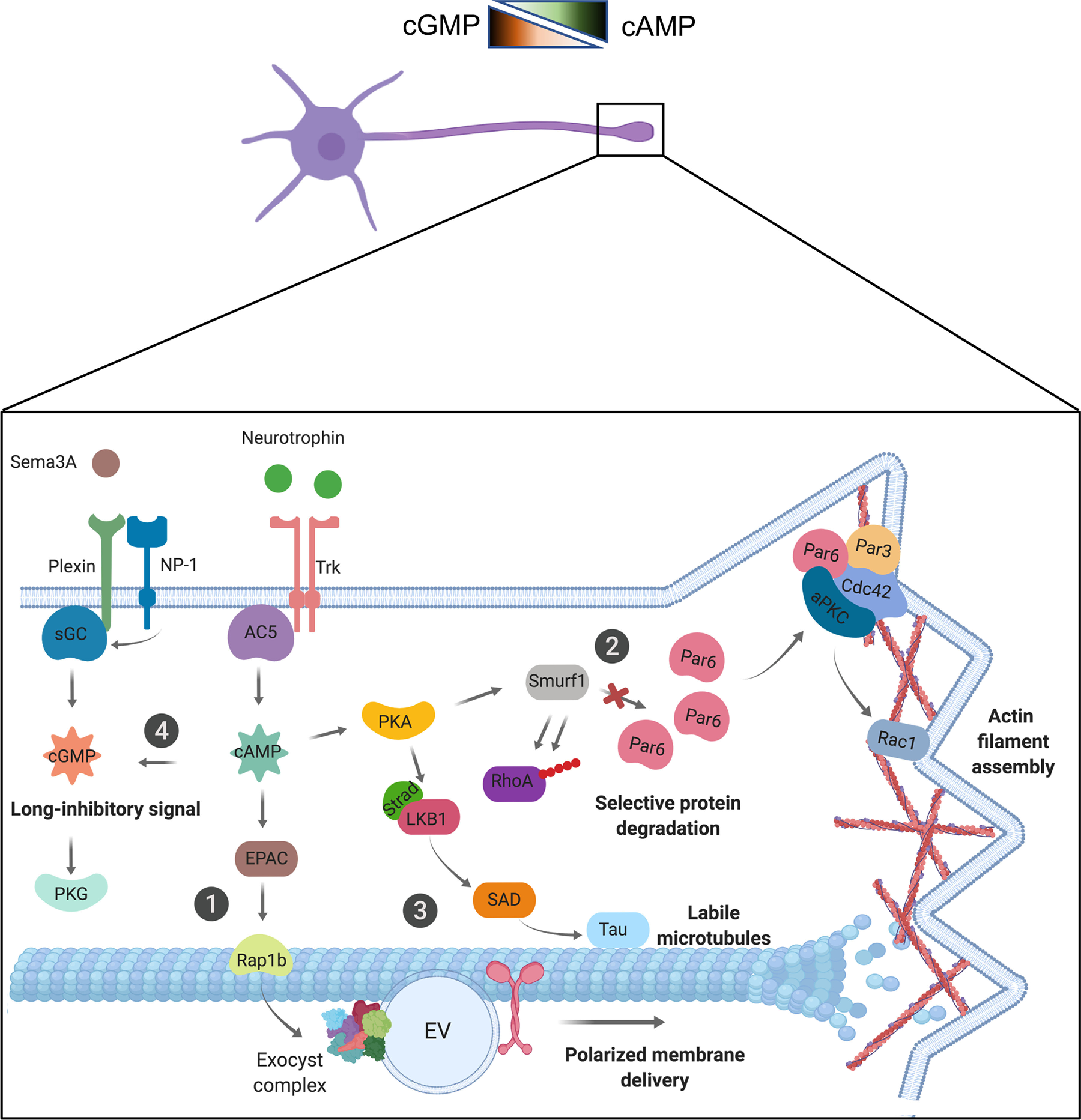
cAMP and cGMP signaling pathways involved in neuronal polarization. Top, Schematic representation of cAMP and cGMP gradients in neurons. Bottom, Diagram showing a growth cone. Trk activation by ligand binding activates adenylyl cyclase 5 (AC5), inducing the generation of cAMP. In contrast, inhibitory signals like Plexin or NP-1 induce the activation of soluble GC (sGC), triggering cGMP production. cAMP and cGMP impact several effectors to regulate axon elongation. ①, cAMP activates the EPAC-Rap1b signaling pathway controlling exocyst complex-mediated polarized membrane delivery, and ② PKA-Par complex-Rac1 signaling pathway, triggering actin filament assembly. ③, PKA also activates LKB1-tau signaling, increasing the labile microtubule fraction. ④, Finally, cAMP production induces a cGMP long-inhibitory signal, reducing cAMP levels in the soma and other neurites. EV, Exocytic vesicle.
Extracellular molecules that regulate axonal specification by modulating cAMP levels include neurotrophins. Local exposure of undifferentiated neurites to BDNF promotes axon differentiation regulated by PKA-dependent LKB1 phosphorylation (Shelly et al., 2007). It induces additional BDNF release together with an increased plasma membrane insertion of the BDNF receptor TrkB, resulting in continued cAMP and PKA signaling and generating a nested positive-feedback loop in the nascent axon (Cheng et al., 2011b).
Modulation of cAMP levels has also been observed downstream of activation of metabotropic GABAA receptors (GABABR) during neuronal development (Bony et al., 2013). In vivo, GABABR activation in PN progenitors reduces cAMP-dependent phosphorylation of LKB1, inhibiting axonal formation. Furthermore, GABABR knockdown impairs neuronal migration and axon/dendrite morphologic maturation by increasing cAMP, generating multiaxonal neurons, and thus phenocopying the effects of increasing cAMP/LKB1 signaling, both in vivo and in vitro (Bony et al., 2013).
Interestingly, localization of LKB1 and its allosteric activator, the pseudokinase STRAD, is enriched selectively in one neurite before axon differentiation, indicating that LKB1 and STRAD predict axonal fate. Moreover, overexpression of LKB1 or STRAD induces the generation of multiaxonal neurons, while downregulation of either LKB1 or STRAD inhibits axon differentiation without affecting overall neurite growth (Barnes et al., 2007; Shelly et al., 2007). LKB1 activation promotes axonal differentiation by activating SAD-A and SAD-B kinases, which phosphorylate the microtubule-associated protein Tau on S262 (Kishi et al., 2005), enabling axonal microtubules to have long labile domains (Baas and Qiang, 2019). Tau also participates in other processes, including regulation of axonal traffic, and anchoring and linking of kinases and phosphatases to the plasma membrane (Biernat et al., 2002).
PKA activation also phosphorylate the E3-ligase Smurf1 on the Thr306 site, changing its substrate affinities, reducing Par6 degradation. Therefore, Par6 accumulates in the future axon allowing Par3/Par6/aPKC complex formation, which plays a crucial role in axon specification and polarity establishment. In addition, Smurf phosphorylation induces RhoA ubiquitination in the axonal compartment, contributing to selective RhoA protein degradation required for localized subcellular function in axonal development (Cheng et al., 2011a) (Fig. 3).
Otherwise, an alternative cAMP-dependent mechanism regulating axonal development was reported, involving the exchange protein directly activated by cAMP (EPAC). The EPAC protein family is composed of EPAC1 and EPAC2, which are GEFs for the small GTPases Rap1 and Rap2, respectively (de Rooij et al., 1998; Kawasaki et al., 1998; Bos, 2006). Epac1 is enriched at the tip of the longest neurite, while Epac2 is distributed in the whole neuron. Treatment with 8-CPT, an Epac-specific activator, or overexpression of Epac1 in hippocampal neurons increased the number of neurons bearing multiple axons through activation of Rap1B. In contrast, inhibition of Epac impairs axonal specification. The effect of Epac1 on neuronal polarization is independent of PKA signaling (Muñoz-Llancao et al., 2015). Previously, it had been reported that Rap1B is a critical regulator of polarization in hippocampal neurons and migrating cortical neurons (Jossin and Cooper, 2011), by RalA activation, which promotes direct exocyst-Par6 interaction, controlling polarized membrane delivery (Nakamura et al., 2013; Das et al., 2014). Rap1B is also necessary to develop axons upstream of Cdc42 and the Par complex, regulating the actin cytoskeleton (Schwamborn and Puschel, 2004). However, 8-pCPT does not induce activation of Cdc42 (Muñoz-Llancao et al., 2015) (Fig. 3). Therefore, further studies are needed to clarify the downstream effectors of Epac1 in neuronal polarity.
cAMP signaling specificity is mainly achieved by confining increases in cAMP concentration to discrete subcellular compartments (Gervasi et al., 2007; Depry et al., 2011). Previous studies show that AC5 is enriched in the distal axon and growth cone, allowing the local increase of cAMP in that region (del Puerto et al., 2012). In contrast, in the soma, PDE4 activity strongly limits the cAMP/PKA signals elicited by stimulation of β-adrenergic receptors (Castro et al., 2010). In addition, the spatial compartmentalization of cAMP is directly controlled by A-kinase anchoring proteins (AKAPs), which act as scaffolds for PKA holoenzyme, ACs, PDEs, phosphatases, and ion channels. The axon-directed cAMP gradient maintained by AKAP-anchored PKA activity is critical for ensuring proper axonal development (Sanderson and Dell'Acqua, 2011; Murphy et al., 2014; Gorshkov et al., 2017). Finally, a local cAMP compartment produced by muscle-specific (m)AKAPα has been reported to promote neuronal survival and axon growth by restricting cAMP signaling to a perinuclear neuronal compartment, suggesting transcriptional regulation mediated by cAMP (Boczek et al., 2019).
It has also been demonstrated that local axonal cAMP elevations induce long-range inhibitory effects on dendritic cAMP levels in hippocampal neurons (Shelly et al., 2010). This effect is mediated by the production of cyclic guanosine monophosphate (cGMP), which is generated by guanylate cyclases (GCs) and degraded by PDEs, similarly to cAMP. High levels of cAMP in a neurite elicit long-range cGMP elevation together with a decrease in cAMP in the other neurites (Shelly et al., 2010). cGMP exerts antagonistic actions. Sema3A application promotes dendrite formation and suppresses axon differentiation after cGMP production, which decreases local cAMP levels through activating PDE4 (Nishiyama et al., 2008; Shelly et al., 2011). Therefore, high levels of cAMP drive axonal elongation and decrease dendritic growth, while high levels of cGMP reduce axonal elongation and increase minor neurite development (Shelly et al., 2010), suggesting that a local balance of cAMP and cGMP could be sufficient to induce preferential initiation of axons and dendrites, respectively. Accordingly, elevating cAMP levels causes a PKA-dependent increase of LKB1 and GSK-3β phosphorylation, whereas cGMP decreases PKA activity and reduces the phosphorylation and stabilization of LKB1. Therefore, cAMP and cGMP signaling acts as antagonistic second messengers for axon/dendrite development, linking second messenger variations with downstream mechanisms controlling axon specification and elongation, such as the LKB1-SAD/MARK pathway (Shelly et al., 2010; Shelly and Poo, 2011) (Fig. 3).
In all, during neuronal polarization, cAMP induces: (1) activation of the LKB1/STRAD/SAD/tau1 signaling pathway, thereby controlling MT-dynamics; (2) Smurf1 phosphorylation, thereby regulating selective protein degradation; and (3) activation of the EPAC/Rap1B signaling pathway, thereby controlling polarized membrane delivery. cAMP also induces a long-range negative signal mediated by increased cGMP levels in the soma and other neurites, which has antagonist action to ensure the correct neuronal polarization.
Hydrogen peroxide
For many years, ROS were only considered toxic byproducts of aerobic energy metabolism that were associated with the onset of several neurodegenerative diseases (Andersen, 2004; Urrutia et al., 2014; Liu et al., 2017). However, ROS can also be essential mediators of normal cellular functions, particularly as second messengers in multiple signal transduction pathways (Cross and Templeton, 2006; Weidinger and Kozlov, 2015; Borquez et al., 2016; Iqbal and Eftekharpour, 2017; Wilson et al., 2018). In neurons, ROS production modulates redox-sensitive features of several molecules, such as transcription factors, signaling proteins, and cytoskeletal components, defining critical aspects of stem cell proliferation and differentiation, neuronal maturation, and neuronal plasticity (Borquez et al., 2016; Wilson et al., 2016a; Iqbal and Eftekharpour, 2017).
The main ROS produced in cells are superoxide anion radicals (O2•−), hydroxyl radicals (•OH), peroxynitrite (ONOO−), and hydrogen peroxide (H2O2). They display different reactivities, concentrations, diffusion velocities, and lifetimes. Therefore, they play different roles in oxidative stress and in signal transduction. •OH and ONOO− are generated by secondary oxidation-reduction reactions of H2O2 and O2•−, respectively, and are highly reactive with proteins, lipids, and DNA, and are thus mainly associated with irreversible oxidative damage. H2O2 is generated by superoxide dismutase activity or spontaneous dismutation from O2•−, and it participates in the oxidation of cysteine thiol side chains, the most characterized reversible redox post-translational modification in cells (Winterbourn, 2008; Dickinson and Chang, 2011). H2O2 is the most stable ROS molecule, has diffusion capacity, and has high selectivity; of the ROS family members, it is thus the best candidate for ROS signaling in cells.
The two most important sources of ROS in cells are the mitochondrion and the NADPH oxidase family (NOX) of redox-active enzymes. Although the primary source of cellular ROS is the mitochondria, its production is not regulated, unlike that of NOX, which is the most studied cellular ROS signaling molecule in many cell types, including neurons (Bedard and Krause, 2007; Nayernia et al., 2014; Wilson et al., 2015, 2016b). Although many studies concentrate on the role of mitochondria in neuronal polarity, these are mostly focused on the production of ATP (Mattson and Partin, 1999; H. J. Kim et al., 2015). Therefore, the importance of mitochondrial ROS in neuronal polarity remains an open question.
The NOX family includes seven membares: NOX1-5 and Dual oxidase (DUOX) 1 and 2. NOX1, NOX2, NOX3, and NOX5 catalyze NADPH-dependent conversion of O2 to O2•− whereas NOX4, DUOX 1 and 2 produce H2O2 (Geiszt et al., 2003; Gough and Cotter, 2011; Takac et al., 2011). Each NOX family member requires several regulatory subunits for activation. NOX2, the best characterized member of this family, is regulated by p40phox (NCF4), p47phox (NCF1), p67phox (NCF2), and Rac1 (Lambeth et al., 2007). This complex interaction involves the exchange of GDP for GTP on Rac1 and the phosphorylation of the p47phox subunit, which allows p47phox to bind with the p22phox/NOX2 complex (Bedard and Krause, 2007) (Fig. 4).
Figure 4.
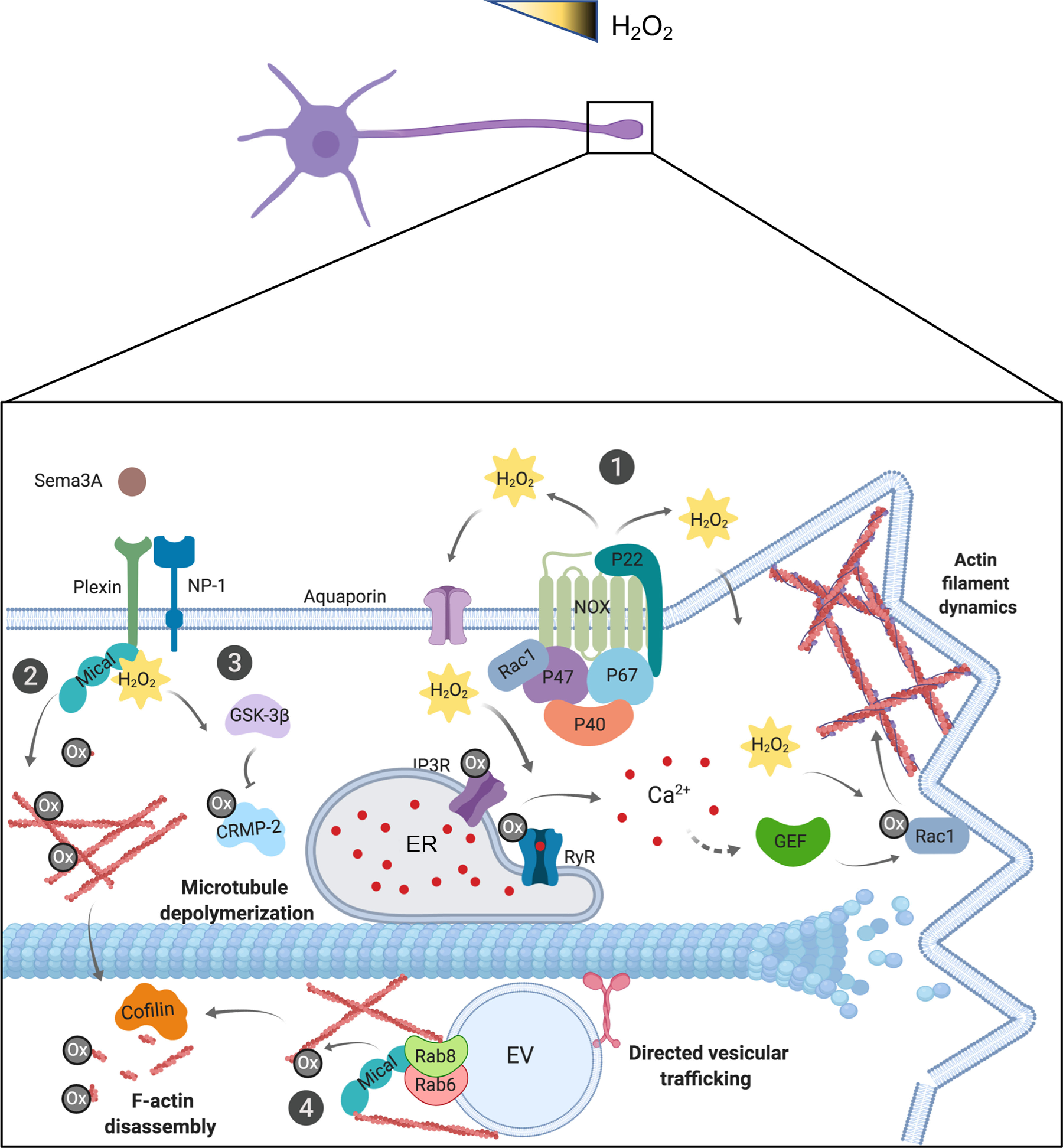
ROS signaling pathways are involved in neuronal polarization. Top, Schematic representation of a proposed ROS gradient in neurons. Bottom, Diagram showing a growth cone. ①, H2O2 production by NOX complex leads to RyR and IP3R activation, producing Ca2+ release, which finally activates Rac1 and NOX complexes, generating a positive loop and enhancing actin filament polymerization. ②, In addition, plexin and NP-1 activate the effector MICAL, which oxidizes G-actin monomers and F-actin, leading to cofilin-mediated F-actin disassembly. ③, MICAL also generates H2O2-mediated oxidation of CRMP-2 promoting its GSK-3β-dependent phosphorylation, inducing MT depolymerization and growth cone collapse. ④, Finally, MICAL could regulate vesicular trafficking by actin disassembly, permitting selective passage of Rab6/Rab8-exocytic vesicles (EV) through the AIS.
NOX1, NOX2, and NOX4 are expressed in the CNS (Serrano et al., 2003; M. J. Kim et al., 2005; Sorce and Krause, 2009), and expression of NOX isoforms has been detected in many neuronal types (Usami et al., 1991; Hilburger et al., 2005; Choi et al., 2012).
Several studies have demonstrated the crucial role played by ROS, particularly H2O2, in neurite outgrowth after treatments with molecules that induce neuronal differentiation in cell lines (Suzukawa et al., 2000; Goldsmit et al., 2001; Min et al., 2006; Nitti et al., 2010; D. S. Kim et al., 2013). However, the first evidence that ROS can regulate neuronal differentiation in neurons was reported in 2009. In this study, using Aplysia neurons treated with pharmacological inhibitors of NOX, it was demonstrated that physiological levels of H2O2 are critical for maintaining a dynamic F-actin cytoskeleton and controlling neurite outgrowth (Munnamalai and Suter, 2009) (Fig. 4). Further studies reported the bidirectional interaction between NOX and the F-actin cytoskeleton in neuronal growth cones. The presence of NOX in this area depended on F-actin and, in turn, NOX regulated F-actin dynamics (Munnamalai et al., 2014).
More recently, it was reported that loss of NOX2 function using genetic and pharmacological strategies alters both neuronal polarity acquisition and axonal growth. These effects were associated with alterations in F-actin dynamics, as found in in Aplysia neurons. Actin changes were associated with a decrease in the global activity of Rac1 and axonal activity of Cdc42 in the neurons (Wilson et al., 2015; Villarroel-Campos et al., 2016; Urrutia et al., 2021) (Fig. 4). Other in vitro studies reported that both ROS and reactive nitrogen species induce the oxidation and glutathionylation of redox-sensitive Rac1 Cys18, promoting nucleotide exchange and Rac activation (Heo and Campbell, 2005; Hobbs et al., 2015). Reduced neurite outgrowth is also reported in cerebellar granule neurons derived from NOX2−/− mice (Olguin-Albuerne and Moran, 2015), in hippocampal neurons derived from p47phox−/− mice (Wilson et at., 2016b), and by pharmacological and genetic inhibition of NOX2 in cultured rat sympathetic neurons (Chandrasekaran et al., 2015).
ROS can also regulate the actin cytoskeleton through G-actin cysteine oxidation by the molecule interacting with CasL (MICAL), a protein effector of the Semaphorin receptor (Terman et al., 2002). Semaphorins exert their effects by binding to cell surface receptors of the Plexin and neuropilin families (Pasterkamp, 2012). The MICAL family of flavoprotein monooxygenases includes MICAL-1, MICAL2, and MICAL-3, whose principal target is actin (Hung et al., 2010, 2011; Giridharan and Caplan, 2014). MICAL oxidizes Met44 and Met47 residues of G-actin monomers, leading to growth cone collapse (Hung et al., 2010). The methionine sulfoxide reductase, MsrB1, can eliminate the oxidative modification in actin induced by MICAL, thus allowing reversible redox regulation of G-actin (Hung et al., 2011; Hung et al., 2013). MICAL can also regulate F-actin stability, as the oxidation of actin improves cofilin-mediated severing, accelerating F-actin disassembly and generating post-translationally modified actin that has altered assembly properties (Grintsevich et al., 2016) (Fig. 4). Additionally, MICAL-2 promotes oxidation-mediated depolymerization of nuclear actin, stimulating transcriptional mechanisms dependent on the serum response factor/myocardin-related transcription factor-A to enhance neurite outgrowth (Lundquist et al., 2014).
MICAL also regulates MT dynamics by H2O2-mediated oxidation of CRMP-2, which generates a cysteine-linked intermediate complex that promotes CRMP-2 phosphorylation by GSK-3β, stimulating MT depolymerization and therefore growth cone collapse (Morinaka et al., 2011) (Fig. 4).
Vesicular traffic can also be regulated by MICAL oxidation. MICAL-3 interacts with Rab8 and regulates the docking and fusion of Rab6/Rab8 exocytic vesicles in a monooxygenase-dependent manner (Grigoriev et al., 2011) controlling trafficking from the trans-Golgi to the plasma membrane. Recently, MICAL3 was found to interact with α2 spectrin in the AIS. In addition, the interaction of MICAL3 with several Rab-GTPases, including Rab8, induces its activation, leading to reduction in the number of AIS actin patches. Therefore, MICAL3 could regulate vesicular trafficking by disassembling actin, permitting the selective passage of several Rab-GTPases-vesicles containing specific axonal cargoes through the AIS (Hamdan et al., 2020) (Fig. 4).
Together, the evidence indicates that physiological hydrogen peroxide levels act as positive signals promoting the establishment of neuronal polarity, neurite growth, and axon specification. The distribution of NOX levels is determined by local compartmentalization of NOX proteins (Fig. 4). Moreover, MICAL-mediated oxidation has multiple roles: it reduces actin stability in the growth cone, controls vesicular traffic by regulating the passage through the AIS of specific vesicles, and stimulates MT depolymerization and growth cone collapse.
Second messenger crosstalk
The coupling of signals associated with more than one second messenger is expected in axon-dendritic polarity (Fig. 5). Extracellular cues, such as Wnt5a and NT-3, induce axon specification by increasing IP3 and activating IP3R, allowing calcium release and activation of CaMKK (Fig. 5, ②) (Li et al., 2009, 2010; Hutchins et al., 2011, 2012; Nakamuta et al., 2011). Extracellular cues can also induce the activation of cAMP-dependent pathways (Fig. 5, ①), which can subsequently induce calcium increases, allowing the coordination between cAMP, calcium, and IP3 signaling pathways (Fig. 5, ①). An additional point of crosstalk involves the activation of Rac1 via a calcium-dependent signaling mechanism, which promotes hydrogen peroxide production, connecting ROS generation with calcium changes in axonal development (Fig. 5, ③). A NOX2 gain of function in primary hippocampal neurons increases axonal and minor neurite length, apparently by inducing RyR2 receptors oxidation that results in Ca2+ release from the ER (Wilson et al., 2016b). In addition, functional coupling between NOX2 and RyR promotes Rac1 activation and axonal elongation, whereby RyR stimulation induces H2O2 production through Rac1-dependent NOX activation (Wilson et al., 2016b). Notably, Rac1 is at the core of mechanisms involved in neuronal polarity since it is a master regulator of actin dynamics and promotes the activation of NOX enzymes (Acevedo and González-Billault, 2018). These results agree with those previously reported regarding the role of calcium gradients in axonal development.
Interestingly, reciprocal interactions between calcium and cAMP signal dynamics are observed in a wide range of non-neuronal cell types (Borodinsky and Spitzer, 2006; Willoughby and Cooper, 2006) (Fig. 5, ①). However, a direct link between cAMP and calcium in the modulation of axon outgrowth and growth cones has not been established so far. In neurons, it is reported that retinal ganglion cell depolarization activates a cAMP/PKA cascade in a calcium-dependent manner, enhancing axonal outgrowth (Dunn et al., 2009). During axon turning, sustained and global manipulations of PKA activity are sufficient to modify the amplitude of fast and local calcium transients dependent on RyR (Stubbs, 1992). Moreover, growth cones stimulated with Netrin-1 (a guidance molecule that modulates both the direction and the speed of axon elongation), which momentarily increases cAMP levels in filopodia, induces filopodial calcium transients following the same dynamics as the cAMP (Nicol et al., 2011). Furthermore, growing neurons in calcium-free culture medium block the Netrin-1–induced increase in cAMP concentration, indicating reciprocal regulation between calcium and cAMP (Nicol et al., 2011). Therefore, spatiotemporal dynamics between calcium and cAMP is a topic that should be extensively explored in polarity development (Fig. 5).
Finally, the coordination between three second messengers in growth cone behavior has been reported. Using DRG neurons, it was demonstrated that endogenous nitric oxide (NO), a reactive nitrogen species, is involved in repulsive axon guidance by activating GCs, increasing activity of the cGMP-PKG pathway, and inactivating the cAMP–PKA pathway over the entire region of the growth cone (Tojima et al., 2009). In addition, there is a counteraction of the NO–cGMP and cAMP pathways that determines the occurrence of RyR-mediated calcium-induced calcium release, thereby switching the direction of growth cone turning, linked with asymmetric Ca2+ elevations. Furthermore, the NO–cGMP–PKG pathway is a negative regulator of RyR-mediated CICR (Tojima et al., 2009). Therefore, a similar mechanism could also participate in axon development.
In conclusion, compelling evidence strongly suggests that multiple second messengers are key players in the local-activation–long-range-inhibition model of single axon generation in polarizing neurons. These molecules allow the coordination of four main molecular events: membrane recruitment, protein transport, an increase in F-actin dynamics, and microtubule assembly.
Ca2+ have a dual role in axonal specification: confined signals of Ca2+ in the growth cone are coupled to the axon elongation machinery, which in turn generates a long-range inhibitory signal, increasing Ca2+ at the soma, ensuring the generation of a single axon (Fig. 5).
This response is mediated by calcium transients of lower frequency, which promotes axon generation by regulating F-actin dynamics and microtubule assembly. At the same time, calcium transients of high-frequency act as long-inhibitory signals via increased RhoA activity, ensuring that only one axon is generated.
cAMP also has a dual role in axonal specification: cAMP signals in the growth cone are coupled to the axon elongation machinery, controlling microtubules dynamics, selective protein degradation, and polarized membrane delivery, and induce a long-range negative signal mediated by cGMP in the soma and other neurites, exerting antagonist action for correct neuronal polarization.
Otherwise, IP3 and ROS are positive signals that promote neuronal polarity, neurite growth, and axon specification. IP3 exerts its action through the generation of calcium transients, whereas ROS regulates actin and microtubule stability and controls directed vesicular traffic.
Hence, all second messengers participate in a highly interconnected network of positive and negative feedback loops that control complex cellular responses, such as the acquisition of neuronal polarity. However, there are still many missing links that need to be addressed. For example, what molecular mechanisms coordinate the timing of local second messenger production? A simultaneous spatiotemporal characterization through newly available molecular sensors and super-resolution microscopy may prove particularly valuable in identifying dynamic patterns of second messenger production that determine neuronal morphology in mechanisms that could be similar to those observed for the coordination of small GTPases in protruding/retraction edge dynamics (Martin et al., 2016).
Footnotes
CONICYT funded this work under the FONDAP Program to Geroscience Center for Brain Health and Metabolism 15150012, Fondecyt 1180419, and 1220414 to C.G.-B., and Fondecyt Postdoctoral 3160630 to P.J.U. The figures were created with www.BioRender.com. We thank Dr. Michael Handford for proofreading this manuscript.
The authors declare no competing financial interests.
References
- Acevedo A, González-Billault C (2018) Crosstalk between Rac1-mediated actin regulation and ROS production. Free Radic Biol Med 116:101–113. 10.1016/j.freeradbiomed.2018.01.008 [DOI] [PubMed] [Google Scholar]
- Akiyama H, Matsu-ura T, Mikoshiba K, Kamiguchi H (2009) Control of neuronal growth cone navigation by asymmetric inositol 1,4,5-trisphosphate signals. Sci Signal 2:ra34. 10.1126/scisignal.2000196 [DOI] [PubMed] [Google Scholar]
- al-Mohanna FA, Cave J, Bolsover SR (1992) A narrow window of intracellular calcium concentration is optimal for neurite outgrowth in rat sensory neurones. Brain Res Dev Brain Res 70:287–290. 10.1016/0165-3806(92)90209-F [DOI] [PubMed] [Google Scholar]
- Andersen JK (2004) Oxidative stress in neurodegeneration: cause or consequence? Nat Med 10 Suppl:S18–S25. 10.1038/nrn1434 [DOI] [PubMed] [Google Scholar]
- Andersen SS, Bi GQ (2000) Axon formation: a molecular model for the generation of neuronal polarity. Bioessays 22:172–179. [DOI] [PubMed] [Google Scholar]
- Arimura N, Kaibuchi K (2007) Neuronal polarity: from extracellular signals to intracellular mechanisms. Nat Rev Neurosci 8:194–205. 10.1038/nrn2056 [DOI] [PubMed] [Google Scholar]
- Arimura N, Kimura T, Nakamuta S, Taya S, Funahashi Y, Hattori A, Shimada A, Menager C, Kawabata S, Fujii K, Iwamatsu A, Segal RA, Fukuda M, Kaibuchi K (2009) Anterograde transport of TrkB in axons is mediated by direct interaction with Slp1 and Rab27. Dev Cell 16:675–686. 10.1016/j.devcel.2009.03.005 [DOI] [PubMed] [Google Scholar]
- Attardo A, Calegari F, Haubensak W, Wilsch-Brauninger M, Huttner WB (2008) Live imaging at the onset of cortical neurogenesis reveals differential appearance of the neuronal phenotype in apical versus basal progenitor progeny. PLoS One 3:e2388. 10.1371/journal.pone.0002388 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baas PW, Qiang L (2019) Tau: it's not what you think. Trends Cell Biol 29:452–461. 10.1016/j.tcb.2019.02.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banker G (2018) The development of neuronal polarity: a retrospective view. J Neurosci 38:1867–1873. 10.1523/JNEUROSCI.1372-16.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes AP, Polleux F (2009) Establishment of axon-dendrite polarity in developing neurons. Annu Rev Neurosci 32:347–381. 10.1146/annurev.neuro.31.060407.125536 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes AP, Lilley BN, Pan YA, Plummer LJ, Powell AW, Raines AN, Sanes JR, Polleux F (2007) LKB1 and SAD kinases define a pathway required for the polarization of cortical neurons. Cell 129:549–563. 10.1016/j.cell.2007.03.025 [DOI] [PubMed] [Google Scholar]
- Batty NJ, Fenrich KK, Fouad K (2017) The role of cAMP and its downstream targets in neurite growth in the adult nervous system. Neurosci Lett 652:56–63. [DOI] [PubMed] [Google Scholar]
- Bedard K, Krause KH (2007) The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev 87:245–313. 10.1152/physrev.00044.2005 [DOI] [PubMed] [Google Scholar]
- Berridge MJ (1998) Neuronal calcium signaling. Neuron 21:13–26. 10.1016/S0896-6273(00)80510-3 [DOI] [PubMed] [Google Scholar]
- Biernat J, Wu YZ, Timm T, Zheng-Fischhofer Q, Mandelkow E, Meijer L, Mandelkow EM (2002) Protein kinase MARK/PAR-1 is required for neurite outgrowth and establishment of neuronal polarity. Mol Biol Cell 13:4013–4028. 10.1091/mbc.02-03-0046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blair LA, Marshall J (1997) IGF-1 modulates N and L calcium channels in a PI 3-kinase-dependent manner. Neuron 19:421–429. 10.1016/S0896-6273(00)80950-2 [DOI] [PubMed] [Google Scholar]
- Boczek T, Cameron EG, Yu W, Xia X, Shah SH, Castillo Chabeco B, Galvao J, Nahmou M, Li J, Thakur H, Goldberg JL, Kapiloff MS (2019) Regulation of neuronal survival and axon growth by a perinuclear cAMP compartment. J Neurosci 39:5466–5480. 10.1523/JNEUROSCI.2752-18.2019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bony G, Szczurkowska J, Tamagno I, Shelly M, Contestabile A, Cancedda L (2013) Non-hyperpolarizing GABAB receptor activation regulates neuronal migration and neurite growth and specification by cAMP/LKB1. Nat Commun 4:1800. 10.1038/ncomms2820 [DOI] [PubMed] [Google Scholar]
- Borodinsky LN, Spitzer NC (2006) Second messenger pas de deux: the coordinated dance between calcium and cAMP. Sci STKE 2006:pe22. 10.1126/stke.3362006pe22 [DOI] [PubMed] [Google Scholar]
- Borquez DA, Urrutia PJ, Wilson C, van Zundert B, Núñez MT, González-Billault C (2016) Dissecting the role of redox signaling in neuronal development. J Neurochem 137:506–517. 10.1111/jnc.13581 [DOI] [PubMed] [Google Scholar]
- Bos JL (2006) Epac proteins: multi-purpose cAMP targets. Trends Biochem Sci 31:680–686. 10.1016/j.tibs.2006.10.002 [DOI] [PubMed] [Google Scholar]
- Bouron A (2020) Transcriptomic profiling of Ca2+ transport systems during the formation of the cerebral cortex in mice. Cells 9:1800. 10.3390/cells9081800 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradke F, Dotti CG (2000) Establishment of neuronal polarity: lessons from cultured hippocampal neurons. Curr Opin Neurobiol 10:574–581. 10.1016/S0959-4388(00)00124-0 [DOI] [PubMed] [Google Scholar]
- Brailoiu E, Hoard JL, Filipeanu CM, Brailoiu GC, Dun SL, Patel S, Dun NJ (2005) Nicotinic acid adenine dinucleotide phosphate potentiates neurite outgrowth. J Biol Chem 280:5646–5650. 10.1074/jbc.M408746200 [DOI] [PubMed] [Google Scholar]
- Buddell T, Quinn CC (2021) An autism-associated calcium channel variant causes defects in neuronal polarity in the ALM neuron of C. elegans. MicroPubl Biol 2021:2021: 10.17912/micropub.biology.000378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caceres A, Ye B, Dotti CG (2012) Neuronal polarity: demarcation, growth and commitment. Curr Opin Cell Biol 24:547–553. 10.1016/j.ceb.2012.05.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calderon de Anda F, Gartner A, Tsai LH, Dotti CG (2008) Pyramidal neuron polarity axis is defined at the bipolar stage. J Cell Sci 121:178–185. 10.1242/jcs.023143 [DOI] [PubMed] [Google Scholar]
- Castro LR, Gervasi N, Guiot E, Cavellini L, Nikolaev VO, Paupardin-Tritsch D, Vincent P (2010) Type 4 phosphodiesterase plays different integrating roles in different cellular domains in pyramidal cortical neurons. J Neurosci 30:6143–6151. 10.1523/JNEUROSCI.5851-09.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan C, Ooashi N, Akiyama H, Fukuda T, Inoue M, Matsu-Ura T, Shimogori T, Mikoshiba K, Kamiguchi H (2020) Inositol 1,4,5-trisphosphate receptor type 3 regulates neuronal growth cone sensitivity to guidance signals. iScience 23:100963. 10.1016/j.isci.2020.100963 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chandrasekaran V, Lea C, Sosa JC, Higgins D, Lein PJ (2015) Reactive oxygen species are involved in BMP-induced dendritic growth in cultured rat sympathetic neurons. Mol Cell Neurosci 67:116–125. 10.1016/j.mcn.2015.06.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y, Beffert U, Ertunc M, Tang TS, Kavalali ET, Bezprozvanny I, Herz J (2005) Reelin modulates NMDA receptor activity in cortical neurons. J Neurosci 25:8209–8216. 10.1523/JNEUROSCI.1951-05.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng PL, Poo MM (2012) Early events in axon/dendrite polarization. Annu Rev Neurosci 35:181–201. 10.1146/annurev-neuro-061010-113618 [DOI] [PubMed] [Google Scholar]
- Cheng PL, Lu H, Shelly M, Gao H, Poo MM (2011a) Phosphorylation of E3 ligase Smurf1 switches its substrate preference in support of axon development. Neuron 69:231–243. 10.1016/j.neuron.2010.12.021 [DOI] [PubMed] [Google Scholar]
- Cheng PL, Song AH, Wong YH, Wang S, Zhang X, Poo MM (2011b) Self-amplifying autocrine actions of BDNF in axon development. Proc Natl Acad Sci USA 108:18430–18435. 10.1073/pnas.1115907108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi DH, Cristovao AC, Guhathakurta S, Lee J, Joh TH, Beal MF, Kim YS (2012) NADPH oxidase 1-mediated oxidative stress leads to dopamine neuron death in Parkinson's disease. Antioxid Redox Signal 16:1033–1045. 10.1089/ars.2011.3960 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ciccolini F, Collins TJ, Sudhoelter J, Lipp P, Berridge MJ, Bootman MD (2003) Local and global spontaneous calcium events regulate neurite outgrowth and onset of GABAergic phenotype during neural precursor differentiation. J Neurosci 23:103–111. 10.1523/JNEUROSCI.23-01-00103.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connor JA (1986) Digital imaging of free calcium changes and of spatial gradients in growing processes in single, mammalian central nervous system cells. Proc Natl Acad Sci USA 83:6179–6183. 10.1073/pnas.83.16.6179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cross JV, Templeton DJ (2006) Regulation of signal transduction through protein cysteine oxidation. Antioxid Redox Signal 8:1819–1827. 10.1089/ars.2006.8.1819 [DOI] [PubMed] [Google Scholar]
- Das A, Gajendra S, Falenta K, Oudin MJ, Peschard P, Feng S, Wu B, Marshall CJ, Doherty P, Guo W, Lalli G (2014) RalA promotes a direct exocyst-Par6 interaction to regulate polarity in neuronal development. J Cell Sci 127:686–699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davare MA, Fortin DA, Saneyoshi T, Nygaard S, Kaech S, Banker G, Soderling TR, Wayman GA (2009) Transient receptor potential canonical 5 channels activate Ca2+/calmodulin kinase Igamma to promote axon formation in hippocampal neurons. J Neurosci 29:9794–9808. 10.1523/JNEUROSCI.1544-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Rooij J, Zwartkruis FJ, Verheijen MH, Cool RH, Nijman SM, Wittinghofer A, Bos JL (1998) Epac is a Rap1 guanine-nucleotide-exchange factor directly activated by cyclic AMP. Nature 396:474–477. [DOI] [PubMed] [Google Scholar]
- del Puerto A, Diaz-Hernandez JI, Tapia M, Gomez-Villafuertes R, Benitez MJ, Zhang J, Miras-Portugal MT, Wandosell F, Diaz-Hernandez M, Garrido JJ (2012) Adenylate cyclase 5 coordinates the action of ADP, P2Y1, P2Y13 and ATP-gated P2X7 receptors on axonal elongation. J Cell Sci 125:176–188. [DOI] [PubMed] [Google Scholar]
- Depry C, Allen MD, Zhang J (2011) Visualization of PKA activity in plasma membrane microdomains. Mol Biosyst 7:52–58. 10.1039/C0MB00079E [DOI] [PubMed] [Google Scholar]
- Dickinson BC, Chang CJ (2011) Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat Chem Biol 7:504–511. 10.1038/nchembio.607 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolmetsch RE, Pajvani U, Fife K, Spotts JM, Greenberg ME (2001) Signaling to the nucleus by an L-type calcium channel-calmodulin complex through the MAP kinase pathway. Science 294:333–339. 10.1126/science.1063395 [DOI] [PubMed] [Google Scholar]
- Dombert B, Balk S, Luningschror P, Moradi M, Sivadasan R, Saal-Bauernschubert L, Jablonka S (2017) BDNF/trkB Induction of calcium transients through Cav2.2 calcium channels in motoneurons corresponds to F-actin assembly and growth cone formation on beta2-chain laminin (221). Front Mol Neurosci 10:346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dotti CG, Banker GA (1987) Experimentally induced alteration in the polarity of developing neurons. Nature 330:254–256. 10.1038/330254a0 [DOI] [PubMed] [Google Scholar]
- Dotti CG, Sullivan CA, Banker GA (1988) The establishment of polarity by hippocampal neurons in culture. J Neurosci 8:1454–1468. 10.1523/JNEUROSCI.08-04-01454.1988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunn TA, Storm DR, Feller MB (2009) Calcium-dependent increases in protein kinase-A activity in mouse retinal ganglion cells are mediated by multiple adenylate cyclases. PLoS One 4:e7877. 10.1371/journal.pone.0007877 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Figge C, Loers G, Schachner M, Tilling T (2012) Neurite outgrowth triggered by the cell adhesion molecule L1 requires activation and inactivation of the cytoskeletal protein cofilin. Mol Cell Neurosci 49:196–204. [DOI] [PubMed] [Google Scholar]
- Funahashi Y, Namba T, Nakamuta S, Kaibuchi K (2014) Neuronal polarization in vivo: growing in a complex environment. Curr Opin Neurobiol 27:215–223. 10.1016/j.conb.2014.04.009 [DOI] [PubMed] [Google Scholar]
- Gao WQ, Hatten ME (1993) Neuronal differentiation rescued by implantation of Weaver granule cell precursors into wild-type cerebellar cortex. Science 260:367–369. [DOI] [PubMed] [Google Scholar]
- Geiszt M, Witta J, Baffi J, Lekstrom K, Leto TL (2003) Dual oxidases represent novel hydrogen peroxide sources supporting mucosal surface host defense. FASEB J 17:1502–1504. 10.1096/fj.02-1104fje [DOI] [PubMed] [Google Scholar]
- Gervasi N, Hepp R, Tricoire L, Zhang J, Lambolez B, Paupardin-Tritsch D, Vincent P (2007) Dynamics of protein kinase A signaling at the membrane, in the cytosol, and in the nucleus of neurons in mouse brain slices. J Neurosci 27:2744–2750. 10.1523/JNEUROSCI.5352-06.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giandomenico SL, Alvarez-Castelao B, Schuman EM (2022) Proteostatic regulation in neuronal compartments. Trends Neurosci 45:41–52. 10.1016/j.tins.2021.08.002 [DOI] [PubMed] [Google Scholar]
- Giridharan SS, Caplan S (2014) MICAL-family proteins: complex regulators of the actin cytoskeleton. Antioxid Redox Signal 20:2059–2073. 10.1089/ars.2013.5487 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldsmit Y, Erlich S, Pinkas-Kramarski R (2001) Neuregulin induces sustained reactive oxygen species generation to mediate neuronal differentiation. Cell Mol Neurobiol 21:753–769. 10.1023/A:1015108306171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez TM, Zheng JQ (2006) The molecular basis for calcium-dependent axon pathfinding. Nat Rev Neurosci 7:115–125. 10.1038/nrn1844 [DOI] [PubMed] [Google Scholar]
- Gomis-Ruth S, Wierenga CJ, Bradke F (2008) Plasticity of polarization: changing dendrites into axons in neurons integrated in neuronal circuits. Curr Biol 18:992–1000. 10.1016/j.cub.2008.06.026 [DOI] [PubMed] [Google Scholar]
- González-Billault C, Muñoz-Llancao P, Henríquez DR, Wojnacki J, Conde C, Caceres A (2012) The role of small GTPases in neuronal morphogenesis and polarity. Cytoskeleton (Hoboken) 69:464–485. 10.1002/cm.21034 [DOI] [PubMed] [Google Scholar]
- Gorshkov K, Mehta S, Ramamurthy S, Ronnett GV, Zhou FQ, Zhang J (2017) AKAP-mediated feedback control of cAMP gradients in developing hippocampal neurons. Nat Chem Biol 13:425–431. 10.1038/nchembio.2298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gough DR, Cotter TG (2011) Hydrogen peroxide: a Jekyll and Hyde signalling molecule. Cell Death Dis 2:e213. 10.1038/cddis.2011.96 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grigoriev I, Yu KL, Martinez-Sanchez E, Serra-Marques A, Smal I, Meijering E, Demmers J, Peranen J, Pasterkamp RJ, van der Sluijs P, Hoogenraad CC, Akhmanova A (2011) Rab6, Rab8, and MICAL3 cooperate in controlling docking and fusion of exocytotic carriers. Curr Biol 21:967–974. 10.1016/j.cub.2011.04.030 [DOI] [PubMed] [Google Scholar]
- Grintsevich EE, Yesilyurt HG, Rich SK, Hung RJ, Terman JR, Reisler E (2016) F-actin dismantling through a redox-driven synergy between Mical and cofilin. Nat Cell Biol 18:876–885. 10.1038/ncb3390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo C, Cho KS, Li Y, Tchedre K, Antolik C, Ma J, Chew J, Utheim TP, Huang XA, Yu H, Malik MTA, Anzak N, Chen DF (2018) IGFBPL1 regulates axon growth through IGF-1-mediated signaling cascades. Sci Rep 8:2054. 10.1038/s41598-018-20463-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamdan H, Lim BC, Torii T, Joshi A, Konning M, Smith C, Palmer DJ, Ng P, Leterrier C, Oses-Prieto JA, Burlingame AL, Rasband MN (2020) Mapping axon initial segment structure and function by multiplexed proximity biotinylation. Nat Commun 11:100. 10.1038/s41467-019-13658-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatanaka Y, Yamauchi K (2013) Excitatory cortical neurons with multipolar shape establish neuronal polarity by forming a tangentially oriented axon in the intermediate zone. Cereb Cortex 23:105–113. 10.1093/cercor/bhr383 [DOI] [PubMed] [Google Scholar]
- Hell JW, Westenbroek RE, Warner C, Ahlijanian MK, Prystay W, Gilbert MM, Snutch TP, Catterall WA (1993) Identification and differential subcellular localization of the neuronal class C and class D L-type calcium channel alpha 1 subunits. J Cell Biol 123:949–962. 10.1083/jcb.123.4.949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heo J, Campbell SL (2005) Mechanism of redox-mediated guanine nucleotide exchange on redox-active Rho GTPases. J Biol Chem 280:31003–31010. 10.1074/jbc.M504768200 [DOI] [PubMed] [Google Scholar]
- Herkert M, Rottger S, Becker CM (1998) The NMDA receptor subunit NR2B of neonatal rat brain: complex formation and enrichment in axonal growth cones. Eur J Neurosci 10:1553–1562. 10.1046/j.1460-9568.1998.00164.x [DOI] [PubMed] [Google Scholar]
- Hevner RF (2006) From radial glia to pyramidal-projection neuron: transcription factor cascades in cerebral cortex development. Mol Neurobiol 33:33–50. [DOI] [PubMed] [Google Scholar]
- Hilburger EW, Conte EJ, McGee DW, Tammariello SP (2005) Localization of NADPH oxidase subunits in neonatal sympathetic neurons. Neurosci Lett 377:16–19. [DOI] [PubMed] [Google Scholar]
- Hobbs GA, Mitchell LE, Arrington ME, Gunawardena HP, DeCristo MJ, Loeser RF, Chen X, Cox AD, Campbell SL (2015) Redox regulation of Rac1 by thiol oxidation. Free Radic Biol Med 79:237–250. 10.1016/j.freeradbiomed.2014.09.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holliday J, Adams RJ, Sejnowski TJ, Spitzer NC (1991) Calcium-induced release of calcium regulates differentiation of cultured spinal neurons. Neuron 7:787–796. 10.1016/0896-6273(91)90281-4 [DOI] [PubMed] [Google Scholar]
- Horigane S, Ageta-Ishihara N, Kamijo S, Fujii H, Okamura M, Kinoshita M, Takemoto-Kimura S, Bito H (2016) Facilitation of axon outgrowth via a Wnt5a-CaMKK-CaMKIalpha pathway during neuronal polarization. Mol Brain 9:8. 10.1186/s13041-016-0189-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hung RJ, Yazdani U, Yoon J, Wu H, Yang T, Gupta N, Huang Z, van Berkel WJ, Terman JR (2010) Mical links semaphorins to F-actin disassembly. Nature 463:823–827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hung RJ, Pak CW, Terman JR (2011) Direct redox regulation of F-actin assembly and disassembly by Mical. Science 334:1710–1713. 10.1126/science.1211956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hung RJ, Spaeth CS, Yesilyurt HG, Terman JR (2013) SelR reverses Mical-mediated oxidation of actin to regulate F-actin dynamics. Nat Cell Biol 15:1445–1454. 10.1038/ncb2871 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchins BI, Kalil K (2008) Differential outgrowth of axons and their branches is regulated by localized calcium transients. J Neurosci 28:143–153. 10.1523/JNEUROSCI.4548-07.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchins BI, Li L, Kalil K (2011) Wnt/calcium signaling mediates axon growth and guidance in the developing corpus callosum. Dev Neurobiol 71:269–283. 10.1002/dneu.20846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchins BI, Li L, Kalil K (2012) Wnt-induced calcium signaling mediates axon growth and guidance in the developing corpus callosum. Sci Signal 5:pt1. [DOI] [PubMed] [Google Scholar]
- Iketani M, Imaizumi C, Nakamura F, Jeromin A, Mikoshiba K, Goshima Y, Takei K (2009) Regulation of neurite outgrowth mediated by neuronal calcium sensor-1 and inositol 1,4,5-trisphosphate receptor in nerve growth cones. Neuroscience 161:743–752. [DOI] [PubMed] [Google Scholar]
- Iqbal MA, Eftekharpour E (2017) Regulatory role of redox balance in determination of neural precursor cell fate. Stem Cells Int 2017:9209127. 10.1155/2017/9209127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang H, Guo W, Liang X, Rao Y (2005) Both the establishment and the maintenance of neuronal polarity require active mechanisms: critical roles of GSK-3beta and its upstream regulators. Cell 120:123–135. [DOI] [PubMed] [Google Scholar]
- Johnson HW, Schell MJ (2009) Neuronal IP3 3-kinase is an F-actin-bundling protein: role in dendritic targeting and regulation of spine morphology. Mol Biol Cell 20:5166–5180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jossin Y, Cooper JA (2011) Reelin, Rap1 and N-cadherin orient the migration of multipolar neurons in the developing neocortex. Nat Neurosci 14:697–703. 10.1038/nn.2816 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamijo S, Ishii Y, Horigane SI, Suzuki K, Ohkura M, Nakai J, Fujii H, Takemoto-Kimura S, Bito H (2018) A critical neurodevelopmental role for L-type voltage-gated calcium channels in neurite extension and radial migration. J Neurosci 38:5551–5566. 10.1523/JNEUROSCI.2357-17.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kater SB, Mattson MP, Cohan C, Connor J (1988) Calcium regulation of the neuronal growth cone. Trends Neurosci 11:315–321. 10.1016/0166-2236(88)90094-X [DOI] [PubMed] [Google Scholar]
- Kawasaki H, Springett GM, Mochizuki N, Toki S, Nakaya M, Matsuda M, Housman DE, Graybiel AM (1998) A family of cAMP-binding proteins that directly activate Rap1. Science 282:2275–2279. 10.1126/science.282.5397.2275 [DOI] [PubMed] [Google Scholar]
- Kim DS, An JM, Lee HG, Seo SR, Kim SS, Kim JY, Kang JW, Bae YS, Seo JT (2013) Activation of Rac1-dependent redox signaling is critically involved in staurosporine-induced neurite outgrowth in PC12 cells. Free Radic Res 47:95–103. 10.3109/10715762.2012.748193 [DOI] [PubMed] [Google Scholar]
- Kim HJ, Shaker MR, Cho B, Cho HM, Kim H, Kim JY, Sun W (2015) Dynamin-related protein 1 controls the migration and neuronal differentiation of subventricular zone-derived neural progenitor cells. Sci Rep 5:15962. 10.1038/srep15962 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim MJ, Shin KS, Chung YB, Jung KW, Cha CI, Shin DH (2005) Immunohistochemical study of p47Phox and gp91Phox distributions in rat brain. Brain Res 1040:178–186. 10.1016/j.brainres.2005.01.066 [DOI] [PubMed] [Google Scholar]
- Kim WY, Zhou FQ, Zhou J, Yokota Y, Wang YM, Yoshimura T, Kaibuchi K, Woodgett JR, Anton ES, Snider WD (2006) Essential roles for GSK-3s and GSK-3-primed substrates in neurotrophin-induced and hippocampal axon growth. Neuron 52:981–996. 10.1016/j.neuron.2006.10.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- King JR, Kabbani N (2018) Alpha 7 nicotinic receptors attenuate neurite development through calcium activation of calpain at the growth cone. PLoS One 13:e0197247. 10.1371/journal.pone.0197247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kishi M, Pan YA, Crump JG, Sanes JR (2005) Mammalian SAD kinases are required for neuronal polarization. Science 307:929–932. 10.1126/science.1107403 [DOI] [PubMed] [Google Scholar]
- Komuro H, Yacubova E, Yacubova E, Rakic P (2001) Mode and tempo of tangential cell migration in the cerebellar external granular layer. J Neurosci 21:527–540. 10.1523/JNEUROSCI.21-02-00527.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kriegstein A, Alvarez-Buylla A (2009) The glial nature of embryonic and adult neural stem cells. Annu Rev Neurosci 32:149–184. 10.1146/annurev.neuro.051508.135600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambeth JD, Kawahara T, Diebold B (2007) Regulation of Nox and Duox enzymatic activity and expression. Free Radic Biol Med 43:319–331. 10.1016/j.freeradbiomed.2007.03.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau PM, Zucker RS, Bentley D (1999) Induction of filopodia by direct local elevation of intracellular calcium ion concentration. J Cell Biol 145:1265–1275. 10.1083/jcb.145.6.1265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lautermilch NJ, Spitzer NC (2000) Regulation of calcineurin by growth cone calcium waves controls neurite extension. J Neurosci 20:315–325. 10.1523/JNEUROSCI.20-01-00315.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leemhuis J, Bouche E, Frotscher M, Henle F, Hein L, Herz J, Meyer DK, Pichler M, Roth G, Schwan C, Bock HH (2010) Reelin signals through apolipoprotein E receptor 2 and Cdc42 to increase growth cone motility and filopodia formation. J Neurosci 30:14759–14772. 10.1523/JNEUROSCI.4036-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Letourneau PC (1996) The cytoskeleton in nerve growth cone motility and axonal pathfinding. Perspect Dev Neurobiol 4:111–123. [PubMed] [Google Scholar]
- Li L, Hutchins BI, Kalil K (2009) Wnt5a induces simultaneous cortical axon outgrowth and repulsive axon guidance through distinct signaling mechanisms. J Neurosci 29:5873–5883. 10.1523/JNEUROSCI.0183-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Hutchins BI, Kalil K (2010) Wnt5a induces simultaneous cortical axon outgrowth and repulsive turning through distinct signaling mechanisms. Sci Signal 3:pt2. [DOI] [PubMed] [Google Scholar]
- Li L, Fothergill T, Hutchins BI, Dent EW, Kalil K (2014) Wnt5a evokes cortical axon outgrowth and repulsive guidance by tau mediated reorganization of dynamic microtubules. Dev Neurobiol 74:797–817. 10.1002/dneu.22102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Z, Zhou T, Ziegler AC, Dimitrion P, Zuo L (2017) Oxidative stress in neurodegenerative diseases: from molecular mechanisms to clinical applications. Oxid Med Cell Longev 2017:2525967. 10.1155/2017/2525967 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lundquist MR, Storaska AJ, Liu TC, Larsen SD, Evans T, Neubig RR, Jaffrey SR (2014) Redox modification of nuclear actin by MICAL-2 regulates SRF signaling. Cell 156:563–576. 10.1016/j.cell.2013.12.035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin K, Reimann A, Fritz RD, Ryu H, Jeon NL, Pertz O (2016) Spatio-temporal co-ordination of RhoA, Rac1 and Cdc42 activation during prototypical edge protrusion and retraction dynamics. Sci Rep 6:21901. 10.1038/srep21901 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattson MP, Kater SB (1987) Calcium regulation of neurite elongation and growth cone motility. J Neurosci 7:4034–4043. 10.1523/JNEUROSCI.07-12-04034.1987 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattson MP, Partin J (1999) Evidence for mitochondrial control of neuronal polarity. J Neurosci Res 56:8–20. [DOI] [PubMed] [Google Scholar]
- Mattson MP, Guthrie PB, Kater SB (1988a) Components of neurite outgrowth that determine neuronal cytoarchitecture: influence of calcium and the growth substrate. J Neurosci Res 20:331–345. [DOI] [PubMed] [Google Scholar]
- Mattson MP, Taylor-Hunter A, Kater SB (1988b) Neurite outgrowth in individual neurons of a neuronal population is differentially regulated by calcium and cyclic AMP. J Neurosci 8:1704–1711. 10.1523/JNEUROSCI.08-05-01704.1988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattson MP, Murain M, Guthrie PB (1990) Localized calcium influx orients axon formation in embryonic hippocampal pyramidal neurons. Brain Res Dev Brain Res 52:201–209. [DOI] [PubMed] [Google Scholar]
- McCaffrey LM, Macara IG (2009) The Par3/aPKC interaction is essential for end bud remodeling and progenitor differentiation during mammary gland morphogenesis. Genes Dev 23:1450–1460. 10.1101/gad.1795909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menager C, Arimura N, Fukata Y, Kaibuchi K (2004) PIP3 is involved in neuronal polarization and axon formation. J Neurochem 89:109–118. [DOI] [PubMed] [Google Scholar]
- Min JY, Park MH, Park MK, Park KW, Lee NW, Kim T, Kim HJ, Lee DH (2006) Staurosporin induces neurite outgrowth through ROS generation in HN33 hippocampal cell lines. J Neural Transm (Vienna) 113:1821–1826. 10.1007/s00702-006-0500-z [DOI] [PubMed] [Google Scholar]
- Morgan JL, Dhingra A, Vardi N, Wong RO (2006) Axons and dendrites originate from neuroepithelial-like processes of retinal bipolar cells. Nat Neurosci 9:85–92. 10.1038/nn1615 [DOI] [PubMed] [Google Scholar]
- Morinaka A, Yamada M, Itofusa R, Funato Y, Yoshimura Y, Nakamura F, Yoshimura T, Kaibuchi K, Goshima Y, Hoshino M, Kamiguchi H, Miki H (2011) Thioredoxin mediates oxidation-dependent phosphorylation of CRMP2 and growth cone collapse. Sci Signal 4:ra26. 10.1126/scisignal.2001127 [DOI] [PubMed] [Google Scholar]
- Munnamalai V, Suter DM (2009) Reactive oxygen species regulate F-actin dynamics in neuronal growth cones and neurite outgrowth. J Neurochem 108:644–661. 10.1111/j.1471-4159.2008.05787.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munnamalai V, Weaver CJ, Weisheit CE, Venkatraman P, Agim ZS, Quinn MT, Suter DM (2014) Bidirectional interactions between NOX2-type NADPH oxidase and the F-actin cytoskeleton in neuronal growth cones. J Neurochem 130:526–540. 10.1111/jnc.12734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muñoz-Llancao P, Henríquez DR, Wilson C, Bodaleo F, Boddeke EW, Lezoualc'h F, Schmidt M, González-Billault C (2015) Exchange protein directly activated by cAMP (EPAC) regulates neuronal polarization through Rap1B. J Neurosci 35:11315–11329. 10.1523/JNEUROSCI.3645-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy JG, Sanderson JL, Gorski JA, Scott JD, Catterall WA, Sather WA, Dell'Acqua ML (2014) AKAP-anchored PKA maintains neuronal L-type calcium channel activity and NFAT transcriptional signaling. Cell Rep 7:1577–1588. 10.1016/j.celrep.2014.04.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura T, Yasuda S, Nagai H, Koinuma S, Morishita S, Goto A, Kinashi T, Wada N (2013) Longest neurite-specific activation of Rap1B in hippocampal neurons contributes to polarity formation through RalA and Nore1A in addition to PI3-kinase. Genes Cells 18:1020–1031. 10.1111/gtc.12097 [DOI] [PubMed] [Google Scholar]
- Nakamuta S, Funahashi Y, Namba T, Arimura N, Picciotto MR, Tokumitsu H, Soderling TR, Sakakibara A, Miyata T, Kamiguchi H, Kaibuchi K (2011) Local application of neurotrophins specifies axons through inositol 1,4,5-trisphosphate, calcium, and Ca2+/calmodulin-dependent protein kinases. Sci Signal 4:ra76. 10.1126/scisignal.2002011 [DOI] [PubMed] [Google Scholar]
- Namba T, Funahashi Y, Nakamuta S, Xu C, Takano T, Kaibuchi K (2015) Extracellular and intracellular signaling for neuronal polarity. Physiol Rev 95:995–1024. 10.1152/physrev.00025.2014 [DOI] [PubMed] [Google Scholar]
- Namba T, Kibe Y, Funahashi Y, Nakamuta S, Takano T, Ueno T, Shimada A, Kozawa S, Okamoto M, Shimoda Y, Oda K, Wada Y, Masuda T, Sakakibara A, Igarashi M, Miyata T, Faivre-Sarrailh C, Takeuchi K, Kaibuchi K (2014) Pioneering axons regulate neuronal polarization in the developing cerebral cortex. Neuron 81:814–829. [DOI] [PubMed] [Google Scholar]
- Naoki H, Ishii S (2014) Mathematical modeling of neuronal polarization during development. Prog Mol Biol Transl Sci 123:127–141. [DOI] [PubMed] [Google Scholar]
- Naoki H, Nakamuta S, Kaibuchi K, Ishii S (2011) Flexible search for single-axon morphology during neuronal spontaneous polarization. PLoS One 6:e19034. 10.1371/journal.pone.0019034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nayernia Z, Jaquet V, Krause KH (2014) New insights on NOX enzymes in the central nervous system. Antioxid Redox Signal 20:2815–2837. 10.1089/ars.2013.5703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicol X, Hong KP, Spitzer NC (2011) Spatial and temporal second messenger codes for growth cone turning. Proc Natl Acad Sci USA 108:13776–13781. 10.1073/pnas.1100247108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieto Guil AF, Oksdath M, Weiss LA, Grassi DJ, Sosa LJ, Nieto M, Quiroga S (2017) IGF-1 receptor regulates dynamic changes in neuronal polarity during cerebral cortical migration. Sci Rep 7:7703. 10.1038/s41598-017-08140-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura T, Yamaguchi T, Kato K, Yoshizawa M, Nabeshima Y, Ohno S, Hoshino M, Kaibuchi K (2005) PAR-6-PAR-3 mediates Cdc42-induced Rac activation through the Rac GEFs STEF/Tiam1. Nat Cell Biol 7:270–277. 10.1038/ncb1227 [DOI] [PubMed] [Google Scholar]
- Nishiyama M, von Schimmelmann MJ, Togashi K, Findley WM, Hong K (2008) Membrane potential shifts caused by diffusible guidance signals direct growth-cone turning. Nat Neurosci 11:762–771. 10.1038/nn.2130 [DOI] [PubMed] [Google Scholar]
- Nitti M, Furfaro AL, Cevasco C, Traverso N, Marinari UM, Pronzato MA, Domenicotti C (2010) PKC delta and NADPH oxidase in retinoic acid-induced neuroblastoma cell differentiation. Cell Signal 22:828–835. 10.1016/j.cellsig.2010.01.007 [DOI] [PubMed] [Google Scholar]
- Noctor SC, Flint AC, Weissman TA, Dammerman RS, Kriegstein AR (2001) Neurons derived from radial glial cells establish radial units in neocortex. Nature 409:714–720. 10.1038/35055553 [DOI] [PubMed] [Google Scholar]
- Noctor SC, Martinez-Cerdeno V, Ivic L, Kriegstein AR (2004) Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat Neurosci 7:136–144. 10.1038/nn1172 [DOI] [PubMed] [Google Scholar]
- Olguin-Albuerne M, Moran J (2015) ROS produced by NOX2 control in vitro development of cerebellar granule neurons development. ASN Neuro 7:175909141557871. 10.1177/1759091415578712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasterkamp RJ (2012) Getting neural circuits into shape with semaphorins. Nat Rev Neurosci 13:605–618. 10.1038/nrn3302 [DOI] [PubMed] [Google Scholar]
- Peters T (1986) Calcium in physiological and pathological cell function. Eur Neurol Suppl 1:27–44. [DOI] [PubMed] [Google Scholar]
- Randlett O, Norden C, Harris WA (2011) The vertebrate retina: a model for neuronal polarization in vivo. Dev Neurobiol 71:567–583. 10.1002/dneu.20841 [DOI] [PubMed] [Google Scholar]
- Rehder V, Kater SB (1992) Regulation of neuronal growth cone filopodia by intracellular calcium. J Neurosci 12:3175–3186. 10.1523/JNEUROSCI.12-08-03175.1992 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reichardt LF (2006) Neurotrophin-regulated signalling pathways. Philos Trans R Soc Lond B Biol Sci 361:1545–1564. 10.1098/rstb.2006.1894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saito A, Miyajima K, Akatsuka J, Kondo H, Mashiko T, Kiuchi T, Ohashi K, Mizuno K (2013) CaMKIIbeta-mediated LIM-kinase activation plays a crucial role in BDNF-induced neuritogenesis. Genes Cells 18:533–543. 10.1111/gtc.12054 [DOI] [PubMed] [Google Scholar]
- Sakakibara A, Sato T, Ando R, Noguchi N, Masaoka M, Miyata T (2014) Dynamics of centrosome translocation and microtubule organization in neocortical neurons during distinct modes of polarization. Cereb Cortex 24:1301–1310. 10.1093/cercor/bhs411 [DOI] [PubMed] [Google Scholar]
- Sanderson JL, Dell'Acqua ML (2011) AKAP signaling complexes in regulation of excitatory synaptic plasticity. Neuroscientist 17:321–336. 10.1177/1073858410384740 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saneyoshi T, Wayman G, Fortin D, Davare M, Hoshi N, Nozaki N, Natsume T, Soderling TR (2008) Activity-dependent synaptogenesis: regulation by a CaM-kinase kinase/CaM-kinase I/betaPIX signaling complex. Neuron 57:94–107. 10.1016/j.neuron.2007.11.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schelski M, Bradke F (2017) Neuronal polarization: from spatiotemporal signaling to cytoskeletal dynamics. Mol Cell Neurosci 84:11–28. 10.1016/j.mcn.2017.03.008 [DOI] [PubMed] [Google Scholar]
- Schwamborn JC, Puschel AW (2004) The sequential activity of the GTPases Rap1B and Cdc42 determines neuronal polarity. Nat Neurosci 7:923–929. 10.1038/nn1295 [DOI] [PubMed] [Google Scholar]
- Serrano F, Kolluri NS, Wientjes FB, Card JP, Klann E (2003) NADPH oxidase immunoreactivity in the mouse brain. Brain Res 988:193–198. 10.1016/s0006-8993(03)03364-x [DOI] [PubMed] [Google Scholar]
- Shelly M, Poo MM (2011) Role of LKB1-SAD/MARK pathway in neuronal polarization. Dev Neurobiol 71:508–527. 10.1002/dneu.20884 [DOI] [PubMed] [Google Scholar]
- Shelly M, Cancedda L, Heilshorn S, Sumbre G, Poo MM (2007) LKB1/STRAD promotes axon initiation during neuronal polarization. Cell 129:565–577. 10.1016/j.cell.2007.04.012 [DOI] [PubMed] [Google Scholar]
- Shelly M, Lim BK, Cancedda L, Heilshorn SC, Gao H, Poo MM (2010) Local and long-range reciprocal regulation of cAMP and cGMP in axon/dendrite formation. Science 327:547–552. [DOI] [PubMed] [Google Scholar]
- Shelly M, Cancedda L, Lim BK, Popescu AT, Cheng PL, Gao H, Poo MM (2011) Semaphorin3A regulates neuronal polarization by suppressing axon formation and promoting dendrite growth. Neuron 71:433–446. 10.1016/j.neuron.2011.06.041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi SH, Jan LY, Jan YN (2003) Hippocampal neuronal polarity specified by spatially localized mPar3/mPar6 and PI 3-kinase activity. Cell 112:63–75. 10.1016/S0092-8674(02)01249-7 [DOI] [PubMed] [Google Scholar]
- Singh TJ, Wang JZ, Novak M, Kontzekova E, Grundke-Iqbal I, Iqbal K (1996) Calcium/calmodulin-dependent protein kinase II phosphorylates tau at Ser-262 but only partially inhibits its binding to microtubules. FEBS Lett 387:145–148. 10.1016/0014-5793(96)00485-1 [DOI] [PubMed] [Google Scholar]
- Song B, Lai B, Zheng Z, Zhang Y, Luo J, Wang C, Chen Y, Woodgett JR, Li M (2010) Inhibitory phosphorylation of GSK-3 by CaMKII couples depolarization to neuronal survival. J Biol Chem 285:41122–41134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sorce S, Krause KH (2009) NOX enzymes in the central nervous system: from signaling to disease. Antioxid Redox Signal 11:2481–2504. 10.1089/ars.2009.2578 [DOI] [PubMed] [Google Scholar]
- Sosa L, Dupraz S, Laurino L, Bollati F, Bisbal M, Caceres A, Pfenninger KH, Quiroga S (2006) IGF-1 receptor is essential for the establishment of hippocampal neuronal polarity. Nat Neurosci 9:993–995. 10.1038/nn1742 [DOI] [PubMed] [Google Scholar]
- Stubbs L (1992) Long-range walking techniques in positional cloning strategies. Mamm Genome 3:127–142. 10.1007/BF00352457 [DOI] [PubMed] [Google Scholar]
- Suzukawa K, Miura K, Mitsushita J, Resau J, Hirose K, Crystal R, Kamata T (2000) Nerve growth factor-induced neuronal differentiation requires generation of Rac1-regulated reactive oxygen species. J Biol Chem 275:13175–13178. 10.1074/jbc.275.18.13175 [DOI] [PubMed] [Google Scholar]
- Suzuki A, Ohno S (2006) The PAR-aPKC system: lessons in polarity. J Cell Sci 119:979–987. 10.1242/jcs.02898 [DOI] [PubMed] [Google Scholar]
- Takac I, Schroder K, Zhang L, Lardy B, Anilkumar N, Lambeth JD, Shah AM, Morel F, Brandes RP (2011) The E-loop is involved in hydrogen peroxide formation by the NADPH oxidase Nox4. J Biol Chem 286:13304–13313. 10.1074/jbc.M110.192138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takano T, Wu M, Nakamuta S, Naoki H, Ishizawa N, Namba T, Watanabe T, Xu C, Hamaguchi T, Yura Y, Amano M, Hahn KM, Kaibuchi K (2017) Discovery of long-range inhibitory signaling to ensure single axon formation. Nat Commun 8:33. 10.1038/s41467-017-00044-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takei K, Shin RM, Inoue T, Kato K, Mikoshiba K (1998) Regulation of nerve growth mediated by inositol 1,4,5-trisphosphate receptors in growth cones. Science 282:1705–1708. 10.1126/science.282.5394.1705 [DOI] [PubMed] [Google Scholar]
- Takemoto-Kimura S, Ageta-Ishihara N, Nonaka M, Adachi-Morishima A, Mano T, Okamura M, Fujii H, Fuse T, Hoshino M, Suzuki S, Kojima M, Mishina M, Okuno H, Bito H (2007) Regulation of dendritogenesis via a lipid-raft-associated Ca2+/calmodulin-dependent protein kinase CLICK-III/CaMKIgamma. Neuron 54:755–770. 10.1016/j.neuron.2007.05.021 [DOI] [PubMed] [Google Scholar]
- Tan M, Ma S, Huang Q, Hu K, Song B, Li M (2013) GSK-3alpha/beta-mediated phosphorylation of CRMP-2 regulates activity-dependent dendritic growth. J Neurochem 125:685–697. [DOI] [PubMed] [Google Scholar]
- Tang F, Dent EW, Kalil K (2003) Spontaneous calcium transients in developing cortical neurons regulate axon outgrowth. J Neurosci 23:927–936. 10.1523/JNEUROSCI.23-03-00927.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terman JR, Mao T, Pasterkamp RJ, Yu HH, Kolodkin AL (2002) MICALs, a family of conserved flavoprotein oxidoreductases, function in plexin-mediated axonal repulsion. Cell 109:887–900. 10.1016/s0092-8674(02)00794-8 [DOI] [PubMed] [Google Scholar]
- Timmermann DB, Westenbroek RE, Schousboe A, Catterall WA (2002) Distribution of high-voltage-activated calcium channels in cultured gamma-aminobutyric acidergic neurons from mouse cerebral cortex. J Neurosci Res 67:48–61. 10.1002/jnr.10074 [DOI] [PubMed] [Google Scholar]
- Toescu EC (2007) Altered calcium homeostasis in old neurons. In: Brain aging: models, methods, and mechanisms (Riddle DR, ed). Boca Raton, FL: CRC. [Google Scholar]
- Tojima T, Takahashi M, Ito E (2003) Dual regulation of LIM kinase 1 expression by cyclic AMP and calcium determines cofilin phosphorylation states during neuritogenesis in NG108-15 cells. Brain Res 985:43–55. 10.1016/s0006-8993(03)03113-5 [DOI] [PubMed] [Google Scholar]
- Tojima T, Itofusa R, Kamiguchi H (2009) The nitric oxide-cGMP pathway controls the directional polarity of growth cone guidance via modulating cytosolic Ca2+ signals. J Neurosci 29:7886–7897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uboha NV, Flajolet M, Nairn AC, Piccioto MR (2007) A calcium- and calmodulin-dependent kinase Ialpha/microtubule affinity regulating kinase 2 signaling cascade mediates calcium-dependent neurite outgrowth. J Neurosci 27:4413–4423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urrutia PJ, Mena NP, Núñez MT (2014) The interplay between iron accumulation, mitochondrial dysfunction, and inflammation during the execution step of neurodegenerative disorders. Front Pharmacol 5:38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urrutia PJ, Bodaleo F, Bórquez DA, Homma Y, Rozes-Salvador V, Villablanca C, Conde C, Fukuda M, González-Billault C (2021) Tuba activates Cdc42 during neuronal polarization downstream of the small GTPase Rab8. J Neurosci 41:1636–1649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Usami M, Nishimatsu S, Ohyanagi H, Shiroiwa H, Takeyama Y, Kasahara H, Saitoh Y (1991) Changes of serum bilirubin fraction in postoperative liver failure cases treated by plasmapheresis: an effective marker for evaluation of bilirubin removal. Artif Organs 15:241–243. [DOI] [PubMed] [Google Scholar]
- Villarroel-Campos D, Henríquez DR, Bodaleo FJ, Oguchi ME, Bronfman FC, Fukuda M, Gonzalez-Billault C (2016) Rab35 functions in axon elongation are regulated by p53-related protein kinase in a mechanism that involves Rab35 protein degradation and the microtubule-associated protein 1B. J Neurosci 36:7298–7313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang PY, Petralia RS, Wang YX, Wenthold RJ, Brenowitz SD (2011) Functional NMDA receptors at axonal growth cones of young hippocampal neurons. J Neurosci 31:9289–9297. 10.1523/JNEUROSCI.5639-10.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Tang JL, Xu X, Zhou XP, Du J, Wang X, Zhou Y, Zhu Q, Yao LL, Wang YG, Hou S, Huang Z (2018) NMDA receptors inhibit axonal outgrowth by inactivating Akt and activating GSK-3beta via calcineurin in cultured immature hippocampal neurons. Exp Cell Res 371:389–398. 10.1016/j.yexcr.2018.08.033 [DOI] [PubMed] [Google Scholar]
- Wayman GA, Kaech S, Grant WF, Davare M, Impey S, Tokumitsu H, Nozaki N, Banker G, Soderling TR (2004) Regulation of axonal extension and growth cone motility by calmodulin-dependent protein kinase I. J Neurosci 24:3786–3794. 10.1523/JNEUROSCI.3294-03.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber JT (2012) Altered calcium signaling following traumatic brain injury. Front Pharmacol 3:60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weidinger A, Kozlov AV (2015) Biological activities of reactive oxygen and nitrogen species: oxidative stress versus signal transduction. Biomolecules 5:472–484. 10.3390/biom5020472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wen Z, Guirland C, Ming GL, Zheng JQ (2004) A CaMKII/calcineurin switch controls the direction of Ca(2+)-dependent growth cone guidance. Neuron 43:835–846. 10.1016/j.neuron.2004.08.037 [DOI] [PubMed] [Google Scholar]
- Willoughby D, Cooper DM (2006) Ca2+ stimulation of adenylyl cyclase generates dynamic oscillations in cyclic AMP. J Cell Sci 119:828–836. 10.1242/jcs.02812 [DOI] [PubMed] [Google Scholar]
- Wilson C, Núñez MT, González-Billault C (2015) Contribution of NADPH oxidase to the establishment of hippocampal neuronal polarity in culture. J Cell Sci 128:2989–2995. [DOI] [PubMed] [Google Scholar]
- Wilson C, Muñoz-Palma E, González-Billault C (2018) From birth to death: a role for reactive oxygen species in neuronal development. Semin Cell Dev Biol 80:43–49. 10.1016/j.semcdb.2017.09.012 [DOI] [PubMed] [Google Scholar]
- Wilson C, Terman JR, González-Billault C, Ahmed G (2016a) Actin filaments: a target for redox regulation. Cytoskeleton (Hoboken) 73:577–595. 10.1002/cm.21315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson C, Muñoz-Palma E, Henríquez DR, Palmisano I, Núñez MT, Di Giovanni S, González-Billault C (2016b) A feed-forward mechanism involving the NOX complex and RyR-mediated Ca2+ release during axonal specification. J Neurosci 36:11107–11119. 10.1523/JNEUROSCI.1455-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winterbourn CC (2008) Reconciling the chemistry and biology of reactive oxygen species. Nat Chem Biol 4:278–286. 10.1038/nchembio.85 [DOI] [PubMed] [Google Scholar]
- Xi F, Xu RJ, Xu JH, Ma JJ, Wang WH, Wang F, Ma YX, Qi SB, Zhang HC, Zhang HN, Qin XZ, Chen JQ, Li B, Liu CM, Yang HL, Meng B, Saijilafu (2019) Calcium/calmodulin-dependent protein kinase II regulates mammalian axon growth by affecting F-actin length in growth cone. J Cell Physiol 234:23053–23065. 10.1002/jcp.28867 [DOI] [PubMed] [Google Scholar]
- Yamamoto H, Yamauchi E, Taniguchi H, Ono T, Miyamoto E (2002) Phosphorylation of microtubule-associated protein tau by Ca2+/calmodulin-dependent protein kinase II in its tubulin binding sites. Arch Biochem Biophys 408:255–262. 10.1016/s0003-9861(02)00556-8 [DOI] [PubMed] [Google Scholar]
- Yano H, Chao MV (2000) Neurotrophin receptor structure and interactions. Pharm Acta Helv 74:253–260. [DOI] [PubMed] [Google Scholar]
- Yi JJ, Barnes AP, Hand R, Polleux F, Ehlers MD (2010) TGF-beta signaling specifies axons during brain development. Cell 142:144–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshimura T, Kawano Y, Arimura N, Kawabata S, Kikuchi A, Kaibuchi K (2005) GSK-3beta regulates phosphorylation of CRMP-2 and neuronal polarity. Cell 120:137–149. 10.1016/j.cell.2004.11.012 [DOI] [PubMed] [Google Scholar]
- Zhang X, Zhu J, Yang GY, Wang QJ, Qian L, Chen YM, Chen F, Tao Y, Hu HS, Wang T, Luo ZG (2007) Dishevelled promotes axon differentiation by regulating atypical protein kinase C. Nat Cell Biol 9:743–754. 10.1038/ncb1603 [DOI] [PubMed] [Google Scholar]
- Zheng JQ, Zheng Z, Poo M (1994) Long-range signaling in growing neurons after local elevation of cyclic AMP-dependent activity. J Cell Biol 127:1693–1701. 10.1083/jcb.127.6.1693 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zolessi FR, Poggi L, Wilkinson CJ, Chien CB, Harris WA (2006) Polarization and orientation of retinal ganglion cells in vivo. Neural Dev 1:2. 10.1186/1749-8104-1-2 [DOI] [PMC free article] [PubMed] [Google Scholar]