

Abstract
Background
Pseudomonas aeruginosa is a common co-infecting pathogen recognized among COVID-19 patients. We aimed to investigate the antimicrobial resistance patterns and molecular typing of Pseudomonas aeruginosa isolates among Coronavirus disease-19 patients.
Methods
Between December 2020 and July 2021, 15 Pseudomonas aeruginosa were isolated from COVID-19 patients in the intensive care unit at Sina Hospital in Hamadan, west of Iran. The antimicrobial resistance of the isolates was determined by disk diffusion and broth microdilution methods. The double-disk synergy method, Modified Hodge test, and polymerase chain reaction were utilized to detect Pseudomonas aeruginosa extended spectrum beta-lactamase and carbapenemase producers. Microtiter plate assay was performed to evaluate the biofilm formation ability of the isolates. The isolates phylogenetic relatedness was revealed using the multilocus variable-number tandem-repeat analysis method.
Results
The results showed Pseudomonas aeruginosa isolates had the most elevated resistance to imipenem (93.3%), trimethoprim-sulfamethoxazole (93.3%), ceftriaxone (80%), ceftazidime (80%), gentamicin (60%), levofloxacin (60%), ciprofloxacin (60%), and cefepime (60%). In the broth microdilution method, 100%, 100%, 20%, and 13.3% of isolates showed resistance to imipenem, meropenem, polymyxin B, and colistin, respectively. Ten (66.6%) isolates were identified as multiple drug resistance. Carbapenemase enzymes and extended spectrum beta-lactamases were identified in 66.6% and 20% of the isolates, respectively and the biofilm formation was detected in 100% of the isolates. The blaOXA-48, blaTEM, blaIMP, blaSPM, blaPER, blaVEB, blaNDM, blaSHV, and blaCTX-M genes were detected in 100%, 86.6%, 86.6%, 40%, 20%, 20%, 13.3%, 6.6%, and 6.6% of the isolates, respectively. The blaVIM, blaGIM, blaGES, and blaMCR-1 genes were not identified in any of the isolates. The MLVA typing technique showed 11 types and seven main clusters and most isolates belong to cluster I, V and VII.
Conclusion
Due to the high rate of antimicrobial resistance, as well as the genetic diversity of Pseudomonas aeruginosa isolates from COVID-19 patients, it is indispensable to monitor the antimicrobial resistance pattern and epidemiology of the isolates on a regular basis.
Keywords: COVID-19, Co-infection, Pseudomonas aeruginosa, Antimicrobial resistance, Biofilm formation, MLVA
Background
The Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) is bringing forth Coronavirus Disease 2019 (COVID-19), giving health systems and clinicians a difficult medical challenge [1]. Hospitalized patients, specifically COVID-19 patients, receive antibiotics without sufficient scientific evidence and clinical experience. Although co-infections between viruses and bacteria can have severe consequences, not much information is available on the coinfection of bacteria with SARS-CoV-2 [2]. COVID-19 patients are affected by a number of bacteria, including P. aeruginosa [3]. P. aeruginosa is an opportunistic organism that causes nosocomial infections (such as pneumonia, urinary tract infections, bloodstream infections, surgical site infections and burn wound infections), and infections in immunocompromised patients (especially neutropenia and malignancy blood) as well as one of the leading causes of disability and death for patients with cystic fibrosis (CF) and non-CF bronchiectasis [4]. In P. aeruginosa, pathogenesis is conducted by adhesions (flagella and type IV pilli), secreted toxins, proteases, effector proteins (such as ExoS, ExoT, ExoU, and ExoY produced by the type III secretion system) and pigments that induce adhesion, regulate or interrupt host cell pathways, and interact with the external matrix. Its ability to cause severe infections is also enhanced by quorum sensing and biofilm formation [5]. Multidrug resistance has increased worldwide, which is considered a threat to public health. Several recent studies have reported the emergence of multidrug-resistant bacterial pathogens from different origins, which increases the need for proper use of antibiotics. In addition, the routine use of antimicrobial susceptibility testing is necessary to detect the antibiotic of choice as well as to screen for emerging MDR strains [6–8].
The rising prevalence of nosocomial infections caused by MDR P. aeruginosa is related to a considerable increase in morbidity and fatality due to limitations in the selection of appropriate antibiotics [9, 10]. The selective treatment of P. aeruginosa-infections is the use of beta-lactam antibiotics [11]. Resistance to the mentioned antibiotics is extending and is done by diverse resistance mechanisms, including the breakdown of antibiotics by β-lactamase enzymes such as Extended-spectrum-β-lactamases (ESBLs), the excretion of antibiotics by efflux pumps, and the reduction of drug absorption [12, 13]. The biofilm formation by P. aeruginosa is responsible for hospital-acquired infections and contributes to persistent colonization in tissues. Biofilms protect bacteria from antibiotics and host immune reactions and contribute to the interchange of resistance genes among microorganisms [14, 15]. Besides routine susceptibility tests to antimicrobial agents, typing pathogenic microorganisms isolated from hospitalized patients, particularly patients with COVID-19, can provide helpful information for physicians. Several genotyping procedures have been applied to investigate the epidemiology and genetic relatedness of P. aeruginosa isolates in the primary phase of infection. Due to the faster, less complicated, and inexpensive Multilocus Variable Number Tandem Repeat Analysis (MLVA) technique, it has become increasingly popular to characterize microorganisms [16, 17]. The foundation of the MLVA method is the proliferation of sequences containing a variable number of tandem repeats (VNTRs) in particular loci on microorganisms' genomes. The difference in the numeral of repetitions from a VNTR allows the strain's discrimination [18]. Therefore, the purpose of this study was to investigate the antibiotic resistance patterns and molecular typing of Pseudomonas aeruginosa strains that were isolated from patients with Covid-19.
Methods
Isolation and identification of P. aeruginosa
From December 2020 to July 2021, fifteen P. aeruginosa were isolated from COVID-19 patients in the intensive care unit (ICU) at Sina Hospital in Hamadan, west of Iran. The collected P. aeruginosa isolates were cultured on blood agar, MacConkey agar and cetrimide agar (Merck, Germany). After incubation for 24 h at 37 ºC, the grown colonies were examined in terms of morphology and pigment production and gram staining. Lactose negative bacteria were purified to perform confirmatory tests. The growth at 42 ºC, oxidase test, TSI, urease, simon citrate and oxidative/fermentative media (Merck, Germany) were used to confirm the P. aeruginosa isolates [19, 20].
Antimicrobial susceptibility testing
The disc diffusion method was accomplished using guided by the Clinical and Laboratory Standards Institute (CLSI) criteria [21]. Briefly, a standardized inoculum was cultured onto the surface of Mueller–Hinton (MH) agar (Merck, Germany) and antibiotic disks were placed on the surface of the agar, and the size of the zone of inhibition around the disk was measured after overnight incubation at 37 °C. The following antibiotic disks (Condalab, Spain) were used: carbapenems (imipenem 10 μg), cephalosporins (cefepime 30 μg, ceftazidime 30 μg, and ceftriaxone 30 μg), fluoroquinolones (ciprofloxacin 5 μg, and levofloxacin 5 μg), aminoglycosides (gentamicin 10 μg), and trimethoprim-sulfamethoxazole (1.25—23.75 μg). P. aeruginosa ATCC 27,853 was used as control positive organism. The MDR, extensively drug-resistant (XDR), and pandrug-resistant (PDR) isolates categorized based on criteria defined by Magiorakos et al. [22]. When performing routine antimicrobial susceptibility testing on bacterial isolates in clinical microbiology laboratories, the limited number of agents generally tested will result in many MDR bacteria being categorized as ‘possible XDR’ or possible PDR’ [22].
The broth microdilution method was conducted to determine the minimum inhibitory concentration (MIC) of the imipenem, meropenem, colistin, and polymyxin B (Sigma-Aldrich/USA) and interpreted as CLSI guidelines [21]. Briefly, the bacterial isolates were inoculated into a MH broth in the presence of different concentrations of an antimicrobial agent and the growth of bacteria was assessed after incubation (16–20 h) at 37 °C and the MIC value was determined.
Phenotypic ESBLs examination
Combination Disk Test (CDT) was applied to identify ESBL-producing isolates using ceftazidime, ceftazidime-clavulanic acid, cefotaxime, and cefotaxime-clavulanic acid disks (Oxoid, UK). After incubation, the difference of > 5 mm in zones of growth inhibition for a disk with clavulanic acid compared to a disc without clavulanic acid is indicative of the presence of ESBLs [23].
Phenotypic carbapenemase examination
The Modified Hodge test (MHT) was conducted according to CLSI guidelines [21]. Briefly, a suspension of the indicator organism of Escherichia coli ATCC 25,922 was prepared and lawn cultured. The meropenem disk (Condalab, Spain) was put in the center of the plate. After that, a colony of the test organisms was inoculated onto the plate and incubated at 37 °C for 18 h. The cloverleaf-like structure indicated the production of carbapenemase.
Biofilm assay
Microtiter plate (MTP) assay evaluated biofilm formation as described previously [24]. Briefly, the isolates were cultured in Luria–Bertani broth (LB) medium (Merck, Germany) overnight and adjusted to 1.5 × 108 CFU/mL, then were diluted 1:100 and inoculated into a 96-well microtiter plate. Each isolate was investigated three times. The un-inoculated LB medium was used as a negative control. Following incubation of the microplate, the wells' contents were discharged and flushed with saline solution. Wells were stained with 0.1% crystal violet (Sigma–Aldrich, St Louis, USA). The crystal violet was aspirated and it’s remaining in the wells was solubilized by adding 95% ethanol (Flucka, Germany). The optical density (OD) of wells was measured at 570 nm and biofilm formation was assayed.
Polymerase chain reaction (PCR) detection of resistance genes
DNA extraction was done from the P. aeruginosa isolates by the salting out method [25]. PCR assay was conducted on all the extracted DNA of the isolates using specific primers (Table 1). Electrophoresis detected amplicons on a 1% agarose (CinnaGen, Iran) gel in TBE (Tris–borate-EDTA) Buffer (CinnaGen, Iran). A 50 bp DNA ladder (MBI Fermentas, France) was utilized for comparisons. A representative for each positive PCR result was sequenced using the Applied Biosystems 3500.
Table 1.
Primer sequences used for genes amplification by PCR
| Genes | Primer Sequences (5’-3’) | Annealing temperature (°C) | Product Size (bp) | Refrences |
|---|---|---|---|---|
| blaOXA-48 |
F-GCGTGGTTAAGGATGAACAC R-CATCAAGTTCAACCCAACCG |
58 | 438 | [26] |
| blaPER |
F-AATTTGGGCTTAGGGCAGAA R-ATGAATGTCATTATAAAAGC |
56 | 925 | [27] |
| blaVEB |
F-CGACTTCCATTTCCCGATGC R-GGACTCTGCAACAAATACGC |
55 | 643 | [28] |
| blaCTX-M |
F-TCTTCCAGAATAAGGAATCCC R-CCGTTTCCGCTATTACAAAC |
55 | 909 | [29] |
| blaTEM |
F-TTTCGTGTCGCCCTTATTCC R-ATCGTTGTCAGAAGTAAGTTGG |
60 | 403 | [30] |
| blaSHV |
F-TCAGCGAAAAACACCTTG R-TCCCGCAGATAAATCACC |
52 | 472 | [31] |
| blaVIM |
F- AGTGGTGAGTATCCGACA R- ATGAAAGTGCGTGGAGAC |
53 | 261 | [27] |
| blaGIM |
F- TCGACACACCTTGGTCTGAA R- AACTTCCAACTTTGCCATGC |
52 | 477 | [32] |
| blaIMP |
F-ACCGCAGCAGAGTCTTTGCC R-ACAACAAGTTTTGCCTTACC |
55 | 587 | [27] |
| blaGES |
F- ATGCGCTTCATTCACGCAC R- CAAAATTTTAAGACGGATCG |
55 | 864 | [27] |
| blaSPM |
F-AAAATCTGGGTACGCAAACG R-ACATTATCCGCTGGAACAGG |
58 | 271 | [33] |
| blaNDM-1 |
F-GGTTTGGCGATCTGGTTTTC R-CGGAATGGCTCATCACGATC |
52 | 621 | [34] |
| blaMCR-1 |
F- CGGTCAGTCCGTTTGTTC R- CTTGGTCGGTCTGTAGGG |
50 | 309 | [35] |
Genotyping
The PCR method based on MLVA was conducted to amplify the VNTRs in the bacterial genome to determine various variants of P. aeruginosa isolates. The VNTR regions (Table 2) were chosen according to Vu-Thien et al. suggestion [36]. PCR products were dissociated in 1% agarose gel (CinnaGen, Iran). A 50 bp ladder (MBI Fermentas, France) was employed to determine the size of the amplicons. The size of the amplicons was analyzed by gel analyzer software [37]. To analyze the clusters, the Unweighted Pair Group Method with Arithmetic (UPGMA) technique was used with the BioNumerics 7.1 software (Applied Maths, Belgium). In addition, calculating the similarity coefficient of Pearson's correlation and the minimum spanning tree (MST) was implemented in BioNumerics 7.1 software (Applied Maths, Belgium). The dissimilarity of the isolates in conforming to the UPGMA algorithm was shown in the dendrogram. The Hunter-Gaston diversity index (HGDI) was utilized to assess the individual or combined VNTR loci polymorphism index. Observing one difference at any VNTRs was considered as a new genotype number. The clustering analysis using the categorical coefficient correlates with an interval of 85 to 100% similarity.
Table 2.
Primer sequences used for multiple VNTRs loci in the PCR reactions
| Locus name | Primer name | Primer Sequences (5’-3’) | Repeat unit size (bp) | HGDI indexa |
|---|---|---|---|---|
| ms142 |
ms142L ms142R |
AGCAGTGCCAGTTGATGTTG GTGGGGCGAAGGAGTGAG |
115 | 0.81 |
| ms211 |
ms111L ms111R |
ACAAGCGCCAGCCGAACCTGT CTTCGAACAGGTGCTGACCGC |
101 | 0.76 |
| ms212 |
ms112L ms112R |
TGCTGGTCGACTACTTCGGCAA ACTACGAGAACGACCCGGTGTT |
40 | 0.75 |
| ms213 |
ms113L ms113R |
CTGGGCAAGTGTTGGTGGATC TGGCGTACTCCGAGCTGATG |
103 | 0.85 |
| ms214 |
ms114L ms114R |
AAACGCTGTTCGCCAACCTCTA CCATCATCCTCCTACTGGGTT |
115 | 0.81 |
| ms215 |
ms115L ms115R |
GACGAAACCCGTCGCGAACA CTGTACAACGCCGAGCCGTA |
129 | 0.80 |
| ms216 |
ms116L ms116R |
ACTACTACGTCGAACACGCCA GATCGAAGACAAGAACCTCG |
113 | 0.64 |
| ms217 |
ms117L ms117R |
TTCTGGCTGTCGCGACTGAT GAACAGCGTCTTTTCCTCGC |
109 | 0.79 |
| ms222 |
ms122L ms122R |
AGAGGTGCTTAACGACGGAT TGCAGTTCTGCGAGGAAGGCG |
101 | 0.76 |
| ms223 |
ms123L ms123R |
TTGGCAATATGCCGGTTCGC TGAGCTGATCGCCTACTGG |
106 | 0.77 |
aHunter-Gaston discriminatory index
Statistical analyses
To analysis the data of this study, descriptive statistics (frequency and percentage) were used, and for this purpose Statistical Package for the Social Sciences (SPSS) version 16 was utilized. BioNumerics 7.1 software was used to analyze MLVA results.
Results
Multidrug resistance P. aeruginosa has increased worldwide, which is considered a threat to public health. Our study investigated the phenotypic and genotypic antimicrobial resistance and molecular typing of P. aeruginosa isolates from Coronavirus disease-19 patients. Overall, 15 clinical isolates of P. aeruginosa were collected from blood cultures and endotracheal aspirates of COVID-19 patients in the ICU. The hypertension (60%), diabetes mellitus (33.3%), and ischemic heart disease (33.3%) were the most prevalent comorbidities. Six (40%) of the patients did not have any underlying disease. Table 3 showed the demographic characteristics of the patients hospitalized in ICU and infected with P. aeruginosa in the COVID-19 pandemic waves in Iran.
Table 3.
Demographic characteristics of patients and the frequency of P. aeruginosa isolated from COVID-19 pandemic waves
| Characteristics | P. aeruginosa isolates No (%) |
|---|---|
| Age groups | |
| 15–25 | 0(0) |
| 26–35 | 1(6.67) |
| 36–45 | 0(0) |
| 46–55 | 3(20) |
| 56–65 | 4(26.67) |
| 66–75 | 5(33.33) |
| 76–85 | 0(0) |
| 86–95 | 1(6.67) |
| 96–105 | 1(6.67) |
| Gender | |
| Male | 10(66.67) |
| Female | 5(33.33) |
| COVID-19 pandemic waves in Iran | |
| Third (November, 2020) | 5(33.33) |
| Fourth (April, 2021) | 4(26.67) |
| Fifth (August, 2021) | 6(40) |
| Underlining disease | |
| 0 | 6(40) |
| 1 | 2(13.33) |
| 1, 2 | 2(13.33) |
| 1, 2, 3 | 3(20) |
| 1, 3 | 1(6.67) |
| 1,3, 4 | 1(6.67) |
| Fate | |
| Deceased | 7(46.67) |
| Discharged | 8(53.33) |
0: No underlying disease; 1: Hypertension; 2: Diabetes; 3: Heart failure; 4: Brain aneurysm
Phenotypic characteristics of P. aeruginosa isolates
Using the biochemical tests, all 15 isolates were confirmed as P. aeruginosa.
Antimicrobial susceptibility testing
The antimicrobial susceptibility of P. aeruginosa isolates were reported as follows: 14(93.3%) isolates resistant to imipenem, 14(93.3%) to co-trimaxazole, 12(80%) to ceftriaxone, 12(80%) to ceftazidime, 9(60%) to gentamicin, 9(60%) to levofloxacin, 9(60%) to ciprofloxacin, and 9(60%) isolates resistant to cefepime. Ten (66.6%) isolates were identified as MDR (Table 4). In the MIC method, 15(100%), 15(100%), 3(20%), and 2(13.3%) P. aeruginosa isolates were resistant to imipenem, meropenem, polymyxin B, and colistin, respectively.
Table 4.
Antimicrobial categories and agents used to define MDR, XDR and PDR P. aeruginosa isolates
| Antimicrobial category | Antimicrobial agent | No (%) of resistant isolates | Type of resistance No (%) | antibiotic-resistance genes | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No (%) of positive ESBL isolates | No (%) of positive Carbapenemase isolates | No (%) of Colistin resistance isolates | ||||||||
| Aminoglycosides | Gentamicin | 9 (60) | Non-MDR | 5 (33.3) | P | EGa | P | CGb | P | MG |
| Carbapenems | Imipenem | 14 (93.3) | ||||||||
| Cephalosporins | Ceftazidime | 12 (80) | MDR | 1 (6.6) | 3 (20) | 3 (100) | 10 (66.6) | 10 (100) | 2 (13.3) | 0 |
| Cefepime | 9 (60) | |||||||||
| Fluoroquinolones | Ciprofloxacin | 9 (60) | ||||||||
| Levofloxacin | 9 (60) | MDR, possible XDR | 8 (53.3) | |||||||
| Penicillins + β-lactamase inhibitors | NT | - | ||||||||
| Monobactams | NT | - | ||||||||
| Phosphonic acids | NT | - | MDR, possible XDR, possible PDR | 1 (6.6) | ||||||
| Polymyxins | Colistin | 2 (13.3) | ||||||||
| Polymyxin B | 3 (20) | |||||||||
NT not tested for susceptibility to antimicrobial agent in this category, P Phenotypic, EG ESBL genes (Positive for at least one gene in phenotypically positive isolates), CG Carbapenemase genes (Positive for at least one gene in phenotypically positive isolates)
aESBL genes (blaTEM, blaSHV, blaPER, blaCTX-M, and blaVEB)
bCarbapenemase genes (blaOXA-48, blaIMP, blaSPM, blaVIM, blaGIM, blaGES, and blaNDM); MG: Colistin resistance gen (blaMCR-1)
MDR: non-susceptible to ≥ 1 agent in ≥ 3 antimicrobial categories
XDR: non-susceptible to ≥ 1 agent in all but ≤ 2 categories
PDR: non-susceptible to all antimicrobial agents listed
ESBLs and carbapenemases producing P. aeruginosa
Detection of carbapenemases by MHT demonstrated that 10(66.6%) P. aeruginosa isolate were positive (Fig. 1) and ESBLs were identified in 3(20%) P. aeruginosa isolates.
Fig. 1.
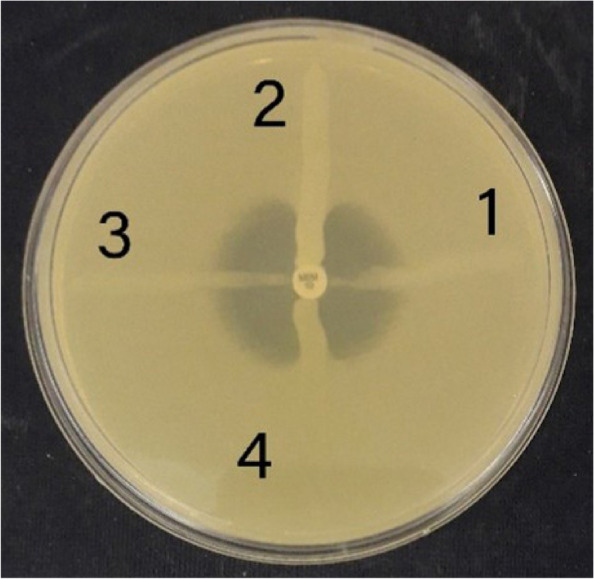
Modified Hodge test; isolates 2 and 4 show positive results, isolates 1 and 3 show negative results
Biofilm formation and quantification
The results demonstrate that 15(100%) P. aeruginosa isolates were positive for biofilm formation, which 11(73.3%) and 4(26.7%) isolates were strong and moderate biofilm producers, respectively.
PCR results
The results of PCR for ESBLs blaTEM, blaPER, blaSHV, and blaCTX-M genes were detected in 13(86.6%), 3(20%), 1(6.6%), and 1(6.6%) of the isolates, respectively. The blaVIM, blaGIM, blaGES, and blaMCR-1 genes were not identified in any of the isolates. The blaOXA-48, blaIMP, blaSPM, blaVEB, and blaNDM were detected in 15(100%), 13(86.6%), 6(40%), 3(20%), and 2(13.3%) of the isolates, respectively. Eleven (73.3%) isolates carried blaOXA-48, blaIMP, and blaTEM genes, and 6 (40%) isolates also carried blaOXA-48, blaTEM, and blaSPM genes, simultaneously.
MLVA typing
The molecular typing of 15 clinical isolates of P. aeruginosa were evaluated by the MLVA method regarding the amplification of ten different VNTR regions. Generally, there were 11 different MLVA types of P. aeruginosa, which the most frequent types belonged to types 6 with 4 isolates, and type 9 with 2 isolates. Eleven various MLVA types of P. aeruginosa isolates were allocated to seven clusters (Fig. 2). The MST algorithm originated from the MLVA genotyping for the clinical isolates of P. aeruginosa shown in Fig. 3.
Fig. 2.

Inferred dendrogram from the clustering analysis of MLVA results of 15 P. aeruginosa isolates using the UPGMA algorithm. A: third wave of COVID-19, B: fourth wave of COVID-19, C: fifth wave of COVID-19, Key: isolates ID, T.A.C: tracheal aspirates culture, B.C: blood culture, ESBL: Extended-spectrum-β-lactamases, MHT: Modified Hodge Test, N: negative, P: positive, St: strong biofilm producer, Mod: moderate biofilm producer, F: female, M: male, None: No underlying disease
Fig. 3.

Minimum Spanning Tree (MST) algorithm obtained from the 15 P. aeruginosa isolates by MLVA. Each circle indicates a single type, the size showing the number of isolates with this specific type. The numbers on a single circle are 100% identical to each other. Thick black lines connecting pairs of MLVA-types display that they differ in one VNTR locus, thin black lines connecting pairs of MLVA-types show that they differ in two VNTR loci, and dashed lines connecting pairs of MLVA-types show that they differ in three VNTR loci. Pink and light green zones surround MLVA-types that belong to the same MLVA clonal complexes (MLVA-CC)
Discussion
COVID-19 patients are particularly prone to superinfection and secondary bacterial infections. However, the pattern of bacterial co-infections associated with COVID-19 and the microbiological profile in these cases are not widely studied [38]. Therefore, it is essential to monitor bacterial co-infection in COVID-19 patients, especially with multi-drug resistant bacteria, to control hospital infections. Antibiotic resistance is a serious crisis that threatens global health and needs urgent action. In prior investigations, resistance to carbapenem varied significantly from 17.5% to 100% [39–41]. A recent study reported that P. aeruginosa isolated from COVID-19 patients showed 100% resistance to ciprofloxacin, levofloxacin, co-trimaxazole, cefotaxime, cefepime, meropenem, and imipenem and 50% resistance to gentamicin [42]. In a recent study done in Iraq by Tizkam et al., P. aeruginosa isolated from COVID-19 patients showed 100% resistance to ceftriaxone and gentamicin, 48.9% and 50% resistance to levofloxacin and meropenem, respectively [43]. Based on Jamnani et al. study, P.aeruginosa isolated from ICU-admitted COVID-19 patients was 100% resistant to cefixime and co-trimaxazole, 50% resistant to ciprofloxacin, 25% resistant to gentamycin and colistin [44]. In this study, colistin (86.6%) showed the highest rate of susceptibility, similar to prior investigations [45, 46]. It can be attributed to factors such as the high cost of colistin and its limited use outside hospitals. Discrepancies in antibiotic susceptibility patterns between isolates in various countries can be explained by the source of isolates, the rise of empiric antibiotic use, the existence or lack of antibiotic use supervision schedules, horizontal gene transfer, and discrepancies in the region's epidemiology.
Multiple mechanisms of resistance to antibiotics exist in bacteria, including reduced permeability, expression of efflux pumps, generation of antibiotic-inactivating enzymes, and target modifications. Most of these resistance mechanisms are present in P. aeruginosa. By producing these mechanisms simultaneously, MDR, XDR, and PDR strains emerge [47]. A recent study showed the most abundant species of bacteria isolated from severe COVID-19 patients were P. aeruginosa (39.5%), and 64.7% of these isolates also were multi-drug-resistant strains [48]. Another study detected MDR isolates in 64.5% of COVID-19 patients [49]. Typical resistance mechanisms in pathogens isolated from COVID-19 patients have rarely been examined. Similarly, low production (27.3%) of ESBL was reported by Dutta et al., while Farhan et al. study presented ESBL production in 54% MDR P. aeruginosa [50, 51]. In the other study, the production rate of ESBL and carbapenemase enzymes was stated at 24.7% and 25.7% in P.aeruginosa isolates, respectively [52]. The emergence and rapid spread of β-lactamase enzymes producing bacteria are of serious concern and threat. Thus, it is essential to examine β-lactamase enzymes producing isolates.
Hence, for COVID-19 subjects that exhibit co-infections with other respiratory infections, the immediate administration of antimicrobial agents relevant to the Antibiotic sensitivity test results and also accurate application of infection control protocols are required to alleviate mortality and hospital spread [53–55]. Moreover, the isolates were assessed for the presence of several resistance genes. A recent report indicated that the gram-negative clinical isolates from COVID-19 Patients were mostly multidrug-resistant and ESBL and/or carbapenemase producers and carried different resistance-associated genes, including blaNDM-1, blaTEM, blaCTX-M, and blaSHV. According to their results, 100% of P. aeruginosa isolates carried blaNDM-1 and blaTEM, and no isolates had blaCTX-M and blaSHV [56]. In contrast to our study, in Farhan et al. Study, blaCTX-M15 was detected in 55.5% positive ESBL P. aeruginosa, and blaIMP, blaVIM, and blaGIM were found in 42.8%, 52.3%, and 52.3% of carbapenem-resistant P. aeruginosa, respectively. In a study in Iraq, out of 20 P. aeruginosa isolates isolated from Covid-19 patients hospitalized in ICU, 16 (80%) isolates were positive for blaCTX-M,but blaSHV and blaTEM were not found in any of the tested isolates [57]. Furthermore, Similar to our findings, reported for incidence of blaSPM (38%) among P. aeruginosa isolates [51]. Ahmed et al. reprted a low prevalence of positive blaCTX-M P. aeruginosa isolates(10.7%), which is consistent with the results of our study [58]. In contrast to current study, Tawfik et al. reported 68% and 20% the prevalence of blaVEB, and blaGES in P. aeruginosa, respectively. Also, ESBLs blaTEM, blaSHV, blaPER, and blaCTX-M genes were not reported [59]. Based on the findings of Bianco et al., among the 1242 clinical isolates of Enterobacterales during the Covid-19 epidemic, 1034 (83.2%), 114 (9.2%), 53 (4.3%) and 51 (4.1%) isolates were positive for KPC, blaVIM, blaOXA-48 and blaNDM, respectively [60]. According to Miftode et al. Study, 72 (82.7%) of the 87 Enterobacterales isolated from covid-19 patients produced carbapenemases and 26 (36.1%), 25 (34.7%), 13 (18%), and 2 (2.7%) isolates were positive for blaOXA-48, blaNDM, KPC, and blaVIM, respectively [61]. However, resistance genes rate vary significantly between various studies, which can lead to diversity in infection management guidelines. Prior studies showed that viral infections, including COVID-19, promote bacterial biofilm formation [62–64]. In recent studies, 92.7% and 94% P. aeruginosa isolates were reported as biofilm producers [39, 65]. Typing methods have a substantial role in comprehending the epidemiology relevant to severe nosocomial infections caused by P. aeruginosa [66]. Detection of diverse strains of various bacterial types is necessary for the investigation of the prevalence and control of bacterial infections [67]. Regarding the great genetic variety of MDR P. aeruginosa isolates, especially seen in ICU, implementing appropriate infection management procedures is challenging. In the present study, the amplification of the ten distinct VNTR loci showed all studied P. aeruginosa isolates were typeable. In contrast, Lalancette et al. study reported three not-typable strains [68]. Regarding observed high genetic diversity among P. aeruginosa isolates, separated from COVID-19 patients, it is essential the continuous monitoring of the molecular epidemiology of P. aeruginosa isolates in the COVID-19 epidemic. The current study has some limitations. We have no information about COVID-19 patients who did not develop co-infections (control patients), the type and number of antimicrobials prescribed during the COVID-19 pandemic, and the length of hospitalization. We suggest that subsequent work comprise such information to allow comparative analysis.
Conclusions
At the beginning of the Covid-19 pandemic, the existing guidelines for COVID-19 patients did not include specific recommendations for the use of antibiotics or specific management measures to prevent nosocomial infections in these patients. Due to the excessive use of antibiotics during the pandemic, there was a significant increase in antibiotic resistance. It was worried that the existent overuse of antimicrobial agents during the COVID-19 pandemic could accelerate the emersion of the subsequent global public health crisis caused by the resistance of microorganisms to a variety of drugs. Due to the high rate of antimicrobial resistance, as well as the genetic diversity of Pseudomonas aeruginosa isolates from COVID-19 patients, the current study emphasizes the significance of monitoring local epidemiology, which might be helpful in antimicrobial agents use and surveillance programs.
Acknowledgements
The authors express their gratitude to the Vice-chancellor of Research of Hamadan University of Medical Sciences, Hamadan, Iran, for advocating this research.
Abbreviations
- COVID-19
Coronavirus disease 2019
- MLVA
Multilocus variable number tandem repeat analysis
- ICU
Intensive care unit
- MDR
Multidrug-resistant
- SARS-CoV-2
Severe acute respiratory syndrome coronavirus 2
- VNTRs
Variable number of tandem repeats
- ESBLs
Extended-spectrum-β-lactamases
- CLSI
Clinical and Laboratory Standards Institute
- MH
Mueller–Hinton
- MIC
Minimum inhibitory concentration
- CDT
Combination disk test
- MHT
Modified Hodge test
- LB
Luria–Bertani broth
- MTP
Microtiter plates
- HGDI
Hunter-Gaston discriminatory index
- UPGMA
Unweighted Pair Group Method with Arithmetic
- MST
Minimum spanning tree
Authors’ contributions
MYA designed and supervised the study. MYA and MA performed data interpretation. MA, SS, MSA, PK, and ZR were responsible for data collection and doing experiments. FK, SHH and MMM performed clinical examination. All authors approved the final version of the manuscript.
Funding
This study was funded by the Hamadan University of medical sciences, Hamadan, IRAN, under Grant numbers: 140012031272, 14000124434 and 140111049570.
Availability of data and materials
Data is available from the corresponding author upon request.
Declarations
Ethics approval and consent to participate
All experimental protocols were approved by the Ethic comity of the Hamadan University of Medical sciences, Hamadan, Iran (Ethic approval codes: IR.UMSHA.REC.1400.876, IR.UMSHA.REC.1399.1059 and IR.UMSHA.REC.1401.716). All methods were carried out in accordance with relevant guidelines and regulations. Ethical Review Board approved informed consent taken from all the participants and their legal guardians.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflicts of interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Zhou F, Yu T, Du R, Fan G, Liu Y, Liu Z, et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. The lancet. 2020;395(10229):1054–1062. doi: 10.1016/S0140-6736(20)30566-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Abelenda-Alonso G, Rombauts A, Gudiol C, Meije Y, Ortega L, Clemente M, et al., editors. Influenza and bacterial coinfection in adults with community-acquired pneumonia admitted to conventional wards: risk factors, clinical features, and outcomes. Open Forum Infectious Diseases; 2020: Oxford University Press US. [DOI] [PMC free article] [PubMed]
- 3.Lansbury L, Lim B, Baskaran V, Lim WS. Co-infections in people with COVID-19: a systematic review and meta-analysis. J Infect. 2020;81(2):266–275. doi: 10.1016/j.jinf.2020.05.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Alhazmi A. Pseudomonas aeruginosa-pathogenesis and pathogenic mechanisms. Int J Biol. 2015;7(2):44. [Google Scholar]
- 5.Reynolds D, Kollef M. The epidemiology and pathogenesis and treatment of Pseudomonas aeruginosa infections: an update. Drugs. 2021;81(18):2117–2131. doi: 10.1007/s40265-021-01635-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Elbehiry A, Marzouk E, Aldubaib M, Moussa I, Abalkhail A, Ibrahem M, et al. Pseudomonas species prevalence, protein analysis, and antibiotic resistance: an evolving public health challenge. AMB Express. 2022;12(1):1–14. doi: 10.1186/s13568-022-01390-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Algammal AM, Mabrok M, Sivaramasamy E, Youssef FM, Atwa MH, El-Kholy AW, et al. Emerging MDR-Pseudomonas aeruginosa in fish commonly harbor oprL and toxA virulence genes and blaTEM, blaCTX-M, and tetA antibiotic-resistance genes. Sci Rep. 2020;10(1):1–12. doi: 10.1038/s41598-020-72264-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Algammal AM, Ibrahim RA, Alfifi KJ, Ghabban H, Alghamdi S, Kabrah A, et al. A first report of molecular typing, virulence traits, and phenotypic and genotypic resistance patterns of newly emerging XDR and MDR aeromonas veronii in Mugil seheli. Pathogens. 2022;11(11):1262. doi: 10.3390/pathogens11111262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lister PD, Wolter DJ, Hanson ND. Antibacterial-resistant Pseudomonas aeruginosa: clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin Microbiol Rev. 2009;22(4):582–610. doi: 10.1128/CMR.00040-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zarei O, Shokoohizadeh L, Hossainpour H, Alikhani MY. Molecular analysis of Pseudomonas aeruginosa isolated from clinical, environmental and cockroach sources by ERIC-PCR. BMC Res Notes. 2018;11(1):1–7. doi: 10.1186/s13104-018-3765-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ibrahim D, Jabbour J-F, Kanj SS. Current choices of antibiotic treatment for Pseudomonas aeruginosa infections. Curr Opin Infect Dis. 2020;33(6):464–473. doi: 10.1097/QCO.0000000000000677. [DOI] [PubMed] [Google Scholar]
- 12.Pfeifer Y, Cullik A, Witte W. Resistance to cephalosporins and carbapenems in Gram-negative bacterial pathogens. Int J Med Microbiol. 2010;300(6):371–379. doi: 10.1016/j.ijmm.2010.04.005. [DOI] [PubMed] [Google Scholar]
- 13.Zilberberg MD, Jacobsen T, Tillotson G, editors. The burden of hospitalizations and emergency department visits with hereditary angioedema and angioedema in the United States, 2007. Allergy & Asthma Proceedings; 2010. [DOI] [PubMed]
- 14.Stepanović S, Vuković D, Hola V, Bonaventura GD, Djukić S, Ćirković I, et al. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS. 2007;115(8):891–899. doi: 10.1111/j.1600-0463.2007.apm_630.x. [DOI] [PubMed] [Google Scholar]
- 15.Ghorbanzadeh R, Assadian H, Chiniforush N, Parker S, Pourakbari B, Ehsani B, et al. Modulation of virulence in Enterococcus faecalis cells surviving antimicrobial photodynamic inactivation with reduced graphene oxide-curcumin: An ex vivo biofilm model. Photodiagn Photodyn Ther. 2020;29:101643. doi: 10.1016/j.pdpdt.2019.101643. [DOI] [PubMed] [Google Scholar]
- 16.Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, et al. Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28(12):1647–1649. doi: 10.1093/bioinformatics/bts199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pitman TL, Philbrook RN, Vetterli MR, Warren JG. First report of Pythium ultimum causing crown rot in greenhouse-grown Cannabis sativa in California. Plant Dis. 2021;105(4):1230. doi: 10.1094/PDIS-10-20-2228-PDN. [DOI] [PubMed] [Google Scholar]
- 18.Larsson J, Torpdahl M, Petersen R, Sørensen G, Lindstedt B, Nielsen E. Development of a new nomenclature for Salmonella Typhimurium multilocus variable number of tandem repeats analysis (MLVA) Eurosurveillance. 2009;14(15):19174. doi: 10.2807/ese.14.15.19174-en. [DOI] [PubMed] [Google Scholar]
- 19.Koneman EW, Allen SD, Janda W, Schreckenberger P, Winn W. Diagnostic microbiology. The nonfermentative gram-negative bacilli Philedelphia: Lippincott-Raven Publishers; 1997. pp. 253–320. [Google Scholar]
- 20.Parte A, Whitman WB, Goodfellow M, Kämpfer P, Busse H-J, Trujillo ME, et al. Bergey's manual of systematic bacteriology: volume 5: the Actinobacteria: Springer Science & Business Media; 2012.
- 21.Clinical and Laboratory Standards Institute . M100: performance standards for antimicrobial susceptibility testing. 31. Washington: CLSI; 2021. [Google Scholar]
- 22.Magiorakos A-P, Srinivasan A, Carey RB, Carmeli Y, Falagas M, Giske C, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. doi: 10.1111/j.1469-0691.2011.03570.x. [DOI] [PubMed] [Google Scholar]
- 23.Salihu M, Yarima A, Atta H. Methods for the phenotypic detection of extended spectrum beta lactamase-producing bacteria. Niger J Biotechnol. 2020;37(2):113–125. [Google Scholar]
- 24.Ramos-Vivas J, Chapartegui-González I, Fernández-Martínez M, González-Rico C, Fortún J, Escudero R, et al. Biofilm formation by multidrug resistant Enterobacteriaceae strains isolated from solid organ transplant recipients. Sci Rep. 2019;9(1):1–10. doi: 10.1038/s41598-019-45060-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dairawan M, Shetty PJ. The evolution of DNA extraction methods. Am J Biomed Sci Res. 2020;8:39–45. doi: 10.34297/AJBSR.2020.08.001234. [DOI] [Google Scholar]
- 26.Poirel L, Bonnin RA, Nordmann P. Analysis of the resistome of a multidrug-resistant NDM-1-producing Escherichia coli strain by high-throughput genome sequencing. Antimicrob Agents Chemother. 2011;55(9):4224–4229. doi: 10.1128/AAC.00165-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shahcheraghi F, Nobari S, Rahmati Ghezelgeh F, Nasiri S, Owlia P, Nikbin VS, et al. First report of new Delhi metallo-beta-lactamase-1-producing Klebsiella pneumoniae in Iran. Microb Drug Resist. 2013;19(1):30–36. doi: 10.1089/mdr.2012.0078. [DOI] [PubMed] [Google Scholar]
- 28.Naas T, Bogaerts P, Bauraing C, Degheldre Y, Glupczynski Y, Nordmann P. Emergence of PER and VEB extended-spectrum β-lactamases in Acinetobacter baumannii in Belgium. J Antimicrob Chemother. 2006;58(1):178–182. doi: 10.1093/jac/dkl178. [DOI] [PubMed] [Google Scholar]
- 29.Stürenburg E, Kühn A, Mack D, Laufs R. A novel extended-spectrum β-lactamase CTX-M-23 with a P167T substitution in the active-site omega loop associated with ceftazidime resistance. J Antimicrob Chemother. 2004;54(2):406–409. doi: 10.1093/jac/dkh334. [DOI] [PubMed] [Google Scholar]
- 30.Bali EB, Acik L, Sultan N. Phenotypic and molecular characterization of SHV, TEM, CTX-M and extended-spectrum beta-lactamase produced by Escherichia coli, Acinobacter baumannii and Klebsiella isolates in a Turkish hospital. Afr J Microbiol Res. 2010;4(8):650–654. [Google Scholar]
- 31.Lal P, Kapil A, Das BK, Sood S. Occurrence of TEM & SHV gene in extended spectrum b-lactamases (ESBLs) producing Klebsiella sp. isolated from a tertiary care hospital. Indian J Med Res. 2007;125(2):173. [PubMed] [Google Scholar]
- 32.Aksoy MD, Çavuşlu Ş, Tuğrul HM. Investigation of metallo beta lactamases and oxacilinases in carbapenem resistant Acinetobacter baumannii strains isolated from inpatients. Balkan Med J. 2015;32(1):79–83. doi: 10.5152/balkanmedj.2015.15302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Poirel L, Walsh TR, Cuvillier V, Nordmann P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn Microbiol Infect Dis. 2011;70(1):119–123. doi: 10.1016/j.diagmicrobio.2010.12.002. [DOI] [PubMed] [Google Scholar]
- 34.Khorsi K, Messai Y, Hamidi M, Ammari H, Bakour R. High prevalence of multidrug-resistance in Acinetobacter baumannii and dissemination of carbapenemase-encoding genes blaOXA-23-like, blaOXA-24-like and blaNDM-1 in Algiers hospitals. Asian Pac J Trop Med. 2015;8(6):438–446. doi: 10.1016/j.apjtm.2015.05.011. [DOI] [PubMed] [Google Scholar]
- 35.Liu Y-Y, Wang Y, Walsh TR, Yi L-X, Zhang R, Spencer J, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi: 10.1016/S1473-3099(15)00424-7. [DOI] [PubMed] [Google Scholar]
- 36.Vu-Thien H, Corbineau G, Hormigos K, Fauroux B, Corvol H, Clément A, et al. Multiple-locus variable-number tandem-repeat analysis for longitudinal survey of sources of Pseudomonas aeruginosa infection in cystic fibrosis patients. J Clin Microbiol. 2007;45(10):3175–3183. doi: 10.1128/JCM.00702-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Shirani K, Ataei B, Roshandel F. Antibiotic resistance pattern and evaluation of metallo-beta lactamase genes (VIM and IMP) in Pseudomonas aeruginosa strains producing MBL enzyme, isolated from patients with secondary immunodeficiency. Adv Biomed Res. 2016;5:124. doi: 10.4103/2277-9175.186986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Senok A, Alfaresi M, Khansaheb H, Nassar R, Hachim M, Al Suwaidi H, et al. Coinfections in patients hospitalized with COVID-19: a descriptive study from the United Arab Emirates. Infect Drug Resist. 2021;14:2289. doi: 10.2147/IDR.S314029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.El-Mahdy R, El-Kannishy G. Virulence factors of carbapenem-resistant Pseudomonas aeruginosa in hospital-acquired infections in Mansoura, Egypt. Infect Drug Resist. 2019;12:3455. doi: 10.2147/IDR.S222329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rouhi S, Ramazanzadeh R. Prevalence of blaOxacillinase-23and blaOxacillinase-24/40-type Carbapenemases in Pseudomonas aeruginosa species isolated from patients with nosocomial and non-nosocomial infections in the West of Iran. Iran J Pathol. 2018;13(3):348. [PMC free article] [PubMed] [Google Scholar]
- 41.Kresken M, Körber-Irrgang B, Korte-Berwanger M, Pfennigwerth N, Gatermann SG, Seifert H, et al. Dissemination of carbapenem-resistant Pseudomonas aeruginosa isolates and their susceptibilities to ceftolozane-tazobactam in Germany. Int J Antimicrob Agents. 2020;55(6):105959. doi: 10.1016/j.ijantimicag.2020.105959. [DOI] [PubMed] [Google Scholar]
- 42.Ramadan HK-A, Mahmoud MA, Aburahma MZ, Elkhawaga AA, El-Mokhtar MA, Sayed IM, et al. Predictors of severity and co-infection resistance profile in COVID-19 patients: first report from upper Egypt. Infect Drug Resist. 2020;13:3409. doi: 10.2147/IDR.S272605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tizkam HH, Fadhil OQ, Ghazy E. Effect of COVID-19 on bacterial resistance. Sys Rev Pharm. 2020;11(11):423–427. [Google Scholar]
- 44.Jamnani AN, Montazeri M, Mirzakhani M, Moosazadeh M, Haghighi M. Evaluation of bacterial coinfection and antibiotic resistance in patients with COVID-19 under mechanical ventilation. SN Compr Clin Med. 2022;4(1):1–5. doi: 10.1007/s42399-021-01114-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Baniya B, Pant ND, Neupane S, Khatiwada S, Yadav UN, Bhandari N, et al. Biofilm and metallo beta-lactamase production among the strains of Pseudomonas aeruginosa and Acinetobacter spp. at a Tertiary Care Hospital in Kathmandu, Nepal. Ann Clin Microbiol Antimicrob. 2017;16(1):1–4. doi: 10.1186/s12941-017-0245-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Heidari R, Farajzadeh Sheikh A, Hashemzadeh M, Farshadzadeh Z, Salmanzadeh S, Saki M. Antibiotic resistance, biofilm production ability and genetic diversity of carbapenem-resistant Pseudomonas aeruginosa strains isolated from nosocomial infections in southwestern Iran. Mol Biol Rep. 2022;49(5):3811–22. [DOI] [PMC free article] [PubMed]
- 47.Bassetti M, Vena A, Croxatto A, Righi E, Guery B. How to manage Pseudomonas aeruginosa infections. Drugs Context. 2018;7:212527. doi: 10.7573/dic.212527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mazzariol A, Benini A, Unali I, Nocini R, Smania M, Bertoncelli A, et al. Dynamics of SARS-CoV2 infection and multi-drug resistant bacteria superinfection in patients with assisted mechanical ventilation. Front Cell Infect Microbiol. 2021;11:683409. [DOI] [PMC free article] [PubMed]
- 49.Temperoni C, Caiazzo L, Barchiesi F. High prevalence of antibiotic resistance among opportunistic pathogens isolated from patients with COVID-19 under mechanical ventilation: results of a single-center study. Antibiotics. 2021;10(9):1080. doi: 10.3390/antibiotics10091080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Dutta H, Nath R, Saikia L. Multi-drug resistance in clinical isolates of Gram-negative bacilli in a tertiary care hospital of Assam. Indian J Med Res. 2014;139(4):643. [PMC free article] [PubMed] [Google Scholar]
- 51.Farhan SM, Ibrahim RA, Mahran KM, Hetta HF, Abd El-Baky RM. Antimicrobial resistance pattern and molecular genetic distribution of metallo-β-lactamases producing Pseudomonas aeruginosa isolated from hospitals in Minia, Egypt. Infect Drug Resist. 2019;12:2125. doi: 10.2147/IDR.S198373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Tahmasebi H, Dehbashi S, Arabestani MR. Prevalence and molecular typing of colistin-resistant Pseudomonas aeruginosa (CRPA) among β-lactamase-producing isolates: a study based on high-resolution melting curve analysis method. Infect Drug Resist. 2020;13:2943. doi: 10.2147/IDR.S264796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bills CB, Newberry JA, Rao GR, Matheson LW, Rao S, Mahadevan SV, et al. Acute respiratory illness among a prospective cohort of pediatric patients using emergency medical services in India: demographic and prehospital clinical predictors of mortality. PLoS ONE. 2020;15(4):e0230911. doi: 10.1371/journal.pone.0230911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Rawson TM, Moore LS, Zhu N, Ranganathan N, Skolimowska K, Gilchrist M, et al. Bacterial and fungal coinfection in individuals with coronavirus: a rapid review to support COVID-19 antimicrobial prescribing. Clin Infect Dis. 2020;71(9):2459–2468. doi: 10.1093/cid/ciaa530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ruan Q, Yang K, Wang W, Jiang L, Song J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med. 2020;46(5):846–848. doi: 10.1007/s00134-020-05991-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ramadan HK-A, Mahmoud MA, Aburahma MZ, Elkhawaga AA, El-Mokhtar MA, Sayed IM, et al. Predictors of severity and co-infection resistance profile in COVID-19 patients: First report from upper Egypt. Infect Drug Resist. 2020:3409–22. [DOI] [PMC free article] [PubMed]
- 57.Miri Mizher Al Muhana B, A Ali S, Sabri Jabbar A. Isolation of Pseudomonas aeruginosa from persistent bacterial coinfection of a COVID-19 patients with molecular detection of antibiotics resistance genes. Arch. Razi Inst. 2023;78(1):397–403. [DOI] [PMC free article] [PubMed]
- 58.Ahmed OB, Asghar AH, Bahwerth FS. Prevalence of ESBL genes of Pseudomonas aeruginosa strains isolated from Makkah Hospitals, Saudi Arabia. Euro J Biol Med Sci Res. 2015;3(6):12–18. [Google Scholar]
- 59.Tawfik AF, Shibl AM, Aljohi MA, Altammami MA, Al-Agamy MH. Distribution of Ambler class A, B and D β-lactamases among Pseudomonas aeruginosa isolates. Burns. 2012;38(6):855–860. doi: 10.1016/j.burns.2012.01.005. [DOI] [PubMed] [Google Scholar]
- 60.Bianco G, Boattini M, Comini S, Casale R, Iannaccone M, Cavallo R, et al. Occurrence of multi-carbapenemases producers among carbapenemase-producing Enterobacterales and in vitro activity of combinations including cefiderocol, ceftazidime-avibactam, meropenem-vaborbactam, and aztreonam in the COVID-19 era. Eur J Clin Microbiol Infect Dis. 2022;41(4):573–580. doi: 10.1007/s10096-022-04408-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Miftode I-L, Leca D, Miftode R-S, Roşu F, Plesca C, Loghin I, et al. The clash of the titans: COVID-19, carbapenem-resistant enterobacterales, and first mcr-1-mediated colistin resistance in humans in Romania. Antibiotics. 2023;12(2):324. doi: 10.3390/antibiotics12020324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hendricks MR, Lashua LP, Fischer DK, Flitter BA, Eichinger KM, Durbin JE, et al. Respiratory syncytial virus infection enhances Pseudomonas aeruginosa biofilm growth through dysregulation of nutritional immunity. Proc Natl Acad Sci. 2016;113(6):1642–1647. doi: 10.1073/pnas.1516979113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Li N, Ren A, Wang X, Fan X, Zhao Y, Gao GF, et al. Influenza viral neuraminidase primes bacterial coinfection through TGF-β–mediated expression of host cell receptors. Proc Natl Acad Sci. 2015;112(1):238–243. doi: 10.1073/pnas.1414422112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Qu J, Cai Z, Liu Y, Duan X, Han S, Liu J, et al. Persistent bacterial coinfection of a COVID-19 patient caused by a genetically adapted Pseudomonas aeruginosa chronic colonizer. Front Cell Infect Microbiol. 2021;11:641920. [DOI] [PMC free article] [PubMed]
- 65.Cho HH, Kwon KC, Kim S, Park Y, Koo SH. Association between biofilm formation and antimicrobial resistance in carbapenem-resistant Pseudomonas aeruginosa. Ann Clin Lab Sci. 2018;48(3):363–368. [PubMed] [Google Scholar]
- 66.Livermore DM. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: our worst nightmare? Clin Infect Dis. 2002;34(5):634–640. doi: 10.1086/338782. [DOI] [PubMed] [Google Scholar]
- 67.Babenko D, Turmuhambetova A, Sandle T, Pestrea SA, Moraru D, CHEŞCĂ In silico comparison of different types of MLVA with PFGE based on Pseudomonas aeruginosa genomes. Acta Medica . 2017;33:347. [Google Scholar]
- 68.Lalancette C, Charron D, Laferrière C, Dolcé P, Déziel E, Prévost M, et al. Hospital drains as reservoirs of Pseudomonas aeruginosa: multiple-locus variable-number of tandem repeats analysis genotypes recovered from faucets, sink surfaces and patients. Pathogens. 2017;6(3):36. doi: 10.3390/pathogens6030036. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data is available from the corresponding author upon request.