

Abstract
Background
Buffaloes are important contributors to the livestock economy in many countries, particularly in Asia, and tick-borne pathogens (TBPs) commonly infect buffaloes, giving rise to serious pathologies other than their zoonotic potential.
Methods
The present investigation focuses on the prevalence of TBPs infecting buffaloes worldwide. All published global data on TBPs in buffaloes were collected from different databases (e.g., PubMed, Scopus, ScienceDirect, and Google Scholar) and subjected to various meta-analyses using OpenMeta[Analyst] software, and all analyses were conducted based on a 95% confidence interval.
Results
Over 100 articles discussing the prevalence and species diversity of TBPs in buffaloes were retrieved. Most of these reports focused on water buffaloes (Bubalus bubalis), whereas a few reports on TBPs in African buffaloes (Syncerus caffer) had been published. The pooled global prevalence of the apicomplexan parasites Babesia and Theileria, as well as the bacterial pathogens Anaplasma, Coxiella burnetii, Borrelia, Bartonella, and Ehrlichia in addition to Crimean-Congo hemorrhagic fever virus, were all evaluated based on the detection methods and 95% confidence intervals. Interestingly, no Rickettsia spp. were detected in buffaloes with scarce data. TBPs of buffaloes displayed a fairly high species diversity, which underlines the high infection risk to other animals, especially cattle. Babesia bovis, B. bigemina, B. orientalis, B. occultans and B. naoakii, Theileria annulata, T. orientalis complex (orientalis/sergenti/buffeli), T. parva, T. mutans, T. sinensis, T. velifera, T. lestoquardi-like, T. taurotragi, T. sp. (buffalo) and T. ovis, and Anaplasma marginale, A. centrale, A. platys, A. platys-like and “Candidatus Anaplasma boleense” were all were identified from naturally infected buffaloes.
Conclusions
Several important aspects were highlighted for the status of TBPs, which have serious economic implications for the buffalo as well as cattle industries, particularly in Asian and African countries, which should aid in the development and implementation of prevention and control methods for veterinary care practitioners, and animal owners.
Graphical Abstract

Supplementary Information
The online version contains supplementary material available at 10.1186/s13071-023-05727-y.
Keywords: Tick-borne pathogens, Buffaloes, Meta-analysis, Babesia, Theileria, Anaplasma
Background
Along with cattle, buffaloes are important members of the subfamily Bovinae, and include various species, of which water buffaloes (Bubalus bubalis) are the most important and widely distributed worldwide over 77 countries on five continents, with a population exceeding 200 million [1, 2]. Water buffaloes (also known as domestic water buffalo or Asian buffalo) have two genetically distinct subspecies: river buffalo (81.5% of the global population) and swamp buffalo (18.5%) [2, 3]. The former is native to the Indian subcontinent [4] and has spread west, across southwestern Asia, Egypt, and Anatolia, and reached the Balkans and the Italian peninsula [5, 6]. Swamp buffaloes are predominant in Southeast Asia and Australia [4, 5, 7, 8]. The global water buffalo population is concentrated mostly in Asia (196 million) [4]. Smaller numbers are reared in Africa (3.4 million) and South America (2.0 million). Another buffalo species, Syncerus caffer (the African buffalo), lives in sub-Saharan Africa [9]; however, its numbers have been severely reduced since the eighteenth century as a result of the combined effects of anthropogenic pressures (e.g., land conversion), disease outbreaks, and climatic changes [10]. Although water buffaloes are mostly reared for milk production, they also contribute to the global meat sector, with around 4.3 million tons annually [11]. In some regions, buffaloes serve as working animals in various agricultural fields.
Tick-borne diseases (TBDs) are significant factors limiting the development of livestock industries worldwide, resulting in annual economic losses that can be estimated in billions of dollars [12, 13]. For example, the annual economic loss due to TBDs infecting cattle in Tanzania has been estimated at US$ 364 million [14]. Tick-borne pathogens (TBPs) pose a significant threat to buffalo health and production other than their zoonotic risks. In recent years, much progress has been made in the characterization and taxonomic justification of TBPs infecting buffaloes worldwide. Nonetheless, studies detailing various epidemiological aspects of TBPs infecting buffaloes are scarce. The present study provides a systematic review and meta-analysis of the global published data on TBPs infecting buffaloes worldwide, which should be useful for interpreting the epidemiology of this important group of pathogens.
Methods
Search strategy
The international databases PubMed, Scopus, ScienceDirect, and Google Scholar were systematically searched for studies on TBPs infecting buffaloes, with no date limit. The search was refined by article language (English) and type (research articles). Various keywords were used for the search: ticks; tick-borne pathogens; tick-borne diseases; Anaplasma, anaplasmosis; Babesia, babesiosis; Theileria, theileriosis; CCHF, Crimean-Congo hemorrhagic fever; Coxiella burnetii, Q fever; Ehrlichia, ehrlichiosis; Rickettsia, rickettsioses; Borrelia, borreliosis. These keywords were used in combination with the word “buffaloes,” and connected using the Boolean operators “AND” and “OR.”
Eligibility criteria
The collected publications were screened for inclusion independently by two of the authors, and any article with disagreement was discussed with a third author (Fig. 1). Studies were considered eligible to be included in this meta-analysis when (1) the study investigated TBPs in samples from buffaloes, either water (B. bubalis) or African (S. caffer) species, and (2) the study defined the number of examined buffaloes and number of positives. Studies on TBPs in animals other than buffaloes or those with inadequate methodologies were considered ineligible, and articles of non-original contributions including reviews, book chapters, and seminars were also excluded.
Fig. 1 .

Flow diagram established according to PRISMA [Preferred Reporting Items for Systematic Reviews and Meta-Analyses] guidelines and displaying the search and selection methodology
Data extraction
Data from eligible studies on TBPs of buffaloes were independently extracted and organized in a Microsoft Excel® spreadsheet by two of the authors. Any disagreement was resolved by consensus. The following information was extracted: authors, publication year, study subregion/country, number of tested samples, number of positive samples, detection methods employed, and various TBPs detected as well as the genetic markers (where recorded). Authors of the included articles were not contacted for further information, and no data conversions were conducted in the present study.
Meta-analysis
Data tabulated in the Excel spreadsheets were used for various meta-analyses conducted in the present study using OpenMeta[Analyst] software [15], and all analyses were conducted based on a 95% confidence interval (CI). The prevalence for various TBPs was calculated as “a pooled estimate” using the random-effects model associated with the DerSimonian–Laird method [16]. The heterogeneity between studies was estimated based on the I2 statistic. The heterogeneity values were considered high when I2 exceeded 50% [17, 18]. Subgroup analyses were conducted to detect the source of heterogeneity and included the variation in prevalence among various worldwide regions and the detection method. Publication bias was not assessed in the present study because it is not considered relevant for prevalence studies [19].
Results and discussion
Eligible studies
Data from 116 studies were found eligible to describe the TBPs infecting buffaloes worldwide, employing various detection methods [20–135]. Of these studies, 107 were used for meta-analysis (Fig. 1), and nine were not included [42, 69, 91, 103, 104, 108, 116, 119, 123] for the following reasons: the piroplasm was not identified even to the genus (n = 2), undefined mixed infections (n = 4), and double testing of the samples to identify two separate species of the same genus (n = 3). The country-wise distribution of these studies correlated with the number of reared buffaloes. The majority of these studies (n = 75) came from Asia [43–117], including India (21), Pakistan (13), China (10), Thailand (8), the Philippines (6), Vietnam (6), Iran (3), Sri Lanka (3), Iraq (1), Malaysia (1), Myanmar (1), Turkey (1), and Laos (1). Twenty-three studies were conducted in Africa [20–42], around two thirds (n = 15) of them came from Egypt [21–35], and a few studies were conducted in South Africa (3), Botswana (1), Kenya (1), Namibia (1), Tanzania (1), and Uganda (1). A few surveys (n = 12) were conducted on buffaloes from South America [124–135] including Brazil (9), Colombia (2), and Argentina (1). The lowest number (n = 4) of the eligible studies came from Europe [118–121]: Hungary (2), the Czech Republic (1), and Italy (1). Likewise, isolated surveys were conducted on buffaloes from North America (2), Cuba (1), and Mexico (1) (Additional file 1: Table S1). These studies comprised 145 datasets according to the detection method used, which included 57 datasets on Babesia spp., 48 on Theileria spp., 41 for Anaplasma spp., 15 for Coxiella burnetii, and 22 for miscellaneous TBPs (e.g., bacterial pathogens; Borrelia, Bartonella, Ehrlichia, and Rickettsia spp., and Crimean-Congo hemorrhagic fever virus [CCHFV]). Although data were divided into subgroups of investigated regions by continent, many studies did not include the buffalo species. As a result, data were not classified according to buffalo species, particularly African (S. caffer) and Asian (B. bubalis) buffaloes.
Piroplasmids
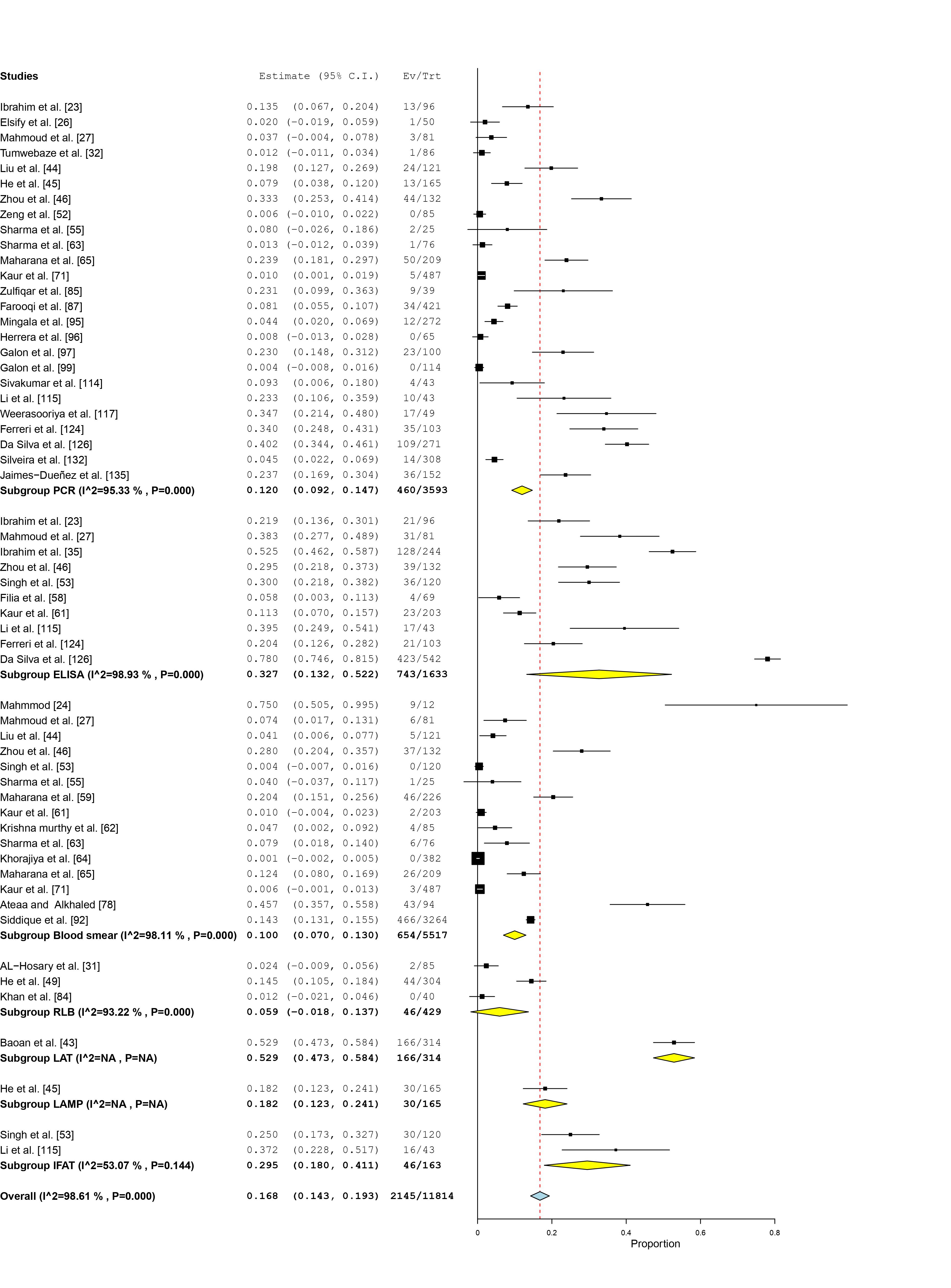
Ticks can transmit a variety of Babesia and Theileria piroplasmids, which can cause serious infections as well as economic consequences in livestock worldwide [136–138]. Microscopy detection of these piroplasmids has little significance in the identification of various species [136, 139]. In contrast, molecular diagnostic technologies provide rapid, sensitive, and accurate species delimitation [140, 141]. Alternatively, to detect parasite-specific antibodies, serological diagnostic procedures such as the enzyme-linked immunosorbent assay (ELISA) and indirect immunofluorescence antibody test (IFAT) can be used [142, 143]. However, the presence of antibodies as a marker of earlier parasite exposure does not always correspond to the current infectious condition, as antibodies usually persist for variable periods of time [143]. Four methods have been used for the direct detection of Babesia piroplasm in the blood of infected buffaloes worldwide (Additional file 2: Fig. S1). In total, 25 datasets screened Babesia spp. DNA in the blood of 3593 buffaloes worldwide, and 460 cases were found infected, resulting in a pooled global prevalence of 12.0% (95% CI 9.2–14.7%). This prevalence was slightly higher than that estimated for 5517 buffaloes diagnosed using microscopy (i.e., stained blood smears) in 15 datasets; 654 animals were found positive, with a pooled prevalence of 10.0% (95% CI 7.0–13.0%). Asymptomatic carrier animals in which no parasitemia could be found by microscopic examination led to a high false-negative diagnosis rate [137, 144]. Contrastingly, polymerase chain reaction (PCR) enabled sensitive and precise detection of various Theileria and Babesia species in carrier animals [144–148].
Three datasets used the reverse line blot hybridization (RLB) assay to test 429 animals, yielding a lower prevalence of 5.9%. On the contrary, a comparatively high prevalence (18.2%) was detected when a single dataset used the loop-mediated isothermal amplification (LAMP) assay on 165 animals (Table 1). Sero-surveys were also conducted to detect Babesia in the blood of buffaloes using various tests (e.g., ELISA, IFAT, and latex agglutination test [LAT]). Enzyme-linked immunosorbent assay was used to test 1633 serum samples in 10 datasets, and 743 were found positive for antibodies against Babesia spp., with a pooled prevalence of 32.7% (95% CI 13.2–52.2%). The highest prevalence was estimated with serology, which is similar to a previous report [149]. This could be due to the nature of the disease, since naturally infected animals that survive the acute disease continue to be healthy carriers of the parasites with low parasitemia [143]. Therefore, the animals may be seropositive while evading detection by direct tests; in terms of sensitivity, microscopy has the lowest, followed by direct PCR, and nested PCR [142, 143]. Among molecular techniques, real-time PCR (qPCR) has shown better sensitivity and specificity [143]; however, the sensitivity depends on many factors, including target gene, origin of the sample, and level of parasitemia. Overall, the number of datasets detailing the prevalence of Babesia in buffaloes from Asian countries (n = 41) was four times that obtained from African (n = 10) or South American (n = 6) countries (Table 1). Not all datasets named the species of buffaloes tested; however, all datasets (n = 10) in Africa came from Egypt, and it can be assumed that all investigated buffaloes belonged to the Asian species (B. bubalis).
Table 1.
Worldwide prevalence of Babesia infection in buffaloes, and variation in prevalence in relation to the detection method
| Parameter | No. datasets | No. tested | No. positive | Pooled estimate % based on 95% CI | Heterogeneity I2% |
|---|---|---|---|---|---|
| Overall prevalence | 57 | ||||
| Blood smear | 15 | 5517 | 654 | 10.0 (7.0–13.0) | 98.11 |
| PCR | 25 | 3593 | 460 | 12.0 (9.2–14.7) | 95.33% |
| RLB | 3 | 429 | 46 | 5.9 (−1.8 to 13.7) | 93.22% |
| LAMP | 1 | 165 | 30 | 18.2 (12.3–24.1) | NA |
| ELISA | 10 | 1633 | 743 | 32.7 (13.2–52.2) | 98.93% |
| IFAT | 2 | 163 | 46 | 29.5 (18.0–41.1) | 53.07% |
| LAT | 1 | 314 | 166 | 52.9 (47.3–58.4) | NA |
| Regional prevalence | |||||
| Africa | 10 | ||||
| Blood smear | 2 | 93 | 15 | 40.1 (−26.1 to 106.3) | 96.39% |
| PCR | 4 | 313 | 18 | 4.1 (0.2–8.0) | 74.42% |
| RLB | 1 | 85 | 2 | 2.4 (−0.9 to 5.6) | NA |
| ELISA | 3 | 421 | 180 | 37.7 (18.2–57.1) | 94.05% |
| Asia | 41 | ||||
| Blood smear | 13 | 5424 | 639 | 9.2 (6.1–12.3) | 98.29% |
| PCR | 17 | 2446 | 248 | 10.2 (7.4–13.0) | 93.79% |
| RLB | 2 | 344 | 44 | 7.8 (−5.2 to 20.8) | 96.01% |
| LAMP | 1 | 165 | 30 | 18.2 (12.3–24.1) | NA |
| ELISA | 5 | 567 | 119 | 22.2 (11.1–33.2) | 92.05% |
| IFAT | 2 | 163 | 46 | 29.5 (18.0–41.1) | 53.07% |
| LAT | 1 | 314 | 166 | 52.9 (47.3–58.4) | NA |
| South America | 6 | ||||
| PCR | 4 | 834 | 194 | 25.4 (5.6–45.3) | 98.16% |
| ELISA | 2 | 645 | 444 | 49.3 (3.5–63.5) | 99.43% |
NA not applicable
Babesia bovis and B. bigemina were the most frequently investigated and detected species in buffaloes based on either PCR sequencing or serological protocols, and B. orientalis were identified to a lesser extent (Additional file 1: Table S1). Babesia bovis and B. bigemina were the most common species in bovines worldwide. They are primarily found in tropical and subtropical regions of the world, including Australia, Africa, Asia, and the Americas, and are transmitted by the tick vectors Rhipicephalus (Boophilus) microplus and R. annulatus, and R. decoloratus for B. bigemina alone [149, 150]. Other species included B. occultans and Babesia sp. Mymensingh. Of note, Babesia sp. Mymensingh was recently named Babesia naoakii [151].
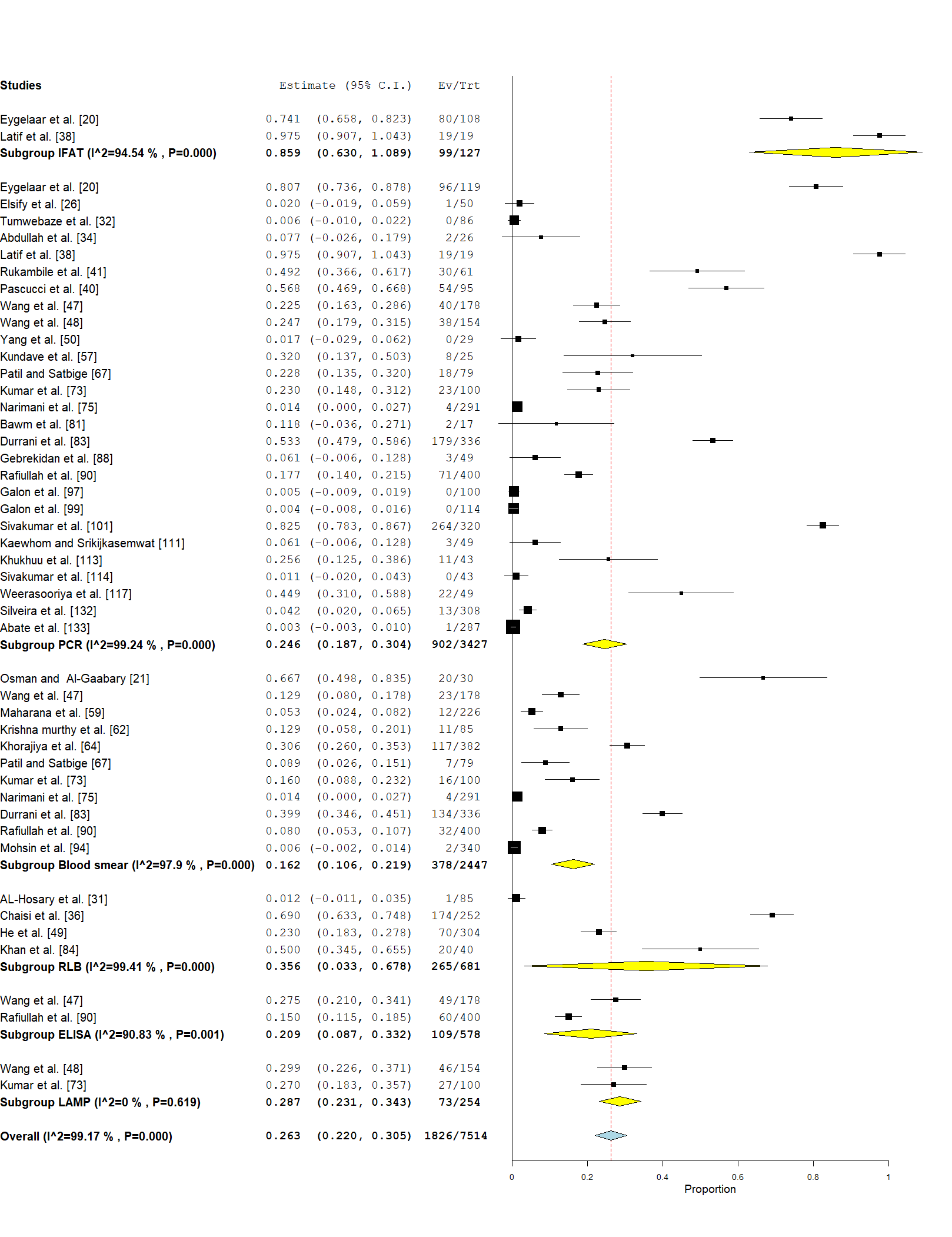
A large number of datasets were also found describing Theileria infections in buffaloes based either on direct detection methods (e.g., blood smear, PCR, RLB, LAMP) or serological assays for screening antibodies (e.g., ELISA and IFAT) (Additional file 3: Fig. S2). Like Babesia, the pooled global prevalence based on 3427 samples tested using PCR (24.6%; 95% CI 18.7–30.4%) was higher than that detected in 2447 samples tested microscopically (16.2%; 95% CI 10.6–21.9%) (Table 2). On the other hand, the estimated prevalence for ELISA-tested samples (n = 578) was 20.9% (95% CI 8.7–33.2%). Overall, the prevalence rates were much higher in Africa (12 datasets) than in Asia (34 datasets) for samples tested using either microscopy (66.7% and 13.4%, respectively) or PCR (42.0% and 20.7%, respectively); however, similar prevalence was estimated for both continents based on samples tested using the RLB (35.1 and 35.5%, respectively) (Table 2). Notably, the regional variation in the prevalence of Theileria spp. and Babesia spp. between Africa and Asia may be attributed to the difference in the number of datasets used and the species of buffaloes reared in Africa (B. bubalis and S. caffer) in comparison with the species reared in Asia (B. bubalis).
Table 2.
Worldwide prevalence of Theileria infection in buffaloes, and variation in prevalence in relation to the detection method
| Parameter | No. datasets | No. tested | No. positive | Pooled estimate % based on 95% CI | Heterogeneity I2 % |
|---|---|---|---|---|---|
| Overall prevalence | 48 | ||||
| Blood smear | 11 | 2447 | 378 | 16.2 (10.6–21.9) | 97.9% |
| PCR | 27 | 3427 | 902 | 24.6 (18.7–30.4) | 99.24% |
| RLB | 4 | 681 | 265 | 35.6 (3.3–67.8) | 99.41% |
| LAMP | 2 | 254 | 73 | 28.7 (23.1–34.3) | 0% |
| ELISA | 2 | 578 | 109 | 20.9 (8.7–33.2) | 90.83% |
| IFAT | 2 | 127 | 99 | 85.9 (63.0–108.9) | 94.54% |
| Regional prevalence | |||||
| Africa | 12 | ||||
| Blood smear | 1 | 30 | 20 | 66.7 (49.8–83.5) | NA |
| PCR | 7 | 456 | 202 | 42.0 (13.2–70.8) | 99.53% |
| RLB | 2 | 337 | 175 | 35.1 (−31.5 to 101.6) | 99.79% |
| IFAT | 2 | 127 | 99 | 85.9 (63.0–108.9) | 94.54% |
| Asia | 34 | ||||
| Blood smear | 10 | 2417 | 358 | 13.4 (7.9–18.9) | 97.87% |
| PCR | 18 | 2376 | 686 | 20.7 (12.6–28.8) | 99.14% |
| RLB | 2 | 344 | 90 | 35.5 (9.1–61.8) | 90.61% |
| LAMP | 2 | 254 | 73 | 28.7 (23.1–34.3) | 0% |
| ELISA | 2 | 578 | 109 | 20.9 (8.7–33.2) | 90.83% |
| South America (PCR) | 2 | 595 | 14 | 2.1 (−1.7 to 5.9) | 90.44% |
NA not applicable
Buffaloes have been found infected with different Theileria species, including Theileria annulata, T. orientalis complex (orientalis/sergenti/buffeli), and T. parva among the most frequently detected. Theileria annulata and T. parva (the causative agents of tropical or Mediterranean and East Coast fevers, respectively) are the most pathogenic species in bovines, whereas other species frequently cause asymptomatic infections in this host group [152]. Based on 18S ribosomal RNA sequences, phylogenetic analysis of Theileria genotypes reveals that the T. buffeli clade has the most genotypes and is found on all major continents, infecting cattle, African buffalo, water buffalo, and yak [137, 153, 154]. Members of this clade have previously been given a variety of species names, including T. buffeli, T. orientalis, and T. sergenti, and their taxonomy is debatable, but they have been proposed to constitute a single species known as T. buffeli [137, 153, 155] or T. orientalis complex in many other reports. Theileria mutans, T. sinensis, T. velifera, T. lestoquardi-like, T. taurotragi, T. sp. (buffalo), and T. ovis were also detected in naturally infected buffaloes.
Anaplasma species
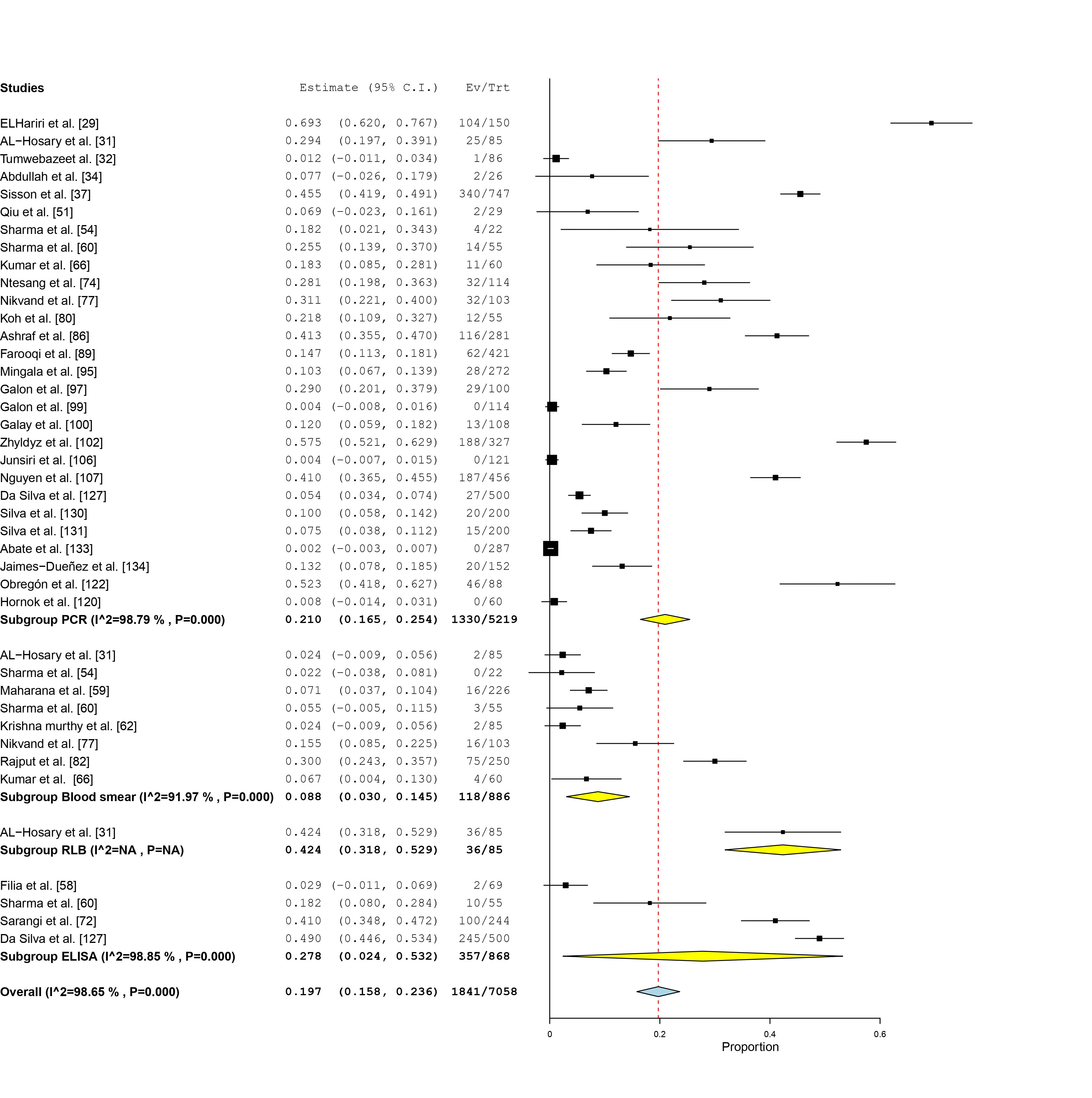
Anaplasmosis is an emerging infection that is gaining attention around the world because it affects animal body weight, causes abortions, reduces milk production, and leads to the death of animals [156]. Clinical disease is most common in cattle, although other ruminants, including water buffalo, could become persistently infected [157]. Anaplasma marginale was the most common tick-borne infection in buffaloes, and was considered the most widespread TBP globally in bovines, producing mild to severe hemolytic disease with significant economic loss [12, 158]. Forty-one datasets described Anaplasma infections in buffaloes, which represented the third most frequently tested TBPs in buffaloes worldwide after Babesia and Theileria, respectively (Additional file 4: Fig. S3). Anaplasma spp. were PCR-detected at a much higher rate than when microscopically detected (Table 3). Using blood smears, 886 samples in eight datasets were examined, and 118 were found positive resulting in pooled prevalence of 8.8% (95% CI 3.0–14.5%), whereas 5219 blood samples in 28 datasets were screened for Anaplasma DNA and 1330 were found positive with a pooled prevalence of 21.0% (95% CI 16.5–25.4%). Additionally, four datasets tested 868 serum samples using ELISA for the detection of Anaplasma spp., and 357 were found positive, giving rise to a prevalence of 27.8% (95% CI 2.4–53.2%). Moreover, RLB was used in a single study from Egypt for detection of A. marginale, with a prevalence rate of 42.4% (36/85), which is much higher than that of any other method.
Table 3.
Worldwide prevalence of Anaplasma infection in buffaloes, and variation in prevalence in relation to the detection method
| Parameter | No. datasets | No. tested | No. positive | Pooled estimate % based on 95% CI | Heterogeneity I2 % |
|---|---|---|---|---|---|
| Overall prevalence | 41 | ||||
| Blood smear | 8 | 886 | 118 | 8.8 (3.0–14.5) | 91.97% |
| PCR | 28 | 5219 | 1330 | 21.0 (16.5–25.4) | 98.79% |
| RLB | 1 | 85 | 36 | 42.4 (31.8–52.9) | NA |
| ELISA | 4 | 868 | 357 | 27.8 (2.4–53.2) | 98.85% |
| Regional prevalence | |||||
| Africa | 7 | ||||
| Blood smear | 1 | 85 | 2 | 2.4 (−0.9 to 5.6) | NA |
| PCR | 5 | 1094 | 472 | 30.6 (3.1–58.2) | 99.37% |
| RLB | 1 | 85 | 36 | 42.4 (31.8–52.9) | NA |
| Asia | 26 | ||||
| Blood smear | 7 | 801 | 116 | 9.8 (3.0–16.5) | 92.37% |
| PCR | 16 | 2638 | 730 | 22.2 (14.8–29.6) | 98.57% |
| ELISA | 3 | 368 | 112 | 20.6 (−5.5 to 46.8 | 98.08% |
| South America | 6 | ||||
| PCR | 5 | 1339 | 82 | 6.9 (2.2–11.5) | 94.96% |
| ELISA | 1 | 500 | 245 | 49.0 (44.6–53.4) | NA |
| North America (PCR) | 1 | 88 | 46 | 52.3 (41.8–62.7) | NA |
| Europe (PCR) | 1 | 60 | 0 | 0.8 (−1.4 to 3.1) | NA |
NA not applicable
In Africa, most studies relied on PCR for diagnosis of Anaplasma species, where 472 out of 1094 samples were found infected, with a pooled prevalence of 30.6% (95% CI 3.1–58.2%). The African buffalo was investigated in a single study from South Africa (Additional file 1: Table S1). Regarding Asia, PCR and ELISA were used in 16 and three datasets, respectively, with a nearly equal prevalence (22.2% and 20.6%, respectively), while lower prevalence was estimated based on microscopy (9.8%). Anaplasma marginale, A. centrale, A. platys, A. platys-like, and “Candidatus Anaplasma boleense” were recovered from infected buffaloes, with A. marginale being the most frequently investigated and detected species. Anaplasma platys and A. platys-like were detected in infected water buffaloes from Egypt, Thailand, and Malaysia [31, 34, 80, 107]. Anaplasma centrale was found infecting water buffaloes from Pakistan [82], and African buffaloes in South Africa and Uganda [37, 42]. Additionally, “Candidatus A. boleense” was recorded in a single study in water buffaloes from Malaysia [80]. However, A. phagocytophilum has a broad host range including humans [159]; only one study investigated the parasite in 60 water buffaloes in Hungary, and no infection was detected by qPCR [120].
Coxiella burnetii
The intracellular Gram-negative bacterium Coxiella burnetii, which can be excreted in tick feces and saliva, is the cause of Q fever, a zoonotic infection that is globally transmitted [160, 161]. Because of the prevalence of this bacterium in ticks from different bioclimatic zones and socioeconomic contexts, it is clear that ticks play a significant role in the epidemiology of Q fever [162, 163]. Coxiella burnetii infection leads mainly to reproductive disorders including abortions, premature birth, stillbirth, and poor calf deliveries, which have a negative economic impact on livestock [164–166]. Infection with C. burnetii in humans is typically asymptomatic or appears like the flu and can result in acute and chronic fever disease and pneumonia. It is mostly transmitted to humans by aerosolized dry, contaminated soil or animal products [160, 162, 166, 167].
Coxiella burnetii infections were tested in sera of buffaloes in six datasets worldwide using ELISA (five datasets) and IFAT (one set). A total of 987 sera were examined using ELISA, and 63 harbored the antibodies against C. burnetii, resulting in a pooled prevalence of 5.3% (95% CI 1.4–9.2%) (Table 4). The prevalence of Coxiella burnetii in buffaloes from Africa was higher (7.5%) than that in Asia (3.8%) based on ELISA. A higher number of samples were examined using PCR (1360 samples) including buffaloes’ blood, milk, vaginal discharges, vaginal and preputial swabs, placental cotyledons, and aborted fetuses (Additional file 1: Table S1), which were used for estimating the pooled prevalence (4.5%; 95% CI 1.9–7.1%). The detection of Coxiella burnetii in buffaloes’ sera along with the detection in raw milk [68, 69, 76] suggests that they can play a role in the transmission of Q fever to humans, but more extensive prevalence studies need to be carried out to define the role of buffaloes as reservoirs for this pathogen. Furthermore, the role of C. burnetii as an abortive agent in buffaloes has been suggested [121].
Table 4.
Worldwide prevalence of C. burnetii infection in buffaloes, and variation in prevalence in relation to the detection method
| Parameter | No. datasets | No. tested | No. positive | Pooled estimate % based on 95% CI | Heterogeneity I2 % |
|---|---|---|---|---|---|
| Overall prevalence | 15 | ||||
| PCR | 9 | 1360 | 106 | 4.5 (1.9–7.1) | 92.71% |
| ELISA | 5 | 987 | 63 | 5.3 (1.4–9.2) | 91.54% |
| IFAT | 1 | 156 | 7 | 4.5 (1.2–7.7) | NA |
| Regional prevalence | |||||
| Africa | 4 | ||||
| PCR | 2 | 52 | 0 | 1.9 (−1.7 to 5.4) | 0% |
| ELISA | 2 | 457 | 40 | 7.5 (0.4–14.6) | 89.14% |
| Asia | 10 | ||||
| PCR | 6 | 1144 | 92 | 4.6 (1.5–7.6) | 94.92% |
| ELISA | 3 | 530 | 23 | 3.8 (−0.3 to 7.9) | 88.84% |
| IFAT | 1 | 156 | 7 | 4.5 (1.2–7.7) | NA |
| Europe (PCR) | 1 | 164 | 14 | 8.5 (4.3–12.8) | NA |
NA not applicable
Miscellaneous pathogens
Isolated surveys have detected a few potentially zoonotic TBPs infecting buffaloes worldwide (Table 5). Notably, 5617 sera from three datasets in Brazil were examined for Borrelia burgdorferi and 4157 were found to harbor antibodies, resulting in a pooled prevalence of 68.2% (95% CI 53.2–83.1%). PCR was used to screen B. burgdorferi sensu lato in eight buffaloes in the Czech Republic, and a single case was found infected (12.5%) in contrast to the examined 26 buffaloes in Egypt for Borrelia theileri, where no cases were found infected (Additional file 1: Table S1). Borrelia burgdorferi is a tick-borne obligatory parasite with a natural reservoir of a range of mammals, although, infection of these natural hosts does not result in disease; nonetheless, infection of humans can result in Lyme disease [168–170].
Table 5.
Worldwide prevalence of miscellaneous tick-borne pathogens of buffaloes, and variation in prevalence in relation to the detection method
| Pathogen | Parameter | No. datasets | No. tested | No. positive | Pooled estimate % based on 95% CI | Heterogeneity I2 % |
|---|---|---|---|---|---|---|
| Borrelia spp. | Overall prevalence | 5 | ||||
| PCR | 2 | 34 | 1 | 2.4 (−2.6 to 7.3) | 0% | |
| iELISA | 3 | 5617 | 4157 | 68.2 (53.2–83.1) | 98.67% | |
| Regional prevalence | ||||||
| Africa (PCR) | 1 | 26 | 0 | 1.9 (−3.2 to 6.9) | NA | |
| South America (iELISA) | 3 | 5617 | 4157 | 68.2 (53.2–83.1) | 98.67% | |
| Europe (PCR) | 1 | 8 | 1 | 12.5 (−10.4 to 35.4) | NA | |
| Bartonella spp. | Overall prevalence | 5 | ||||
| PCR | 3 | 208 | 3 | 1.5 (−1.9 to 5.0) | 42.93% | |
| Culture | 1 | 103 | 7 | 6.8 (1.9–11.7) | NA | |
| IFAT | 1 | 156 | 25 | 16.0 (10.3–21.8) | NA | |
| Regional prevalence | ||||||
| Africa (PCR) | 2 | 52 | 3 | 5.0 (−3.9 to 13.9) | 50.99% | |
| Asia | 3 | |||||
| PCR | 1 | 156 | 0 | 0.3 (−0.6 to 1.2) | NA | |
| Culture | 1 | 103 | 7 | 6.8 (1.9–11.7) | NA | |
| IFAT | 1 | 156 | 25 | 16.0 (10.3–21.8) | NA | |
| Ehrlichia spp. | Overall prevalence (PCR) | 4 | 328 | 13 | 3.1 (−0.3 to 6.5) | 73.39% |
| Regional prevalence | ||||||
| Africa | 1 | 95 | 3 | 3.2 (−0.4 to 6.7) | NA | |
| Asia | 3 | 233 | 10 | 3.5 (−1.5 to 8.5) | 79.77% | |
| CCHFV | Overall prevalence (ELISA) | 4 | 880 | 145 | 16.0 (9.9–22.1) | 99.48% |
| Regional prevalence | ||||||
| Africa | 3 | 608 | 145 | 23.8 (12.4–35.3) | 99.65% | |
| Asia | 1 | 272 | 0 | 0.2 (−0.3 to 0.7) | NA | |
| Rickettsiae | Overall prevalence (PCR) | 4 | 220 | 0 | 0.7 (−0.4 to 1.7) | 0% |
iELISA indirect ELISA, NA not applicable
Bartonella species are arthropod-borne Gram-negative bacteria that infect erythrocytes, endothelial cells, and macrophages, frequently resulting in chronic blood-borne infections [171]. They are the causative agents of multiple human diseases, and their main vectors include fleas, keds, lice, sand flies, bed bugs, biting flies, and ticks [171–173]. Ticks were previously postulated but not confirmed as a vector for Bartonella transmission; nevertheless, there is increasing evidence of transovarial and transstadial transmission of bartonellae in ixodid ticks [174–176]. A pooled prevalence of 1.5% (95% CI −1.9 to 5.0%) was estimated for Bartonella spp. in buffaloes from three datasets based on PCR. Other methods were used based on traditional culture and IFAT for the detection of Bartonella spp. and resulted in higher prevalence (6.8% and 16%, respectively) from one dataset for each method. Bartonella bovis was identified in water buffaloes from Thailand based on multi-locus sequence typing in a single study [105]. Bartonella henselae, B. vinsonii subsp. berkhoffii, and B. tamiae antibodies were found in buffaloes from Thailand using indirect IFAT; however, PCR results were negative [109].
Much lower prevalence of 3.1% (95% CI −0.3 to 6.5%) was estimated for 328 buffaloes tested for ehrlichiosis with PCR from four datasets, which constitutes a potentially fatal emerging zoonosis of global veterinary concern [177]. Ehrlichia ruminantium was the species identified from African buffaloes in Namibia [40]. In South Africa, a buffalo calf (S. caffer) was suspected of having died as a result of heartwater (E. ruminantium) [178]. However, small samples were used for the detection of Rickettsia species; 220 buffaloes from three continents, namely Africa (Egypt), Asia (Philippines), and Europe (Hungary), were found free from infection based on PCR [33, 34, 98, 120].
Crimean-Congo hemorrhagic fever (CCHF) is a viral infection spread by ticks. Humans are the only known hosts that develop sickness following CCHFV infection, owing to the virus's asymptomatic presence in animal reservoirs, which allows it to circulate undetected [179]. Humans can become infected with CCHFV through tick bites or contact with contaminated animal tissues during and immediately after slaughter [180, 181]. Notably, humans who had high CCHF seropositivity had a history of animal contact, animal husbandry, farming, and tick bites [182]. Buffaloes can act as silent reservoirs of CCHFV; the sera of 880 buffaloes were examined using ELISA from four datasets and 145 were found infected, with a pooled prevalence of 16.0% (95% CI 9.9–22.1%).
Conclusions
The present article provides the first meta-analysis of the published data on TBPs in buffaloes. The analyses conducted have some limitations sometimes due to the limited number of available studies (e.g., rickettsioses) as well as variability in the relevance of the reported data and diagnostic methods used. Nonetheless, the close interaction of varied animal species, the availability of mixed animal shelters, and unregulated animal movements in many African and Asian countries all increase the risk of a pathogen crossing the species barrier. This is evident in many species and genotypes of Babesia, Theileria, and Anaplasma that are circulating between buffaloes and cattle worldwide. Consequently, detection of the pathogenic species is considered critical not only for buffaloes but also for cattle raised in the same areas, with economic implications. There is evidence of high species diversity of Babesia, Theileria, and Anaplasma infecting buffaloes, which suggests susceptibility to a wide range of TBPs, and adequate control measures should be applied to prevent their circulation.
Supplementary Information
Additional file 1: Table S1. Study characteristics of tick-borne pathogen surveys in buffaloes worldwide.
{kind=link}
Additional file 2: Figure S1. Forest plot diagrams for random effects in the meta-analysis of the prevalence of Babesia spp. infections in buffaloes worldwide. The middle point of each line indicates the prevalence, while the length of the line is the 95% confidence interval for each study. Diamonds refers to the prevalence in accordance with detection methods.
{kind=link}
Additional file 3: Figure S2. Forest plot diagrams for random effects in the meta-analysis of the prevalence of Theileria spp. infections in buffaloes worldwide. The middle point of each line indicates the prevalence, while the length of the line is the 95% confidence interval of each study. Diamonds refers to the prevalence in accordance with detection methods.
{kind=link}
Additional file 4: Figure S3. Forest plot diagrams for random effects in the meta-analysis of the prevalence of Anaplasma spp. infections in buffaloes worldwide. The middle point of each line indicates the prevalence, while the length of the line is the 95% confidence interval of each study. Diamonds refers to the prevalence in accordance with detection methods.
Acknowledgements
Not applicable.
Author contributions
Conceptualization, EE and MAR; literature search, EE, RE and SS; data extraction and organization, RE, EE, and IA; meta-analysis, IA and SS; writing—original draft preparation, EE, SAESES and MAR; writing—review and editing, IA, MAR and EE. All authors read and approved the final manuscript.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). Open access funding was provided by the Transformative Agreement between Springer Nature and the Science, Technology and Innovation Funding Authority (STDF) in cooperation with the Egyptian Knowledge Bank (EKB).
Availability of data and materials
All data generated or analysed during this study are included in this published article and its supplementary information files.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
El-Sayed El-Alfy, Email: sydnabil@mans.edu.eg.
Ibrahim Abbas, Email: ielsayed@mans.edu.eg.
Rana Elseadawy, Email: ranamagdy6666@gmail.com.
Somaya Saleh, Email: soma.saleh2010@gmail.com.
Bassem Elmishmishy, Email: dr_b_elmishmishy@mans.edu.eg.
Shimaa Abd El-Salam El-Sayed, Email: Shimaa_a@mans.edu.eg.
Mohamed Abdo Rizk, Email: dr_moh_abdo2008@mans.edu.eg.
References
- 1.Borghese A. Buffalo production and research. FAO Regional Office for Europe. In: Borghese A, editor. Technical Series 67. Italy: Food and Agriculture Organization Rome; 2005. [Google Scholar]
- 2.Minervino AHH, Zava M, Vecchio D, Borghese A. Bubalus bubalis: a short story. Front Vet Sci. 2020;7:570413. doi: 10.3389/fvets.2020.570413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Colli L, Milanesi M, Vajana E, Iamartino D, Bomba L, Puglisi F, et al. New insights on water buffalo genomic diversity and post-domestication migration routes from medium density SNP chip data. Front Genet. 2018;2:9–53. doi: 10.3389/fgene.2018.00053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zhang Y, Colli L, Barker JSF. Asian water buffalo: domestication, history and genetics. Anim Genet. 2020;51:177–191. doi: 10.1111/age.12911. [DOI] [PubMed] [Google Scholar]
- 5.Cockrill WR 1974 ed. The husbandry and health of the domestic buffalo. Rome: Food and Agriculture Organization of the United Nations. 1974.
- 6.Kumar S, Nagarajan M, Sandhu JS, Kumar N, Behl V. Phylogeography and domestication of Indian river buffalo. BMC Evol Biol. 2007;7:186. doi: 10.1186/1471-2148-7-186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cockrill WR. The water buffalo Rome: Animal production and health series no. 4. Food and Agriculture Organization of the United Nations. 1977.
- 8.Cockrill WR. The water buffalo: a review. Br Vet J. 1981;137:8–16. doi: 10.1016/S0007-1935(17)31782-7. [DOI] [PubMed] [Google Scholar]
- 9.Megaze A, Balakrishnan M, Belay G. Current population estimate and distribution of the African buffalo in Chebera Churchura National Park Ethiopia. Afr J Ecol. 2017;56:12–19. doi: 10.1111/aje.12411. [DOI] [Google Scholar]
- 10.Cornélis D, Melletti M, Korte L, Ryan SJ, Mirabile M, Prin T, Prins HH. 2014 African buffalo Syncerus caffer (Sparrman, 1779) Ecology, evolution and behaviour of wild cattle: implications for conservation 326–372.
- 11.Di Stasio L, Brugiapaglia A. Current knowledge on river buffalo meat: a critical analysis. Animals. 2021;11:2111. doi: 10.3390/ani11072111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Suarez CE, Noh S. Emerging perspectives in the research of bovine babesiosis and anaplasmosis. Vet Parasitol. 2011;180:109–125. doi: 10.1016/j.vetpar.2011.05.032. [DOI] [PubMed] [Google Scholar]
- 13.Garcia K, Weakley M, Do T, Mir S. Current and future molecular diagnostics of tick-borne diseases in cattle. Vet Sci. 2022;9:241. doi: 10.3390/vetsci9050241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kivaria FM. Estimated direct economic costs associated with tick-borne diseases on cattle in Tanzania. Trop Anim Health Prod. 2006;38:291–299. doi: 10.1007/s11250-006-4181-2. [DOI] [PubMed] [Google Scholar]
- 15.Wallace BC, Dahabreh IJ, Trikalinos TA, Lau J, Trow P, Schmid CH. Closing the gap between methodologists and end-users: R as a computational back-end. J Stat Softw. 2012;49:1–15. doi: 10.18637/jss.v049.i05. [DOI] [Google Scholar]
- 16.DerSimonian R, Laird N. Meta-analysis in clinical trials revisited. Contemp Clin Trials. 2015;45:139–145. doi: 10.1016/j.cct.2015.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Higgins JP, Thompson SG. Quantifying heterogeneity in a meta-analysis. Stat Med. 2002;21:1539–1558. doi: 10.1002/sim.1186. [DOI] [PubMed] [Google Scholar]
- 18.Higgins JP, Thompson SG, Deeks JJ, Altman DG. Measuring inconsistency in meta-analyses. BMJ. 2003;327:557–560. doi: 10.1136/bmj.327.7414.557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hunter JP, Saratzis A, Sutton AJ, Boucher RH, Sayers RD, Bown MJ. In meta-analyses of proportion studies, funnel plots were found to be an inaccurate method of assessing publication bias. J Clin Epidemiol. 2014;67:897–903. doi: 10.1016/j.jclinepi.2014.03.003. [DOI] [PubMed] [Google Scholar]
- 20.Eygelaar D, Jori F, Mokopasetso M, Sibeko KP, Collins NE, Vorster I, et al. Tick-borne haemoparasites in African buffalo (Syncerus caffer) from two wildlife areas in Northern Botswana. Parasit Vectors. 2015;8:26. doi: 10.1186/s13071-014-0627-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Osman SA, Al-Gaabary MH. Clinical, haematological and therapeutic studies on tropical theileriosis in water buffaloes (Bubalus bubalis) in Egypt. Vet Parasitol. 2007;146:337–340. doi: 10.1016/j.vetpar.2007.03.012. [DOI] [PubMed] [Google Scholar]
- 22.Mohamed M, Said AR, Murad A, Graham R. A serological survey of Crimean-Congo haemorrhagic fever in animals in the Sharkia governorate of Egypt. Vet Ital. 2008;44:513–517. [PubMed] [Google Scholar]
- 23.Ibrahim HM, Moumouni PFA, Mohammed-Geba K, Sheir SK, Hashem IS, Cao S, et al. Molecular and serological prevalence of Babesia bigemina and Babesia bovis in cattle and water buffalos under small-scale dairy farming in Beheira and Faiyum Provinces. Egypt Vet Parasitol. 2013;198:187–192. doi: 10.1016/j.vetpar.2013.08.028. [DOI] [PubMed] [Google Scholar]
- 24.Mahmmod Y. Natural Babesia bovis infection in water buffaloes (Bubalus bubalis) and crossbred cattle under field conditions in Egypt: a preliminary study. J Arthropod Borne Dis. 2014;8:1–9. [PMC free article] [PubMed] [Google Scholar]
- 25.Horton KC, Wasfy M, Samaha H, Abdel-Rahman B, Safwat S, Abdel Fadeel M, et al. Serosurvey for zoonotic viral and bacterial pathogens among slaughtered livestock in Egypt. Vector Borne Zoonotic Dis. 2014;14:633–639. doi: 10.1089/vbz.2013.1525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Elsify A, Sivakumar T, Nayel M, Salama A, Elkhtam A, Rizk M, et al. An epidemiological survey of bovine Babesia and Theileria parasites in cattle, buffaloes, and sheep in Egypt. Parasitol Int. 2015;64:79–85. doi: 10.1016/j.parint.2014.10.002. [DOI] [PubMed] [Google Scholar]
- 27.Mahmoud MS, Kandil OM, Nasr SM, Hendawy SH, Habeeb SM, Mabrouk DM, et al. Serological and molecular diagnostic surveys combined with examining hematological profiles suggests increased levels of infection and hematological response of cattle to babesiosis infections compared to native buffaloes in Egypt. Parasit Vectors. 2015;8:319. doi: 10.1186/s13071-015-0928-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Abdel-Moein KA, Hamza DA. The burden of Coxiella burnetii among aborted dairy animals in Egypt and its public health implications. Acta Trop. 2017;166:92–95. doi: 10.1016/j.actatropica.2016.11.011. [DOI] [PubMed] [Google Scholar]
- 29.ELHariri MD, ELHelw RA, Hamza DA, Soliman DE. Molecular detection of Anaplasma marginale in the Egyptian water buffaloes (Bubalus bubalis) based on major surface protein 1α. J Egypt Soc Parasitol. 2017;47:247–252. doi: 10.21608/jesp.2017.77758. [DOI] [Google Scholar]
- 30.Klemmer J, Njeru J, Emam A, El-Sayed A, Moawad AA, Henning K, et al. Q fever in Egypt: Epidemiological survey of Coxiella burnetii specific antibodies in cattle, buffaloes, sheep, goats and camels. PLoS ONE. 2018;13:e0192188. doi: 10.1371/journal.pone.0192188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Al-Hosary A, Răileanu C, Tauchmann O, Fischer S, Nijhof AM, Silaghi C. Epidemiology and genotyping of Anaplasma marginale and co-infection with piroplasms and other Anaplasmataceae in cattle and buffaloes from Egypt. Parasit Vectors. 2020;13:495. doi: 10.1186/s13071-020-04372-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tumwebaze MA, Lee SH, Moumouni PFA, Mohammed-Geba K, Sheir SK, Galal-Khallaf A, et al. First detection of Anaplasma ovis in sheep and Anaplasma platys-like variants from cattle in Menoufia governorate. Egypt Parasitol Int. 2020;78:102150. doi: 10.1016/j.parint.2020.102150. [DOI] [PubMed] [Google Scholar]
- 33.Abdullah HH, Elbayoumy MK, Allam AM, Ashry HM, Abdel-Shafy S. Molecular epidemiology of certain vector-borne bacterial microorganisms in domestic animals and their ectoparasites in Egypt. Trop Anim Health Prod. 2021;53:484. doi: 10.1007/s11250-021-02911-z. [DOI] [PubMed] [Google Scholar]
- 34.Abdullah HH, Amanzougaghene N, Dahmana H, Louni M, Raoult D, Mediannikov O. Multiple vector-borne pathogens of domestic animals in Egypt. PLoS Negl Trop Dis. 2021;15:e0009767. doi: 10.1371/journal.pntd.0009767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ibrahim HM, Galon EMS, Tumwebaze MA, Byamukama B, Liu M, Mohammed-Geba K, et al. Serological survey of Babesia bigemina and Babesia bovis in cattle and water Buffaloes from Menoufia Province. Egypt Acta Parasitol. 2021;66:1458–1465. doi: 10.1007/s11686-021-00338-y. [DOI] [PubMed] [Google Scholar]
- 36.Chaisi ME, Sibeko KP, Collins NE, Potgieter FT, Oosthuizen MC. Identification of Theileria parva and Theileria sp. (buffalo) 18S rRNA gene sequence variants in the African Buffalo (Syncerus caffer) in southern Africa. Vet Parasitol. 2011;182:150–162. doi: 10.1016/j.vetpar.2011.05.041. [DOI] [PubMed] [Google Scholar]
- 37.Sisson D, Hufschmid J, Jolles A, Beechler B, Jabbar A. Molecular characterisation of Anaplasma species from African buffalo (Syncerus caffer) in Kruger national park. South Africa Ticks Tick Borne Dis. 2017;8:400–406. doi: 10.1016/j.ttbdis.2017.01.003. [DOI] [PubMed] [Google Scholar]
- 38.Latif AA, Troskie PC, Peba SB, Maboko BB, Pienaar R, Mans BJ. Corridor disease (buffalo-associated Theileria parva) outbreak in cattle introduced onto a game ranch and investigations into their carrier-state. Vet Parasitol Reg Stud Reports. 2019;18:100331. doi: 10.1016/j.vprsr.2019.100331. [DOI] [PubMed] [Google Scholar]
- 39.Obanda V, Agwanda B, Blanco-Penedo I, Mwangi IA, King'ori E, Omondi GP, et al. Livestock presence influences the seroprevalence of Crimean Congo hemorrhagic fever virus on sympatric wildlife in Kenya. Vector Borne Zoonotic Dis. 2021;21:809–816. doi: 10.1089/vbz.2021.0024. [DOI] [PubMed] [Google Scholar]
- 40.Pascucci I, Dondona AC, Cammà C, Marcacci M, Di Domenico M, Lelli R, et al. Survey of ixodid ticks and two tick-borne pathogens in African buffaloes, Syncerus caffer, from the Caprivi Strip. Namibia J Zoo Wildl Med. 2011;42:634–640. doi: 10.1638/2011-0014.1. [DOI] [PubMed] [Google Scholar]
- 41.Rukambile E, Machuka E, Njahira M, Kyalo M, Skilton R, Mwega E, et al. Population genetic analysis of Theileria parva isolated in cattle and buffaloes in Tanzania using minisatellite and microsatellite markers. Vet Parasitol. 2016;224:20–26. doi: 10.1016/j.vetpar.2016.04.038. [DOI] [PubMed] [Google Scholar]
- 42.Oura CAL, Tait A, Asiimwe B, Lubega GW, Weir W. Haemoparasite prevalence and Theileria parva strain diversity in Cape buffalo (Syncerus caffer) in Uganda. Vet Parasitol. 2011;175:212–219. doi: 10.1016/j.vetpar.2010.10.032. [DOI] [PubMed] [Google Scholar]
- 43.Baoan Y, Junlong Z, Enyong L, Sutian D, Junhua S, Zhongling L. Serological investigations on Babesia orientalis infection status of water buffaloes in Hubei Province. Parasitol Res. 2002;88:11–12. doi: 10.1007/s00436-001-0561-8. [DOI] [PubMed] [Google Scholar]
- 44.Liu Q, Zhou YQ, Zhou DN, Liu EY, Du K, Chen SG, et al. Semi-nested PCR detection of Babesia orientalis in its natural hosts Rhipicephalus haemaphysaloides and buffalo. Vet Parasitol. 2007;143:260–266. doi: 10.1016/j.vetpar.2006.08.016. [DOI] [PubMed] [Google Scholar]
- 45.He L, Zhou YQ, Oosthuizen MC, Zhao JL. Loop-mediated isothermal amplification (LAMP) detection of Babesia orientalis in water buffalo (Bubalus babalis, Linnaeus, 1758) in China. Vet Parasitol. 2009;165:36–40. doi: 10.1016/j.vetpar.2009.06.036. [DOI] [PubMed] [Google Scholar]
- 46.Zhou DN, Du F, Liu Q, Zhou YQ, Zhao JL. A 29-kDa merozoite protein is a prospective antigen for serological diagnosis of Babesia orientalis infection in buffaloes. Vet Parasitol. 2009;162:1–6. doi: 10.1016/j.vetpar.2009.03.001. [DOI] [PubMed] [Google Scholar]
- 47.Wang LX, Zhao JH, He L, Liu Q, Zhou DN, Zhou YQ, Zhao JL. An indirect ELISA for detection of Theileria sergenti antibodies in water buffalo using a recombinant major piroplasm surface protein. Vet Parasitol. 2010;170:323–326. doi: 10.1016/j.vetpar.2010.02.009. [DOI] [PubMed] [Google Scholar]
- 48.Wang LX, He L, Fang R, Song QQ, Tu P, Jenkins A, et al. Loop-mediated isothermal amplification (LAMP) assay for detection of Theileria sergenti infection targeting the p33 gene. Vet Parasitol. 2010;171:159–162. doi: 10.1016/j.vetpar.2010.02.046. [DOI] [PubMed] [Google Scholar]
- 49.He L, Feng HH, Zhang WJ, Zhang QL, Fang R, Wang LX, et al. Occurrence of Theileria and Babesia species in water buffalo (Bubalus babalis, Linnaeus, 1758) in the Hubei province. South China Vet Parasitol. 2012;186:490–496. doi: 10.1016/j.vetpar.2011.11.021. [DOI] [PubMed] [Google Scholar]
- 50.Yang Y, Mao Y, Kelly P, Yang Z, Luan L, Zhang J, et al. A pan-Theileria FRET-qPCR survey for Theileria spp in ruminants from nine provinces of China. Parasit Vectors. 2014;7:413. doi: 10.1186/1756-3305-7-413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Qiu H, Kelly PJ, Zhang J, Luo Q, Yang Y, Mao Y, et al. Molecular detection of Anaplasma spp and Ehrlichia spp in ruminants from twelve provinces of China. Can J Infect Dis Med Microbiol. 2016;2016:9183861. doi: 10.1155/2016/9183861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zeng Z, Zhou S, Xu G, Liu W, Han T, Liu J, et al. Prevalence and phylogenetic analysis of Babesia parasites in reservoir host species in Fujian province Southeast China. Zoonoses Publ Health. 2022;69:915–924. doi: 10.1111/zph.12988. [DOI] [PubMed] [Google Scholar]
- 53.Singh H, Mishra AK, Rao JR, Tewari AK. Comparison of indirect fluorescent antibody test (IFAT) and slide enzyme linked immunosorbent assay (SELISA) for diagnosis of Babesia bigemina infection in bovines. Trop Anim Health Prod. 2009;41:153–159. doi: 10.1007/s11250-008-9170-1. [DOI] [PubMed] [Google Scholar]
- 54.Sharma A, Singla LD, Kaur P, Bal MS, Batth BK, Juyal PD. Prevalence and haemato-biochemical profile of Anaplasma marginale infection in dairy animals of Punjab (India) Asian Pac J Trop Med. 2013;6:139–144. doi: 10.1016/S1995-7645(13)60010-3. [DOI] [PubMed] [Google Scholar]
- 55.Sharma A, Singla LD, Tuli A, Kaur P, Batth BK, Javed M, Juyal PD. Molecular prevalence of Babesia bigemina and Trypanosoma evansi in dairy animals from Punjab, India, by duplex PCR: a step forward to the detection and management of concurrent latent infections. Biomed Res Int. 2013;2013:893862. doi: 10.1155/2013/893862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Das DP, Malik SVS, Rawool DB, Das S, Shoukat S, Gandham RK, et al. Isolation of Coxiella burnetii from bovines with history of reproductive disorders in India and phylogenetic inference based on the partial sequencing of IS1111 element. Infect Genet Evol. 2014;22:67–71. doi: 10.1016/j.meegid.2013.12.017. [DOI] [PubMed] [Google Scholar]
- 57.Kundave VR, Patel AK, Patel PV, Hasnani JJ, Joshi CG. Qualitative and quantitative assessment of Theileria annulata in cattle and buffaloes polymerase chain reaction. Trop Biomed. 2014;31:728–735. [PubMed] [Google Scholar]
- 58.Filia G, Mahajan V, Bal MS, Leishangthem GD, Singh A. Seroprevalence of babesiosis and anaplasmosis in apparently healthy large ruminants of Punjab, India. Proc Natl Acad Sci India Sect B Biol Sci. 2015;85:885–888. doi: 10.1007/s40011-015-0555-4. [DOI] [Google Scholar]
- 59.Maharana BR, Kumar B, Prasad A, Patbandha TK, Sudhakar NR, Joseph JP, Patel BR. Prevalence and assessment of risk factors for haemoprotozoan infections in cattle and buffaloes of South-West Gujarat India. Indian J Anim Res. 2016;50:733–739. [Google Scholar]
- 60.Sharma A, Singla LD, Kaur P, Bal MS. PCR and ELISA vis-à-vis microscopy for detection of bovine anaplasmosis: a study on associated risk of an upcoming problem in North India. Sci World J. 2015;2015:352519. doi: 10.1155/2015/352519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kaur P, Juyal PD, Sharma A, Bal MS, Singla LD. Seroprevalence of Babesia bigemina in dairy animals from low lying regions of Punjab India. Indian J Anim Res. 2016;50:406–410. doi: 10.18805/ijar.10716. [DOI] [Google Scholar]
- 62.Krishnamurthy CM, Ananda KJ, Adeppa J. Prevalence of Haemoprotozoan infections in bovines of Shimoga region of Karnataka state. J Parasit Dis. 2016;40:890–892. doi: 10.1007/s12639-014-0599-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sharma A, Singla LD, Batth BK, Kaur P. Clinicopatho-biochemical alterations associated with subclinical babesiosis in dairy animals. J Arthropod Borne Dis. 2016;10:258–266. [PMC free article] [PubMed] [Google Scholar]
- 64.Khorajiya J, Srivastava MK, Kumar R, Panigrahi PN. Hospital based prevalence of haemoprotozoan infections in cattle and buffalo during summer season in Mathura region of Uttar Pradesh. Vet Pract. 2017;18:204–206. [Google Scholar]
- 65.Maharana BR, Kumar B, Joseph JP, Patbandha TK. A comparative analysis of microscopy and PCR based detection methods for Babesia and Trypanosoma infecting bovines and assessment of risk factors. Indian J Anim Res. 2019;53:382–387. [Google Scholar]
- 66.Kumar N, Solanki JB, Varghese A, Jadav MM, Das B, Patel MD, Patel DC. Molecular assessment of Anaplasma marginale in bovine and Rhipicephalus (Boophilus) microplus tick of endemic tribal belt of coastal South Gujarat. India Acta Parasitol. 2019;64:700–709. doi: 10.2478/s11686-019-00041-z. [DOI] [PubMed] [Google Scholar]
- 67.Patil NA, Satbige AS. Molecular detection, haematological and therapeutic studies on theileriosis in buffaloes. Buffalo Bull. 2019;38:141–146. [Google Scholar]
- 68.Keshavamurthy R, Singh BB, Kalambhe DG, Aulakh RS, Dhand NK. Prevalence of Coxiella burnetii in cattle and buffalo populations in Punjab India. Prev Vet Med. 2019;166:16–20. doi: 10.1016/j.prevetmed.2019.03.003. [DOI] [PubMed] [Google Scholar]
- 69.Keshavamurthy R, Singh BB, Kalambhe DG, Aulakh RS, Dhand NK. Identification of risk factors associated with Coxiella burnetii infection in cattle and buffaloes in India. Prev Vet Med. 2020;181:105081. doi: 10.1016/j.prevetmed.2020.105081. [DOI] [PubMed] [Google Scholar]
- 70.Dhaka P, Malik SVS, Yadav JP, Kumar M, Barbuddhe SB, Rawool DB. Apparent prevalence and risk factors of coxiellosis (Q fever) among dairy herds in India. PLoS ONE. 2020;15:e0239260. doi: 10.1371/journal.pone.0239260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kaur P, Sharma A, Juyal PD, Bal MS, Singh C, Singla LD. Comparative epidemiology and pathophysiology of patent and latent babesiosis caused by Babesia bigemina in buffaloes and cattle from different agroclimatic zones of Punjab state. India Trop Anim Health Prod. 2021;53:264. doi: 10.1007/s11250-021-02717-z. [DOI] [PubMed] [Google Scholar]
- 72.Sarangi LN, Rana SK, Prasad A, Ponnanna NM, Sharma GK. Prevalence of antibodies to Anaplasma in cattle and buffaloes of different organized herds in India. J Parasit Dis. 2021;45:359–365. doi: 10.1007/s12639-020-01312-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kumar S, Paliwal S, Sudan V, Shanker D, Singh SK. An endpoint visualization loop-mediated isothermal amplification (LAMP) for detecting bubaline theileriosis. Beni-Suef Univ J Basic Appl Sci. 2022;11:67. doi: 10.1186/s43088-022-00251-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ntesang K, Singla LD, Kaur P, Arora JS, Kashyap N. Molecular epidemiology, phylogenetic analysis and risk assessment of Anaplasma marginale from naturally infected bovines of Punjab (India) Acta Trop. 2022;232:106499. doi: 10.1016/j.actatropica.2022.106499. [DOI] [PubMed] [Google Scholar]
- 75.Narimani B, Hoghooghi-Rad N, Shayan P, Rahbari S. Molecular and microscopic detection of Theileria spp. among cattle and buffaloes in West Azarbaijan. Iran. Arch Razi Inst. 2017;72:189–195. doi: 10.22092/ari.2017.111605. [DOI] [PubMed] [Google Scholar]
- 76.Khademi P, Ownagh A, Mardani K, Khalili M. Prevalence of Coxiella burnetii in milk collected from buffalo (water buffalo) and cattle dairy farms in Northwest of Iran. Comp Immunol Microbiol Infect Dis. 2019;67:101368. doi: 10.1016/j.cimid.2019.101368. [DOI] [PubMed] [Google Scholar]
- 77.Nikvand AA, Besati EH, Gharibi D, Jalali SM. Molecular and hematologic survey on Anaplasma marginale Infection in Slaughtered Water Buffaloes (Bubalous bubalis) in Ahvaz City. Iran. J Vet Res. 2020;75:192–199. [Google Scholar]
- 78.Ateaa R, Alkhaled M. Microscopic identification, molecular and phylogenetic analysis of Babesia species in buffalo from slaughterhouse in Al-Najaf city of Iraq. Iraqi J Vet Sci. 2019;33:251–258. doi: 10.33899/ijvs.2019.162882. [DOI] [Google Scholar]
- 79.Douangngeun B, Theppangna W, Soukvilay V, Senaphanh C, Phithacthep K, Phomhaksa S, et al. Seroprevalence of Q fever, brucellosis, and blue tongue in selected provinces in lao people's democratic republic. Am J Trop Med Hyg. 2016;95:558–561. doi: 10.4269/ajtmh.15-0913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Koh FX, Panchadcharam C, Sitam FT, Tay ST. Molecular investigation of Anaplasma spp. in domestic and wildlife animals in Peninsular Malaysia. Vet Parasitol Reg Stud Reports. 2018;13:141–147. doi: 10.1016/j.vprsr.2018.05.006. [DOI] [PubMed] [Google Scholar]
- 81.Bawm S, Sagara R, Kakisaka K, Thu MJ, Hmoon MM, Htun LL, et al. Identification, genetic variation, and structural analysis of 18S rRNA of Theileria orientalis and Theileria velifera-like isolates from Myanmar. Parasitol Int. 2021;82:102299. doi: 10.1016/j.parint.2021.102299. [DOI] [PubMed] [Google Scholar]
- 82.Rajput ZI, Hu SH, Arijo AG, Habib M, Khalid M. Comparative study of Anaplasma parasites in tick carrying buffaloes and cattle. J Zhejiang Univ Sci B. 2005;6:1057–1062. doi: 10.1631/jzus.2005.B1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Durrani AZ, Ahmad M, Ashraf M, Khan MS, Khan JA, Kamal N, Mumtaz N. Prevalence of theileriosis in buffaloes and detection through blood smear examination and polymerase chain reaction test in district Lahore. J Anim Plant Sci. 2008;18:59–62. [Google Scholar]
- 84.Khan MK, He L, Hussain A, Azam S, Zhang WJ, Wang LX, et al. Molecular epidemiology of Theileria annulata and identification of 18S rRNA gene and ITS regions sequences variants in apparently healthy buffaloes and cattle in Pakistan. Infect Genet Evol. 2013;13:124–132. doi: 10.1016/j.meegid.2012.09.007. [DOI] [PubMed] [Google Scholar]
- 85.Zulfiqar S, Shahnawaz S, Ali M, Bhutta AM, Iqbal S, Hayat S, et al. Detection of Babesia bovis in blood samples and its effect on the hematological and serum biochemical profile in large ruminants from Southern Punjab Asian Pac. J Trop Biomed. 2012;2:104–108. doi: 10.1016/S2221-1691(11)60202-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Ashraf QU, Khan AU, Khattak RM, Ali M, Shaikh RS, Iqbal F. A report on the high prevalence of Anaplasma sp. in buffaloes from two provinces in Pakistan. Ticks Tick Borne Dis. 2013;4:395–398. doi: 10.1016/j.ttbdis.2013.04.001. [DOI] [PubMed] [Google Scholar]
- 87.Farooqi SH, Ijaz M, Rashid MI, Aqib AI, Ahmad Z, Saleem MH, et al. Molecular epidemiology of Babesia bovis in bovine of Khyber Pakhtunkhwa. Pakistan Pak Vet J. 2017;37:275–280. [Google Scholar]
- 88.Gebrekidan H, Abbas T, Wajid M, Ali A, Gasser RB, Jabbar A. Molecular characterisation of Theileria orientalis in imported and native bovines from Pakistan. Infect Genet Evol. 2017;47:19–25. doi: 10.1016/j.meegid.2016.11.005. [DOI] [PubMed] [Google Scholar]
- 89.Farooqi SH, Ijaz M, Rashid MI, Nabi H, Islam S, Aqib AI, et al. Molecular epidemiology of bovine anaplasmosis in Khyber Pakhtunkhwa. Pakistan Trop Anim Health Prod. 2018;50:1591–1598. doi: 10.1007/s11250-018-1599-2. [DOI] [PubMed] [Google Scholar]
- 90.Rafiullah A, Rahman K, Khan A, Ali A, Khan A, Sajid NK. Prevalence of Theileria Parva in large ruminants through conventional and molecular techniques in district Lakki Marwat and Peshawar (Pakistan) Sarhad J Agric. 2019;35:320–329. [Google Scholar]
- 91.Ghafar A, Koehler AV, Hall RS, Gauci CG, Gasser RB, Jabbar A. Targeted next-generation sequencing and informatics as an effective tool to establish the composition of bovine piroplasm populations in endemic regions. Microorganisms. 2020;9:21. doi: 10.3390/microorganisms9010021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Siddique RM, Sajid MS, Iqbal Z, Saqib M. Association of different risk factors with the prevalence of babesiosis in cattle and buffalos. Pak J Agric Sci. 2020;57:517–524. [Google Scholar]
- 93.Basit MA, Ijaz M, Khan JA, Ashraf K, Abbas RZ. molecular evidence and hematological profile of bovines naturally infected with Ehrlichiosis in Southern Punjab Pakistan. Acta Parasitol. 2022;67:72–78. doi: 10.1007/s11686-021-00433-0. [DOI] [PubMed] [Google Scholar]
- 94.Mohsin M, Hameed K, Kamal M, Ali A, Rafiq N, Usman T, et al. Prevalence and risk factors assessment of theileriosis in livestock of Malakand division Pakistan. J Saudi Soc Agric Sci. 2022;21:242–247. doi: 10.1016/j.sjbs.2021.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Mingala CN, Konnai S, Cruz LC, Onuma M, Ohashi K. Comparative moleculo-immunological analysis of swamp- and riverine-type water buffaloes responses. Cytokine. 2009;46:273–282. doi: 10.1016/j.cyto.2009.02.006. [DOI] [PubMed] [Google Scholar]
- 96.Herrera PCT, Viloria VV, Balbin MM, Mingala CN. Prevalence of babesiosis (Babesia bovis and Babesia bigemina) in cattle and water buffalo in Nueva Ecija, Philippines using nested polymerase chain reaction. Ann Parasitol. 2017;63:309–316. doi: 10.17420/ap6304.117. [DOI] [PubMed] [Google Scholar]
- 97.Galon EMS, Moumouni PFA, Ybañez RHD, Ringo AE, Efstratiou A, Lee SH, et al. First molecular detection and characterization of tick-borne pathogens in water buffaloes in Bohol Philippines. Ticks Tick Borne Dis. 2019;10:815–821. doi: 10.1016/j.ttbdis.2019.03.016. [DOI] [PubMed] [Google Scholar]
- 98.Galay RL, Talactac MR, Ambita-Salem BV, Chu DMM, delaCosta LMO, Salangsang CMA, et al. Molecular detection of Rickettsia spp and Coxiella burnetii in cattle, water buffalo, and Rhipicephalus (Boophilus) microplus ticks in Luzon Island of the Philippines. Trop. Med. Infect. Dis. 2020;5:54. doi: 10.3390/tropicalmed5020054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Galon EMS, Ybañez RHD, Moumouni PFA, Tumwebaze MA, Fabon RJA, Callanta MRR, et al. Molecular survey of tick-borne pathogens infecting backyard cattle and water buffaloes in Quezon province Philippines. J Vet Med Sci. 2020;82:886–890. doi: 10.1292/jvms.19-0636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Galay RL, Llaneta CR, Monreal MKFB, Armero AL, Baluyut ABD, Regino CMF, et al. Molecular Prevalence of Anaplasma marginale and Ehrlichia in domestic large ruminants and Rhipicephalus (Boophilus) microplus ticks from Southern Luzon Philippines. Front Vet Sci. 2021;8:746705. doi: 10.3389/fvets.2021.746705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Sivakumar T, Tattiyapong M, Fukushi S, Hayashida K, Kothalawala H, Silva SSP, et al. Genetic characterization of Babesia and Theileria parasites in water buffaloes in Sri Lanka. Vet Parasitol. 2014;200:24–30. doi: 10.1016/j.vetpar.2013.11.029. [DOI] [PubMed] [Google Scholar]
- 102.Zhyldyz A, Sivakumar T, Igarashi I, Gunasekara E, Kothalawala H, Silva SSP, Yokoyama N. Epidemiological survey of Anaplasma marginale in cattle and buffalo in Sri Lanka. J Vet Med Sci. 2019;81:1601–1605. doi: 10.1292/jvms.19-0242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Sivakumar T, Tuvshintulga B, Kothalawala H, Silva SS, Lan DT, Long PT, et al. Host range and geographical distribution of Babesia sp. Mymensingh Transbound Emerg Dis. 2020;67:2233–2239. doi: 10.1111/tbed.13546. [DOI] [PubMed] [Google Scholar]
- 104.Terkawi MA, Huyen NX, Shinuo C, Inpankaew T, Maklon K, Aboulaila M, et al. Molecular and serological prevalence of Babesia bovis and Babesia bigemina in water buffaloes in the northeast region of Thailand. Vet Parasitol. 2011;178:201–207. doi: 10.1016/j.vetpar.2011.01.041. [DOI] [PubMed] [Google Scholar]
- 105.Bai Y, Malania L, Alvarez Castillo D, Moran D, Boonmar S, Chanlun A, et al. Global distribution of Bartonella infections in domestic bovine and characterization of Bartonella bovis strains using multi-locus sequence typing. PLoS ONE. 2013;8:e80894. doi: 10.1371/journal.pone.0080894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Junsiri W, Watthanadirek A, Poolsawat N, Kaewmongkol S, Jittapalapong S, et al. Molecular detection and genetic diversity of Anaplasma marginale based on the major surface protein genes in Thailand. Acta Trop. 2020;205:105338. doi: 10.1016/j.actatropica.2020.105338. [DOI] [PubMed] [Google Scholar]
- 107.Nguyen AH, Tiawsirisup S, Kaewthamasorn M. Molecular detection and genetic characterization of Anaplasma marginale and Anaplasma platys-like (Rickettsiales: Anaplasmataceae) in water buffalo from eight provinces of Thailand. BMC Vet Res. 2020;16:380. doi: 10.1186/s12917-020-02585-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Nguyen AH, Tiawsirisup S, Kaewthamasorn M. Low level of genetic diversity and high occurrence of vector-borne protozoa in water buffaloes in Thailand based on 18S ribosomal RNA and mitochondrial cytochrome b genes. Infect Genet Evol. 2020;82:104304. doi: 10.1016/j.meegid.2020.104304. [DOI] [PubMed] [Google Scholar]
- 109.Boonmar S, Saengsawang P, Mitsuwan W, Panjai D, Kidsin K, Sansamur C, Wichianrat I. The first report of the seroprevalence of antibodies against Bartonella spp. in water buffaloes (Bubalus bubalis) from South Thailand. Vet World. 2021;14:3144–3148. doi: 10.14202/vetworld.2021.3144-3148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Kidsin K, Panjai D, Boonmar S. The first report of seroprevalence of Q fever in water buffaloes (Bubalus bubalis) in Phatthalung. Thailand Vet World. 2021;14:2574–2578. doi: 10.14202/vetworld.2021.2574-2578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Kaewhom P, Srikijkasemwat K. A molecular survey of Theileria spp. in ruminants in the Thailand-Cambodia border region. Technology. 2022;18:205–214. [Google Scholar]
- 112.Okur-Gumusova S, Tamer C, Ozan E, Cavunt A, Kadi H, Muftuoglu B, et al. An investigation of the seroprevalence of crimean-congo hemorrhagic fever and lumpy skin disease in domesticated water buffaloes in Northern Turkey. Trop Biomed. 2020;37:165–173. [PubMed] [Google Scholar]
- 113.Khukhuu A, Lan DTB, Long PT, Ueno A, Li Y, Luo Y, et al. Molecular epidemiological survey of Theileria orientalis in Thua Thien Hue province Vietnam. J Vet Med Sci. 2011;73:701–705. doi: 10.1292/jvms.10-0472. [DOI] [PubMed] [Google Scholar]
- 114.Sivakumar T, Lan DTB, Long PT, Yoshinari T, Tattiyapong M, Guswanto A, et al. PCR detection and genetic diversity of bovine hemoprotozoan parasites in Vietnam. J Vet Med Sci. 2013;75:1455–1462. doi: 10.1292/jvms.13-0221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Li Y, Luo Y, Cao S, Terkawi MA, Lan DT, Long PT, et al. Molecular and seroepidemiological survey of Babesia bovis and Babesia bigemina infections in cattle and water buffaloes in the central region of Vietnam. Trop Biomed. 2014;31:406–413. [PubMed] [Google Scholar]
- 116.Yokoyama N, Sivakumar T, Tuvshintulga B, Hayashida K, Igarashi I, Inoue N, et al. Genetic variations in merozoite surface antigen genes of Babesia bovis detected in Vietnamese cattle and water buffaloes. Infect Genet Evol. 2015;30:288–295. doi: 10.1016/j.meegid.2014.12.035. [DOI] [PubMed] [Google Scholar]
- 117.Weerasooriya G, Sivakumar T, Lan DTB, Long PT, Takemae H, Igarashi I, et al. Epidemiology of bovine hemoprotozoa parasites in cattle and water buffalo in Vietnam. J Vet Med Sci. 2016;78:1361–1367. doi: 10.1292/jvms.16-0099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Hrnková J, Golovchenko M, Musa AS, Needham T, Italiya J, Ceacero F, et al. Borrelia spirochetes in European exotic farm animals. Front Vet Sci. 2022;9:996015. doi: 10.3389/fvets.2022.996015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Hornok S, Sugár L, Horváth G, Kovács T, Micsutka A, Gönczi E, et al. Evidence for host specificity of Theileria capreoli genotypes in cervids. Parasit Vectors. 2017;10:473. doi: 10.1186/s13071-017-2403-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Hornok S, Sugár L, Fernández de Mera IG, De La Fuente J, Horváth G, Kovács T, et al. Tick-and fly-borne bacteria in ungulates: the prevalence of Anaplasma phagocytophilum, haemoplasmas and rickettsiae in water buffalo and deer species in Central Europe Hungary. BMC Vet Res. 2018;14:98. doi: 10.1186/s12917-018-1403-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Perugini AG, Capuano F, Esposito A, Marianelli C, Martucciello A, Iovane G, Galiero G. Detection of Coxiella burnetii in buffaloes aborted fetuses by IS111 DNA amplification: a preliminary report. Res Vet Sci. 2009;87:189–191. doi: 10.1016/j.rvsc.2009.01.005. [DOI] [PubMed] [Google Scholar]
- 122.Obregón D, Corona BG, De La Fuente J, Cabezas-Cruz A, Gonçalves LR, Matos CA, et al. Molecular evidence of the reservoir competence of water buffalo (Bubalus bubalis) for Anaplasma marginale in Cuba. Vet Parasitol Reg Stud Reports. 2018;13:180–187. doi: 10.1016/j.vprsr.2018.06.007. [DOI] [PubMed] [Google Scholar]
- 123.Romero-Salas D, Mira A, Mosqueda J, García-Vázquez Z, Hidalgo-Ruiz M, Vela NAO, et al. Molecular and serological detection of Babesia bovis-and Babesia bigemina-infection in bovines and water buffaloes raised jointly in an endemic field. Vet Parasitol. 2016;217:101–107. doi: 10.1016/j.vetpar.2015.12.030. [DOI] [PubMed] [Google Scholar]
- 124.Ferreri L, Benitez D, Dominguez M, Rodriguez A, Asenzo G, Mesplet M, et al. Water buffalos as carriers of Babesia bovis in Argentina. Ann N Y Acad Sci. 2008;1149:149–151. doi: 10.1196/annals.1428.036. [DOI] [PubMed] [Google Scholar]
- 125.Corrêa FDN, Teixeira RC, Oliveira CMC, Barbosa JD, Fonseca AHD. Detection of anti-Borrelia burgdorferi antibodies in buffaloes (Bubalus bubalis) in the state of Pará. Brazil Rev Bras Parasitol Vet. 2012;21:338–341. doi: 10.1590/S1984-29612012000300033. [DOI] [PubMed] [Google Scholar]
- 126.Da Silva JB, André MR, da Fonseca AH, de Albuquerque Lopes CT, da Silva Lima DH, et al. Molecular and serological prevalence of Babesia bovis and Babesia bigemina in water buffaloes in the north region of Brazil. Vet Parasitol. 2013;197:678–681. doi: 10.1016/j.vetpar.2013.05.020. [DOI] [PubMed] [Google Scholar]
- 127.da Silva JB, Vinhote WMS, Oliveira CMC, André MR, Machado RZ, da Fonseca AH, Barbosa JD. Molecular and serological prevalence of Anaplasma marginale in water buffaloes in northern Brazil. Ticks Tick Borne Dis. 2014;5:100–104. doi: 10.1016/j.ttbdis.2013.09.007. [DOI] [PubMed] [Google Scholar]
- 128.da Silva JB, Baêta BA, Lopes CA, Manier BSML, de Castro GNS, dos Santos PN, et al. Serological prevalence and risk factors of Borrelia burgdorferi in water buffaloes (Bubalus bubalis, Linnaeus, 1758) on Marajó Island, northern region of Brazil. Rev Salud Anim. 2014;36:147–151. [Google Scholar]
- 129.da Silva JB, da Fonseca AH, Barbosa JD. Serological survey of Mycobacterium bovis, Brucella abortus and Borrelia burgdorferi in water buffaloes in the northern region of Brazil. Rev Salud Anim. 2014;36:35–39. [Google Scholar]
- 130.Silva JB, Cabezas-Cruz A, Fonseca AH, Barbosa JD, de la Fuente J. Infection of water buffalo in Rio de Janeiro Brazil with Anaplasma marginale strains also reported in cattle. Vet Parasitol. 2014;205:730–734. doi: 10.1016/j.vetpar.2014.09.009. [DOI] [PubMed] [Google Scholar]
- 131.Silva JB, Fonseca AH, Barbosa JD, Cabezas-Cruz A, De La Fuente J. Low genetic diversity associated with low prevalence of Anaplasma marginale in water buffaloes in Marajó Island. Brazil Ticks Tick Borne Dis. 2014;5:801–804. doi: 10.1016/j.ttbdis.2014.06.003. [DOI] [PubMed] [Google Scholar]
- 132.Silveira JA, de Oliveira CH, Silvestre BT, Albernaz TT, Leite RC, Barbosa JD, et al. Molecular assays reveal the presence of Theileria spp. and Babesia spp. in Asian water buffaloes (Bubalus bubalis, Linnaeus, 1758) in the Amazon region of Brazil. Ticks Tick Borne Dis. 2016;7:1017–1023. doi: 10.1016/j.ttbdis.2016.05.009. [DOI] [PubMed] [Google Scholar]
- 133.Abate HL, Santos NJRD, Brito DRB, Valente JDM, Vieira TSWJ, Garcia JL, et al. Theileria sp. in water buffaloes from Maranhão State, northeastern Brazil. Braz J Vet Parasitol. 2018;27:593–596. doi: 10.1590/s1984-296120180075. [DOI] [PubMed] [Google Scholar]
- 134.Jaimes-Dueñez J, Triana-Chávez O, Mejía-Jaramillo AM. Genetic, host and environmental factors associated with a high prevalence of Anaplasma marginale. Ticks Tick Borne Dis. 2018;9:1286–1295. doi: 10.1016/j.ttbdis.2018.05.009. [DOI] [PubMed] [Google Scholar]
- 135.Jaimes-Dueñez J, Triana-Chávez O, Holguín-Rocha A, Tobon-Castaño A, Mejía-Jaramillo AM. Molecular surveillance and phylogenetic traits of Babesia bigemina and Babesia bovis in cattle (Bos taurus) and water buffaloes (Bubalus bubalis) from Colombia. Parasit Vectors. 2018;11:510. doi: 10.1186/s13071-018-3091-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Schnittger L, Rodriguez AE, Florin-Christensen M, Morrison DA. Babesia: a world emerging. Infect Genet Evol. 2012;12:1788–1809. doi: 10.1016/j.meegid.2012.07.004. [DOI] [PubMed] [Google Scholar]
- 137.Mans BJ, Pienaar R, Latif AA. A review of Theileria diagnostics and epidemiology. Int J Parasitol Parasites Wildl. 2015;4:104–118. doi: 10.1016/j.ijppaw.2014.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Schnittger L, Ganzinelli S, Bhoora R, Omondi D, Nijhof AM, Florin-Christensen M. The Piroplasmida Babesia, Cytauxzoon, and Theileria in farm and companion animals: species compilation, molecular phylogeny, and evolutionary insights. Parasitol Res. 2022;121:1207–1245. doi: 10.1007/s00436-022-07424-8. [DOI] [PubMed] [Google Scholar]
- 139.Schreeg ME, Marr HS, Tarigo JL, Cohn LA, Bird DM, Scholl EH, et al. Mitochondrial genome sequences and structures aid in the resolution of Piroplasmida phylogeny. PLoS ONE. 2016;11:e0165702. doi: 10.1371/journal.pone.0165702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Mans BJ. The basis of molecular diagnostics for piroplasmids: Do the sequences lie? Ticks Tick Borne Dis. 2022;13:101907. doi: 10.1016/j.ttbdis.2022.101907. [DOI] [PubMed] [Google Scholar]
- 141.Uilenberg G, Gray J, Kahl O. Research on Piroplasmorida and other tick-borne agents: are we going the right way? Ticks Tick Borne Dis. 2018;9:860–863. doi: 10.1016/j.ttbdis.2018.03.005. [DOI] [PubMed] [Google Scholar]
- 142.Mosqueda J, Olvera-Ramirez A, Aguilar-Tipacamu G, Canto J, G. Current advances in detection and treatment of babesiosis. Curr Med Chem. 2012;19:1504–1518. doi: 10.2174/092986712799828355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Alvarez JA, Rojas C, Figueroa JV. Diagnostic tools for the identification of Babesia sp in persistently infected cattle. Pathogens. 2019;8:143. doi: 10.3390/pathogens8030143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Almería S, Castella J, Ferrer D, Ortuno A, Estrada-Peña A, Gutierrez JF. Bovine piroplasms in Minorca (Balearic Islands, Spain): a comparison of PCR-based and light microscopy detection. Vet Parasitol. 2001;99:249–259. doi: 10.1016/S0304-4017(01)00464-2. [DOI] [PubMed] [Google Scholar]
- 145.Bishop R, Sohanpal B, Kariuki DP, Young AS, Nene V, Baylis H, et al. Detection of a carrier state in Theileria parva-infected cattle by the polymerase chain reaction. Parasitology. 1992;104:215–232. doi: 10.1017/S0031182000061655. [DOI] [PubMed] [Google Scholar]
- 146.Fahrimal Y, Goff WL, Jasmer DP. Detection of Babesia bovis carrier cattle by using polymerase chain reaction amplification of parasite DNA. J Clin Microbiol. 1992;30:1374–1379. doi: 10.1128/jcm.30.6.1374-1379.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Figueroa JV, Chieves LP, Johnson GS, Buening G. Detection of Babesia bigemina-infected carriers by polymerase chain reaction amplification. J Clin Microbiol. 1992;30:2576–2582. doi: 10.1128/jcm.30.10.2576-2582.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.d'Oliveira C, Van Der Weide M, Habela MA, Jacquiet P, Jongejan F. Detection of Theileria annulata in blood samples of carrier cattle by PCR. J Clin Microbiol. 1995;33:2665–2669. doi: 10.1128/jcm.33.10.2665-2669.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Jacob SS, Sengupta PP, Paramanandham K, Suresh KP, Chamuah JK, Rudramurthy GR, Roy P. Bovine babesiosis: an insight into the global perspective on the disease distribution by systematic review and meta-analysis. Vet Parasitol. 2020;283:109136. doi: 10.1016/j.vetpar.2020.109136. [DOI] [PubMed] [Google Scholar]
- 150.Gohil S, Herrmann S, Günther S, Cooke BM. Bovine babesiosis in the 21st century: advances in biology and functional genomics. Int J Parasitol. 2013;43:125–132. doi: 10.1016/j.ijpara.2012.09.008. [DOI] [PubMed] [Google Scholar]
- 151.Sivakumar T, Tuvshintulga B, Otgonsuren D, Batmagnai E, Ahedor B, Kothalawala H, et al. Phylogenetic analyses of the mitochondrial, plastid, and nuclear genes of Babesia sp Mymensingh and its naming as Babesia naoakii n sp. Parasit Vectors. 2022;15:299. doi: 10.1186/s13071-022-05374-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Jabbar A, Abbas T, Sandhu ZUD, Saddiqi HA, Qamar MF, Gasser RB. Tick-borne diseases of bovines in Pakistan: major scope for future research and improved control. Parasit Vectors. 2015;8:283. doi: 10.1186/s13071-015-0894-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Gubbels MJ, Yin H, Bai Q, Liu G, Nijman IJ, Jongejan F. The phylogenetic position of the Theileria buffeli group in relation to other Theileria species. Parasitol Res. 2002;88:S28–S32. doi: 10.1007/s00436-001-0566-3. [DOI] [PubMed] [Google Scholar]
- 154.Sivakumar T, Hayashida K, Sugimoto C, Yokoyama N. Evolution and genetic diversity of Theileria. Infect Genet Evol. 2014;27:250–263. doi: 10.1016/j.meegid.2014.07.013. [DOI] [PubMed] [Google Scholar]
- 155.Gubbels MJ, Hong Y, van der Weide M, Qi B, Nijman IJ, Guangyuan L, Jongejan F. Molecular characterisation of the Theileria buffeli/orientalis group. Int J Parasitol. 2000;30:943–952. doi: 10.1016/S0020-7519(00)00074-6. [DOI] [PubMed] [Google Scholar]
- 156.Rymaszewska A, Grenda S. Bacteria of the genus Anaplasma–characteristics of Anaplasma and their vectors: a review. Vet Med. 2008;53:573–584. doi: 10.17221/1861-VETMED. [DOI] [Google Scholar]
- 157.Aubry P, Geale DW. A review of bovine anaplasmosis. Transbound Emerg Dis. 2011;58:1–30. doi: 10.1111/j.1865-1682.2010.01173.x. [DOI] [PubMed] [Google Scholar]
- 158.Kocan KM, de la Fuente J, Blouin EF, Coetzee JF, Ewing SA. The natural history of Anaplasma marginale. Vet Parasitol. 2010;167:95–107. doi: 10.1016/j.vetpar.2009.09.012. [DOI] [PubMed] [Google Scholar]
- 159.Stuen S, Granquist EG, Silaghi C. Anaplasma phagocytophilum—a widespread multi-host pathogen with highly adaptive strategies. Front Cell Infect Microbiol. 2013;3:31. doi: 10.3389/fcimb.2013.00031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Van Schaik EJ, Chen C, Mertens K, Weber MM, Samuel JE. Molecular pathogenesis of the obligate intracellular bacterium Coxiella burnetii. Nat Rev Microbiol. 2013;11:561–573. doi: 10.1038/nrmicro3049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Körner S, Makert GR, Ulbert S, Pfeffer M, Mertens-Scholz K. The prevalence of Coxiella burnetii in hard ticks in europe and their role in q fever transmission revisited—a systematic review. Front Vet Sci. 2021;8:655715. doi: 10.3389/fvets.2021.655715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Kazar J. Coxiella burnetii infection. Ann N Y Acad Sci. 2005;1063:105–114. doi: 10.1196/annals.1355.018. [DOI] [PubMed] [Google Scholar]
- 163.Yessinou RE, Katja MS, Heinrich N, Farougou S. Prevalence of Coxiella-infections in ticks-review and meta-analysis. Ticks Tick Borne Dis. 2022;13:101926. doi: 10.1016/j.ttbdis.2022.101926. [DOI] [PubMed] [Google Scholar]
- 164.Arricau-Bouvery N, Rodolakis A. Is Q fever an emerging or re-emerging zoonosis? Vet Res. 2005;36:327–349. doi: 10.1051/vetres:2005010. [DOI] [PubMed] [Google Scholar]
- 165.Guatteo R, Seegers H, Taurel AF, Joly A, Beaudeau F. Prevalence of Coxiella burnetii infection in domestic ruminants: a critical review. Vet Microbiol. 2011;149:1–16. doi: 10.1016/j.vetmic.2010.10.007. [DOI] [PubMed] [Google Scholar]
- 166.Agerholm JS. Coxiella burnetii associated reproductive disorders in domestic animals-a critical review. Acta Vet Scand. 2013;55:1–11. doi: 10.1186/1751-0147-55-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Vanderburg S, Rubach MP, Halliday JE, Cleaveland S, Reddy EA, Crump JA. Epidemiology of Coxiella burnetii infection in Africa: a one health systematic review. PLoS Negl Trop Dis. 2014;8:e2787. doi: 10.1371/journal.pntd.0002787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Tilly K, Rosa PA, Stewart PE. Biology of infection with Borrelia burgdorferi. Infect Dis Clin N Am. 2008;22:217–234. doi: 10.1016/j.idc.2007.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Rudenko N, Golovchenko M, Grubhoffer L, Oliver JR. Updates on Borrelia burgdorferi sensu lato complex with respect to public health. Ticks Tick Borne Dis. 2011;2:123–128. doi: 10.1016/j.ttbdis.2011.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Hyde JA. Borrelia burgdorferi keeps moving and carries on: a review of borrelial dissemination and invasion. Front Immunol. 2017;8:114. doi: 10.3389/fimmu.2017.00114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Billeter SA, Levy MG, Chomel BB, Breitschwerdt EB. Vector transmission of Bartonella species with emphasis on the potential for tick transmission. Med Vet Entomol. 2008;22:1–15. doi: 10.1111/j.1365-2915.2008.00713.x. [DOI] [PubMed] [Google Scholar]
- 172.Reis C, Cote M, Le Rhun D, Lecuelle B, Levin ML, Vayssier-Taussat M, Bonnet SI. Vector competence of the tick Ixodes ricinus for transmission of Bartonella birtlesii. PLoS Negl Trop Dis. 2011;5:e1186. doi: 10.1371/journal.pntd.0001186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Cheslock MA, Embers ME. Human bartonellosis: an underappreciated public health problem? Trop Med Infect Dis. 2019;4:69. doi: 10.3390/tropicalmed4020069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Wechtaisong W, Bonnet SI, Lien YY, Chuang ST, Tsai YL. Transmission of Bartonella henselae within Rhipicephalus sanguineus: data on the potential vector role of the tick. PLoS Negl Trop Dis. 2020;14:e0008664. doi: 10.1371/journal.pntd.0008664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Wechtaisong W, Bonnet SI, Chomel BB, Lien YY, Chuang ST, Tsai YL. investigation of transovarial transmission of Bartonella henselae in Rhipicephalus sanguineus sensu lato ticks using artificial feeding. Microorganisms. 2021;9:2501. doi: 10.3390/microorganisms9122501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Król N, Militzer N, Stöbe E, Nijhof AM, Pfeffer M, Kempf VA, Obiegala A. Evaluating transmission paths for three different Bartonella spp in Ixodes ricinus ticks using artificial feeding. Microorganisms. 2021;9:901. doi: 10.3390/microorganisms9050901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Esemu SN, Ndip LM, Ndip RN. Ehrlichia species, probable emerging human pathogens in sub-Saharan Africa: environmental exacerbation. Rev Environ Health. 2011;26:269–279. doi: 10.1515/REVEH.2011.034. [DOI] [PubMed] [Google Scholar]
- 178.Pfitzer S, Last R, De Waal DT. Possible death of a buffalo calf (Syncercus caffer) due to suspected heartwater (Ehrlichia ruminantium): clinical communication. J S Afr Vet Assoc. 2004;75:54–57. doi: 10.4102/jsava.v75i1.451. [DOI] [PubMed] [Google Scholar]
- 179.Gilbride C, Saunders J, Sharpe H, Maze EA, Limon G, Ludi AB, Lambe T, et al. The integration of human and veterinary studies for better understanding and management of crimean-congo haemorrhagic fever. Front Immunol. 2021;12:629636. doi: 10.3389/fimmu.2021.629636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Camp JV, Kannan DO, Osman BM, Shah MS, Howarth B, Khafaga T, et al. Crimean-Congo hemorrhagic fever virus endemicity in United Arab Emirates, 2019. Emerg Infect Dis. 2020;26:1019–1021. doi: 10.3201/eid2605.191414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Lado S, Futas J, Plasil M, Loney T, Weidinger P, Camp JV, et al. Crimean-congo hemorrhagic fever virus past infections are associated with two innate immune response candidate genes in dromedaries. Cells. 2022;11:8. doi: 10.3390/cells11010008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Nasirian H. Crimean-congo hemorrhagic fever (CCHF) seroprevalence: a systematic review and meta-analysis. Acta Trop. 2019;196:102–120. doi: 10.1016/j.actatropica.2019.05.019. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1. Study characteristics of tick-borne pathogen surveys in buffaloes worldwide.
Additional file 2: Figure S1. Forest plot diagrams for random effects in the meta-analysis of the prevalence of Babesia spp. infections in buffaloes worldwide. The middle point of each line indicates the prevalence, while the length of the line is the 95% confidence interval for each study. Diamonds refers to the prevalence in accordance with detection methods.
Additional file 3: Figure S2. Forest plot diagrams for random effects in the meta-analysis of the prevalence of Theileria spp. infections in buffaloes worldwide. The middle point of each line indicates the prevalence, while the length of the line is the 95% confidence interval of each study. Diamonds refers to the prevalence in accordance with detection methods.
Additional file 4: Figure S3. Forest plot diagrams for random effects in the meta-analysis of the prevalence of Anaplasma spp. infections in buffaloes worldwide. The middle point of each line indicates the prevalence, while the length of the line is the 95% confidence interval of each study. Diamonds refers to the prevalence in accordance with detection methods.
Data Availability Statement
All data generated or analysed during this study are included in this published article and its supplementary information files.