

Abstract
A changing climate is altering ecosystem carbon dynamics with consequences for natural systems and human economies, but there are few tools available for land managers to meaningfully incorporate carbon trajectories into planning efforts. To address uncertainties wrought by rapidly changing conditions, many practitioners adopt resistance and resilience as ecosystem management goals, but these concepts have proven difficult to monitor across landscapes. Here, we address the growing need to understand and plan for ecosystem carbon with concepts of resistance and resilience. Using time series of carbon fixation (n = 103), we evaluate forest management treatments and their relative impacts on resistance and resilience in the context of an expansive and severe natural disturbance. Using subalpine spruce–fir forest with a known management history as a study system, we match metrics of ecosystem productivity (net primary production, g C m−2 year−1) with site‐level forest structural measurements to evaluate (1) whether past management efforts impacted forest resistance and resilience during a spruce beetle (Dendroctonus rufipennis) outbreak, and (2) how forest structure and physiography contribute to anomalies in carbon trajectories. Our analyses have several important implications. First, we show that the framework we applied was robust for detecting forest treatment impacts on carbon trajectories, closely tracked changes in site‐level biomass, and was supported by multiple evaluation methods converging on similar management effects on resistance and resilience. Second, we found that stand species composition, site productivity, and elevation predicted resistance, but resilience was only related to elevation and aspect. Our analyses demonstrate application of a practical approach for comparing forest treatments and isolating specific site and physiographic factors associated with resistance and resilience to biotic disturbance in a forest system, which can be used by managers to monitor and plan for both outcomes. More broadly, the approach we take here can be applied to many scenarios, which can facilitate integrated management and monitoring efforts.
Keywords: ecosystem function, forest carbon, forest disturbance, forest management, spruce beetle
INTRODUCTION
Climate change continues to increase both the magnitude and variability of disturbances globally, with consequences for ecosystem carbon storage (Seidl et al., 2016) and subsequent patterns of radiative forcing (O'Halloran et al., 2012). Quantifying and predicting changes in carbon fixation rates due to changing disturbance regimes is important for many reasons, but particularly in relation to how policy and management actions effect ecosystem services (Morris et al., 2018). Carbon sequestration for control of greenhouse gas emissions has emerged as a key focus in the forestry sector (Miner et al., 2014), and the stabilization of forest carbon is a central concern in climate change mitigation policy (Lamb et al., 2021). For instance, recent changes to international greenhouse gas guidelines charge land management agencies with formative and summative assessment of forest carbon inventories (IPCC, 2019); paramount to achieving this is appropriate management of forest productivity. Many ecologists use “resistance” and “resilience” as key principles to track shifts in productivity at ecosystem scales relative to a variety of biotic and abiotic factors (e.g., Gunderson, 2000; Holling, 1996; Isbell et al., 2015; Peterson et al., 1998). However, these concepts are challenging to apply to the evaluation and monitoring of forest management outcomes from a systems‐thinking perspective (Yi & Jackson, 2021), especially in the context of subsequent natural disturbances (Hurteau et al., 2019) but could be broadly important for federal carbon sequestration goals.
Ecosystem resistance is historically defined as the degree to which process variables of interest depart from stable equilibria due to disturbance (Grimm & Wissel, 1997). As such, resistance is measured in terms of the magnitude of change in some ecosystem process variables (Isbell et al., 2015), whereas resilience is defined by the rate at which process variables recover from disturbance to an equilibrium or pre‐disturbance state (Holling, 1973). More recently, both concepts are generally considered in terms of shifts in the structure, composition, or demography of dominant vegetation, as well as transitions between ecological communities and the ability to retain adaptive capacity (Angeler & Allen, 2016; Coop et al., 2020; Johnstone et al., 2016). For example, in a forest landscape “resistance” may be characterized by rate or extent of mortality of a dominant tree species, or by shifts in tree or herbaceous species composition following disturbance (Jactel et al., 2017; Larson et al., 2013; Stevens‐Rumann & Morgan, 2016). In the same system, “resilience” might be quantified by tree regeneration rates (e.g., Stevens‐Rumann & Morgan, 2016) or a rapid return to a similar forest structure, canopy cover, and species composition (DeRose & Long, 2014; Yi & Jackson, 2021).
Consequently, the evaluation of resistance and resilience concepts is often considered in terms of population vital rates of dominant vegetation. For instance, a discussion of resistant or resilient forest structures must consider differences in forest cover type, physiography, climate and disturbance regimes, and the inevitable question “Resistant or resilient to what?” Acknowledging this variability recognizes that conditions consistent with ecological resilience vary widely across ecosystems and should relate to specific management objectives and disturbances. To apply the concepts of resistance and resilience for ecosystem management (e.g., Millar et al., 2007), it is useful to consider process variables that are quantifiable within and across ecosystems and are consistent across different forest and vegetation types (Yi & Jackson, 2021). Large‐scale process variables can be replicated across systems using remote platforms; this synoptic approach has several advantages, including limiting observer bias, consistency in data collection methods, and rapid evaluation of ecosystem management techniques or responses to disturbance.
Net primary productivity (NPP) is one such ecosystem process variable that can be readily extracted across large spatial scales using remotely sensed data (Running et al., 2004) and directly reflects key biological activity and carbon dynamics. In terrestrial ecosystems, NPP typically refers to the per unit area conversion of CO2 by vascular plants into plant dry matter through the process of photosynthesis, accounting for loss of carbon‐fixation due to cellular respiration processes (Roxburgh et al., 2005). By monitoring NPP, inferences can be made about the productivity of dominant vegetation (and resultant carbon storage or fixation dynamics) without relying on a detailed description of population‐level structure or demography. In addition, patterns in NPP can be described in terms of objective functions based on interannual variation. For example, change over a single period or time‐step in a variable of interest can be estimated by displacement from a steady state and relates to relative ecosystem resistance in response to perturbation, a large negative change in NPP is consistent with loss of productivity due to disturbance. In contrast, resilience is estimable by a rate of change and can be used to model NPP trajectories following management or disturbance. These concepts can be respectively represented as the maximum displacement from the equilibrium (dNPPmax) and the rate of change (dy/dt or slope; αNPP) in NPP across a time series, where disturbances are represented by reasonably discrete events (Figure 1; Holling, 1973, Isbell et al., 2015, Yi & Jackson, 2021).
FIGURE 1.
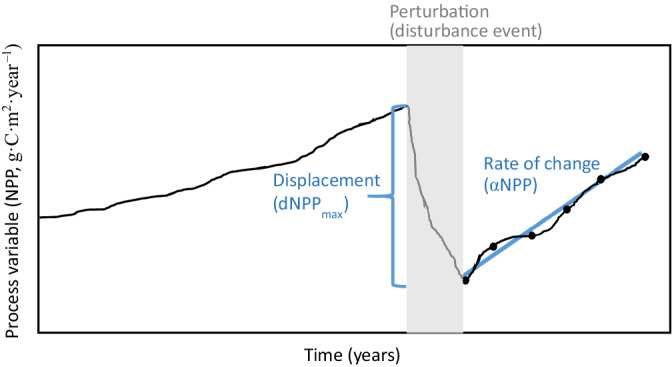
Conceptual model of ecosystem resistance and resilience to perturbation, shown as a function of carbon fixation trajectory (net primary productivity, NPP). Here, we represent the change in pre‐ and post‐disturbance NPP (dNPPmax) as the relative “resistance” to disturbance, where a greater departure of dNPPmax from zero is consistent with lower resistance to change. The y = mx + b slope of post‐disturbance NPP (αNPP) represents the “resilience” to disturbance; a steeper slope is consistent with a more rapid rate of return to pre‐disturbance productivity.
We use NPP to quantify resistance and resilience and evaluate the relative effects of two commonly implemented silvicultural systems in mediating responses to a spatially expansive and extreme disturbance across a subalpine forest landscape. In the southern Rocky Mountains, a recent outbreak of the spruce bark beetle (Dendroctonus rufipennis Kirby) resulted in the death of millions of mature trees, and some agencies estimate a loss of ~1 M ha of forest canopy cover in the state of Colorado alone due to these events (Colorado State Forest Service, 2018). The tree species primarily impacted by this event is Engelmann spruce (Picea engelmannii), a slow‐growing, shade‐tolerant conifer. There is general concern among forest managers, the scientific community, and the public about whether this outbreak will have long‐term consequences for forest growth and composition (Schapira et al., 2021).
In our study region (Figure 2), shelterwood silvicultural systems are commonly implemented and intended to increase establishment of seedling cohorts (Windmuller‐Campione et al., 2017); shelterwood is also a potential management approach for increasing resistance and resilience to bark beetle outbreaks. The shelterwood method requires a series of entries, typically (1) a preparatory cut that removes up to 10% of the stand basal area, keeping species intended for recruitment in a regular spatial arrangement and removing undesirable species, (2) a second establishment cut removes another 20%–30% is intended to prepare a seedbed and increase growing space for recruitment, and (3) an overstory removal cut to release established seedlings/saplings from competition with dominant trees (Alexander, 1987; Deal, 2018). Shelterwood treatments might therefore be associated with resistance or resilience of spruce forests to bark beetle outbreaks if pre‐outbreak reductions in tree density, competitive release of surviving trees, or the rapid regrowth of seedling cohorts following outbreak are associated with detectable losses or gains in forest productivity, respectively. Approximately 20 years prior to the outbreak, shelterwood cuts were implemented across the study landscape; at the time of the outbreak shelterwood stands were early in their sequence of treatments (i.e., only preparatory/establishment cuts had been implemented) and forest structure resembled a multi‐aged system. In addition, salvage logging was implemented throughout the region in some post‐outbreak stands to recover economic value from timber resources damaged by spruce beetle. This pre‐existing patchwork of silvicultural prescriptions (shelterwood preparatory/establishment cuts and salvage logging) with differing objectives, along with nearby non‐treated stands in wilderness areas, provides a tractable opportunity for examining how treatments interact with resistance and resilience in relation to forest carbon fixation.
FIGURE 2.
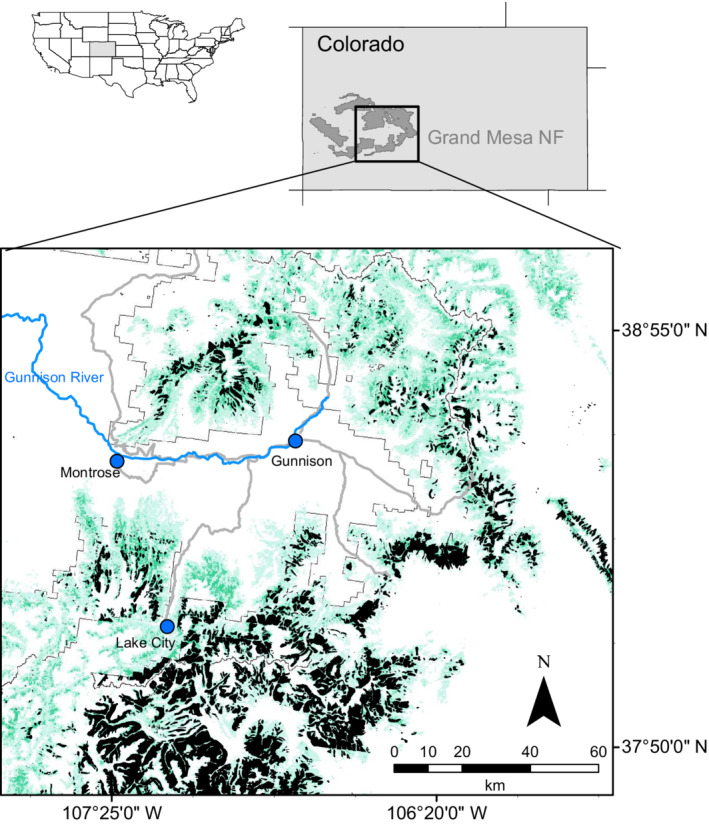
Map of the study region within the state of Colorado, USA. The study landscape was located within the Grand Mesa‐Uncompaghre‐Gunnison National Forest; green shading indicates the density of Picea engelmannii and black polygons denote aerially mapped observations of spruce beetle mortality. Gray lines show major regional roadways.
The goal of this work is to apply an approach for monitoring resistance and resilience using readily available datasets. We model variability in NPP across a 35‐year time series (1986–2019) to assess how forest management, site‐level vegetation structure, composition, and physiography impact forest resistance and resilience to a spruce beetle outbreak, applying a framework initially developed in Isbell et al. (2015) to evaluate biodiversity–productivity relationships. We address two specific objectives concerned with optimizing treatment methods: (1) we determine whether different patterns of resistance and resilience are detectable for different silvicultural treatments and (2) we characterize how variability in forest structure and physiography impact resistance and resilience metrics. The analysis framework can be readily applied by forest managers to monitor resistance and resilience as treatment outcomes and plan for forest carbon, as similar objectives are often written into forest management plans.
MATERIALS AND METHODS
Study area
Our study area is located on the Grand Mesa‐Uncompahgre‐Gunnison National Forest (GMUG) in the La Garita Mountains of southwestern Colorado (Figure 2). At elevations above ~2600 m, forest cover in the region is predominated by spruce–fir ecosystems that include Engelmann spruce (Picea engelmannii Parry ex Engelm) and subalpine fir (Abies lasiocarpa (Hooker) Nuttall) as foundation species; however, lodgepole pine (Pinus contorta Douglas), quaking aspen (Populus tremuloides Michx.), and Colorado blue spruce (Picea pungens Engelm.) are also a component of these ecosystems. Spruce beetle (D. rufipennis) is endemic to the region and populations typically occur at low densities, relying on weakened or stressed host trees for survival. However, environmental triggers such as drought, wind events, and avalanches can create high abundances of stressed trees and decouple population densities from host vigor, resulting in subsequent outbreaks (e.g., Hart et al., 2017). A large regional outbreak of spruce beetle, initiated by drought conditions, occurred in the region prior to our study. Aerial surveyors repeatedly mapped canopy tree mortality beginning in 2009–2010 and occurring through 2015–2016, though some smaller, endemic‐level patches of spruce mortality were mapped as early as 2005 (U. S. Forest Service, 2021).
Spruce–fir forests within the study area have experienced a range of forest management activities. Management history of the study area was identified with the vegetation and management history using GIS layers provided by the GMUG. Areas within the La Garita wilderness had no recorded management since Euro‐American colonization and served as control stands. Within the treated areas, the preparatory/establishment cut (hereafter, referred to as “shelterwood cuts”) was implemented in the late 1980s to early 1990s, and no further entries occurred prior to the outbreak. After spruce beetle impacted the study area, several salvage‐logging operations were implemented from 2014 to 2016 in the footprint of the 1980s/90s shelterwood stands. The salvage prescription removed most dead trees over 20 cm diameter at breast height (dbh; nearly all Engelmann spruce) but attempted to protect live trees within the stand in mature size classes, as well as trees of sapling and seedling size.
Field methodology
GIS data containing vegetation types and management history from the USFS‐GMUG was available to identify areas that were dominated by either Engelmann spruce or a mix of Engelmann spruce, subalpine fir, and aspen within the designated study area. Polygons were designated by management activity, including untreated controls, previously harvested (i.e., shelterwood cuts), or salvage logged. Plot locations were randomly generated within the different management activity polygons. At each of these points, permanent forest inventory plots were installed. At each forest inventory plot, a circular fixed‐radius (0.05 ha) sampling plot was established. Within each sampling plot, tree species, dbh, and status (live or dead) were censused. Control and previously harvested stands were measured in the summer of 2015 or 2016. Of the 103 unique plots, 53 plots were located in stands that had no history of prior treatment and were considered as “control” stands, 29 plots were located in stands treated with shelterwood cuts between 1986 and 1993, and 21 plots were located in stands treated with salvage logging in 2014 to 2016. We used field data to calculate the amount of tree mortality as a percentage of the total plot biomass, using the total aboveground biomass (percentage) of the live trees and spruce beetle‐killed trees estimated from the Fire and Fuel extension of Forest Vegetation Simulator Central Rockies variant (FVS Staff, 2008; Rebain, 2010). We then calculated spruce beetle outbreak severity as percent biomass mortality.
Remote sensing procedures
We used multitemporal NPP trajectories to develop our resilience and resistance metrics. The Numerical Terradynamic Simulation Group at the University of Montana has developed a vegetation productivity product that estimates yearly vegetation productivity at 30‐m resolution for the contiguous United States from 1986 to 2018 (Robinson et al., 2018). We downloaded the annual Landsat NPP time series data from 1986 to 2018 across the landscape of interest that experienced a spruce beetle outbreak from 2008 to 2015. The product includes annual NPP and 16 daily gross primary productivity (GPP) estimates. GPP (g C m−2 year−1) is derived using a light use efficiency model based on meteorological, Landsat data, and biome parameter lookup tables, following this equation:
| (1) |
LUE is the biome‐specific maximum light use efficiency (in g C/MJ), f Tmin and f vpd are biome‐specific temperature and vapor pressure deficit down regulators. SWrad is the incoming shortwave radiation (45% of that is available for photosynthesis), and FPAR is the fraction of photosynthetically active radiation, which is derived from the Landsat imagery. This GPP product is calculated for every 16‐day period throughout the year. Annual NPP is calculated as the total sum of the 16‐day GPP estimates within a given year minus the autotrophic respiration (Robinson et al., 2018). As the Landsat data record is inherently noisy due to atmospheric effects, sensor differences, clouds, and retrieval errors, we adopted the methods of Robinson et al. (2018) to perform a gap‐filling method, employing climatology and smoothing approaches to harmonize the satellite data record (for greater detail see Robinson et al., 2017).
To assess the resistance and resilience functions of the plot locations following spruce beetle disturbance, we developed NPP anomalies from an undisturbed reference mean. First, we randomly selected a set of 60 locations in the vicinity of our plot locations (i.e., south‐central Colorado) that did not show any mortality. These undisturbed locations were selected using aerial survey data (i.e., selection of locations that were outside areas identified as affected by the spruce beetle) and placed outside known management areas, but within Engelmann spruce dominated forest as determined by the Individual Tree Species Parameter Maps (U. S. Forest Service, 2020). Next, we plotted these reference sites over time and removed plots (n = 35) that showed a deviation from a temporal mean at the end of the time series; such locations likely experienced other unaccounted disturbances (such as harvest) or experienced spruce beetle mortality but might have been omitted by the aerial surveys.
To compare sites and calculate our resilience and resistance metrics (Figure 1), we calculated an NPP anomaly at the pixel‐level, that is, the difference of NPP from a temporal mean. The NPP anomaly (dNPP) for each plot was then calculated as follows:
| (2) |
where NPPplot,year was the NPP (g C m−2 year−1) for a given year (1986–2018) for a given plot and Mean(NPP_undisturbedplot,year) was the mean NPP for the undisturbed plots for a given year. We corrected for differences in productivity between disturbed and undisturbed plots by adjusting the anomalies with the mean difference of NPP over the five years before the outbreak event. We compared the maximum NPP reduction (dNPPmax) with the reduction of the percent biomass from field observations, and this comparison verified that the NPP data product was sensitive to the effects of the spruce beetle outbreak and reflected changes in carbon fixation due to loss of photosynthetic biomass (R 2 = 0.804, RMSE = 1195 g C m−2 year−1; Appendix S1: Figure S1).
Following the establishment of the anomalies, we calculated the maximum decrease in dNPP in a single year after the initial outbreak; we used this metric as the resistance metric, that is, the maximum reduction of productivity (dNPPmax). For most stands this occurred between 2011 and 2012. Therefore, dNPPmax reflects the degree to which the ecosystem departs from equilibrium due to a disturbance (in this case, a spruce beetle outbreak). Subsequently, we determined the year at which the maximum reduction of NPP occurred and fitted a linear model of dNPP across the years from the maximum reduction of NPP to the final year in the time series. The slope of that fitted line was analyzed as the resilience metric (Figure 1). In other words, we treat resilience as the rate (g C m−2 year−2) at which ecosystem processes recover from disturbance towards the prior steady state. These metrics where then used to assess ecosystem resistance and resilience for the plot locations and we explored how different management, environmental, and topographical characteristics affected the resilience and resistance metrics.
Data analysis
Objective 1: Determine whether different patterns of resistance and resilience are detectable for different silvicultural treatments
A three‐tiered approach was taken to evaluate responsiveness of carbon trajectories and our derived metrics to silvicultural treatments. First, we visually examined the mean productivity curves (NPP) for each treatment type to determine whether these matched with patterns of resistance and resilience predicted in Figure 1. Next, we used a one‐way ANOVA to analyze the fixed effect of silvicultural treatment (shelterwood cuts, salvage logging, and non‐treated control stands) on variance in resistance (dNPPmax) and resilience (αNPP) metrics. Since salvage logging occurred post‐outbreak, dNPPmax is only compared between non‐treated and shelterwood cut stands, but all three treatment types are used in comparisons of αNPP. Last, to validate the robustness of these results, another analysis was done to compare the empirical distribution functions of dNPPmax and αNPP across silvicultural treatments using a nonparametric Kolmogorov–Smirnov test (Kolmogorov's D statistic, Young, 1977). This second analytical approach (Kolmogorov–Smirnov test) allowed us to broaden our evaluation of differences in resistance and resilience metrics due to treatment effects by comparing the relative “shapes” of frequency distributions to one another, rather than relying solely on comparing sample means and variances.
Objective 2: Characterize how variability in forest structure and physiography impact resistance and resilience metrics
In order to analyze how variation in forest structure and physiography impact resistance and resilience in the absence of variation due to forest management history, residuals were extracted from ANOVA models testing fixed effects of forest treatment and used as the response in linear models (Appendix S1: Figure S2). A generalized linear model framework was used to analyze the relative effect sizes of forest structure (tree density per hectare, TPH; quadratic mean diameter, QMD), tree species composition (represented as percent basal area of Engelmann spruce), and site physiography (aspect, slope, elevation, heat load index; McCune & Keon, 2002, and latitude) on extracted residuals. Prior to analysis, aspect was modified using the methods of Roberts and Cooper (1989) to transform aspect from a circular aspect (ranging from 0 to 360°, were 0 and 360 are similar) to an index (trasp) ranging from 0 to 1 that reflects transition from northeasterly (coolest and wettest, value = 0) to south‐southwesterly slopes (hottest and driest, value = 1; Appendix S1: Figure S3).
We also evaluated whether mean pre‐disturbance NPP or pre‐disturbance variance in NPP were associated with forest resistance and resilience by testing two directional hypotheses: (1) the pre‐disturbance productivity of a stand mediates resistance and resilience to disturbance, with the expectation that dNPPmax will be closer to zero (i.e., a high pre‐disturbance NPP may be associated with reduced loss to overall NPP) and αNPP will be positively correlated with pre‐disturbance mean NPP and (2) high interannual variation (variance) in NPP is associated with reduced resilience. The reasoning behind this second prediction is that variance in NPP may be a signal for drought conditions (Peng et al., 2016), which are associated with a combination of complex factors including topography, aspect, and precipitation. Therefore, no single predictor variable may be associated with resilience, but a resilience signal could be detectable by year‐to‐year variation in NPP. Accordingly, the ten‐year average site NPP prior to outbreak initiation and variance in annual pre‐disturbance NPP over those 10 years were included as factors in our modeling framework.
We initially included site in our models as a random effect, but site identity accounted for a negligible proportion of residual variance in modeled responses and was subsequently removed to simplify interpretation. A Type I error rate of ɑ = 0.05 was used for assigning statistical significance to modeled effects, but effects significant at a ɑ = 0.10 level were interpreted as marginally significant. All statistical analyses were performed in the R statistical programming environment (R Core Team, 2020).
RESULTS
Objective 1: Determine whether different patterns of resistance and resilience are detectable for different silvicultural treatments
Mean NPP trajectories of study sites were generally similar between non‐treated and shelterwood cut stands from the period of 1985–2010 (Figure 3), prior to spruce beetle outbreak. In 2012–2013 mortality of canopy trees from the outbreak is observable in control and shelterwood stands, when NPP precipitously dropped as expected for all stand types, consistent with the predicted change in of the NPP signal. After 2015 there was a clear shift in the NPP signal consistent with resilience, with a trend of increasing NPP in non‐treated control stands and those treated with shelterwood cuts. However, this pattern was not the same for salvage‐logged stands, which showed a trend of ongoing reduction in NPP (Figure 3a). Resistance (dNPPmax) was similar with no significant differences among non‐treated control stands or those treated with shelterwood cuts (F 1,101 = 2.412, p = 0.124, Figure 3b). However, a different pattern was found for resilience (αNPP), where trajectories were similar (and positive) between non‐treated control and shelterwood cuts following the end of the outbreak period, but mean αNPP was reduced by fourfold on average in salvage‐logged stands and remained negative rather than showing an upward trajectory (F 2,100 = 33.162, p < 0.001, Figure 3c).
FIGURE 3.
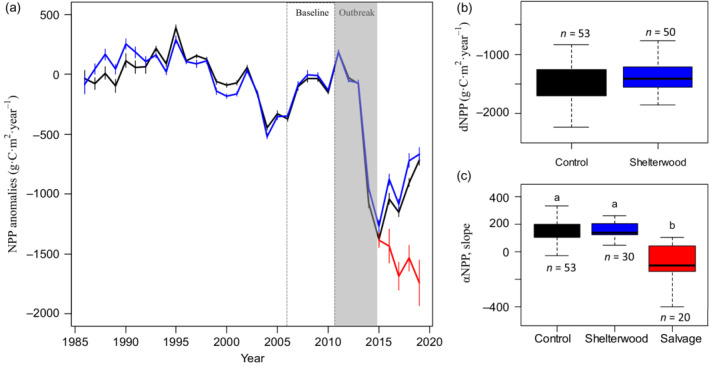
(a) Time series of mean net primary productivity (NPP) anomaly (±SE) aggregated across forest management treatments. A sharp decline in NPP is observable following initial forest mortality from spruce beetle outbreak (ca. 2011–2015), with clear differences in the recovery rate of carbon trajectories across treatments. The open box denotes the period over which baseline anomalies in NPP were computed, and the gray shading shows the period where the outbreak occurred. Analysis of variance also showed no clear differences in (b) resistance, but there were significant differences in resilience metrics (c) due to different silvicultural treatments.
Nonparametric tests (K‐S test) comparing empirical distribution functions converged on similar results for dNPPmax distributions and indicated no difference between non‐treated control stands versus those treated with shelterwood cuts (D = 0.213, p = 0.189) (Figure 4a). The empirical distributions of αNPP did not differ between non‐treated control stands and shelterwood cuts (D = 0.117, p = 0.923), but distributions of αNPP from salvage‐logged sites significantly differed from both non‐treated control stands (D = 0.754, p < 0.001) and those treated with shelterwood cuts (D = 0.793, p < 0.001) (Figure 4b).
FIGURE 4.
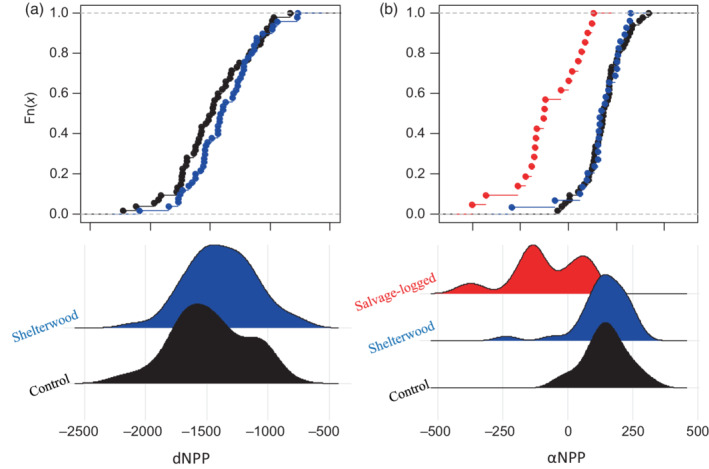
Empirical cumulative density functions of (a) resistance (dNPPmax) and (b) resilience metrics (αNPP), showing variation in the distribution of resilience (but not resistance) due to effects of forest management. NPP, net primary productivity.
Objective 2: Characterize how variability in forest structure and physiography impact resistance and resilience metrics
Once silvicultural treatment effects were accounted for, residual variance in resistance and resilience metrics were responsive to variation in elements of forest structure and physiography, but patterns of significance differed, indicating different drivers of resistance and resilience.
Our model of residual variance in resistance (dNPPmax) indicated that both stand species composition and physiographic factors were important in predicting resistance (Table 1a). Elevation had the largest overall effect size and indicated that stands at higher elevations exhibited more negative changes in NPP due to spruce beetle outbreak (β = −0.286, Figure 5a). The percent of stand basal area comprised of Engelmann spruce followed a similar pattern, though the effect size was smaller (β = −0.016, Figure 5b), and higher elevation stands were correlated with an increased spruce basal area component (Pearson's r = 0.545, p < 0.001). Stands comprised of a higher proportion of aspen basal area tended to have higher resistance than stands with little or no aspen component (Appendix S1: Figure S4). Pre‐disturbance mean NPP was also negatively related to resistance and stands with higher productivity prior to outbreak had a greater overall decline in NPP due to tree mortality (β = −0.258, Figure 5c).
TABLE 1.
Summary of a generalized linear model analysis comparing the effect sizes of forest structure, composition, and physiography on (a) resistance (dNPPmax) and (b) resilience (αNPP) of Engelmann spruce forest to a regional spruce beetle outbreak (n = 103 sample locations).
| Metric and parameter | Estimate (β) | SE | t | p |
|---|---|---|---|---|
| (a) dNPPmax | ||||
| (Intercept) | 1.479 | 0.451 | 3.277 | 0.001 |
| Tree density (stems/ha−1) | −0.025 | 0.118 | −0.216 | 0.829 |
| QMD (cm) | −0.158 | 0.130 | −1.213 | 0.228 |
| Transformed aspect | −0.323 | 0.263 | −1.225 | 0.224 |
| Slope (%) | −0.087 | 0.127 | −0.688 | 0.493 |
| Elevation (m) | −0.309 | 0.107 | −2.892 | 0.005 |
| HLI (MJ/cm2/year) | −0.110 | 0.139 | −0.787 | 0.433 |
| Spruce basal area (%) | −0.016 | 0.005 | −3.110 | 0.002 |
| Pre‐disturbance NPP mean | −0.258 | 0.088 | −2.945 | 0.004 |
| Pre‐disturbance NPP variance | 0.088 | 0.085 | 1.038 | 0.302 |
| (b) αNPP | ||||
| (Intercept) | −0.234 | 0.113 | −2.080 | 0.040 |
| Tree density (stems/ha−1) | 0.076 | 0.094 | 0.806 | 0.422 |
| QMD (cm) | 0.050 | 0.129 | 0.386 | 0.701 |
| Transformed aspect | 0.595 | 0.227 | 2.622 | 0.010 |
| Slope (%) | −0.015 | 0.108 | −0.143 | 0.887 |
| Elevation (m) | 0.162 | 0.096 | 1.683 | 0.096 |
| HLI (MJ/cm2/year) | −0.018 | 0.116 | −0.158 | 0.875 |
| Spruce basal area (%) | 0.110 | 0.137 | 0.804 | 0.424 |
| Pre‐disturbance NPP mean | 0.035 | 0.076 | 0.457 | 0.649 |
| Pre‐disturbance NPP variance | 0.072 | 0.076 | 0.941 | 0.349 |
Note: Significant (p < 0.05) and marginally significant (p < 0.10) effects are denoted in boldface text.
Abbreviations: HLI, heat‐load index; NPP, net primary productivity; QMD, quadratic mean diameter.
FIGURE 5.
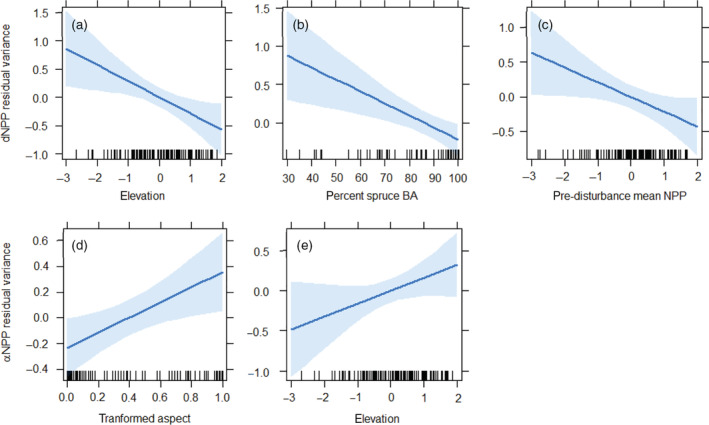
Effects plots showing linear relationships between residual variance in resistance (dNPPmax) and (a) elevation, (b) percent spruce basal area (BA), and (c) pre‐disturbance mean NPP. Effects are also shown for residual variance in resilience (αNPP) for (d) aspect and (e) elevation. Shading represents the 95% confidence interval of modeled effects. Rug plots show distributions of respective independent variables along the x‐axis. NPP, net primary productivity.
The factors that predicted residual variance in resilience (αNPP) were physiographic, and no stand structural variables were associated with resilience (Table 1b). None of the modeled parameters were significantly negatively related to resilience. Unlike the model for resistance, aspect had the largest overall effect size and stands at southwesterly trended towards recovering productivity more quickly than stands in northeasterly aspects (β = 0.595, Figure 5d); however, there was no correlation between aspect and percent spruce basal area in our dataset (Pearson's r = −0.064, p = 0.514). In addition, elevation exerted a positive effect on resilience (β = 0.162) such that stands at higher elevations recovered productivity more rapidly than lower elevation stands (Figure 5e).
DISCUSSION
Applying remotely sensed products, such as we do here with NPP, to derive resistance and resilience metrics is a useful approach for monitoring responses of forests and other systems to perturbation. Here, we model carbon trajectories over a time series to monitor responses to biotic disturbance in the context of previous forest management history. Our approach was robust to parametric (Figure 3) and nonparametric analyses (Figure 4), indicating that departure from equilibrium (resistance) and rate of change (resilience) (Figure 1) are good signals for evaluating ecosystem carbon trajectories and reliably reflect variation in forest productivity (Appendix S1: Figure S1) due to both prior management history and biotic disturbances. Our analysis expands on the work of Isbell et al. (2015) by applying a carbon dynamics framework to forest management before and after a large biotic disturbance (bark beetle outbreak) and can help practitioners to identify (and prioritize for treatment) areas of vulnerability based on forest structural and physiographic factors.
In our study system, there were no clear differences in productivity loss due to management history in stand carbon trajectories following spruce beetle; stands that were non‐treated or treated with shelterwood cut exhibited similar resistance to productivity losses. This is similar to others who examined resistance from the perspective of tree survival and regeneration using field methods (Temperli et al., 2014; Windmuller‐Campione et al., 2017). Note however, that the shelterwood cuts were implemented in the late 1980s to early 1990s, with the spruce beetle outbreak occurring approximately 25 years later. Although stand structure prior to the outbreak differed between non‐treated and managed stands (non‐treated stands had lower tree densities and higher quadratic mean diameters on average), this was not associated with differences in resistance (Table 1).
Salvage‐logged stands did not recover productivity at the same rate as non‐treated stands or shelterwood cuts and exhibited substantially reduced productivity trajectories (Figure 3). These differences have implications for forest managers concerned with controlling stand growth or carbon storage, and our approach enables comparison of ecosystem process variables across forest treatments at an ecosystem scale. Our data suggest that over short timescales (3–4 years) following spruce beetle outbreak, salvage logging treatments are likely to be associated with further losses in stand productivity, in addition to reductions in shrub and forb cover (Mattson et al., 2019). These losses to stand productivity are important for managing ecosystem carbon stocks by changing site‐level (live) carbon pools (Dobor et al., 2020), though salvage logging may also result in conversion of harvested trees to permanent products with long‐term carbon storage potential. In addition, federal directives (i.e., National Forest Management Act 1976, NFMA) require that post‐salvage stands be planted if little or no seedling recruitment is observed after five years, which has implications for recovery time. Salvage logging represents a subsequent disturbance to the stand, and it will be important to quantify if and how salvage‐logged stands eventually recover productivity equivalent to non‐treated or shelterwood cut stands over longer post‐disturbance timeframes, with and without planting seedlings.
Shelterwood cuts have been recommended by others to increase stand resilience (e.g., Temperli et al., 2014; Windmuller‐Campione et al., 2017), but our data indicate that shelterwood treatments were not different from non‐treated control stands in terms of short‐term resilience. However, over a longer time‐since‐outbreak time series, this result could potentially change as Engelmann spruce recruitment continues to increase. Future tests of management effects on resilience in this system can benefit from examining a longer post‐outbreak time series to evaluate whether carbon trajectories in shelterwood cut stands eventually outpace those of non‐treated stands.
By analyzing residual variability in carbon fixation, we were also able to consider how site‐level components, including stand structural factors and species composition, as well as physiography, impacted stand resistance and resilience in the absence of variance due to prior forest management. Such a comparison can help managers to prioritize treatments by identifying stand conditions that can be modified to alter resistance or resilience (Churchill et al., 2013). For example, we found that increased aspen basal area led to a predicted increase in resistance to spruce beetle outbreak (Appendix S1: Figure S4). Increases of non‐host basal area results in reduced forest canopy area killed during an outbreak, which has been previously observed for spruce beetle disturbances (e.g., Schmid & Frye, 1976). Managers could focus on maintaining tree species diversity across the landscape, thereby increasing the forest resistance to future beetle attacks (Fettig et al., 2007), although managers should also be aware of spill‐over effects towards non‐host species (Berthelot et al., 2021). Generally, in our study region high‐elevation sites on northerly aspects that were highly productive prior to outbreak were likely to be the least resistant to spruce beetle. One potential application of these findings is that stands fitting this description could be prioritized for manipulation of species composition (e.g., Figure 6).
FIGURE 6.
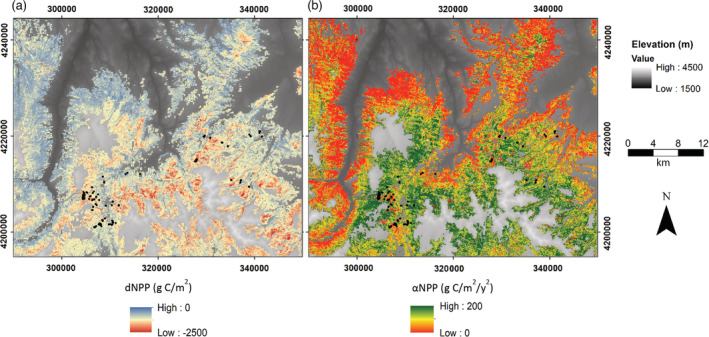
Raster products projecting metrics of (a) resistance (dNPPmax) and (b) resilience (αNPP) across the study landscape, located in the Grand Mesa‐Uncompaghre‐Gunnison National Forest. Black dots denote plots where data were collected on stand structure and composition. NPP, net primary productivity.
In contrast, responses of resilience were predicted only by physiography, which may indicate specific areas on the landscape that are likely to recover productivity rapidly following spruce beetle disturbance. The most rapid rates of productivity recovery were observed in stands at higher elevations and on southwesterly aspects. We hypothesize that this is due to a combination of two factors, specifically (1) higher elevation stands may receive more winter precipitation that buffers soil moisture across the growing season allowing for more successful germination, subsequent tree growth, and potentially enhanced species diversity and (2) southwesterly aspects are generally warmer and receive more insolation, which likely drives a longer growing season. This outcome of our modeling approach (i.e., identifying high elevation stands on southwesterly aspects as most resilient; Figure 6) may help managers to identify specific areas, based on topography, that require little intervention following biotic disturbance.
Our analysis has several implications for the management of subalpine spruce–fir forests in the southern Rocky Mountains. First, the productivity of stands treated with shelterwood did not clearly differ from non‐treated control stands prior to spruce beetle outbreak; shelterwood was also not associated with ecosystem resistance or resilience. Second, even a small aspen component is likely to improve ecosystem resistance to spruce beetle in mixed‐species stands. This is not surprising since aspen is a non‐host for spruce beetle, however, if the management goal is to preserve spruce dominance, then our data indicates managing for a species composition with ~20% non‐host basal area for above‐average stand resistance to spruce beetle (Figure 7, Millar et al., 2007). Third, salvage operations may result in reduced resilience of these systems in the years immediately following outbreak, but continued monitoring of this landscape will be important for conclusively determining effects of shelterwood and salvage logging on resilience at longer (decadal) time scales. Last, the recovery rate of productivity is strongly determined by landscape position (i.e., aspect and elevation), and implementations aimed at increasing resilience might be most effective at spruce‐dominant (near 100% basal area composition) high elevation sites, which in our landscape was areas exceeding ~3350 m (~10,900 ft). Elevation‐mediated ecosystem resilience, such as we report here, might also indicate that higher elevation areas are the most vulnerable to species turnover (i.e., “critical transition,” Johnstone et al., 2016) following large‐scale disturbance.
FIGURE 7.
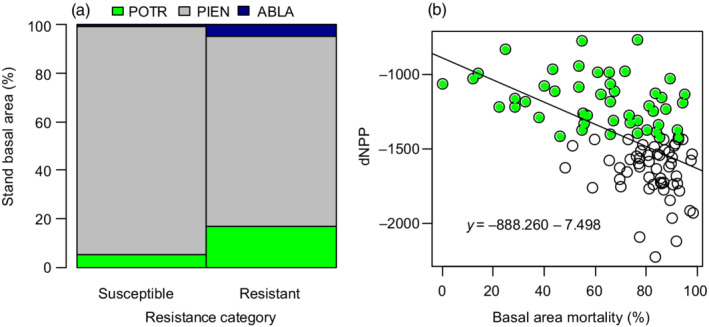
(a) Comparison of stand composition associated with two categories: “resistant” stands that were below average in dNPPmax (i.e., stands that did not have large drops in productivity, n = 49), and “susceptible” stands that were above average in dNPPmax (i.e., had a larger negative departure from the long‐term NPP average following spruce beetle outbreak, n = 54). PIEN, Engelmann spruce (Picea engelmannii); POTR, aspen (Populus tremuloides); ABLA, fir (Abies lasiocarpa). (b) A comparison of the relationship between basal area mortality and dNPPmax; green‐shaded points represent the “resistant” category shown in (a). NPP, net primary productivity.
Under a changing climate, rates of ecosystem carbon fixation are expected to diverge from historical conditions (You et al., 2020) with consequences for emissions and disturbance‐driven feedbacks to global biogeochemical cycles (Kautz et al., 2018; Kesselmeier et al., 2002; Kurz et al., 2008). As a result, there is a growing demand for policy around the management of forest carbon (Fahey et al., 2010) and a corresponding need for methods to link management practices to carbon storage. The workflow we apply here can help to meet this need using satellite‐derived carbon storage data for large landscapes, and these data can objectively quantify resistance and resilience across multiple ecosystems and disturbance types using comparable metrics. However, practitioners should be aware that ecosystem‐scale data such as those we analyze here are limited in their ability to inform small‐scale processes such as microsite variation in biodiversity or species turnover, and trends primarily reflect the growth patterns of dominant species.
CONCLUSION
We address the growing need to understand and plan for ecosystem carbon dynamics by applying a framework (after Isbell et al., 2015) that quantifies resistance and resilience as an emergent ecosystem property, irrespective of vegetation type. We used a well‐established carbon assimilation data product (Robinson et al., 2018) to model effects of a recent spruce beetle outbreak across a region with different management activities. We showed that NPP anomalies closely tracked bark beetle activity across time, and we investigated the drivers of resistance (maximum NPP deviation due to spruce beetle‐caused tree mortality) and resilience (recovery of NPP post spruce beetle disturbance) across the landscape. We found that stand species composition and physiography predicted resistance after the variance associated with non‐significant effect of prior management was accounted for, but resilience was less responsive to these factors and was only related to elevation and variation in pre‐disturbance productivity. This approach can be applied to other scenarios across different ecosystems and disturbance types (e.g., fire, drought, defoliation events) while applying consistent ecosystem‐level definitions of resilience and resistance, potentially leading to the ability to better manage forest dynamics across space and time.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
Supporting information
Appendix S1
ACKNOWLEDGMENTS
This project was supported by funds from the Western Wildland Environmental Threat Assessment Center (WWETAC; U.S. Forest Service agreement no. 18‐CS‐11221633‐142) to Mike A. Battaglia, Arjan J. H. Meddens, and Thomas S. Davis. Numerous field crews contributed to the on‐the‐ground field data collection, which was funded through partnerships with the Grand Mesa, Uncompahgre, and Gunnison National Forest, Colorado Forest Restoration Institute, Colorado State University, and U.S.D.A. Forest Service Rocky Mountain Research Station. This manuscript was written and prepared by U.S. Government employees on official time, and therefore it is in the public domain and not subject to copyright.
Davis, Thomas S. , Meddens Arjan J. H., Stevens‐Rumann Camille S., Jansen Vincent S., Sibold Jason S., and Battaglia Mike A.. 2022. “Monitoring Resistance and Resilience Using Carbon Trajectories: Analysis of Forest Management–Disturbance Interactions.” Ecological Applications 32(8): e2704. 10.1002/eap.2704
Handling Editor: Jason P. Kaye
Funding information U.S. Forest Service, Grant/Award Number: 18‐CS‐11221633‐142
DATA AVAILABILITY STATEMENT
Data (Davis et al., 2022) are available in Dryad at https://doi.org/10.5061/dryad.jwstqjqb0.
REFERENCES
- Alexander, R. 1987. “Ecology, Silviculture, and Management of the Engelmann Spruce ‐ Subalpine Fir Type in the Central and Southern Rocky Mountains.” U.S.D.A. Forest Service, Agriculture Handbook No. 659. Washington, D.C.: U.S.D.A. Forest Service.
- Angeler, D. G. , and Allen C. R.. 2016. “Quantifying Resilience.” Journal of Applied Ecology 53: 617–24. [Google Scholar]
- Berthelot, S. , Frühbrodt T., Hajek P., Nock C. A., Dormann C. F., Bauhus J., and Fründ J.. 2021. “Tree Diversity Reduces the Risk of Bark Beetle Infestation for Preferred Conifer Species, but Increases the Risk for Less Preferred Hosts.” Journal of Ecology 109: 2649–61. [Google Scholar]
- Churchill, D. J. , Larson A. J., Dahlgreen M. C., Franklin J. F., Hessburg P. F., and Lutz J. A.. 2013. “Restoring Forest Resilience: From Reference Spatial Patterns to Silvicultural Prescriptions and Monitoring.” Forest Ecology and Management 291: 442–57. [Google Scholar]
- Colorado State Forest Service . 2018. “Report on the Health of Colorado's Forests.” Colorado State Forest Service. Colorado State University, Fort Collins, Colorado. [Google Scholar]
- Coop, J. D. , Parks S. A., Stevens‐Rumann C., Crausbay S. D., Higuera P. E., Hurteau M. D., Tepley A., et al. 2020. “Wildfire‐Driven Forest Conversion of in Western North American Landscapes.” BioScience 70: 659–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis, T. S. , Meddens A. J. H., Stevens‐Rumann C. S., Jansen V. S., Sibold J. S., and Battaglia M. A.. 2022. “Monitoring Resistance and Resilience using Carbon Trajectories: Analysis of Forest Management‐Disturbance Interactions.” Dryad, Dataset. 10.5061/dryad.jwstqjqb0. [DOI] [PMC free article] [PubMed]
- Deal, R. 2018. The Dictionary of Forestry. Bethesda, MD: Society of American Foresters. [Google Scholar]
- DeRose, R. J. , and Long J. N.. 2014. “Resistance and Resilience: A Conceptual Framework for Silviculture.” Forest Science 60: 1205–12. [Google Scholar]
- Dobor, L. , Hlásny T., Rammer W., Zimová S., Barka I., and Seidl R.. 2020. “Is Salvage Logging Effectively Dampening Bark Beetle Outbreaks and Preserving Forest Carbon Stocks?” Journal of Applied Ecology 57: 67–76. [Google Scholar]
- Fahey, T. J. , Woodbury P. B., Battles J. J., Goodale C. L., Hamburg S. P., Ollinger S. V., and Woodall C. W.. 2010. “Forest Carbon Storage: Ecology, Management, and Policy.” Frontiers in Ecology and the Environment 8: 245–52. [Google Scholar]
- Fettig, C. J. , Klepzig K. D., Billings R. F., Munson A. S., Nebeker T. E., Negrón J. F., and Nowak J. T.. 2007. “The Effectiveness of Vegetation Management Practices for Prevention and Control of Bark Beetle Infestations in Coniferous Forests of the Western and Southern United States.” Forest Ecology and Management 238: 24–53. [Google Scholar]
- FVS Staff . 2008. (revised June 2021). “Central Rockies (CR) Variant Overview—Forest Vegetation Simulator.” Internal Report. Fort Collins, CO: U. S. Department of Agriculture, Forest Service, Forest Management Service Center.
- Grimm, V. , and Wissel C.. 1997. “Babel, or the Ecological Stability Discussions: An Inventory and Analysis of Terminology and a Guide for Avoiding Confusion.” Oecologia 109: 323–34. [DOI] [PubMed] [Google Scholar]
- Gunderson, L. H. 2000. “Ecological Resilience—In Theory and Application.” Annual Review of Ecology and Systematics 31: 425–39. [Google Scholar]
- Hart, S. J. , Veblen T. T., Schneider D., and Molotch N. P.. 2017. “Summer and Winter Drought Drive the Initiation and Spread of Spruce Beetle Outbreak.” Ecology 98: 2698–707. [DOI] [PubMed] [Google Scholar]
- Holling, C. S. 1973. “Resilience and Stability of Ecological Systems.” Annual Review of Ecology and Systematics 4: 1–23. [Google Scholar]
- Holling, C. S. 1996. “Engineering Resilience versus Ecological Resilience.” In Engineering within Ecological Constraints, edited by Schulze P., 31–44. Washington, DC: National Academy Press. [Google Scholar]
- Hurteau, M. D. , North M. P., Koch G. W., and Hungate B. A.. 2019. “Managing for Disturbances Stabilizes Forest Carbon.” Proceedings of the National Academy of Sciences USA 116: 10193–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- IPCC . 2019. Climate Change and Land: an IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems. in press. [Google Scholar]
- Isbell, F. , Craven D., Connolly J., Loreau M., Schmid B., Beierkuhnlein C., Bezemer T. M., et al. 2015. “Biodiversity Increases the Resistance of Ecosystem Productivity to Climate Extremes.” Nature 526: 574–7. [DOI] [PubMed] [Google Scholar]
- Jactel, H. , Bauhus J., Boberg J., Bonal D., Castagneyrol B., Gardiner B., Gonzalez‐Olabarria J. R., Koricheva J., Meurisse N., and Brockerhoff E. G.. 2017. “Tree Diversity Drives Forest Stand Resistance to Natural Disturbances.” Current Forestry Reports 3: 223–43. [Google Scholar]
- Johnstone, J. F. , Allen C. D., Franklin J., Frelich L. E., Harvey B. J., Higuera P. E., et. al. 2016. “Changing Disturbance Regimes, Ecological Memory, and Forest Resilience.” Frontiers in Ecology and the Environment 14: 369–78. [Google Scholar]
- Kautz, M. , Anthoni P., Meddens A. J. H., Pugh T. A. M., and Arneth A.. 2018. “Simulating the Recent Impacts of Multiple Biotic Disturbances on Forest Carbon Cycling across the United States.” Global Change Biology 24: 2079–92. [DOI] [PubMed] [Google Scholar]
- Kesselmeier, J. , Ciccioli P., Kuhn U., Stefani P., Biesenthal T., Rottenberger S., Wolf A., et al. 2002. “Volatile Organic Compound Emissions in Relation to Plant Carbon Fixation and the Terrestrial Carbon Budget.” Global Biogeochemical Cycles 16: 73–9. [Google Scholar]
- Kurz, W. A. , Dymond C. C., Stinson G., Rampley G. J., Neilson E. T., Carroll A. L., Ebata T., and Safranyik L.. 2008. “Mountain Pine Beetle and Forest Carbon Feedback to Climate Change.” Nature 452: 987–90. [DOI] [PubMed] [Google Scholar]
- Lamb, R. L. , Hurtt G. C., Boudreau T. J., Campbell E., Sepúlveda Carlo E. A., Chu H.‐H., de Mooy J., et al. 2021. “Context and Future Directions for Integrating Forest Carbon into Sub‐national Climate Mitigation Planning in the RGGI Region of the U.S.” Environmental Research Letters 16: 063001. [Google Scholar]
- Larson, A. J. , Belote T. R., Cansler C. A., Parks S. A., and Dietz M. S.. 2013. “Latent Resilience in Ponderosa Pine Forest: Effects of Resumed Frequent Fire.” Ecological Applications 23: 1243–9. [DOI] [PubMed] [Google Scholar]
- Mattson, L. R. , Coop J. D., Battaglia M. A., Cheng A. S., Sibold J. S., and Viner S.. 2019. “Post‐spruce Beetle Timber Salvage Drives Short‐term Surface Fuel Increases and Understory Vegetation Shifts.” Forest Ecology and Management 437: 348–59. [Google Scholar]
- McCune, B. , and Keon D.. 2002. “Equations for Potential Annual Direct Incident Radiation and Heat Load.” Journal of Vegetation Science 13: 603–6. [Google Scholar]
- Millar, C. I. , Stephenson N. L., and Stephens S. L.. 2007. “Climate Change and Forests of the Future: Managing Inface of Uncertainty.” Ecological Applications 17: 2145–51. [DOI] [PubMed] [Google Scholar]
- Miner, R. A. , Abt R. C., Bowyer J. L., Buford M. A., Malmsheimer R. W., O'Laughlin J., Oneil E. E., Sedjo R. A., and Skog K. E.. 2014. “Forest Carbon Accounting Considerations in U.S. Bioenergy Policy.” Journal of Forestry 112: 591–606. [Google Scholar]
- Morris, J. L. , Cottrell S., Fettig C. J., DeRose R. J., Mattor K. M., Carter V. A., Clear J., et al. 2018. “Bark Beetles as Agents of Change in Social–Ecological Systems.” Frontiers in Ecology and the Environment 16: S34–43. [Google Scholar]
- O'Halloran, T. L. , Law B. E., Goulden M. L., Wang Z., Barr J. G., Schaaf C., Brown M., et al. 2012. “Radiative Forcing of Natural Disturbances.” Global Change Biology 18: 555–65. [Google Scholar]
- Peng, D. , Wu C., Zhang B., Huete A., Zhang X., Sun R., and Lei L.. 2016. “The Influences of Drought and Land‐cover Conversion on Inter‐annual Variation of NPP in the Three‐North Shelterbelt Program zone of China based on MODIS data.” PLOS ONE 11: e0158173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peterson, G. , Allen C. R., and Holling C. S.. 1998. “Ecological Resilience: Biodiversity and Scale.” Ecosystems 1: 6–18. [Google Scholar]
- R Core Team . 2020. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/. [Google Scholar]
- Rebain, S. A. 2010. (revised June 2021). “The Fire and Fuels Extension to the Forest Vegetation Simulator: Updated Model Documentation.” Internal Report. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Forest Management Service Center.
- Roberts, D. W. , and Cooper S. V.. 1989. “Concepts and Techniques of Vegetation Mapping.” Land classifications based on vegetation: applications for resource management: 90–6. [Google Scholar]
- Robinson, N. P. , Allred B. W., Jones M. O., Moreno A., Kimball J. S., Naugle D. E., Erickson T. A., and Richardson A. D.. 2017. “A Dynamic Landsat Derived Normalized Difference Vegetation Index (NDVI) Product for the Conterminous United States.” Remote Sensing 9: 863. [Google Scholar]
- Robinson, N. P. , Allred B. W., Smith W. K., Jones M. O., Moreno A., Erickson T. A., Naugle D. E., and Running S. W.. 2018. “Terrestrial Primary Production for the Conterminous United States Derived from Landsat 30 m and MODIS 250 m.” Remote Sensing in Ecology and Conservation 4: 264–80. [Google Scholar]
- Roxburgh, S. H. , Berry S. L., Buckley T. N., Barnes B., and Roderick M. L.. 2005. “What is NPP? Inconsistent Accounting of Respiratory Fluxes in the Definition of Net Primary Production.” Functional Ecology 19: 378–82. [Google Scholar]
- Running, S. W. , Ramakrishna R. N., Heinsch F. A., Maosheng Z., Reeves M., and Hashimoto H.. 2004. “A Continuous Satellite‐Derived Measure of Global Terrestrial Primary Production.” Bioscience 54: 547–60. [Google Scholar]
- Schapira, Z. , Stevens‐Rumann C., Shorrock D., Hoffman C., and Chambers A.. 2021. “Beetlemania: Is the Bark Worse than the Bite? Effects of Bark Beetles and Wildfires in Rocky Mountain Subalpine Forests.” Forest Ecology and Management 482: 118879. [Google Scholar]
- Schmid, J. , and Frye R.. 1976. “Stand Ratings for Spruce Beetles.” USDA Forest Service, Rocky Mountain Research Station, Research Paper RM‐309.
- Seidl, R. , Spies T. A., Peterson D. L., Stephens S. L., and Hicke J. A.. 2016. “Searching for Resilience: Addressing the Impacts of Changing Disturbance Regimes on Forest Ecosystem Services.” Journal of Applied Ecology 53: 120–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens‐Rumann, C. S. , and Morgan P.. 2016. “Repeated Wildfires Alter Forest Recovery of Mixed‐Conifer Ecosystems.” Ecological Applications 26: 1842–53. [DOI] [PubMed] [Google Scholar]
- Temperli, C. , Hart S. J., Veblen T. T., Kulakowski D., Hicks J., and Andrus R.. 2014. “Are Density Reduction Treatments Effective at Managing for Resistance or Resilience to Spruce Beetle Disturbance in the Southern Rocky Mountains?” Forest Ecology and Management 334: 53–63. [Google Scholar]
- U. S. Forest Service . 2020. “Individual Tree Species Parameter Maps.” www.fs.fed.us/foresthealth/applied-sciences/mapping-reporting/.
- U. S. Forest Service . 2021. “Forest Insect and Disease Conditions in the Rocky Mountain Region, 2020.” Fort Collins, CO: USDA Forest Service, Rocky Mountain Region, Forest Health, R2–RO–21‐01.
- Windmuller‐Campione, M. A. , Page D. H., and Long J. N.. 2017. “Does the Practice of Silviculture Build Resilience to Spruce Beetle? A Case Study of Treated and Untreated Spruce‐Fir Stands in Northern Utah.” Journal of Forestry 115: 559–67. [Google Scholar]
- Yi, C. , and Jackson N.. 2021. “A Review of Measuring Ecosystem Resilience to Disturbance.” Environmental Research Letters 16: 053008. [Google Scholar]
- You, Y. , Wang S., Pan N., Ma Y., and Liu W.. 2020. “Growth Stage‐Dependent Responses of Carbon Fixation Process of Alpine Grasslands to Climate Change over the Tibetan Plateau, China.” Agricultural and Forest Meteorology 291: 108085. [Google Scholar]
- Young, I. T. 1977. “Proof Without Prejudice: Use of the Kolmogorov–Smirnov Test for the Analysis of Histograms from Flow Systems and Other Sources.” Journal of Histochemistry and Cytochemistry 25: 935–41. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix S1
Data Availability Statement
Data (Davis et al., 2022) are available in Dryad at https://doi.org/10.5061/dryad.jwstqjqb0.