

Abstract
Inflammaging refers to a chronic, systemic, low-grade inflammation, driven by immune (mainly macrophages) and non-immune cells stimulated by endogenous/self, misplaced or altered molecules, belonging to physiological aging. This age-related inflammatory status is characterized by increased inflammation and decreased macroautophagy/autophagy (a degradation process that removes unnecessary or dysfunctional cell components). Inflammaging predisposes to age-related diseases, including obesity, type-2 diabetes, cancer, cardiovascular and neurodegenerative disorders, as well as vulnerability to infectious diseases and vaccine failure, representing thus a major target for anti-aging strategies. Phenolic compounds—found in extra-virgin olive oil (EVOO)—are well known for their beneficial effect on longevity. Among them, hydroxytyrosol (HTyr) appears to greatly contribute to healthy aging by its documented potent antioxidant activity. In addition, HTyr can modulate inflammation and autophagy, thus possibly counteracting and reducing inflammaging. In this review, we reference the literature on pure HTyr as a modulatory agent of inflammation and autophagy, in order to highlight its possible interference with inflammaging. This HTyr-mediated activity might contribute to healthy aging and delay the development or progression of diseases related to aging.
Keywords: polyphenols, extra-virgin olive oil, olive vegetation waste, hydroxytyrosol, aging, inflammaging, inflammation, autophagy, cytokines, age-related diseases
1. Introduction
Aging is a complex physiological condition, derived by the time-dependent lowering of cell functions and dysregulation of molecular mechanisms underlying many cellular homeostatic processes [1].
A well-established hallmark of aging is the chronic state of immune activation causing low-grade inflammation in several tissues and organs, referred to as inflammaging [2,3]. The mechanisms underlying inflammaging are not yet completely understood. However, continuous immune stimulation associated with cellular stress and genetic factors appears to play a key role in generating this condition [4,5,6]. It has been proposed that a major source of immune stimulation in aging consists of endogenous/self, misplaced or altered molecules resulting from aged damaged/dead cells and organelles, and cell debris which, recognized by innate immune receptors, induces activation of inflammatory signaling pathways [4]. A key role in inflammaging is played by the activation of innate immune cells (mostly recruited monocyte-derived macrophages and tissue-resident macrophages, such as microglia), associated with upregulation of innate immune receptors and increased activity of pro-inflammatory signaling pathways [4,6]. Particular emphasis has been given to the balance of macrophage polarization into classically pro-inflammatory M1 and alternatively anti-inflammatory M2 activated macrophages, which is critical for immune homeostasis [4]. In aging, continuous immune stimulation is also associated with a decline in macroautophagy (herein referred to as autophagy)—a lysosome-mediated degradation process appointed to remove unnecessary or dysfunctional cell components [7,8,9]. Autophagy can regulate the function of immune and non-immune cells, affecting their inflammatory response [10,11]. Decreased autophagic activity can contribute to inflammaging in different ways, such as the direct activation of inflammatory signaling pathways as well as the impairment of cellular housekeeping, leading to the accumulation of damaged cellular and protein components, which represent inflammatory stimuli promoting inflammation [12]. Such a systemic imbalance between increased immune stimulation and reduced autophagy represents a major driving force for frailty, leading to aged dysfunctional immune and non-immune cells, declined immune functions and thus vulnerability to infectious diseases and vaccine failure.
The systemic pro-inflammatory status in inflammaging results in the rise of multiple soluble inflammatory mediators, secreted by immune and non-immune cells in the tissues and serum of the elderly. The inflammatory biomarkers of inflammaging include cytokines such as IL-1β, IL-18, IL-6, TNF-α, IL-17, IFN-γ, macrophage migration inhibitory factor (MIF), chemokines such as C-X-C motif ligand (CXCL) 8/IL-8, CXCL10/interferon-induced protein 10 (IP-10), CXCL13, C-C motif ligand (CCL)2/monocyte chemoattractant protein 1 (MCP-1) and CCL5/regulated on activation normal T cell expressed and secreted chemokine (RANTES), as well as other proteins such as C-reactive protein (CRP) [13,14,15,16,17]. Increased levels of these inflammatory mediators in the elderly represent a highly significant risk factor for most age-related diseases, including obesity, type 2 diabetes, osteoporosis, cancer, cardiovascular and neurodegenerative diseases [18,19,20,21,22]. For example, TNF-α, IL-1β and IL-6 cytokine levels are increased in the parkinsonian brain and serum [22,23,24,25], while the CCL2 chemokine level is raised in neuroinflammation associated to neurodegenerative diseases [26,27]. CXCL10 has been shown to be a biomarker for heart failure development and left ventricular dysfunction [28]. In addition, aging can also be associated with the dysregulated production of anti-inflammatory mediators, including cytokines such as IL-10 and IL-4 [21,29,30]. Therefore, inflammaging constitutes the basis for the onset or progression of chronic inflammatory diseases, which accelerate and propagate the aging process locally and systemically [31,32,33]. Consequently, inflammaging can be considered a major target for anti-aging strategies.
The Mediterranean diet (MD), based on fruit and vegetable intake, has been associated with a low occurrence of the diseases related to aging [34]. Daily consumption of extra-virgin olive oil (EVOO) is highly recommended for the beneficial role of polyphenols found in Olea europaea L. on human health [35,36,37,38]. The main phenolic compound is hydroxytyrosol (HTyr) (Figure 1). The central role of HTyr and its derivatives in the prevention of oxidative stress has been recognized by the European Food Safety Authority (EFSA), which recommends a daily intake of 20 g of EVOO containing at least 5 mg of these polyphenols [39].
Figure 1.
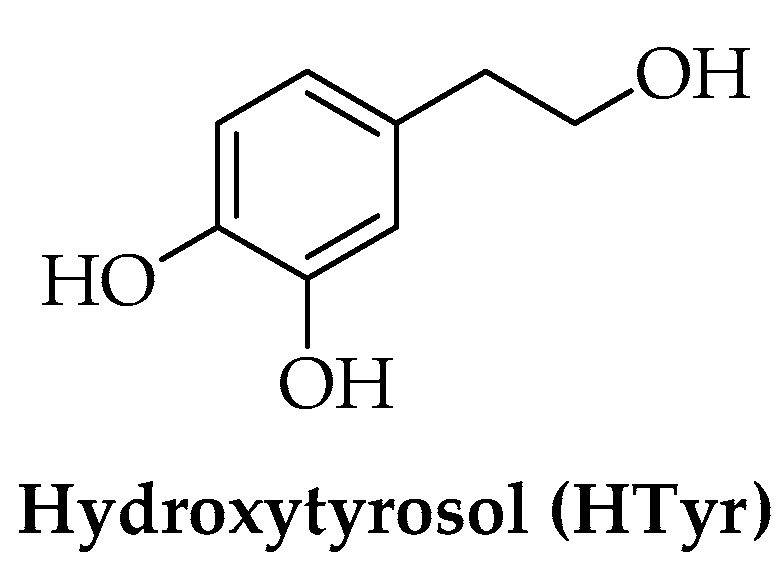
Chemical structure of hydroxytyrosol (HTyr).
HTyr is an amphiphilic compound; then, during olive oil processing, a fair amount spills into olive vegetation waste (OVW), such as olive oil wastewaters and olive pomace, which represents valuable sources from which to recover HTyr according to the “circular economy” model [40]. In recent times, HTyr has also been found in red wine, although with a lower concentration than in EVOO [41].
Our review is focalized on the potential effect of HTyr in inflammaging. In particular, we reference to literature investigating the role of purified HTyr as a modulatory agent of inflammation and autophagy in order to highlight its possible beneficial interference with inflammaging, an activity that might contribute to healthy aging and delay the development or progression of diseases related to aging.
2. Hydroxytyrosol (HTyr)
HTyr is the main phenol present in EVOO. It is a small, catecholic compound exhibiting antioxidant, cardioprotective, anticancer, neuroprotective and anti-inflammatory activities [42,43,44,45,46,47]. Because of these interesting properties, it has a high therapeutic potential and is used as a supplement and preservative for cosmeceutical, nutraceutical and food applications.
Despite the biological relevance of HTyr, its biosynthesis is still not fully clear. According to Scheme 1, the proposed pathway in Rhodiola crenulate starts from L-DOPA, which is decarboxylated to afford dopamine; dopamine is firstly deaminated to 3,4-dihydroxyphenylacetaldehyde and finally reduced to HTyr [48].
Scheme 1.

Proposed biosynthetic pathway of HTyr [48].
As olives ripen and during their processing, the amount of HTyr increases due to the hydrolysis of oleuropein, which produces glucose, elenolic acid and HTyr, according to Scheme 2 [49]. The same reaction occurs after intake of oleuropein under gastric conditions [50] and by the action of the microbiota [51].
Scheme 2.

Being a hydrophilic compound, during the production of EVOO, HTyr splits between the oil and liquid wastes such as olive oil wastewater or in solid wastes such as olive pomace, depending on the partition coefficients. These wastes represent a valuable source of HTyr, from which it can be recovered according to the concept of the “circular economy” [40]. Membrane technologies are sustainable and efficient to produce HTyr-enriched extracts [52,53,54] and the content of HTyr can be increased, treating them with tyrosinase to convert tyrosol to HTyr [55]. However, these extracts contain other phenolic compounds, and the observed biological activities can be influenced by them. In recent times, HTyr has been identified in red wine [41] because of the action of yeasts, which convert tyrosine to HTyr during alcoholic fermentation [56].
Many in vitro and in vivo studies are focused on the ADMET (absorption, distribution, metabolism, excretion and toxicity) processes of HTyr responsible for its beneficial role on health [43,44]. The absorption occurs in the small intestine, but the efficiency of the process depends on the food matrix, age and gender. Experimental results have demonstrated that absorption is higher if HTyr is administered as EVOO [57] and in female rats [58]. If administered as red wine, the levels of HTyr increase in urine due to its production by ethanol and dopamine through dopaminergic pathways [59]. After the absorption, HTyr reaches its highest plasma concentration in 5–30 min, and it is distributed among the kidney, liver, muscle, and brain. These peculiar properties, along with the strong antioxidant effect, candidate HTyr as a dopaminergic neuronal protector. The metabolism of HTyr is mainly related to the production of HTyr acetate (HTyr-Ac), homovanillyl alcohol and homovanillic acid, which were converted into the corresponding sulfate and glucuronide by conjugation reactions catalyzed by uridine 5-diphosphoglucuronosyl transferases and sulfotransferases. Some HTyr metabolites are depicted in Figure 2 [43]. Additionally, the excretion of HTyr depends on the food matrix [60] and a gender-dependent relationship was observed [58]. The main urinary HTyr metabolites are glucuronides.
Figure 2.

Chemical structures of HTyr metabolites [43].
To the best of our knowledge, the toxicity of pure HTyr has not yet been studied. However, the safety profile has been studied for Hidrox®, an HTyr-enriched extract obtained by a patented process. In vivo studies have demonstrated a No Observed Adverse Effect Level (NOAEL) of 500 mg/kg/day of HTyr [61]. At this concentration, HTyr has no adverse effects related to genotoxicity and mutagenicity [62]. This formulation has been designated as Generally Recognized as Safe (GRAS) and has been evaluated on oxidative stress, neuroinflammation, apoptosis and inflammasomes [63].
Purified HTyr can be obtained after chromatographic separations of EVOO or HTyr-enriched extracts or after chemical or enzymatic synthesis. Several synthetic procedures have been optimized in recent years using tyrosol and homovanillic alcohol as starting materials and efficient oxidative stoichiometric or enzymatic oxidants such as 2-iodoxybenzoic acid or tyrosinase [64,65,66].
Synthesis also offers the solution to improve the bioavailability of HTyr. In fact, several lipophilic derivatives have been prepared [67,68,69,70]. The most interesting one is HTyr-Ac depicted in Figure 2 [71]. Dominguez-Perles et al. carried out an in vivo experiment demonstrating the higher absorption of HTyr-Ac compared to HTyr through the corresponding metabolites obtained after oral administration [58]. Recently, the more lipophilic compound HTyr triacetate, called by the authors peracetylated HTyr (Per-HTyr) (Figure 3), was tested as an anti-inflammatory agent [72].
Figure 3.
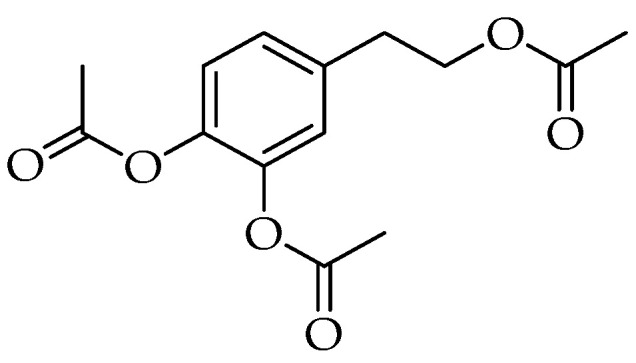
Chemical structure of HTyr triacetate (peracetyted HTyr, Per-HTyr) [72].
3. Inflammation, Inflammaging and HTyr
3.1. Inflammation and Inflammaging
Inflammation represents a crucial line of defense against pathogens. It is an evolutionarily conserved protective process designed to maintain organismal homeostasis against acute perturbations and serves as an adaptive response to infections and injuries [73]. However, chronic, systemic inflammation develops progressively with age in healthy individuals and contributes to organismal deterioration through a process termed inflammaging [5,74,75].
The major inflammatory cells involved in inflammaging are macrophages, including recruited monocyte-derived macrophages and tissue-resident macrophages such as microglia in the brain and spinal cord [4,76,77]. However, recent reports indicate also the involvement of other immune cells, such as T lymphocytes, in eliciting aging-related tissue inflammation [78]. For example, myocardial inflammaging is related to increased accumulation of dysfunctional CD4+T cells secreting massive amounts of inflammatory interferon (IFN)-γ [79,80]. Therefore, we can assume that, in inflammaging, both innate and adaptive immune cells are characterized by enhanced expression and secretion of pro-inflammatory mediators, often associated with reduced production of anti-inflammatory molecules.
The raise of pro-inflammatory mediators in inflammaging is the result of the increased activation of intracellular signaling molecules, such as those belonging to the mitogen-activated protein kinase (MAPK) pathway [81,82], as well as transcription factors, with a pivotal role played by the nuclear factor kappa-light-chain enhancer of activated B cells (NF-κB) [83,84,85]. The MAPK signaling pathway is involved in transmitting extracellular signals to the nucleus, playing critical regulatory roles in the production of pro-inflammatory cytokines. The MAPK family consists of the following three subfamilies: the extracellular signal-regulated kinases (ERKs; ERK1/2 and ERK5), the c-Jun N-terminal kinases (JNKs; JNK1-3) and the p38MAPK, which activates transcription factors leading to inflammation [81]. Therefore, MAPKs are important targets for the treatment of chronic inflammatory diseases [82]. Then, the transcription factor NF-κB is the master regulator of the expression of pro-inflammatory genes (including those encoding cytokines and chemokines), and NF-κB upregulation has been well documented in age-related diseases [86,87]. NF-κB activation is induced by cell stressor signals such as antigens, toll-like receptor (TLR) ligands and cytokines (e.g., tumor necrosis factor-TNF), which, via intermediate steps, lead to the engagement and activation of the inhibitor of the nuclear factor-κB (IκB) kinase (IKK) complex, which guides the phosphorylation of IκB, its ubiquitination and degradation. Once degraded, the remaining NF-κB dimer, composed of p65/p50 subunits, translocates to the nucleus and activates the transcription of inflammatory genes [81,83]. The MAPK pathway and NF-κB activations, tightly regulated by phosphorylation and ubiquitination, are responsible for the production of inflammatory cytokines such as TNF-α and IL-6. In addition, NF-κB translocation to the nucleus is responsible for the expression of IL-1β and IL-18 pro-forms that are processed and released in their active forms by another critical component of the inflammatory response, such as the nucleotide binding domain-like receptor pyrin domain containing the protein 3 (NLRP3) inflammasome [88]. NLRP3 is the most extensively studied inflammatory activator among the inflammasomes. Inflammasomes are a family of intracellular multi-protein complexes, belonging to the family of cytoplasmic innate immune receptors, which activate inflammatory caspases [88]. The NLRP3 inflammasome is mainly expressed by inflammatory innate immune cells such as monocytes/macrophages, dendritic cells and neutrophils. However, it can be also expressed by T lymphocytes and plasma cells, as well as by non-immune cells, including endothelial cells, fibroblasts, muscle cells, adipose progenitor cells and adipocytes. NLRP3 inflammasome formation, whose signaling events and way of assembly are not fully understood, is triggered by pathogen- or damage- associated molecular patterns, which are generated by endogenous stress, tissue damage or metabolic imbalances. Thus, once the NLRP3 protein complex has formed, it activates caspase-1, which initiates the processing and release of active IL-1β and IL-18 pro-inflammatory cytokines. Increased NLRP3 activity associated with increased IL-1β and IL-18 levels have been demonstrated in the elderly, and contribute to inflammaging [75,88,89,90]. Indeed, increased NLRP3 activation is associated with several age-related pathological conditions, including Alzheimer’s disease, atherosclerosis and type-2 diabetes [91,92], whereas NLRP3 inhibition extends health-spans and diminishes age-dependent degenerative conditions [90].
Another biological system implicated in inflammation and inflammaging is the microbiota, which is the community of bacteria, viruses, fungi and protozoa living in the human body and interacting with it. We refer not only to the gut microbiota but also the microbiota at different body sites, whose composition and diversity change according to age [93,94]. Age-related microbiota changes (referred to as age-related dysbiosis) modulate the immune response and produce inflammatory molecules, contributing to immunosenescence and inflammaging by long-term stimulation of inflammation. On the other hand, by-products of metabolic processes in microbiota, including some short-chain fatty acids, can play a role in inhibiting inflammation [93,94].
Altogether, inflammation in inflammaging can be the result of the imbalance between pro- and anti-inflammatory molecules and underlies several age-related diseases.
3.2. HTyr as Modulatory Agent of Inflammation
Polyphenols have been widely recognized as anti-inflammatory agents [95,96,97,98,99,100]. In addition, it has been shown by our and other’s laboratories that HTyr-enriched preparations, including those derived from EVOO and OVW, as well as multicomponent nutraceutical containing HTyr, can modulate several key processes related to inflammation [55,63,101,102,103,104,105,106,107,108,109]. However, these preparations, containing other phenolic compounds as well as HTyr, do not allow the identification of immunomodulatory activity mediated by the single phenol and do not exclude the potential synergistic effects for the presence of other compounds. Therefore, here we reference studies that investigate the in vitro (Table 1) and in vivo (Table 2) modulation of inflammation by purified HTyr. Moreover, we focus our attention to data concerning the HTyr capability of directly affecting the mentioned inflammatory biomarkers of inflammaging, referring to other reviews for its antioxidant activity [63,96,103,110,111,112,113,114,115,116].
3.2.1. Evidence In Vitro
Considering the central role of macrophages in inflammation, we first reference research investigating the in vitro immunomodulatory activity of HTyr on experimental models of murine and human inflammatory monocytes/macrophages (Table 1). Treatment of the human monocyte/macrophage cell line THP-1 with HTyr (25, 50, and 100 μM) for 10 min, before stimulation with lipopolysaccharide (LPS) for 3 h, resulted in a dose-dependent reduction of TNF-α transcription [117]. Then, 30 min of HTyr (0.25, 0.5, 1 μM) pre-treatment of oxysterol mixture-stimulated human peripheral blood mononuclear cells (PBMCs) from healthy donors reduced the secretion of pro-inflammatory cytokines and chemokines, such as IL-1β, MIF, RANTES as well as MAPK signaling, analyzed as p38MAPK and JNK phosphorylation [118]. In addition, HTyr (10, 20, 40, 80 μM) exerted anti-inflammatory effects on LPS-stimulated RAW264.7 murine macrophages, in that it downregulated LPS-induced expression of IL-1β and TNF-α, as well as suppressed NF-κB phosphorylation and activation [119]. Another study illustrated the high anti-inflammatory activity of Per-HTyr observed at lower doses than that used for HTyr. In fact, HTyr 50 (μM) and Per-HTyr (12.5, 25, 50 μM) significantly reduced the production of pro-inflammatory cytokines such as IL-1β, TNF-α, IL-6, IFN-γ and IL-17 by ex vivo murine peritoneal macrophages stimulated with LPS for 18 h. Both polyphenols also inhibited the activation of the signal transducer and activator of transcription 3 (STAT3), an important transcription factor involved in the regulation of immune response, inflammation and ageing [120]. Moreover, Per-HTyr suppressed the activation of the non-canonical NLRP3 inflammasome (which has the same effect as the canonical NLRP3, but is mediated by caspase-11 instead of caspase-1), decreasing thus the IL-18 pro-inflammatory cytokine level [72]. In another study, 12-h HTyr (50, 100 μM) pre-treatment of 24-h LPS-induced RAW 264.7 cells decreased pro-inflammatory M1/CD11c+ macrophages and increased anti-inflammatory M2/CD206+ macrophages, while reducing mRNA and protein levels of TNF-α, IL-1β, IL-6 pro-inflammatory cytokines and raising IL-10 and IL-4 anti-inflammatory cytokines. These results were also associated with a decrease in ERK1/2 phosphorylation, indicating that HTyr is able to promote macrophage M2 polarization with the increase of anti-inflammatory cytokines through the inhibition of the MAPK signaling pathway [121]. However, in contrast to the anti-inflammatory effects of HTyr reported by different investigators so far, in a small number of studies, HTyr appeared to exert no effect or even promote inflammation. In fact, Bigagli et al. reported that co-treatment of LPS-stimulated RAW 264.7 with low doses (5 and 10 μM) of HTyr for 18 h produced no change in IL-1β and TNF-α gene expression [122]. Then, co-incubation of LPS-activated human monocytes with HTyr (50, 100 μM) for 24 h increased TNF-α production by inflammatory monocytes [123]. Accordingly, the expression of TNF-α was significantly increased in ex vivo murine peritoneal macrophages co-incubated with LPS and HTyr (80 μM) for 24 or 48 h. In this study, HTyr also increased LPS-dependent expression of the IL-10 gene, whereas it did not affect LPS-dependent NF-κB gene expression and NF-κB phosphorylation [124].
Moreover, we reference studies on the immunomodulatory activity of HTyr on experimental models of inflammatory microglia cells (Table 1), specialized macrophages of the central nervous system and principal orchestrators of neuroinflammation. Gallardo-Fernandez et al. showed that HTyr (1, 10, 25 and 50 μM) was able to reduce mRNA levels of pro-inflammatory mediators such as TNF-α, IL-1β, IL-6 and CXCL10 expressed by LPS-induced BV2 murine microglial cells. The analysis of the possible molecular mechanisms involved in the anti-inflammatory effect showed that HTyr decreased the phosphorylated forms of JNK1/2 and p38MAPK, thus reducing MAPK signaling, as well as inhibiting the activation of the NLRP3 inflammasome. In addition, HTyr treatment prevented LPS-induced translocation to the nucleus of NF-κBp65 [125]. Accordingly, another study revealed that HTyr (25, 50, 100 μM) significantly decreased the production of IL-1 β, TNF-α and IL-6 by BV2 microglia and primary microglia cells stimulated with LPS. Moreover, HTyr significantly reduced M1/CD86+ macrophages and increased M2/CD206+ macrophages, decreased phosphorylated ERK and NF-κBp65 levels in a dose-dependent manner and suppressed the expression of toll like receptor 4 (TLR4) in BV2 microglia. These results suggest that HTyr can suppress LPS-induced neuroinflammatory responses via modulation of M1/M2 microglia polarization and downregulation of TLR-4 mediated NF-κB activation and the ERK signaling pathway [126]. Then, a study investigated the ability of HTyr (20, 100 μM) to alleviate neuropathic pain, by using an experimental model of intervertebral disc degeneration (IVDD). TNF-α-stimulated primary human nucleus pulposus cells (HNPCs) were used to simulate the local inflammatory microenvironment observed in IVDD, and LPS was used to stimulate rat microglia cells. HTyr inhibited the secretion of TNF-α, IL-6, IL-1β and the activation of the NLRP3 inflammasome, while it also suppressed NF-κB activation and ERK phosphorylation. These results suggest that HTyr plays a protective role against IVDD and secondary neuropathic pain by inhibiting NF-κB and MAPK inflammatory pathways [127].
Finally, the anti-inflammatory activity of HTyr has also been investigated in experimental models of inflammatory non-immune cells (Table 1), such as senescent fibroblasts [128], keratinocytes [129,130] and colonic epithelial cells [131]. Menicacci et al. investigated the ability of HTyr to affect the inflammatory activity of senescent cells, which display increased secretion of growth factors, inflammatory cytokines and proteolytic enzymes. Chronic (4–6 weeks) pre-treatment with HTyr (1 μM) of pre-senescent MRC5 human lung fibroblasts, neonatal human dermal fibroblasts (NHDF) and TNF-α-stimulated NHDF reduced IL-6 production in all cellular experimental models. In addition, HTyr decreased NFκB protein levels and nuclear localization. These data suggest that the modulation of the senescence-associated inflammatory phenotype might be another important activity underlying the anti-inflammatory activity of HTyr in inflammaging [128]. Then, primary human keratinocytes were pre-treated with HTyr (12.5–100 μM) or HTyr-Ac (12.5–100 μM) for 30 min and then stimulated with the IL-1β or TLR3 ligand (Poly I:C). The expression of TNF-α, IL-6 and IL-8 inflammation-related genes was likewise inhibited by HTyr-Ac and HTyr. Mechanistically, these polyphenols counteracted IκB degradation and translocation of NF-κB to the nucleus, in particular to the critical binding site in the IL-8 promoter [129]. Another study showed that HTyr exerted a potent anti-inflammatory effect on inflammatory keratinocytes in an experimental model of psoriasis, a prevalent chronic inflammatory dermatosis. Indeed, HTyr (25, 50, 100 μM) decreased the expression of IL-6, IL-8 and TNF-α in HaCaT keratinocytes stimulated with a M5 cytokine cocktail (composed of TNF-α, oncostatin-M, IL-17A, IL-1α and IL-22) [130]. Lastly, a recent study demonstrated the anti-inflammatory effect of HTyr on a human model of colonic chemical carcinogenesis. Primary human colonic epithelial cells (HCoEpC) were pre-treated with 1 μM HTyr 45 min before their exposition to benzo[a]pyrene [B[a]P]. Interestingly, all the inflammatory effects induced by B[a]P could be counteracted by HTyr. In fact, HTyr reduced the release of pro-inflammatory cytokines, chemokines and growth factors, including IL-6, IL-8, CXCL13 and vascular endothelial growth factor (VEGF), and molecules able to recruit inflammatory monocytes and activate macrophages, which could contribute to inflammation, angiogenesis and tumorigenesis. Moreover, HTyr efficiently counteracted B[a]P-mediated ERK1/2 phosphorylation. These data suggest that HTyr can play an important role in preventing tumorigenesis by chemical carcinogens [131].
Table 1.
In vitro effects of HTyr on inflammation.
| Treatment and Dose | Model | Effects | Ref. |
|---|---|---|---|
| Pre-treatment 25, 50, 100 μM HTyr |
Human Macrophages: THP-1 + LPS |
↓ TNF-α | [117] |
| Pre-treatment 0.25, 0.5, 1 μM HTyr |
Human Mononuclear Cells: PBMCs + oxysterol mixture |
↓ IL-1β, MIF, RANTES ↓ p38MAPK and JNK phosphorylation |
[118] |
| Co-treatment 10, 20, 40, 80 μM HTyr |
Murine Macrophages: RAW 264.7 + LPS |
↓ IL-1β, TNF-α ↓ NF-κB phosphorylation |
[119] |
| Pre-treatment 50 μM HTyr 12.5, 25, 50 μM Per-HTyr |
Murine Macrophages: ex vivo peritoneal macrophages + LPS |
↓ IL-1β, IL-6, TNF-α, IL-17, IFN-γ ↓ STAT3 activation ↓ IL-18 via non-canonical NLRP3 inflammasome |
[72] |
| Pre-treatment 50, 100 μM HTyr |
Murine Macrophages: RAW 264.7 + LPS |
↓ M1, ↑ M2 macrophages ↓ IL-1β, IL-6, TNF-α ↑ IL-10, IL-4 ↓ ERK1/2 phosphorylation |
[121] |
| Co-treatment 5, 10 μM HTyr |
Human Peripheral Blood Monocytes: ex vivo monocytes + LPS |
No change in IL-1β, TNF-α | [122] |
| Co-treatment 50, 100 μM HTyr |
Murine Macrophages: RAW 264.7 + LPS |
↑ TNF-α | [123] |
| Co-treatment 80 μM HTyr |
Murine Macrophages: ex vivo peritoneal macrophages + LPS |
↑ TNF-α, IL-10 no change in NF-κB expression and phosphorylation |
[124] |
| Co-treatment 1, 10, 25, 50 μM HTyr |
Murine Microglia: BV2 + LPS |
↓ IL-1β, TNF-α, IL-6, CXCL10 ↓ JNK1/2 and p38MAPK phosphorylation ↓ NF-κBp65 translocation to the nucleus ↓ NLRP3 inflammasome |
[125] |
| Pre-treatment 25, 50, 100 μM HTyr |
Murine Microglia: BV2 + LPS primary microglia + LPS |
↓ M1 ↑ M2 ↓ IL-1 β, TNF-α, IL-6 ↓ TLR-4 ↓ NF-κBp65 phosphorylation ↓ ERK1/2 phosphorylation |
[126] |
| Co-treatment 20, 100 μM HTyr |
Human Nucleus Pulposus Cells and Rat Microglia: primary HNPC + TNF-α microglia + LPS |
↓ IL-1β, TNF-α, IL-6 ↓ NLRP3 inflammasome ↓ NF-κB activation ↓ ERK phosphorylation |
[127] |
| Pre-treatment 1 μM HTyr for 4 weeks |
Human Pre-senescent and Senescent Fibroblasts: MRC5 NHDF NHDF + TNF-α |
↓ IL-6 ↓ NF-κB activation |
[128] |
| Pre-treatment 12.5–100 μM HTyr 12.5–100 μM HTyr-Ac |
Human Keratinocytes: primary keratinocytes + IL-1β primary keratinocytes + Poly I:C |
↓ TNF-α, IL-6, IL-8 ↓ NF-κB activation and translocation to binding site in the IL-8 promoter |
[129] |
| Pre-treatment 25, 50, 100 μM HTyr |
Human Psoriatic Keratinocytes: HaCaT + M5 cytokine cocktail |
↓ IL-6, IL-8, TNF-α | [130] |
| Pre-treatment 1 μM HTyr |
Chemical Carcinogenesis in Human Primary Colonic Epithelial Cells: HCoEpC + B[a]P |
↓ IL-6, IL-8, VEGF, CXCL13 ↓ ERK1/2 phosphorylation |
[131] |
Abbreviations: Per-HTyr: peracetylated hydroxytyrosol. HTyr-Ac: hydroxytyrosyl acetate. LPS: lipopolysaccharide. PBMC: peripheral blood mononuclear cells. TNF: tumor necrosis factor. IL: interleukin. MIF: macrophage migration inhibitory factor. RANTES: regulated on activation normal T-cell expressed and secreted. CXCL: C-X-C motif ligand. VEGF: vascular endothelial growth factor. TLR: toll-like receptor. NF-κB: nuclear factor-κB. MAPK: mitogen-activated protein kinase. STAT: signal transducer and activator of transcription. ERK: extracellular signal-regulated kinase. JNK: c-Jun N-terminal kinase. HNPC: human nucleus pulposus cells. MRC5: human fibroblasts from fetal lung tissue. NHDFs: neonatal human dermal fibroblasts. B[a]P: Benzo[a]pyrene. ↓ Decrease. ↑ Increase.
3.2.2. Evidence In Vivo
The modulation of inflammation by HTyr has also been investigated in vivo, in inflammatory experimental animal models. As for in vitro investigations, we start by reporting studies on the immunomodulatory effects of HTyr on innate immune cells, such as macrophages and microglia (Table 2). In an in vivo murine model of pristane-induced systemic lupus erythematosus, oral administration of 100 mg/kg (4 g/day for 6 month) HTyr was able to reduce IL-1β and IL-6 secreted by ex vivo LPS-stimulated splenocytes and macrophages. Studies on renal tissue, in the same experimental model, showed that HTyr also prevented the degradation of IκB, the nuclear translocation of NF-κBp65 and the phosphorylation of MAPK [132]. Then, in C57BL/6 mice with an LPS-induced acute liver injury, where infiltrated inflammatory macrophages play a critical role in liver destruction and dysfunction, HTyr (100 mg/kg orally, once a day daily, for 2 days) decreased inflammatory M1/CD11c+ macrophages and increased anti-inflammatory M2/CD206+ macrophages in the liver tissue, while it reduced liver TNF-α, IL-1β and IL-6 mRNA and augmented serum IL-10 and IL-4 protein levels. These data suggest that HTyr mitigated hepatic inflammation and injury through modulating macrophage-mediated inflammation [121]. In a model of atherosclerosis in apoE−/− mice, HTyr administration (10 mg/kg/day orally, for 16 weeks) significantly reduced the extent of aorta atherosclerotic lesions as well as serum CRP, TNF-α, IL-1β and IL-6 levels, while it increased IL-10. In addition, HTyr reduced the activation of inflammatory signaling molecules, such as p38MAPK phosphorylation and NFκ-B activation in the liver, suggesting that HTyr displayed anti-atherosclerotic action via modulating inflammation through the inhibition of inflammatory signaling molecules [133]. Accordingly, Pirozzi et al. showed that HTyr (10 mg/kg/day orally, for 5 weeks) exerted anti-inflammatory activity in the liver of rats with nonalcoholic fatty liver disease (NAFLD), by inhibiting liver TNF-α and IL-6 mRNA expressions [134]. Another study showed a potent anti-inflammatory activity of HTyr in a mouse model of systemic inflammation. In Balb/c mice pre-treated with HTyr (80 mg/kg/daily, for 2 or 5 days) and stimulated by intra-peritoneal injection of LPS, HTyr prevented the LPS-induced increase of TNF-α plasma levels [135].
Moreover, studies have also shown the capability of HTyr to inhibit neuroinflammation in vivo (Table 2). In this context, it is important to consider that HTyr is capable of crossing the blood brain barrier [136]. In mice pre-treated with HTyr (100 mg/kg by gavage, daily for 2 days) and administered with LPS, HTyr significantly damped the LPS-induced increase of IL-6, IL-1β and TNF-α mRNA levels in the brain [126]. Moreover, Yu et al. reported that intrathecal injection of HTyr (2 μL of HT 100 μM) alleviates neuropathic pain in rats undergoing chronic compression of the dorsal root ganglion. Notably, HTyr reduced the production of IL-1β, IL-6 and TNF-α inflammatory cytokines in the spinal dorsal horn. In addition, HTyr inhibited the activation of the ERK signaling pathway by reducing the levels of ERK phosphorylation in the spinal dorsal horn [127].
The anti-inflammatory activity of HTyr has also been investigated in inflammatory models of murine dextran sulfate sodium (DSS)-induced colitis [137,138]. A low dose (10 mg/kg) and high dose (50 mg/kg) of HTyr intervention significantly reduces the markers of DDS-induced colitis such as IL-6, IL-1β and TNF-α inflammatory cytokines and the NF-κB inflammatory pathway. Moreover, a high dose of HTyr increased the secretion of the IL-10 anti-inflammatory cytokine. It is also noteworthy that HTyr intervention transformed the gut microbiota, leading to a lower abundance of inflammation-related microbes (e.g., Bacteroidaceae and Desulfovibrionaceae) and a higher level of short-chain fatty acids producing bacteria (e.g., Lachnospiraceae, Muribaculaceae, ASF356 and Colidextribacter), indicating that the modulation of inflammation by HTyr is also mediated by its capability to modulate the microbiota [137]. In another study, it was found that HTyr (40 mg/kg/day orally for 14 days) exerted anti-inflammatory activity in murine DSS-induced ulcerative colitis by inhibiting NLRP3 inflammasome activation, through the suppression of the expression levels of NLRP3, caspase-1 and the apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC) mRNA, thus decreasing IL-18 and IL-1β levels. Moreover, this study confirmed the capability of HTyr to modulate gut microbiota by exerting a shift from pathogenic bacteria (Helicobacter, Staphylococcus, Desulfovibrio and Streptococcus) to probiotics (Lactobacillus, Lachnospiraceae NK4A136 group, [Ruminococcus] torques group and Roseburia), and increasing the levels of short-chain fatty acids (total acid, acetate, propionate and butyrate) [138].
Table 2.
In vivo effects of HTyr on inflammation.
| HTyr Treatment and Dose | Model | Effects | Ref. |
|---|---|---|---|
| 100 mg/kg diet: 4 g/day orally for 6 months |
Murine pristane-induced Systemic Lupus Erythematosus: ex vivo splenocytes+LPS ex vivo macrophages+LPS renal tissue |
↓ IL-1β, IL-6 ↓ IκB degradation, p65NF-kB nuclear translocation ↓ MAPK phosphorylation |
[132] |
| 100 mg/kg/day orally for 2 days | Murine LPS-induced Acute Liver Injury: liver tissue liver macrophages serum |
↓ M1 ↑ M2 macrophages ↓ IL-1 β, TNF-α, IL-6 ↓ IL-10, IL-4 |
[121] |
| 10 mg/kg/day orally for 16 weeks | ApoE−/− Mice Atherosclerosis: blood heart tissue liver tissue |
↓ IL-1β, TNF-α, IL-6, CRP ↑ IL-10 ↓ NF-kB activation ↓ p38MAPK phosphorylation |
[133] |
| 10 mg/kg/day orally for 5 weeks | Rats with Nonalcoholic Fatty Liver Disease: liver tissue |
↓ TNF-α, IL-6 | [134] |
| 80 mg/kg/daily for 2 or 5 days |
Murine LPS-induced Systemic Inflammation: plasma |
↓ TNF-α | [135] |
| 100 mg/kg/day orally for 2 days | Murine LPS-induced Brain Inflammation: brain tissue |
↓ IL-6, IL-1 β, and TNF-α | [126] |
| 2 μL of 100 μM injected intrathecally | Rat Chronic Compression of Dorsal Root Ganglion-induced Neuropathic Pain: spinal dorsal horn |
↓ IL-1β, TNF-α, IL-6 ↓ ERK phosphorylation |
[127] |
| 10 and 50 mg/kg/day orally |
Murine DSS-induced Colitis: colon tissue fecal samples |
↓ IL-6, IL-1β, and TNF-α ↑ IL-10 ↓ NF-κB activation ↓ Inflammation-related microbes of gut microbiota |
[137] |
| 40 mg/kg/day orally for 14 days | Murine DSS-induced Colitis: colon tissue fecal samples |
↓ IL-18 and IL-1β via ↓ NLRP3 inflammasome activation ↓ inflammation-related microbes of gut microbiota |
[138] |
Abbreviations: LPS: lipopolysaccharide. IL: interleukin. TNF: tumor necrosis factor. DSS: dextran sodium sulfate. NF-κB: nuclear factor-κB. MAPK: mitogen-activated protein kinase. ERK: extracellular signal-regulated kinase. ↓ Decrease ↑ Increase.
4. Autophagy, Inflammaging and HTyr
4.1. Autophagy and Inflammaging
Autophagy is an evolutionary conserved, highly regulated cellular process, responsible for the removal via lysosomal degradation of damaged cell material, such as misfolded proteins and unnecessary or dysfunctional organelles [139]. It can also regulate cellular energy balance by triggering energy production from its own components during nutrient starvation [140]. The autophagic process has a housekeeping role under physiological conditions and an adaptive, cytoprotective role under stress, controlling thus cellular and tissue homeostasis [141,142]. The autophagic process, its response to stress and its regulation has been described in detail elsewhere [143]. Briefly, autophagy is a dynamic process that is controlled by various signaling molecules, including the two main regulators such as the mammalian/mechanistic target of rapamycin (mTOR) and the 5′ adenosine monophosphate-activated protein kinase (AMPK), which act as autophagic inhibitor and activator, respectively [144,145]. mTOR is a serine–threonine kinase that senses cellular nutrient levels through its connection with different signaling pathways, including the MAPK/ERK1/2 pathway and the phosphatidylinositol 3-kinase (PI3K)/Ak strain transforming (Akt; also known as protein kinase B) pathway, which establish the PI3K/Akt/mTOR axis. Regarding the latter, PI3K, activated by many types of cellular stimuli, phosphorylates the phosphatidylinositol 4,5-bisphosphate (PIP2) to phosphatidylinositol (3,4,5)-trisphosphate (PIP3) that recruits and activates Akt via phosphorylation. Phosphorylated Akt, as a serine/threonine kinase, is able to stimulate mTORC1, which can directly activate a variety of cellular effectors, including the eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1), to inhibit autophagy [81,144]. The autophagic driver AMPK is a serine/threonine protein kinase and a highly conserved energy sensor, which can be activated by allosteric regulation or by upstream kinases. Activated AMPK phosphorylates integrated signaling networks to promote autophagy directly (by the activation of transcription factors) or indirectly (by the inhibition of the mTORC1 pathway) [146]. Other signaling molecules involved in autophagic regulation are sirtuins (SIRT), a family (SIRT1-7) of nicotinamide adenine dinucleotide (NAD+)-dependent enzymes, which play roles in diverse cellular processes including autophagy, senescence and cell survival [147]. Among them, SIRT1 and SIRT6, NAD+-dependent deacetylases belonging to class III histone/protein deacetylases and members of the silent information regulator 2 (Sir2) family, are predominantly localized in the nucleus and share functional similarities [148,149]. They play essential roles in promoting autophagy. SIRT1 can promote autophagy both directly, by interacting with autophagy related molecules (ATGs), and indirectly, by interacting with upstream regulators of autophagy, such as those activating AMPK or inhibiting the Akt/mTOR pathway [150]. SIRT6 can induce autophagy via inhibition of Akt signaling, thus inhibiting the Akt-mTOR pathway [151].
The most common markers used for monitoring autophagy are two proteins, the microtubule-associated protein 1 light chain 3 (LC3) and the sequestosome-1 (SQSTM1)/p62 [152]. LC3 is recruited from the cytosol and associates with the phagophore early in autophagy; this localization serves as a general marker for autophagic membranes and for monitoring the process in its development. The SQSTM1/p62 protein is an ubiquitin-binding scaffold protein, which co-localizes with cargo destined to be degraded by autophagy; itself is degraded in autolysosomes and serves as an index of autophagic degradation.
It is now well known that autophagy contributes to the extension of longevity and prevents age-related pathologies [142,153]. Among the beneficial effects on age-related processes, autophagy orchestrates the differentiation and metabolic state of innate and adaptive immune cells, inducing immune response improving effects [154,155,156]. In particular, autophagy plays a pivotal role in both the prevention of aging in immune and non-immune cells and the suppression of inflammaging [10,11,142,153]. Indeed, autophagy prevents inflammation by downregulating inflammation-related signals, reducing inflammatory cytokine expression and promoting apoptotic corpse clearance [10,11]. Autophagy can suppress inflammatory reactions not only directly, but also indirectly through its cytoprotective activity, by increasing cytoplasmic turnover that antagonizes the degradation of organelles and macromolecular complexes, thus ameliorating immune system homeostasis. Although the regulatory mechanisms for autophagy in inflammation are complex and remain to be fully elucidated, the balance between mTOR and AMPK activation has a central role in innate immune cell homeostasis and function [155,157]. It has been demonstrated that downregulation of autophagy in macrophages leads to inflammation. Autophagy can regulate the metabolic state of macrophages, in that mTOR activation, reducing the autophagic flux, induces a proliferative and pro-inflammatory M1 phenotype in macrophages, whereas AMPK activation, driving autophagy, promotes the function of anti-inflammatory M2 macrophages [141]. In addition, AMPK can inhibit the inflammatory response by inhibiting the pro-inflammatory NF-κB pathway in immune and non-immune cells [158]. Furthermore, enabling the removal of damaged cell material, autophagy also regulates the intracellular danger signal-sensing multiprotein platform such as the NLRP3 inflammasome, inhibiting thus the activation of IL-1β and IL-18 inflammatory cytokines [159,160]. Finally, autophagy regulates also the adaptive immune response, such as T lymphocyte function, including their metabolism, survival, development, proliferation, differentiation and aging. In particular, autophagy plays a pivotal role in regulating the function of T helper lymphocytes, regulating their secretion of cytokines in chronic inflammatory diseases [161].
Unfortunately, the aging process is associated with a decrease in autophagy, which further contributes to inflammaging, senescence and aging itself [153,158]. Decreased autophagy has been linked to a wide range of age-related inflammatory diseases, including cardiomyopathy, neurodegenerative disorders and cancer [141,142,162]. Therefore, compounds capable of promoting autophagy have recently received great attention for the treatment of age-related diseases [12].
4.2. HTyr as a Modulatory Agent of Autophagy
Polyphenols have recently been recognized as agents capable of modulating autophagy, improving thus cell homeostasis and function [163,164,165,166,167,168,169,170,171]. As for investigations concerning the modulation of inflammation by HTyr, most laboratories, including ours, performed investigations on the modulation of autophagy using HTyr-enriched preparations, which do not allow the identification of the effect mediated by the single HTyr compound [55,172,173]. Therefore, here we reference investigations that used purified HTyr to analyze its in vitro (Table 3) and in vivo (Table 4) capability to regulate the inflammaging markers through modulation of autophagy.
Table 3.
In vitro effects of HTyr on autophagy.
| HTyr Treatment and Dose |
Model | Effects | Ref. |
|---|---|---|---|
| Pre-treatment 25, 50, 100 μM |
Rat Arthritis: ex vivo primary chondrocytes + TNF-α |
↓ IL-1β, IL-6 through ↑ autophagy via SIRT6 |
[174] |
| Pre-treatment 75 μM |
Rat Arthritis: ex vivo primary condrocytes + AOPPs |
↓ IL-6, TNF-α through ↑ autophagy via SIRT1 |
[175] |
| Pre-treatment 25, 50, 100 μM |
Rat Vasculitis: ex vivo primary vascular adventitial fibroblasts + TNF-α |
↓ IL-1 β, IL-6, MCP-1 through ↑ autophagy via SIRT1-mediated ↓ AKT/mTOR |
[176] |
| Pre-treatment 1 μM |
Chemical Carcinogenesis in Human Primary Colonic Epithelial Cells: HCoEpC + B[a]P |
↓ IL-6, Il-8, VEGF, CXCL13 ↑ autophagy via ↓ 4EBP1 phosphorylation (↓ mTOR pathway) |
[131] |
| Pre-treatment 75 μM |
Rat Alopecia: primary dermal papilla cells + H2O2 |
↓ IL-6, TNF-α via ↑ autophagy ↑ FGF, PDGF |
[177] |
Abbreviations: TNF: tumor necrosis factor. IL: interleukin. MCP-1: monocyte chemoattractant protein-1. CXCL: C-X-C motif ligand. VEGF: vascular endothelial growth factor. AOPPs: advanced oxidation protein products. SIRT: silent mating type information regulator. AKT/mTOR: Ak strain transforming/mammalian target of rapamycin complex. 4EBP1: eukaryotic translation initiation factor 4E (eIF4E)-binding protein 1. B[a]P: Benzo[a]pyrene. ↓ Decrease. ↑ Increase.
Table 4.
In vivo effects of HTyr on autophagy.
| HTyr Treatment and Dose |
Model | Effects | Ref. |
|---|---|---|---|
| 100 mg/kg intragastrically | Murine Acute Lung Injury intranasal LPS-induced: lung tissue bronchoalveolar lavage fluid |
↓ TNF-α, IL-1β, IL-6, MCP-1, IL-10 ↑ autophagy via SIRT-1 ↓ MAPK phosphorylation ↓ neutrophil and lymphocyte infiltration |
[178] |
| 50 mg/kg of diet for 8 weeks | Murine Alzheimer’s disease in TgCRND8 mice: brain tissues |
↓ TNF-α ↓ amyloid protein load ↑ autophagy ↑ cognitive function: memory |
[179] |
Abbreviations: LPS: lipopolysaccharide. TNF: tumor necrosis factor. IL: interleukin. MCP-1: monocyte chemoattractant protein-1. SIRT: silent mating type information regulator. PI3K/AKT: phosphatidylinositol 3-kinase/Ak strain transforming. MAPK: mitogen-activated protein kinase. Tg: transgenic. ↓ Decrease ↑ Increase.
4.2.1. Evidence In Vitro
Two studies exist indicating the capability of HTyr to affect inflammation and inflammaging markers by modulating autophagy in experimental models of chondrocytes stimulated by different inflammatory stimuli [174,175] (Table 3). Zhi et al. demonstrated that HTyr (25, 50, 100 μM) inhibited, in a dose-dependent manner, the levels of pro-inflammatory cytokines such as IL-1β and IL-6 as well as the chemokine MCP-1 in ex vivo rat primary chondrocytes stimulated with TNF-α. In addition, HTyr promoted autophagy and upregulated the expression of SIRT6 at the mRNA and protein level. Moreover, SIRT6 knockdown reduced the effect of HTyr on autophagy promotion by decreasing the expression of the LC3 autophagic marker. Furthermore, both the inhibition of autophagy by the autophagic inhibitor 3-methyladenine and SIRT6 knockdown decreased the suppressive effects of HTyr on the inflammatory response in TNF-α-induced chondrocytes. Overall, these findings indicate that HTyr inhibits TNF-α-induced inflammatory response in chondrocytes through regulating SIRT6-mediated autophagy, thus protecting chondrocytes from inflammation and suggesting potential beneficial effects of HTyr in osteoarthritis [174]. Then, Sun et al. investigated the anti-inflammatory effect of HTyr (75 μM) pre-treatment in rat primary chondrocytes stimulated with advanced oxidation protein products (AOPPs). The investigators found that HTyr increased the expression of LC3 and autophagy related (ATG) 5 and ATG 7 molecules, while it decreased the expression of p62, thus promoting autophagy in AOPP-stimulated chondrocytes. It was also demonstrated that autophagy was promoted through the SIRT1 pathway. Furthermore, HTyr inhibited the expression of IL-6 and TNF-α inflammatory cytokines at the mRNA and protein level, and this inhibition was suppressed by SIRT1 knockdown, showing that HTyr can inhibit the inflammatory response caused by stressed chondrocytes through the promotion of SIRT1-mediated autophagy [175].
A recent study investigated the mechanisms underlying the anti-inflammatory effect of HTyr in cardiovascular diseases, by analyzing the capability of HTyr to affect the inflammatory response in vascular adventitial fibroblasts (VAFs) through autophagy modulation (Table 3). Pre-treatment of TNF-α-stimulated rat primary VAFs (obtained from thoracic aorta) with HTyr (25, 50, 100 μM) promoted autophagy by increasing LC3 cell expression and the autophagic flux. In addition, HTyr upregulated SIRT1 mRNA and protein expression in TNF-α-stimulated VAFs, in a dose-dependent manner. It was also demonstrated that HTyr decreased Akt phosphorylation, and that the inhibition of both Akt and mTOR inhibited the promotion of autophagy by HTyr, indicating that HTyr modulated autophagy through SIRT1-mediated Akt/mTOR suppression. Furthermore, HTyr inhibited the TNF-α-induced inflammatory response in VAFs by inhibiting the secretion of IL-1 β and the expression of IL-6 and MCP-1 mRNA, while SIRT1 knockdown decreased both autophagy and inflammatory inhibition by HTyr. Therefore, these findings indicate that HTyr inhibits the inflammatory response produced by VAFs via the promotion of autophagy through SIRT1-mediated Akt/mTOR pathway suppression [176].
Another study analyzed the autophagic modulatory activity of HTyr in a human model of colonic chemical carcinogenesis (Table 3). HTyr (1 μM) pre-treatment of HCoEpC cells exposed to [B[a]P] restored B[a]P-mediated reduction in cell autophagy by increasing LC3 and reducing SQSTM1/p62 expression levels. HTyr also counteracted the phosphorylation of 4EBP1 (a downstream target of mTOR), thus counteracting mTOR activation. The modulation of autophagy by HTyr was associated with the inhibition of the release of the IL-6 pro-inflammatory cytokine, IL-8 and CXCL13 chemokines, and VEGF by HCoEpC cells exposed to [B[a]P]. These results suggest that HTyr might exert tumor suppression through the promotion of autophagy, thus playing an important role in preventing tumorigenesis by chemical carcinogens [131].
Lastly, Chen et al. explored the potential protective actions of HTyr-promoted autophagy in a rat model of stress-induced alopecia (Table 3). HTyr (75 μM) pre-treatment of H2O2-exposed rat primary dermal papilla cells promoted autophagy, inhibited mRNA and protein levels of IL-6 and TNF-α and enhanced the levels of hair growth factors such as the fibroblast growth factor (FGF) and platelet-derived growth factor (PDGF). By the use of the chloroquine autophagic inhibitor, it was also found that the anti-inflammatory effect was dependent on the regulation of autophagy by HTyr. These results indicate that HTyr is capable of significantly reducing stress-induced inflammation and that this effect is mediated by its capability to promote autophagy [177].
4.2.2. Evidence In Vivo
The in vivo regulation of inflammation and inflammaging markers through the modulation of autophagy by HTyr has been investigated in two inflammatory experimental animal models [178,179] (Table 4). Yang et al. investigated the potential modulatory effect of HTyr on autophagy in a murine model of LPS-induced acute lung injury. Male BALB/c mice, challenged with intranasal instillations of LPS, were treated with HTyr (100 mg/kg) intragastrically 1 h prior to LPS exposure. Twenty-four hours later, lung and bronchoalveolar lavage fluid samples were obtained and analyzed for inflammatory and autophagic markers. LPS-driven release of TNF-α, IL-1β, IL-6, IL-10 and MCP-1 was strongly suppressed by HTyr. In addition, LPS-stimulated SIRT1 inhibition, MAPK phosphorylation and autophagy suppression were all abolished by HTyr administration. HTyr treatment significantly decreased pulmonary edema and inflammatory cell infiltration into lung tissues as well as inflammatory cell levels in bronchoalveolar fluid, suggesting that the protective effect of HTyr on lung inflammation may be attributed to the promotion of autophagy, which is likely associated with the activation of SIRT and MAPK signaling pathways [178]. Then, Nardiello et al. indicated the capability of HTyr to inhibit inflammation in a murine model of Alzheimer’s disease through the promotion of autophagy (Table 4). Indeed, in 4-month-old Tg (transgenic) CRND8 (overexpressing mutant human amyloid precursor protein) mice, treated for 8 weeks with a low-fat diet supplemented with HTyr (50 mg/kg of diet), a marked reduction of TNF-α expression in hippocampal areas associated with a strong promotion of autophagy in brain tissues was found, as indicated by the bright LC3 staining in the neuronal cell bodies and processes of neurons in the parietal cortex. These results were associated with a significant improvement of murine cognitive status and the reduction of plaque area and number of the amyloid aggregate of Aβ42 peptide and its pyroglutamated 3-42 derivative (pE3-Aβ) in the cortex and in the hippocampus, suggesting that HTyr mediates neuroprotection by modulating autophagy [179].
5. Discussion and Conclusions
In this review, we reference studies investigating the capability of purified HTyr to counteract the rise of multiple inflammatory mediators of inflammaging, by modulating inflammation and autophagy.
Encouraging results have been obtained concerning the modulation of inflammation by the administration of purified HTyr in different experimental models of inflammation in vitro and in vivo, indicating that HTyr can interfere with inflammaging by modulating inflammation. In fact, HTyr decreases inflammatory biomarkers of inflammaging, such as pro-inflammatory cytokines, chemokines and acute-phase proteins, by affecting different steps of the inflammatory process in vitro and in vivo. Indeed, HTyr can downmodulate the expression of the TLR-4 innate immune receptor (Table 1). HTyr also inhibits the activation of different intracellular signaling molecules involved in the inflammatory response, including ERKs, JNK and p38 MAPK, which belong to the MAPK pathway (Table 1 and Table 2). HTyr can suppress the activation of NF-κB, the master regulator of inflammatory gene transcription, as well as the NLRP3 inflammasome, a critical component of the inflammatory response (Table 1 and Table 2). Of note, HTyr can also exert anti-inflammatory activity by modulating the composition and diversity of microbiota, in particular the gut microbiota (Table 2). Furthermore, the inflammation modulating activity of HTyr has been illustrated in vitro in different inflammatory cell experimental models, including innate immune cells such as macrophages and microglia, as well as non-immune cells such as senescent fibroblasts, keratinocytes and colonic epithelial cells (Table 1). However, contrasting results have been reported regarding the capability of HTyr of inhibiting inflammation mediated by monocytes/macrophages (Table 1). We think that the diverse results might be related to the different experimental conditions and protocols (i.e., monocyte/macrophage experimental models, HTyr dose, times of HTyr and LPS incubations, times in which samples were taken) used by independent investigators, while Pojero et al. suggested that it might depend on the timing of HTyr administration (i.e., before, during or after the incubation with LPS inflammatory stimuli) [97]. Further experiments are needed to explain these contrasting results. The inflammation modulating activity of HTyr has also been illustrated in vivo in different experimental animal models of inflammatory diseases, by analyzing ex vivo and in vivo macrophages, blood samples, as well as renal, liver, heart, brain and colon tissues (Table 2). Interestingly, the capability of HTyr to affect M1/M2 macrophage/microglia polarization in vitro and in vivo has also been reported, highlighting the capability of HTyr to increase the activation of the M2 anti-inflammatory macrophage phenotype associated with the rise of anti-inflammatory cytokines (Table 1 and Table 2).
Promising results have also been reported concerning the possible interference of HTyr with inflammaging through the modulation of autophagy (Table 3 and Table 4). However, a small number of studies analyzed this issue, especially in in vivo experimental models (Table 4). It has been clearly shown that HTyr is capable of inhibiting the rise of inflammaging markers such as cytokines, chemokines and growth factors through the promotion of autophagy, by affecting the activation of regulatory molecules of the autophagic process in vitro and in vivo (Table 3 and Table 4). In fact, HTyr can upregulate autophagy by activating AMPK or inhibiting the Akt/mTOR pathway, via SIRT1 and SIRT6 modulation (Table 3 and Table 4). Furthermore, the capability of HTyr to decrease the expression of pro-inflammatory mediators through the promotion of autophagy has been observed in different experimental models, including arthritis, vasculitis and colonic chemical carcinogenesis in vitro, as well as acute lung injury and Alzheimer’s disease in vivo. However, although the capability of upregulating autophagy by HTyr-enriched preparations has been established in inflammatory immune cells such as macrophages [55], to the best of our knowledge, there is no study that has investigated the capability of pure HTyr to modulate autophagy in immune cells, including macrophages. Therefore, there is a need for studies on the capability of pure HTyr to interfere with inflammaging through modulating autophagy in innate and adaptive immune cell-mediated inflammatory responses.
Overall, our knowledge on inflammaging and its relationship with inflammation and autophagy is far from being complete, and there is still much to learn about the capability of pure HTyr to interfere with inflammaging via the modulation of inflammation and autophagy [180]. We consider that a certain number of critical issues, including the effective dosage and time of HTyr administration, have to be addressed before proceeding with clinical trials based on the administration of pure HTyr in elderly. In fact, the low bioavailability of orally ingested polyphenols, including HTyr, greatly limits their therapeutic use, particularly in those organs distant from the gastrointestinal tract. In addition, most of the in vitro and in vivo studies performed so far suggest a possible preventive rather than therapeutic activity of HTyr against inflammaging, in that in most studies, HTyr was administered before rather than after the inflammatory stimuli.
In conclusion, we consider that the promising results, emerged from preclinical investigations, strengthen a need for further studies aimed to better characterize the interference of HTyr with inflammaging to predict the possible effective clinical use of this compound in aging and age-related disorders.
Author Contributions
Conceptualization, F.V.; writing and review, F.V. and R.B.; editing, R.B. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data was used for the research described in the article.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
This review did not receive any specific grant from funding agencies in the public, commercial or not-for-profit sectors.
Footnotes
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
References
- 1.Lopez-Otin C., Blasco M.A., Partridge L., Serrano M., Kroemer G. The hallmarks of aging. Cell. 2013;153:1194–1217. doi: 10.1016/j.cell.2013.05.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Franceschi C., Bonafe M., Valensin S., Olivieri F., De Luca M., Ottaviani E., De Benedictis G. Inflammaging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000;908:244–254. doi: 10.1111/j.1749-6632.2000.tb06651.x. [DOI] [PubMed] [Google Scholar]
- 3.Cevenini E., Caruso C., Candore G., Capri M., Nuzzo D., Duro G., Rizzo C., Colonna-Romano G., Lio D., Di Carlo D., et al. Age-related inflammation: The contribution of different organs, tissues and systems. How to face it for therapeutic approaches. Curr. Pharm. Des. 2010;16:609–618. doi: 10.2174/138161210790883840. [DOI] [PubMed] [Google Scholar]
- 4.Franceschi C., Garagnani P., Vitale G., Capri M., Salvioli S. Inflammaging and Garbaging. Trends Endocrinol. Metab. 2017;28:199–212. doi: 10.1016/j.tem.2016.09.005. [DOI] [PubMed] [Google Scholar]
- 5.Franceschi C., Garagnani P., Parini P., Giuliani C., Santoro A. Inflammaging: A new immune-metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 2018;14:576–590. doi: 10.1038/s41574-018-0059-4. [DOI] [PubMed] [Google Scholar]
- 6.Rasa S.M.M., Annunziata F., Krepelova A., Nunna S., Omrani O., Gebert N., Adam L., Kappel S., Höhn S., Donati G., et al. Inflammaging is driven by upregulation of innate immune receptors and systemic interferon signaling and is ameliorated by dietary restriction. Cell Rep. 2022;39:111017. doi: 10.1016/j.celrep.2022.111017. [DOI] [PubMed] [Google Scholar]
- 7.Salminen A., Kaarniranta K. Regulation of the aging process by autophagy. Trends Mol. Med. 2009;15:217–224. doi: 10.1016/j.molmed.2009.03.004. [DOI] [PubMed] [Google Scholar]
- 8.Rajawat Y.S., Hilioti Z., Bossis I. Aging: Central role for autophagy and the lysosomal degradative system. Ageing Res. Rev. 2009;8:199–213. doi: 10.1016/j.arr.2009.05.001. [DOI] [PubMed] [Google Scholar]
- 9.Hansen M., Rubinsztein D.C., Walker D.W. Autophagy as a promoter of longevity: Insights from model organisms. Nat. Rev. Mol. Cell Biol. 2018;19:579–593. doi: 10.1038/s41580-018-0033-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Levine B., Mizushima N., Virgin H.W. Autophagy in immunity and inflammation. Nature. 2011;469:323–335. doi: 10.1038/nature09782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Deretic V. Autophagy in inflammation, infection, and immunometabolism. Immunity. 2021;54:437–453. doi: 10.1016/j.immuni.2021.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zinecker H., Simon A.K. Autophagy takes it all-autophagy inducers target immune aging. Dis. Model Mech. 2022;15:dmm049345. doi: 10.1242/dmm.049345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Krabbe K.S., Pedersen M., Bruunsgaard H. Inflammatory mediators in the elderly. Exp. Gerontol. 2004;39:687–699. doi: 10.1016/j.exger.2004.01.009. [DOI] [PubMed] [Google Scholar]
- 14.Wei J., Xu H., Davies J.L., Hemmings G.P. Increase of plasma IL-6 concentration with age in healthy subjects. Life Sci. 1992;51:1953–1956. doi: 10.1016/0024-3205(92)90112-3. [DOI] [PubMed] [Google Scholar]
- 15.Singh T., Newman A.B. Inflammatory markers in population studies of aging. Ageing Res. Rev. 2010;10:319–329. doi: 10.1016/j.arr.2010.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Koelman L., Pivovarova-Ramich O., Pfeiffer A.F.H., Grune T., Aleksandrova K. Cytokines for evaluation of chronic inflammatory status in ageing research: Reliability and phenotypic characterisation. Immun. Ageing. 2019;16:11. doi: 10.1186/s12979-019-0151-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hartmann A., Hartmann C., Secci R., Hermann A., Fuellen G., Walter M. Ranking biomarkers of aging by citation profiling and effort scoring. Front. Genet. 2021;12:686320. doi: 10.3389/fgene.2021.686320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kaneko N., Kurata M., Yamamoto T., Morikawa S., Masumoto J. The role of interleukin-1 in general pathology. Inflamm. Regen. 2019;39:12. doi: 10.1186/s41232-019-0101-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sethi G., Sung B., Aggarwal B.B. TNF: A master switch for inflammation to cancer. Front. Biosci. 2008;13:5094–5107. doi: 10.2741/3066. [DOI] [PubMed] [Google Scholar]
- 20.Webster J.D., Vucic D. The balance of TNF mediated pathways regulates inflammatory cell death signaling in healthy and diseased tissues. Front. Cell Dev. Biol. 2020;8:365. doi: 10.3389/fcell.2020.00365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Minciullo P.L., Catalano A., Mandraffino G., Casciaro M., Crucitti A., Maltese G., Morabito N., Lasco A., Gangemi S., Basile G. Inflammaging and anti-Inflammaging: The role of cytokines in extreme longevity. Arch. Immunol. Ther. Exp. (Warsz) 2016;64:111–126. doi: 10.1007/s00005-015-0377-3. [DOI] [PubMed] [Google Scholar]
- 22.Rea I.M., Gibson D.S., McGilligan V., McNerlan S.E., Alexander H.D., Ross O.A. Age and age-related diseases: Role of inflammation triggers and cytokines. Front. Immunol. 2018;9:586. doi: 10.3389/fimmu.2018.00586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Boka G., Anglade P., Wallach D., Javoy-Agid F., Agid Y., Hirsch E.C. Immunocytochemical analysis of tumor necrosis factor and its receptors in Parkinson’s disease. Neurosci. Lett. 1994;172:151–154. doi: 10.1016/0304-3940(94)90684-X. [DOI] [PubMed] [Google Scholar]
- 24.Mogi M., Harada M., Kondo T., Riederer P., Inagaki H., Minami M., Nagatsu T. Interleukin-1 beta, interleukin-6, epidermal growth factor and transforming growthfactor-alpha are elevated in the brain from parkinsonian patients. Neurosci. Lett. 1994;180:147–150. doi: 10.1016/0304-3940(94)90508-8. [DOI] [PubMed] [Google Scholar]
- 25.Diaz K., Kohut M.L., Russell D.W., Stegemoller E.L. Peripheral inflammatory cytokines and motor symptoms in persons with Parkinson’s disease. Brain Behav. Immun. Health. 2022;21:100442. doi: 10.1016/j.bbih.2022.100442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gerard C., Rollins B.J. Chemokines and disease. Nat. Immunol. 2001;2:108–115. doi: 10.1038/84209. [DOI] [PubMed] [Google Scholar]
- 27.Hickman S.E., El Khoury J. Mechanisms of mononuclear phagocyte recruitment in Alzheimer’s disease. CNS Neurol. Disord. Drug Targets. 2010;9:168–173. doi: 10.2174/187152710791011982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Altara R., Manca M., Hessel M.H., Gu Y., van Vark L.C., Akkerhuis K.M., Staessen J.A., Struyker Boudier H., Booz G.W., Blankesteijn M. CXCL10 is a circulating inflammatory marker in patients with advanced heart failure: A pilot study. J. Cardiovasc. Transl. Res. 2016;9:302–314. doi: 10.1007/s12265-016-9703-3. [DOI] [PubMed] [Google Scholar]
- 29.Corsini E., Vismara L., Lucchi L., Viviani B., Govoni S., Galli C.L., Marinovich M., Racchi M. High interleukin-10 production is associated with low antibody response to influenza vaccination in the elderly. J. Leukoc. Biol. 2006;80:376–382. doi: 10.1189/jlb.0306190. [DOI] [PubMed] [Google Scholar]
- 30.Almanan M., Raynor J., Ogunsulire I., Malyshkina A., Mukherjee S., Hummel S.A., Ingram J.T., Saini A., Xie M.M., Alenghat T., et al. IL-10–producing Tfh cells accumulate with age and link inflammation with age-related immune suppression. Sci. Adv. 2020;6:eabb0806. doi: 10.1126/sciadv.abb0806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fulop T., Larbi A., Witkowski J.M. Human inflammaging. Gerontology. 2019;65:495–504. doi: 10.1159/000497375. [DOI] [PubMed] [Google Scholar]
- 32.Ferrucci L., Fabbri E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018;9:505–522. doi: 10.1038/s41569-018-0064-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Barcena M.L., Aslam M., Pozdniakova S., Norman K., Ladilov Y. Cardiovascular inflammaging: Mechanisms and translational aspects. Cells. 2022;11:1010. doi: 10.3390/cells11061010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Aguilera Y., Martin-Cabrejas M.A., Gonzalez de Mejia E. Phenolic compounds in fruits and beverages consumed as part of the Mediterranean diet: Their role in prevention of chronic diseases. Phytochem. Rev. 2016;15:405–423. doi: 10.1007/s11101-015-9443-z. [DOI] [Google Scholar]
- 35.Visioli F., Galli C. Olive oil phenols and their potential effects on human health. J. Agric. Food Chem. 1998;46:4292–4296. doi: 10.1021/jf980049c. [DOI] [Google Scholar]
- 36.Owen R.W., Haubner R., Wurtele G., Hull E., Spiegelhalder B., Bartsch H. Olives and olive oil in cancer prevention. Eur. J. Cancer Prev. 2004;13:319–326. doi: 10.1097/01.cej.0000130221.19480.7e. [DOI] [PubMed] [Google Scholar]
- 37.Servili M., Esposto S., Fabiani R., Urbani S., Taticchi A., Mariucci F., Selvaggini R., Montedoro G.F. Phenolic compounds in olive oil: Antioxidant, health and organoleptic activities according to their chemical structure. Inflammopharmacology. 2009;17:76–84. doi: 10.1007/s10787-008-8014-y. [DOI] [PubMed] [Google Scholar]
- 38.Romani A., Ieri F., Urciuoli S., Noce A., Marrone G., Nediani C., Bernini R. Health effects of phenolic compounds found in extra-virgin olive oil, by-products, and leaf of Olea europaea L. Nutrients. 2019;11:1776. doi: 10.3390/nu11081776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.EFSA Scientific Opinion. [(accessed on 3 March 2023)]. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/2033.
- 40.Lucarini M., Durazzo A., Bernini R., Campo M., Vita C., Souto E.B., Lombardi-Boccia G., Ramadan M.F., Santini A., Romani A. Fruit wastes as valuable source of value-added compounds: A collaborative perspective. Molecules. 2021;26:6338. doi: 10.3390/molecules26216338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pineiro Z., Cantos-Villar E., Palma M., Puertas B. Direct liquid chromatography method for the simultaneous quantification of hydroxytyrosol and tyrosol in red wines. J. Agric. Food Chem. 2011;59:11683–11689. doi: 10.1021/jf202254t. [DOI] [PubMed] [Google Scholar]
- 42.Bernini R., Merendino N., Romani A., Velotti F. Naturally occurring hydroxytyrosol: Synthesis and anticancer potential. Curr. Med. Chem. 2013;20:655–670. doi: 10.2174/092986713804999367. [DOI] [PubMed] [Google Scholar]
- 43.Hu T., He X.W., Jiang J.G., Xu X.L. Hydroxytyrosol and its potential therapeutic effects. J. Agric. Food Chem. 2014;62:1449–1455. doi: 10.1021/jf405820v. [DOI] [PubMed] [Google Scholar]
- 44.Robles-Almazan M., Pulido-Moran M., Moreno-Fernandez J., Ramirez-Tortosa C., Rofriguez-Garcia C., Quiles J.L., Ramirez-Tortosa M.C. Hydroxytyrosol: Bioavailability, toxicity and clinical applications. Food Res. Int. 2018;105:654–667. doi: 10.1016/j.foodres.2017.11.053. [DOI] [PubMed] [Google Scholar]
- 45.D’Andrea G., Ceccarelli M., Bernini R., Clemente M., Santi L., Caruso C., Micheli L., Tirone F. Hydroxytyrosol stimulates neurogenesis in aged dentate gyrus by enhancing stem and progenitor cell proliferation and neuron survival. FASEB J. 2020;34:4512–4526. doi: 10.1096/fj.201902643R. [DOI] [PubMed] [Google Scholar]
- 46.Romanucci V., Garcia-Vinuales S., Tempra S., Bernini R., Zarrelli A., Lolicato F., Minardi D., Di Fabio G. Modulating Aβ aggregation by tyrosol-based ligands: The crucial role of the catechol moiety. Biophys. Chem. 2020;265:106434. doi: 10.1016/j.bpc.2020.106434. [DOI] [PubMed] [Google Scholar]
- 47.Laghezza Masci V., Bernini R., Villanova N., Clemente M., Cicaloni V., Tinti L., Salvini L., Taddei A.R., Tiezzi A., Ovidi E. In vitro anti-proliferative and apoptotic effects of hydroxytyrosyl oleate on SH-SY5Y human neuroblastoma cells. Int. J. Mol. Sci. 2022;23:12348. doi: 10.3390/ijms232012348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lan X., Chang K., Zeng L., Liu X., Qiu F., Zheng W., Quan H., Liao Z., Chen M., Huang W., et al. Engineering salidroside biosynthetic pathway in hairy root cultures of Rhodiola crenulata based on metabolic characterization of tyrosine decarboxylase. PLoS ONE. 2013;8:e75459. doi: 10.1371/journal.pone.0075459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gambacorta A., Tofani D., Bernini R., Migliorini A. High yielding preparation of a stable precursor of hydroxytyrosol by total synthesis and from the natural glucoside oleuropein. J. Agric. Food Chem. 2007;55:3386–3391. doi: 10.1021/jf063353b. [DOI] [PubMed] [Google Scholar]
- 50.Vissers M.N., Zock P.L., Roodenburg A.J.C., Leenen R., Katan M.B. Olive oil phenols are absorbed in humans. J. Nutr. 2002;132:409–417. doi: 10.1093/jn/132.3.409. [DOI] [PubMed] [Google Scholar]
- 51.Mosele J.I., Martin-Pelaez S., Macia A., Farras M., Valls R.-M., Catalan U., Motilva M.-J. Faecal microbial metabolism of olive oil phenolic compounds: In vitro and in vivo approaches. Mol. Nutr. Food Res. 2014;58:1809–1819. doi: 10.1002/mnfr.201400124. [DOI] [PubMed] [Google Scholar]
- 52.Romani A., Pinelli P.F., Ieri Bernini R. Sustainability, innovation and green chemistry in the production and valorization of phenolic extracts from Olea europaea L. Sustainability. 2016;8:1002. doi: 10.3390/su8101002. [DOI] [Google Scholar]
- 53.Bernini R., Carastro I., Palmini G., Tanini A., Zonefrati R., Pinelli P., Brandi M.L., Romani A. Lipophilization of Hydroxytyrosol-enriched fractions from Olea europaea L. by-products and evaluation of the in vitro effects on a model of colorectal cancer cells. J. Agric. Food Chem. 2017;65:6506–6512. doi: 10.1021/acs.jafc.6b05457. [DOI] [PubMed] [Google Scholar]
- 54.Luzi F., Pannucci E., Clemente M., Grande E., Urciuoli S., Romani A., Torre L., Puglia D., Bernini R., Santi L. Hydroxytyrosol and oleuropein-enriched extracts obtained from olive oil wastes and by-products as active antioxidant ingredients for poly(vinyl alcohol)-based films. Molecules. 2021;26:2104. doi: 10.3390/molecules26072104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Meschini R., D’Eliseo D., Filippi S., Bertini L., Bizzarri B.M., Botta L., Saladino R., Velotti F. Tyrosinase-treated hydroxytyrosol-enriched olive vegetation waste with increased antioxidant activity promotes autophagy and inhibits the inflammatory response in human THP-1 monocytes. J. Agric. Food Chem. 2018;66:12274–12284. doi: 10.1021/acs.jafc.8b03630. [DOI] [PubMed] [Google Scholar]
- 56.Dickinson J.R., Salgado L.E.J., Hewlins M.J.E. The catabolism of amino acids to long chain and complex alcohols in Saccharomyces cerevisiae. J. Biol. Chem. 2003;278:8028–8034. doi: 10.1074/jbc.M211914200. [DOI] [PubMed] [Google Scholar]
- 57.Tuck K.L., Freeman M.P., Hayball P.J., Stretch G.L., Stupans I. The in vivo fate of hydroxytyrosol and tyrosol, antioxidant phenolic constituents of olive oil, after intravenous and oral dosing of labeled compounds to rats. J. Nutr. 2001;131:1993–1996. doi: 10.1093/jn/131.7.1993. [DOI] [PubMed] [Google Scholar]
- 58.Dominguez-Perles R., Aunon D., Ferreres F., Gil-Izquierdo A. Gender differences in plasma and urine metabolites from Sprague-Dawley rats after oral administration of normal and high doses of hydroxytyrosol, hydroxytyrosol acetate, and DOPAC. Eur. J. Nutr. 2017;56:215–224. doi: 10.1007/s00394-015-1071-2. [DOI] [PubMed] [Google Scholar]
- 59.Serra A., Rubio L., Borras X., Macia A., Romero M.-P., Motilva M.J. Distribution of olive oil phenolic compounds in rat tissues after administration of a phenolic extract from olive cake. Mol. Nutr. Food Res. 2012;56:486–496. doi: 10.1002/mnfr.201100436. [DOI] [PubMed] [Google Scholar]
- 60.Visioli F., Galli C., Grande S., Colonnelli K., Patelli C., Galli G., Caruso D. Hydroxytyrosol excretion differs between rats and humans and depends on the vehicle of administration. J. Nutr. 2003;133:2612–2615. doi: 10.1093/jn/133.8.2612. [DOI] [PubMed] [Google Scholar]
- 61.Aunon-Calles D., Canut L., Visioli F. Toxicological evaluation of pure hydroxytyrosol. Food Chem Toxicol. 2013;55:498–504. doi: 10.1016/j.fct.2013.01.030. [DOI] [PubMed] [Google Scholar]
- 62.Aunon-Calles D., Canut L., Visioli F. Hydroxytyrosol is not genotoxic in vitro. Pharm. Res. 2013;74:87–93. doi: 10.1016/j.phrs.2013.06.002. [DOI] [PubMed] [Google Scholar]
- 63.Siracusa R., Scuto M., Fusco R., Trovato A., Ontario M.L., Crea R., Di Paola R., Cuzzocrea S., Calabrese V. Anti-inflammatory and antioxidant activity of Hidrox® in rotenone-induced Parkinson’s disease in mice. Antioxidants. 2020;9:824. doi: 10.3390/antiox9090824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bernini R., Mincione E., Barontini M., Crisante F. Convenient synthesis of hydroxytyrosol and its lipophilic derivatives from tyrosol or homovanillyl alcohol. J. Agric. Food Chem. 2008;56:8897–8904. doi: 10.1021/jf801558z. [DOI] [PubMed] [Google Scholar]
- 65.Bernini R., Fabrizi G., Pouysegu L., Deffieux D., Quideau S. Synthesis of biologically active catecholic compounds via ortho-selective oxygenation of phenolic compounds using hypervalent iodine(V) reagents. Curr. Org. Synth. 2012;9:650–669. doi: 10.2174/157017912803251792. [DOI] [Google Scholar]
- 66.Bozzini T., Botta G., Delfino M., Onofri S., Saladino R., Amatore D., Sgarbanti R., Nencioni L., Palamara A.T. Tyrosinase and Layer-by-Layer supported tyrosinases in the synthesis of lipophilic catechols with antiinfluenza activity. Bioorg. Med. Chem. 2013;21:7699–7708. doi: 10.1016/j.bmc.2013.10.026. [DOI] [PubMed] [Google Scholar]
- 67.Bernini R., Crisante F., Merendino N., Molinari R., Soldatelli M.C., Velotti F. Synthesis of a novel ester of hydroxytyrosol and α-lipoic acid exhibiting an antiproliferative effect on human colon cancer HT-29 cells. Eur. J. Med. Chem. 2011;46:439–446. doi: 10.1016/j.ejmech.2010.10.028. [DOI] [PubMed] [Google Scholar]
- 68.Bernini R., Gilardini Montani M.S., Merendino N., Romani A., Velotti F. Hydroxytyrosol-derived compounds: A basis for the creation of new pharmacological agents for cancer prevention and therapy. J. Med. Chem. 2015;58:9089–9107. doi: 10.1021/acs.jmedchem.5b00669. [DOI] [PubMed] [Google Scholar]
- 69.Bernini R., Barontini M., Cis V., Carastro I., Tofani D., Chiodo R.A., Lupattelli P., Incerpi S. Synthesis and evaluation of the antioxidant activity of lipophilic phenethyl trifluoroacetate esters by in vitro ABTS, DPPH and in cell-culture DCF assays. Molecules. 2018;23:208. doi: 10.3390/molecules23010208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bernini R., Carastro I., Santoni F., Clemente M. Synthesis of lipophilic esters of tyrosol, homovanillyl alcohol and hydroxytyrosol. Antioxidants. 2019;8:174. doi: 10.3390/antiox8060174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Trujillo N., Mateos R., Collantes de Teran L., Espartero J.L., Cert R., Jover M., Alcudia F., Bautista J., Cert A., Parrado J. Lipophilic hydroxytyrosyl esters. Antioxidant activity in lipid matrices and biological systems. J. Agric. Food Chem. 2006;54:3779–3785. doi: 10.1021/jf060520z. [DOI] [PubMed] [Google Scholar]
- 72.Montoya T., Aparicio-Soto M., Castejón M.L., Rosillo M.A., Sánchez-Hidalgo M., Begines P., Fernández-Bolaños J.G., Alarcón-de-la-Lastra C. Peracetylated hydroxytyrosol, a new hydroxytyrosol derivate, attenuates LPS-induced inflammatory response in murine peritoneal macrophages via regulation of non-canonical inflammasome, Nrf2/HO1 and JAK/STAT signaling pathways. J. Nutr. Biochem. 2018;57:110–120. doi: 10.1016/j.jnutbio.2018.03.014. [DOI] [PubMed] [Google Scholar]
- 73.Medzhitov R. Origin and physiological roles of inflammation. Nature. 2008;454:428–435. doi: 10.1038/nature07201. [DOI] [PubMed] [Google Scholar]
- 74.Gupta S.C., Kunnumakkara A.B., Aggarwal S., Aggarwal B.B. Inflammation, a double-edge sword for cancer and other age-related diseases. Front. Immunol. 2018;9:2160. doi: 10.3389/fimmu.2018.02160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Franceschi C., Capri M., Monti D., Giunta S., Olivieri F., Sevini F., Panourgia M.P., Invidia L., Celani L., Scurti M., et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007;128:92–105. doi: 10.1016/j.mad.2006.11.016. [DOI] [PubMed] [Google Scholar]
- 76.Kovacs E.J., Palmer J.L., Fortin C.F., Fulop J.T., Goldstein D.R., Linton P.J. Aging and innate immunity in the mouse: Impact of intrinsic and extrinsic factors. Trends Immunol. 2009;30:319–324. doi: 10.1016/j.it.2009.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Bleve A., Motta F., Durante B., Pandolfo C., Selmi C., Sica A. Immunosenescence, inflammaging, and frailty: Role of myeloid cells in age-related diseases. Clin. Rev. Allergy Immunol. 2022;15:1–22. doi: 10.1007/s12016-021-08909-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Desdin-Mico G., Soto-Heredero G., Aranda J.F., Oller J., Carrasco E., Gabande-Rodriguez E., Blanco E.M., Alfranca A., Cusso L., Desco M., et al. T cells with dysfunctional mitochondria induce multimorbidity and premature senescence. Science. 2020;368:1371–1376. doi: 10.1126/science.aax0860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ramos G.C., van den Berg A., Nunes-Silva V., Weirather J., Peters L., Burkard M., Friedrich M., Pinnecker J., Abesser M., Heinze K.G., et al. Myocardial aging as a T-cell-mediated phenomenon. Proc. Natl. Acad. Sci. USA. 2017;114:E2420–E2429. doi: 10.1073/pnas.1621047114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Delgobo M., Heinrichs M., Hapke N., Ashour D., Appel M., Srivastava M., Heckel T., Spyridopoulos I., Hofmann U., Frantz S., et al. Terminally differentiated CD4+ T cells promote myocardial inflammaging. Front. Immunol. 2021;12:584538. doi: 10.3389/fimmu.2021.584538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Tabibzadeh S. Signaling pathways and effectors of aging. Front. Biosci. (Landmark Ed.) 2021;26:50–96. doi: 10.2741/4889. [DOI] [PubMed] [Google Scholar]
- 82.Ronkina N., Gaestel M. MAPK-activated protein kinases: Servant or partner? Annu. Rev. Biochem. 2022;91:505–540. doi: 10.1146/annurev-biochem-081720-114505. [DOI] [PubMed] [Google Scholar]
- 83.Salminen A., Huuskonen J., Ojala J., Kauppinen A., Kaarniranta K., Suuronen T. Activation of innate immunity system during aging: NF-κB signaling is the culprit of inflamm-aging. Ageing Res. Rev. 2008;7:83–105. doi: 10.1016/j.arr.2007.09.002. [DOI] [PubMed] [Google Scholar]
- 84.Tilstra J.S., Clauson C.L., Niedernhofer L.J., Robbins P.D. NF-κB in Aging and Disease. Aging Dis. 2011;2:449–465. [PMC free article] [PubMed] [Google Scholar]
- 85.Osorio F., Soria-Valles C., Santiago-Fernández O., Freije J., López-Otín C. NF-κB signaling as a driver of ageing. Int. Rev. Cell Mol. Biol. 2016;326:133–174. doi: 10.1016/bs.ircmb.2016.04.003. [DOI] [PubMed] [Google Scholar]
- 86.Tavenier J., Rasmussen L.J.H., Houlind M.B., Andersen A.L., Panum I., Andersen O., Petersen J., Langkilde A., Nehlin J.O. Alterations of monocyte NF-kappaB p65/RelA signaling in a cohort of older medical patients, age-matched controls, and healthy young adults. Immun. Ageing. 2020;17:25. doi: 10.1186/s12979-020-00197-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.García-García V., Alameda J., Page A., Casanova M. Role of NF-κB in Ageing and Age-Related Diseases: Lessons from Genetically Modified Mouse Models. Cells. 2021;10:1906. doi: 10.3390/cells10081906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Martinon F., Mayor A., Tschopp J. The inflammasomes: Guardians of the body. Annu. Rev. Immunol. 2009;27:229–265. doi: 10.1146/annurev.immunol.021908.132715. [DOI] [PubMed] [Google Scholar]
- 89.Dinarello C.A. Immunological and inflammatory functions of the interleukin-1 family. Annu. Rev. Immunol. 2009;27:519–550. doi: 10.1146/annurev.immunol.021908.132612. [DOI] [PubMed] [Google Scholar]
- 90.Youm Y.H., Grant R.W., McCabe L.R., Albarado D.C., Nguyen K.Y., Ravussin A., Pistell P., Newman S., Carter R., Laque A., et al. Canonical Nlrp3 inflammasome links systemic low-grade inflammation to functional decline in aging. Cell Metab. 2013;18:519–532. doi: 10.1016/j.cmet.2013.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Heneka M.T., Kummer M.P., Stutz A., Delekate A., Schwartz S., Vieira-Saecker A., Griep A., Axt D., Remus A., Tzeng T.C., et al. NLRP3 is activated in Alzheimer’s disease and contributes to pathology in APP/PS1 mice. Nature. 2013;493:674–678. doi: 10.1038/nature11729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Duewell P., Kono H., Rayner K.J., Sirois C.M., Vladimer G., Bauernfeind F.G., Abela G.S., Franchi L., Nunez G., Schnurr M., et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature. 2010;464:1357–1361. doi: 10.1038/nature08938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Conway J.A., Duggal N. Ageing of the gut microbiome: Potential influences on immune senescence and inflammageing. Ageing Res. Rev. 2021;68:101323. doi: 10.1016/j.arr.2021.101323. [DOI] [PubMed] [Google Scholar]
- 94.Santoro A., Zhao J., Wu L., Carru C., Biagi E., Franceschi C. Microbiomes other than the gut: Inflammaging and age-related diseases. Semin. Immunopathol. 2020;42:589–605. doi: 10.1007/s00281-020-00814-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Yahfoufi N., Alsadi N., Jambi M., Matar C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients. 2018;10:1618. doi: 10.3390/nu10111618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Serreli G., Deiana M. Extra virgin olive oil polyphenols: Modulation of cellular pathways related to oxidant species and inflammation in aging. Cells. 2020;9:478. doi: 10.3390/cells9020478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Pojero F., Aiello A., Gervasi F., Caruso C., Ligotti M.E., Calabrò A., Procopio A., Candore G., Accardi G., Allegra M. Effects of oleuropein and hydroxytyrosol on inflammatory mediators: Consequences on inflammaging. Int. J. Mol. Sci. 2022;24:380. doi: 10.3390/ijms24010380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Chin K.Y., Pang K.L. Therapeutic effects of olive and its derivatives on osteoarthritis: From bench to bedside. Nutrients. 2017;9:1060. doi: 10.3390/nu9101060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Hornedo-Ortega R., Cerezo A.B., de Pablos R.M., Krisa S., Richard T., Garcia-Parrilla M.C., Troncoso A.M. Phenolic compounds characteristic of the Mediterranean diet in mitigating microglia-mediated neuroinflammation. Front. Cell. Neurosci. 2018;12:373. doi: 10.3389/fncel.2018.00373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ontario M.L., Siracusa R., Modafferi S., Scuto M., Sciuto S., Greco V., Bertuccio M.P., Trovato Salinaro A., Crea R., Calabrese E.J., et al. Potential prevention and treatment of neurodegenerative disorders by olive polyphenols and Hidrox. Mech. Ageing Dev. 2022;203:111637. doi: 10.1016/j.mad.2022.111637. [DOI] [PubMed] [Google Scholar]
- 101.Bitler C.M., Viale T.M., Damaj B., Crea R. Hydrolyzed olive vegetation water in mice has anti-inflammatory activity. J. Nutr. 2005;135:1475–1479. doi: 10.1093/jn/135.6.1475. [DOI] [PubMed] [Google Scholar]
- 102.Richard N., Arnold S., Hoeller U., Kilpert C., Wertz K., Schwager J. Hydroxytyrosol is the major anti-inflammatory compound in aqueous olive extracts and impairs cytokine and chemokine production in macrophages. Planta Med. 2011;77:1890–1897. doi: 10.1055/s-0031-1280022. [DOI] [PubMed] [Google Scholar]
- 103.Kondreddy V.K.R., Naidu K.A. Oleic acid, hydroxytyrosol and n-3 fatty acids collectively modulate colitis through reduction of oxidative stress and IL-8 synthesis; in vitro and in vivo studies. Int. Immunopharmacol. 2016;35:29–42. doi: 10.1016/j.intimp.2016.03.019. [DOI] [PubMed] [Google Scholar]
- 104.Mahmoudi A., Hadrich F., Feki I., Ghorbel H., Bouallagui Z., Marrekchi R., Fourati H., Sayadi S. Oleuropein and hydroxytyrosol rich extracts from olive leaves attenuate liver injury and lipid metabolism disturbance in bisphenol A-treated rats. Food Funct. 2018;9:3220–3234. doi: 10.1039/C8FO00248G. [DOI] [PubMed] [Google Scholar]
- 105.Martin M.E., Millan-Linares M.C., Naranjo M.C., Toscano R., Abia R., Muriana F.J.G., Bermudez B., Montserrat-de la Paz S. Minor compounds from virgin olive oil attenuate LPS-induced inflammation via visfatin-related gene modulation on primary human monocytes. J. Food Biochem. 2019;43:e12941. doi: 10.1111/jfbc.12941. [DOI] [PubMed] [Google Scholar]
- 106.Rosillo M.A., Alarcon-de-la-Lastra C., Castejon M.L., Montoya T., Cejudo-Guillen M., Sanchez-Hidalgo M. Polyphenolic extract from extra virgin olive oil inhibits the inflammatory response in IL-1β-activated synovial fibroblasts. Br. J. Nutr. 2019;121:55–62. doi: 10.1017/S0007114518002829. [DOI] [PubMed] [Google Scholar]
- 107.Fki I., Sayadi S., Mahmoudi A., Daoued I., Marrekchi R., Ghorbel H. Comparative study on beneficial effects of hydroxytyrosol- and oleuropein-rich olive leaf extracts on high-fat diet-induced lipid metabolism disturbance and liver injury in rats. BioMed Res. Int. 2020;2020:1315202. doi: 10.1155/2020/1315202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Hioki T., Tokuda H., Kuroyanagi G., Kim W., Tachi J., Matsushima-Nishiwaki R., Iida H., Kozawa O. Olive polyphenols attenuate TNF-α-stimulated M-CSF and IL-6 synthesis in osteoblasts: Suppression of Akt and p44/p42 MAP kinase signaling pathways. Biomed. Pharmacother. 2021;141:111816. doi: 10.1016/j.biopha.2021.111816. [DOI] [PubMed] [Google Scholar]
- 109.Leo M., Muccillo L., Dugo L., Bernini R., Santi L., Sabatino L. Polyphenols extracts from oil production waste products (OPWPs) reduce cell viability and exert anti-inflammatory activity via PPAR induction in colorectal cancer cells. Antioxidants. 2022;11:624. doi: 10.3390/antiox11040624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Manna C., Della Ragione F., Cucciolla V., Borriello A., D’Angelo S., Galletti P., Zappia V. Biological effects of hydroxytyrosol, a polyphenol from olive oil endowed with antioxidant activity. Adv. Exp. Med. Biol. 1999;472:115–130. doi: 10.1007/978-1-4757-3230-6_11. [DOI] [PubMed] [Google Scholar]
- 111.Bulotta S., Celano M., Lepore S.M., Montalcini T., Pujia A., Russo D. Beneficial effects of the olive oil phenolic components oleuropein and hydroxytyrosol: Focus on protection against cardiovascular and metabolic diseases. J. Transl. Med. 2014;12:219. doi: 10.1186/s12967-014-0219-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Silva A.F.R., Resende D., Monteiro M., Coimbra M.A., Silva A.M.S., Cardoso S.M. Application of hydroxytyrosol in the functional foods field: From ingredient to dietary supplements. Antioxidants. 2020;9:1246. doi: 10.3390/antiox9121246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.D’Angelo C., Franceschelli S., Quiles J.L., Speranza L. Wide biological role of hydroxytyrosol: Possible therapeutic and preventive properties in cardiovascular diseases. Cells. 2020;9:1932. doi: 10.3390/cells9091932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Bucciantini M., Leri M., Nardiello P., Casamenti F., Stefani M. Olive polyphenols: Antioxidant and anti-inflammatory properties. Antioxidants. 2021;10:1044. doi: 10.3390/antiox10071044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Terracina S., Petrella C., Francati S., Lucarelli M., Barbato C., Minni A., Ralli M., Greco A., Tarani L., Fiore M., et al. Antioxidant intervention to improve cognition in the aging brain: The example of hydroxytyrosol and resveratrol. Int. J. Mol. Sci. 2022;23:15674. doi: 10.3390/ijms232415674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Rufino-Palomares E.E., Perez-Jimenez A., Garcia-Salguero L., Mokhtari K., Reyes-Zurita F.J., Peragon-Sanchez J., Lupianez J.A. Nutraceutical role of polyphenols and triterpenes present in the extracts of fruits and leaves of Olea europaea as antioxidants, anti-infectives and anticancer agents on healthy growth. Molecules. 2022;27:2341. doi: 10.3390/molecules27072341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Zhang X., Cao J., Zhong L. Hydroxytyrosol inhibits pro-inflammatory cytokines, iNOS, and COX-2 expression in human monocytic cells. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 2009;379:581–586. doi: 10.1007/s00210-009-0399-7. [DOI] [PubMed] [Google Scholar]
- 118.Serra G., Deiana M., Spencer J.P.E., Corona G. Olive oil phenolics prevent oxysterol-induced proinflammatory cytokine secretion and reactive oxygen species production in human peripheral blood mononuclear cells, through modulation of p38 and JNK pathways. Mol. Nutr. Food Res. 2017;61:1700283. doi: 10.1002/mnfr.201700283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Yonezawa Y., Miyashita T., Nejishima H., Takeda Y., Imai K., Ogawa H. Anti-inflammatory effects of olive-derived hydroxytyrosol on lipopolysaccharide-induced inflammation in RAW264.7 cells. J. Vet. Med. Sci. 2018;80:1801–1807. doi: 10.1292/jvms.18-0250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Salminen A., Kaarniranta K., Kauppinen A. Insulin/IGF-1 signaling promotes immunosuppression via the STAT3 pathway: Impact on the aging process and age-related diseases. Inflamm. Res. 2021;70:1043–1061. doi: 10.1007/s00011-021-01498-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Yu Y.B., Zhuang H.Z., Ji X.J., Dong L., Duan M.L. Hydroxytyrosol suppresses LPS-induced intrahepatic inflammatory responses via inhibition of ERK signaling pathway activation in acute liver injury. Eur. Rev. Med. Pharmacol. Sci. 2020;24:6455–6462. doi: 10.26355/eurrev_202006_21544. [DOI] [PubMed] [Google Scholar]
- 122.Bigagli E., Cinci L., Paccosi S., Parenti A., D’Ambrosio M., Luceri C. Nutritionally relevant concentrations of resveratrol and hydroxytyrosol mitigate oxidative burst of human granulocytes and monocytes and the production of pro-inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. Int. Immunopharmacol. 2017;43:147–155. doi: 10.1016/j.intimp.2016.12.012. [DOI] [PubMed] [Google Scholar]
- 123.Rosignoli P., Fuccelli R., Fabiani R., Servili M., Morozzi G. Effect of olive oil phenols on the production of inflammatory mediators in freshly isolated human monocytes. J. Nutr. Biochem. 2013;24:1513–1519. doi: 10.1016/j.jnutbio.2012.12.011. [DOI] [PubMed] [Google Scholar]
- 124.Takeda Y., Bui V.N., Iwasaki K., Kobayashi T., Ogawa H., Imai K. Influence of olive-derived hydroxytyrosol on the toll-like receptor 4-dependent inflammatory response of mouse peritoneal macrophages. Biochem. Biophys. Res. Commun. 2014;446:1225–1230. doi: 10.1016/j.bbrc.2014.03.094. [DOI] [PubMed] [Google Scholar]
- 125.Gallardo-Fernández M., Hornedo-Ortega R., Alonso-Bellido I.M., Rodriguez-Gomez J.A., Troncoso A.M., Garcia-Parrilla M.C., Venero J.L., Espinosa-Oliva A.M., Pablos R.M. Hydroxytyrosol decreases LPS- and α-synuclein-induced microglial activation in vitro. Antioxidants. 2019;9:36. doi: 10.3390/antiox9010036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Zhang L., Zhang J., Jiang X., Yang L., Zhang Q., Wang B., Cui L., Wang X. Hydroxytyrosol inhibits LPS-induced neuroinflammatory responses via suppression of TLR-4-mediated NF-κB P65 activation and ERK aignaling pathway. Neuroscience. 2020;426:189–200. doi: 10.1016/j.neuroscience.2019.12.005. [DOI] [PubMed] [Google Scholar]
- 127.Yu H., Zhang Z., Wei F., Hou G., You Y., Wang X., Ca S., Yang X., Liu W., Zhang S., et al. Hydroxytyrosol ameliorates intervertebral disc degeneration and neuropathic pain by reducing oxidative stress and inflammation. Oxid. Med. Cell Longev. 2022;2022:2240894. doi: 10.1155/2022/2240894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Menicacci B., Cipriani C., Margheri F., Mocali A., Giovannelli L. Modulation of the senescence-associated inflammatory phenotype in human fibroblasts by olive phenols. Int. J. Mol. Sci. 2017;18:2275. doi: 10.3390/ijms18112275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Aparicio-Soto M., Redhu D., Sanchez-Hidalgo M., Fernandez-Bolanos J.G., Alarcon-de-la-Lastra C., Worm M., Babina M. Olive-oil-derived polyphenols effectively attenuate inflammatory responses of human keratinocytes by interfering with the NF-κB pathway. Mol. Nutr. Food Res. 2019;63:e1900019. doi: 10.1002/mnfr.201900019. [DOI] [PubMed] [Google Scholar]
- 130.Chen C., Chen L., Zhou J., Cai R., Ye Z., Zhang D. Anti-psoriasis activities of hydroxytyrosol on HaCaT cells under psoriatic inflammation in vitro. Immunopharmacol. Immunotoxicol. 2022;10:1–6. doi: 10.1080/08923973.2022.2143373. [DOI] [PubMed] [Google Scholar]
- 131.Santarelli R., Pompili C., Gilardini Montani M.S., Evangelista L., Gonnella R., Cirone M. 3,4-Dihydroxyphenylethanol (DPE or hydroxytyrosol) counteracts ERK1/2 and mTOR activation, pro-inflammatory cytokine release, autophagy and mitophagy reduction mediated by benzo[a]pyrene in primary human colonic epithelial cells. Pharmaceutics. 2022;14:663. doi: 10.3390/pharmaceutics14030663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Aparicio-Soto M., Sanchez-Hidalgo M., Cardeno A., Gonzalez-Benjumea A., Fernandez-Bolanos J.G., Alarcon-De-La-Lastra C. Dietary hydroxytyrosol and hydroxytyrosyl acetate supplementation prevent pristane-induced systemic lupus erythematous in mice. J. Funct. Foods. 2017;29:84–92. doi: 10.1016/j.jff.2016.12.001. [DOI] [Google Scholar]
- 133.Zhang X., Qin Y., Wan X., Liu H., Iv C., Ruan W., Lu L., He L., Guo X. Hydroxytyrosol plays antiatherosclerotic effects through regulating lipid metabolism via inhibiting the p38 signal pathway. BioMed Res. Int. 2020;2020:5036572. doi: 10.1155/2020/5036572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Pirozzi C., Lama A., Simeoli R., Paciello O., Pagano T.B., Mollica M.P., Di Guida F., Russo R., Magliocca S., Canani R.B., et al. Hydroxytyrosol prevents metabolic impairment reducing hepatic inflammation and restoring duodenal integrity in a rat model of NAFLD. J. Nutr. Biochem. 2016;30:108–115. doi: 10.1016/j.jnutbio.2015.12.004. [DOI] [PubMed] [Google Scholar]
- 135.Fuccelli R., Fabiani R., Rosignoli P. Hydroxytyrosol exerts anti-inflammatory and antioxidant activities in a mouse model of systemic inflammation. Molecules. 2018;23:3212. doi: 10.3390/molecules23123212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Lopez de las Hazas M.-C., Godinho-Pereira J., Macia A., Filipa Almeida A., Ventura M.R., Motilva M.-J., Santos C.N. Brain uptake of hydroxytyrosol and its main circulating metabolites: Protective potential in neuronal cells. J. Funct. Foods. 2018;46:110–117. doi: 10.1016/j.jff.2018.04.028. [DOI] [Google Scholar]
- 137.Wang Q., Wang C., Abdullah T.W., Qiu Z., Song M., Cao Y., Xiao J. Hydroxytyrosol alleviates dextran sulfate sodium-induced colitis by modulating inflammatory responses, intestinal barrier, and microbiome. J. Agric. Food Chem. 2022;70:2241–2252. doi: 10.1021/acs.jafc.1c07568. [DOI] [PubMed] [Google Scholar]
- 138.Miao F. Hydroxytyrosol alleviates dextran sodium sulfate-induced colitis by inhibiting NLRP3 inflammasome activation and modulating gut microbiota in vivo. Nutrition. 2022;97:111579. doi: 10.1016/j.nut.2021.111579. [DOI] [PubMed] [Google Scholar]
- 139.He C., Klionsky D.J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 2009;43:67–93. doi: 10.1146/annurev-genet-102808-114910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Singh R., Cuervo A.M. Autophagy in the cellular energetic balance. Cell Metab. 2011;13:495–504. doi: 10.1016/j.cmet.2011.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Aman Y., Schmauck-Medina T., Hansen M., Morimoto R.I., Simon A.K., Bjedov I., Palikaras K., Simonsen A., Johansen T., Tavernarakis N., et al. Autophagy in healthy aging and disease. Nat. Aging. 2021;1:634–650. doi: 10.1038/s43587-021-00098-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Klionsky D.J., Petroni G., Amaravadi R.K., Baehrecke E.H., Ballabio A., Boya P., Bravo-San Pedro J.M., Cadwell K., Cecconi F., Choi A.M.K., et al. Autophagy in major human diseases. EMBO J. 2021;40:e108863. doi: 10.15252/embj.2021108863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Dikic I., Elazar Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018;19:349–364. doi: 10.1038/s41580-018-0003-4. [DOI] [PubMed] [Google Scholar]
- 144.Kim J., Kundu M., Viollet B., Guan K.L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011;13:132–141. doi: 10.1038/ncb2152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Ge Y., Zhou M., Chen C., Wu X., Wang X. Role of AMPK mediated pathways in autophagy and aging. Biochimie. 2022;195:100–113. doi: 10.1016/j.biochi.2021.11.008. [DOI] [PubMed] [Google Scholar]
- 146.Wang S., Li H., Yuan M., Fan H., Cai Z. Role of AMPK in autophagy. Front. Physiol. 2022;13:1015500. doi: 10.3389/fphys.2022.1015500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Lee I.H. Mechanisms and disease implications of sirtuin-mediated autophagic regulation. Exp. Mol. Med. 2019;51:1–11. doi: 10.1038/s12276-019-0302-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Imai S., Armstrong C.M., Kaeberlein M., Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403:795–800. doi: 10.1038/35001622. [DOI] [PubMed] [Google Scholar]
- 149.Kawahara T.L., Michishita E., Adler A.S., Damian M., Berber E., Lin M., McCord R.A., Ongaigui K.C., Boxer L.D., Chang H.Y., et al. SIRT6 links histone H3 lysine 9 deacetylation to NF-kappa B-dependent gene expression and organismal life span. Cell. 2009;136:62–74. doi: 10.1016/j.cell.2008.10.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Sadria M., Layton A.T. Interactions among mTORC, AMPK and SIRT: A computational model for cell energy balance and metabolism. Cell Commun. Signal. 2021;19:57. doi: 10.1186/s12964-021-00706-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Wakui H., Yoshii Y., Yumino Y., Fujii S. Autophagy Induction by SIRT6 through attenuation of insulin-like growth factor signaling is involved in the regulation of human bronchial epithelial cell senescence. J. Immunol. 2014;192:958–968. doi: 10.4049/jimmunol.1302341. [DOI] [PubMed] [Google Scholar]
- 152.Klionsky D.J. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophahy. 2016;12:1–222. doi: 10.1080/15548627.2015.1100356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Lopez-Otin C., Kroemer G. Decelerating ageing and biological clocks by autophagy. Nat. Rev. Mol. Cell Biol. 2019;20:385–386. doi: 10.1038/s41580-019-0149-8. [DOI] [PubMed] [Google Scholar]
- 154.Deretic V. Multiple regulatory and effector roles of autophagy in immunity. Curr. Opin. Immunol. 2009;21:53–62. doi: 10.1016/j.coi.2009.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Germic N., Frangez Z., Yousefi S., Simon H.-U. Regulation of the innate immune system by autophagy: Monocytes, macrophages, dendritic cells and antigen presentation. Cell Death Differ. 2019;26:715–727. doi: 10.1038/s41418-019-0297-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Metur S.P., Klionsky D.J. Adaptive immunity at the crossroads of autophagy and metabolism. Cell. Mol. Immunol. 2021;18:1096–1105. doi: 10.1038/s41423-021-00662-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Deretic V., Levine B. Autophagy balances inflammation in innate immunity. Autophagy. 2018;14:243–251. doi: 10.1080/15548627.2017.1402992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Salminen A., Hyttinen J.M., Kaarniranta K. AMP-activated protein kinase inhibits NF-κB signaling and inflammation: Impact on healthspan and lifespan. J. Mol. Med. 2011;89:667–676. doi: 10.1007/s00109-011-0748-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Salminen A., Kaarniranta K., Kauppinen A. Inflammaging: Disturbed interplay between autophagy and inflammasome. Aging. 2022;4:166–175. doi: 10.18632/aging.100444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Lu R., Zhang L., Yang X. Interaction between autophagy and the NLRP3 inflammasome in Alzheimer’s and Parkinson’s disease. Front. Aging Neurosci. 2022;14:1018848. doi: 10.3389/fnagi.2022.1018848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Jeong J., Choi Y.J., Lee H.K. The role of autophagy in the function of CD4+ T cells and the development of chronic inflammatory diseases. Front. Pharmacol. 2022;13:860146. doi: 10.3389/fphar.2022.860146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Choi A.J.S., Ryter S.W. Autophagy in inflammatory diseases. Int. J. Cell Biol. 2011;2011:732798. doi: 10.1155/2011/732798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Pallauf K., Rimbach G. Autophagy, polyphenols and healthy ageing. Ageing Res. Rev. 2013;12:237–252. doi: 10.1016/j.arr.2012.03.008. [DOI] [PubMed] [Google Scholar]
- 164.Nabavi S.F., Sureda A., Dehpour A.R., Shirooie S., Silva A.S., Devi K.P., Ahmed T., Ishaq N., Hashim R., Sobarzo-Sanchez E., et al. Regulation of autophagy by polyphenols: Paving the road for treatment of neurodegeneration. Biotechnol. Adv. 2018;36:1768–1778. doi: 10.1016/j.biotechadv.2017.12.001. [DOI] [PubMed] [Google Scholar]
- 165.Leri M., Scuto M., Ontario M.L., Calabrese V., Calabrese E.J., Bucciantini M., Stefani M. Healthy effects of plant polyphenols: Molecular mechanisms. Int. J. Mol. Sci. 2020;21:1250. doi: 10.3390/ijms21041250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Tian Z., Zhang X., Sun M. Plant phytochemicals mediate autophagy against osteoarthritis by maintaining cartilage homeostasis. Front. Pharmacol. 2021;12:795058. doi: 10.3389/fphar.2021.795058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Brimson J.M., Prasanth M.I., Malar D.S., Thitilertdecha P., Kabra A., Tencomnao T., Prasansuklab A. Polyphenols for aging health: Implication from their autophagy modulating properties in age-associated diseases. Pharmaceuticals. 2021;14:982. doi: 10.3390/ph14100982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.García-Aguilar A., Palomino O., Benito M., Guillén C. Dietary polyphenols in metabolic and neurodegenerative diseases: Molecular targets in autophagy and biological effects. Antioxidants. 2021;10:142. doi: 10.3390/antiox10020142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Wu M., Luo Q., Nie R., Yang X., Tang Z., Chen H. Potential implications of polyphenols on aging considering oxidative stress, inflammation, autophagy, and gut microbiota. Crit. Rev. Food Sci. Nutr. 2021;61:2175–2193. doi: 10.1080/10408398.2020.1773390. [DOI] [PubMed] [Google Scholar]
- 170.Blanco-Benitez M., Calderon-Fernandez A., Canales-Cortes S., Alegre-Cortes E., Uribe-Carretero E., Paredes-Barquero M., Gimenez-Bejarano A., Duque Gonzalez G., Gomez-Suaga P., Ortega-Vidal J., et al. Biological effects of olive oil phenolic compounds on mitochondria. Mol. Cell Oncol. 2022;9:2044263. doi: 10.1080/23723556.2022.2044263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Xu W., Luo Y., Yin J., Huang M., Luo F. Targeting AMPK signaling by polyphenols: A novel strategy for tackling aging. Food Funct. 2023;14:56–73. doi: 10.1039/D2FO02688K. [DOI] [PubMed] [Google Scholar]
- 172.de Pablos R.M., Espinosa-Oliva A.M., Hornedo-Ortega R., Cano M., Arguelles S. Hydroxytyrosol protects from aging process via AMPK and autophagy; a review of its effects on cancer, metabolic syndrome, osteoporosis, immune-mediated and neurodegenerative diseases. Pharmacol. Res. 2019;143:58–72. doi: 10.1016/j.phrs.2019.03.005. [DOI] [PubMed] [Google Scholar]
- 173.Leri M., Bertolini A., Stefani M., Bucciantini M. EVOO polyphenols relieve synergistically autophagy dysregulation in a cellular model of Alzheimer’s disease. Int. J. Mol. Sci. 2021;22:7225. doi: 10.3390/ijms22137225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Zhi L.Q., Yao S.X., Liu H.L., Li M., Duan N., Ma J.B. Hydroxytyrosol inhibits the inflammatory response of osteoarthritis chondrocytes via SIRT6-mediated autophagy. Mol. Med. Rep. 2018;17:4035–4042. doi: 10.3892/mmr.2017.8353. [DOI] [PubMed] [Google Scholar]
- 175.Sun T., Chen Q., Zhu S.Y., Wu Q., Liao C.R., Wang Z., Wu X.H., Wu H.T., Chen J.T. Hydroxytyrosol promotes autophagy by regulating SIRT1 against advanced oxidation protein product induced NADPH oxidase and inflammatory response. Int. J. Mol. Med. 2019;44:1531–1540. doi: 10.3892/ijmm.2019.4300. [DOI] [PubMed] [Google Scholar]
- 176.Wang W., Jing T., Yang X., He Y., Wang B., Xiao Y., Shang C., Zhang J., Lin R. Hydroxytyrosol regulates the autophagy of vascular adventitial fibroblasts through the SIRT1-mediated signaling pathway. Can. J. Physiol. Pharmacol. 2018;96:88–96. doi: 10.1139/cjpp-2016-0676. [DOI] [PubMed] [Google Scholar]
- 177.Chen Q., Sun T., Wang J., Jia J., Yi Y.H., Chen Y.X., Miao Y., Hu Z.Q. Hydroxytyrosol prevents dermal papilla cells inflammation under oxidative stress by inducing autophagy. J. Biochem. Mol. Toxicol. 2019;33:e22377. doi: 10.1002/jbt.22377. [DOI] [PubMed] [Google Scholar]
- 178.Yang X., Jing T., Li Y., He Y., Zhang W., Wang B., Xiao Y., Wang W., Zhang J., Wei J., et al. Hydroxytyrosol attenuates LPS-induced acute lung injury in mice by regulating autophagy and sirtuin sxpression. Curr. Mol. Med. 2017;17:149–159. doi: 10.2174/1566524017666170421151940. [DOI] [PubMed] [Google Scholar]
- 179.Nardiello P., Pantano D., Lapucci A., Stefani M., Casamenti F. Diet supplementation with hydroxytyrosol ameliorates brain pathology and restores cognitive functions in a mouse model of amyloid-β deposition. J. Alzheimer Dis. 2018;63:1161–1172. doi: 10.3233/JAD-171124. [DOI] [PubMed] [Google Scholar]
- 180.Mosca A., Crudele A., Smeriglio A., Braghini M.R., Panera N., Comparcola D., Alterio A., Sartorelli M.R., Tozzi G., Raponi M., et al. Antioxidant activity of Hydroxytyrosol and Vitamin E reduces systemic inflammation in children with paediatric NAFDL. Dig. Liver Dis. 2021;53:1154–1158. doi: 10.1016/j.dld.2020.09.021. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
No data was used for the research described in the article.