

Abstract
Neoantigens generated by non-synonymous mutations of tumor genes can induce activation of neoantigen-reactive T (NRT) cells which have the ability to resist the growth of tumors expressing specific neoantigens. Immunotherapy based on NRT cells has made preeminent achievements in melanoma and other solid tumors. The process of manufacturing NRT cells includes identification of neoantigens, preparation of neoantigen expression vectors or peptides, induction and activation of NRT cells, and analysis of functions and phenotypes. Numerous improvement strategies have been proposed to enhance the potency of NRT cells by engineering TCR, promoting infiltration of T cells and overcoming immunosuppressive factors in the tumor microenvironment. In this review, we outline the improvement of the preparation and the function assessment of NRT cells, and discuss the current status of clinical trials related to NRT cell immunotherapy.
Keywords: Neoantigen-reactive T cell, Adoptive cell therapy, Immunotherapy, Cancer
Background
Recently, groundbreaking immunotherapies have revolutionized the schemes of cancer treatment. Conventional immunotherapies include immune checkpoint inhibitors (ICIs), adoptive cell therapy (ACT) and cancer vaccines, all of which improve the capability of immune system of recognizing and attacking cancer cells [1, 2]. However, due to the heterogeneity of tumors, Immunotherapy targeting single antigen may also result in the generation of target-irrelevant tumor cell clones and tumor immune escape, which has been reviewed in reference [3]. Therefore, it is urgent to develop multi-targeted immunotherapy. The term “neoantigen” means a new epitope of autoantigens generated by somatic non-synonymous mutations [4]. And cancer neoantigens will be generated by this kind of DNA mutations accumulated in tumor cells [5]. These antigens have tumor specificity and are absolutely absent in normal cells; they also possess the ability to stimulate autoimmune response and are not subject to central immune tolerance [6]. Targeting multiple neoantigens can be a significant measure to deal with the challenge of tumor immune escape. Since adoptive therapy with tumor infiltrating lymphocytes (TILs) emerged in the 1980s, neoantigens have been found as the major targets of TILs to exert specific antitumor function. Researches showed that these neoantigens can induce neoantigen-specific T cells, also called “neoantigen-reactive T cells” or “NRT” cells. Researches on NRT-based immunotherapies, including neoantigen vaccine and NRT cell adoptive therapy, have made remarkable achievements in melanoma and other solid tumors [7, 8]. The common point of these therapies is to recognize and kill neoplastic cells with autologous or heterologous NRT cells. However, Zhuting Hu et al. noted that neoantigen vaccine cannot induce adaptive immunity if not combined with appropriate adjuvants in the review of [6]. Even after activation, this vaccine still upregulates the immunosuppressive signaling of cancer, leading to the formation of suppressive tumor microenvironment (TME) [9]. What is more, weak immune induction is the most obvious defect of neoantigen vaccine in the treatment of advanced solid tumors. By contrast, a recent review has revealed that NRT cells can directly infiltrate into tumors after cultivation, and overcome the inhibition from TME by genetic modification of signal molecules [10]. For these reasons, developing NRT cell adoptive therapy can be a more effective method in treating solid tumors. This review focuses on the development history, preparation process, and preclinical as well as clinical researches of NRT cell therapy. It also explores the methods to enhance the anti-tumor effect of NRT cells.
Development history of NRT cell therapy
In the 1980s, De Plaen E. et al. first explored a neoantigen deriving from a single nucleotide variant which could be recognized by cytolytic T cells [11]. Subsequently, numerous cancer-related mutations that can be recognized by T cells were identified, including tumor associated antigens (TAAs), tumor specific antigens (TSAs), and cancer or testis antigens [12–16]. Among them, TSAs, especially neoantigens, are considered as the optimal tumor targets because they are never expressed in normal tissues and have a low probability of inducing tolerance. In a recent review, this group of antigens were divided into three types: Guarding neoantigens can induce antitumor immune response independently; restrained neoantigens have immune checkpoint-dependent immunogenicity; ignored neoantigens lack spontaneous immunogenicity [17]. The majority of neoantigens belong to the “ignored neoantigen” type, regarded as “the reserve of neoantigens”, which can be prepared as vaccine to induce autologous NRT cells [17]. In 2004, Rosenberg and his colleagues completed the first case of adoptive cell therapy, which showed that the tumor in metastatic lesions of patients with malignant melanoma regressed completely after adoptive transfer of TILs [18]. In another study, they also demonstrated that this therapy with two identified neoantigens can promote tumor infiltration of NRT cells and prolong their persistence [19]. The emergence of the next-generation sequencing technology has brought a new dawn for screening tumor neoantigens. This technique, combined with major histocompatibility complex (MHC) binding prediction approach based on silico algorithms, facilitates the selection of optimal missense genes and has become the mainstream method in neoantigen-identification [20, 21]. Patrick A. Ott et al. observed that neoantigen vaccine, another neoantigen-based immunotherapy, induces significant anti-tumor immune response in melanoma patients [22]. This therapy provides another reasonable, safe and durable anti-tumor method in a more individualized mode, but it has failed to achieve clinical benefits in a wider spectrum of cancer patients, which is addressed in the review of [23]. However, the combined treatment of neoantigen vaccines and immune checkpoint inhibitors at least partly provides a reference scheme for enhancing the clinical response of NRT cell treatment [24]. Currently, The focus of NRT cell therapy has shifted from melanoma to other solid tumors [25–29]. However, the efficacy of this therapy in solid tumors is limited, which is related to tumor immune escape, immune cell exhaustion or dysfunction, and immunosuppressive state of the tumor microenvironment (TME). Current NRT therapy mainly stems from improvement on TILs adoptive therapy. Neoantigen vaccines, adoptive transfer of NRT cells, TCR-engineered T cells and chimeric receptor T cell therapy have gradually emerged in clinical individualized antitumor treatment. Encouraging results from clinical studies highlight the importance of NRT cells in antitumor immunity. However, because few researches have studied adopting NRT cell therapy, very limited information of how to increase the efficacy of NRT cells adoptive therapy can be obtained from completed clinical trials hitherto. More endeavors are therefore needed to dissect the relationship between tumor immunity, neoantigens, and immune cells.
Introduction of neoantigen detecting platforms
Unlike overexpressed or abnormally expressed tumor-associated antigens, neoantigens are absent in the normal human genome [30]. The high-throughput sequencing technology and algorithmic prediction platforms render neoantigen identification more rapid and accurate (Fig. 1B, C) [31]. As for high-throughput sequencing, whole exome sequencing (WES) becomes the keystone of neoantigen identification [32]. Besides, mass spectrometry technology provides a large number of peptide data for training of MHC prediction platforms [33] (Fig. 1B). As for algorithmic prediction platforms, machine learning and artificial intelligence platforms (Fig. 2A) help to precisely predict potential MHC binding epitopes and MHC-peptide binding affinity based on sequencing outcomes. Some common prediction software, algorithms, and databases are listed in Table 1. Mutation screening is the first step in neoantigen identification. Mutation calling tools include Burrows-Wheeler Alignment tool (BWA), ANNOVAR, MuTect, SomaticSniper, VarScan, and FusionCatcher [34–38]. Differences between mutant sequence and wild sequences can be identified using the differential agretopic index (DAI) [39]. It is necessary to predict MHC binding ligands and binding affinity to determine whether mutations can form neoantigens. Published software represented by NetMHCpan, MHCflurry, HLAthena, MHCnuggets and ProGeo-Neo shows favorable outcomes in neoantigen prediction [40–44]. Immune Epitope Database (IEDB) is a primary epitope database and nealy all the prediction tools use the data from it [45]. The company of this database recently developed TCRMatch, which can identify T cell epitopes with unknown specificity based on optimized T cell epitope data of IEDB [46]. However, the data of IEDB are mostly from virus resources, which lead to the deviation of cancer neoantigen prediction. Novel database-Tumor Neoantigen Selection Alliance (TESLA)-based on tumor sequencing data will improve the precision of tumor neoantigen prediction [47]. The platforms widely applied in predicting MHC ligands are trained on neoepitope prediction with the data from literature or online databases. Each platform has the limitations of prediction objects and methods, while collaboration of multiple platforms will improve the specificity and accuracy. It is reasonable to prospect that these techniques will help to solve the difficulty in choosing the optimal neoantigen to activate antitumor NRT cells, which may be conducive to the efficacy improvement of immunotherapy.
Fig. 1.

Process of NRT cell manufacturing and adoptive therapy. NRT cells are manufactured via the following steps: A acquisition and cultivation of tumor specimens and peripheral blood mononuclear cells; B mutation identification with WES/WGS/RNA sequencing (seq), potential antigen detection with mass spectrometry; C neoantigen prediction; D design and synthesis of neoantigen-encoding mRNA in tandem minigene configuration or neoantigen peptides; E pulsing DCs directly with peptides, or transfection of neoantigen-encoding mRNA into DCs by electroporation, followed by the co-incubation of neoantigen-loaded DCs and PBMC-derived T cells, F flow cytometry-based neoantigen-specific T cell sorting; G rapid expansion protocol (REP) of NRT cells, H reinfusion of NRT cells into patients or mouse model
Fig. 2.

Feasible improvement for the manufacture of NRT cells. A Optimize neoantigen predicting platforms to promote the efficiency and accuracy of prediction. B Micro-electroporation (a), microinjection (b) and nano-delivery c can be applied to elevate transfection efficiency. Artificial APCs can be used to increase the efficiency of antigen presentation. C Promote NRT cell expansion in rapid expanding protocol (REP) through adding cytokines (IL-2, IL-7 and IL-15) (a) or anti-4-1BB antibody (b), using feeder cells (Bcl-xL, K562) (c), inhibiting AICD signaling cascade (d), or adopting the culture method of “young” T cells (e) or organoids (f). D Screen NRT cells with surface or genetic markers via single-cell transcriptome and TCR sequencing
Table 1.
Common platforms and algorithms for neoantigen prediction
| Tools Name | Function | Tool type | Ref |
|---|---|---|---|
| Burrows-Wheeler Alignment tool (BWA) | Alignment tool for mutation identification | software | [34] |
| ANNOVAR | Mutation identification and functional annotation | software | [35] |
| MuTect,SomaticSniper | Mutation calling and screening | software | [36, 37] |
| VarScan | Mutations and copy number alterations calling | software | [38] |
| FusionCatcher | Fusion gene mutation identification | software | http://code.google.com/p/fusioncatcher/ |
| NetMHCpan/NetMHCpanII, MHCflurry | MHC-I/II binding affinity prediction | software | [41, 48–50] |
| MixMHCpred score, HLAthena | MHC-I binding ligands prediction | algorithm | [42, 51] |
| MHCnuggets | MHC-I or MHC-II binding ligands prediction | software | [43] |
| ProGeo-Neo | Mutation calling, MHC-I and MHC-II binding affinity and binding ligands prediction | software | [44] |
| Differential agretopic index(DAI) | Difference identification between mutant and wild sequences | algorithm | [39] |
| NetCTLpan | T cell epitope and MHC-I binding ligands prediction | software | [52] |
| TCRMatch | T cell epitope prediction | software | [46] |
| PyClone | Clonal population prediction | algorithm | [53] |
| MuPeXI | Neoantigen Immunogenicity identification | software | [54] |
| Immune Epitope Database (IEDB) | Epitope data | database | [45] |
| Tumor Neoantigen Selection Alliance(TESLA) | Epitope and sequencing data | database | [47] |
Process of NRT cell induction
The major purpose of predicting neoantigens precisely is to induce the immune response of NRT cells, which is the critical element for antitumor immunotherapy. To induce NRT cells, the wide accepted protocols mainly include: specimen acquisition and isolation (Fig. 1A), identification of non-synonymous mutation through WES/WGS/RNA sequencing (seq), and detection of potential antigens with mass spectrometry (Fig. 1B), neoantigen prediction utilizing bioinformation technology (Fig. 1C), design and synthesis of neoantigen-encoding mRNA in tandem minigene configuration or neoantigen peptides (Fig. 1D), pulsing DCs directly with peptides, or transfection of neoantigen-encoding mRNA into DCs by electroporation, followed by the co-incubation of neoantigen-loaded DCs and PBMC-derived T cells (Fig. 1E), NRT cell functional assay and sorting through flow cytometry (Fig. 1F), rapid expansion protocol (REP) of NRT cells (Fig. 1G) before reinfusion into patients or mouse model (Fig. 1H) [55, 56]. Then, efficacy assessments of NRT cell adoptive therapy will be performed. In this part, we will introduce the improvement strategies of NRT cell induction specifically.
Ameliorated technologies, such as microinjection, micro-electroporation and nano-delivery, should be taken into account to elevate transfection efficiency (Fig. 2B a, b, c) [57, 58]. Because of the limited capacity of antigen presentation and long duration of the induction process when using autologous DCs, modified strategies of allogeneic APCs have been put forward (Fig. 2B d). Synthetic APCs, including magnetic and polymer compound beads covered with anti-CD3/CD28 antibody and HLA-Ig, also can be used to activate NRT cells [59, 60]. Nanoparticle-based artificial antigen presenting cell can mimic DCs to effectively prime and expand T cells. The following are some ways whereby this APC can be engineered: adding co-receptors, synthesizing nanoparticles coated with molecule-labeled DC membrane and T cell targeted antigens, modifying the shape of nanoparticles and endowing anti-phagocytosis ability [61–64]. Introduction of IL-2, and low-level IL-7 and IL-15 can be a time-saving method for NRT cell priming in expanding process, and these two cytokines promote the formation of the effector phenotype and the central memory phenotype while IL-15 additionally induces the stem cell memory phenotype of T cells (Fig. 2C a) [65, 66]. Cultivating TILs with agonistic CD137 (4-1BB) monoclonal antibodies increases frequency of CD8 + TILs, as well as amplification rate and quantity of T cell subclone types (Fig. 2C b) [67]. The “feeder cell”, including immortalized B cells and K562 cells, can be modified to express signals for T cell proliferation, which wins the favor of researchers in the REP process(Fig. 2C c) [68, 69]. In addition, inhibiting AICD signaling cascade and preventing the aging of T cells (“young” T cultivation method) in the process of REP will enhance the activity and prolong the persistence of adoptively transferred T cells (Fig. 2C d, e) [70, 71]. Another strategy is to cultivate NRT cells with tumor organoids in a personalized manner (Fig. 2C f) [72]. This patient-specific cell culture method develops a platform for better exploring the interaction between T cells, tumor cells and other immune cells from native environment. The study of NRT cells based on multi-omics analysis has confirmed its feasibility [73]. Generally, a more efficient inducing process of NRT cells has important implications for NRT cell therapy. The above-mentioned strategies seek to improve the efficiency of vector transduction, and promote the amplification and activation of T cells.
Detection of neoantigen-reactive T cell populations
Florian Kast et al. reviewed that using pMHC tetramer and dextramer binding assay based on flow cytometry can strengthen the binding forces between single pMHC and TCR, and elevate the efficiency of NRT cells screening (Fig. 1F) [4]. However, due to its low sensitivity, this technique cannot detect rare T cell clones containing NRT cells. Single-cell transcriptome and TCR sequencing play essential roles in the discovery of novel tumor-reactive T cell subclones, and further aid in the analysis and filteration of potential NRT cells within these subclones (Fig. 2E). Currently, the most common single-cell sequencing method of T cells mainly adopts microwell or microfluidic technology [74, 75]. Then, reverse transcription and PCR amplification are performed before transcriptomic and TCR sequencing. The sequencing data can be integrated and analyzed to identify T cell subclones and reconstruct TCR chains for pairing TCR with specific T cell clones [75, 76]. With the support of bioinformatic analysis, it is more convenient to find potential therapeutic targets and novel biomarkers with prognostic value, which facilitates efficacy evaluation of immunotherapies tailored to individuals. The feasibility of this novel detecting method has been demonstrated in several studies, and NRT cell populations have been successfully identified and isolated [77, 78]. The latest research also revealed that NeoTCR signatures can be used to identify specific antitumor NRT cells via single-cell transcriptome and single-cell TCR sequencing [79].
The value of screening signatures in NRT cell identification
Currently accepted surface markers of NRT cell include CTLA-4, PD-1, LAG-3, TIM-3 and TIGIT. 4-1BB/CD137 is transiently expressed on T cells, which is regarded as specific activating signature of NRT cells and extensively used in NRT cell population screening [80, 81]. High frequency of CD39 + tumor reactive T cells is relevant to better prognosis in cancer patients [82]. NRT cells of stem-like double negative (CD39- CD69-) phenotype show stronger antitumor activity and longer persistence despite the rarity of these cells [83]. In addition, other novel NRT cell surface markers have been identified, including CXCL13 and CD200 [84–86]. Particularly, the expression of CXCL13 is significantly different between NRT cells and bystander T cells. Common gene signatures include PDCD1, ENTPD1, LAG3, TIGHT, TNFRSF9, HAVCR2, BATF, GZMA/B/K, IFNA/B/G and CXCL13 gene [79, 84]. In order to elevate sensitivity and specificity of NRT cell screening, a combination of surface markers and transcriptome markers is recommended for identifying NRT cells. This approach not only facilitates the cell screening process, but also circumvents the influence of functional assays on the viability of the cells.
Feasible engineering strategies for NRT cells
The process of TCR recognizing and binding to MHC molecules is of great importance for T cells to perform antitumor function. The challenges are how to make T cells recognize tumor cells more effectively, how to enhance TCR function without increasing toxicity, and how to counteract the problems of T cell exhaustion as well as dysfunction. Therefore, we summarize some feasible engineering strategies for NRT cells to address these issues (Fig. 3A). Currently, the three most common engineering objects are TCR signals, co-stimulated signals and cytokines of T cells. Using transgenic TCR, co-expressing CD8 αβ with TCR and upregulating adhesion molecules can enhance MHC-TCR binding avidity and elevate signal-transducing efficiency [87–89]. Besides, engineering co-stimulatory signals is proposed to prolong T cell persistence and enhance anti-tumor activity, which can be achieved by coupling T cell activating signals (CD3ζ) with co-stimulating signals (CD28, OX40, 4-1BB), or using chimeric switch receptors which link exodomain of CTLA-4, PD-1 or TIGHT to intradomain of CD28 [90–95]. In addition, engineering cytokine receptors represented by orthogonal IL-2 has been found to enhance T cell antitumor function while attenuating the side effects caused by cytokine pleiotropy [96–98]. And T cells engineered to secrete additional cytokines (IL-7/12/15/18/23 and Flt3L) or chemokines (CXCL9/10/11 and CCL19/21) also show enhancement of activity and function [99–109]. These strategies improve the function of autologous tumor-reactive T cells, promote their phenotype switching, and, meanwhile, recruit other immune cells (such as NK cells and DCs) to exert antitumor effects. The abovementioned engineering strategies are used in CAR-T cells. CAR-T cells can also be designed to release enzymes, express multiple immunomodulators, deliver endogenous RNA, maximize the diversity of functions and minimize “off-target” toxicity, which are summarized as comprehensive strategies of armored CAR-T (Fig. 3B). These strategies have been reviewed in [110–113]. Engineering strategies of TCR T cells can also draw on this idea to extend persistence, promote homing and penetration into tumors, and enable T cells to target tumor cells and activate multiple immune cells simultaneously. Overall, the above researches highlight the necessity of T cell signaling in priming, proliferation and exerting function. Engineering strategies for receptors and cytokines expressed by T cells help to improve the persistence and antitumor function of T cells. We propose that these T cell modification methodologies can be also used to improve the antitumor activity as well as persistence of NRT cells, and promote infiltration of immune cells into solid tumors to limit their growth more effectively.
Fig. 3.

Feasible strategies for the improvement of T cell antitumor function. A Engineer T cell signals. Signal 1: edit TCR genes or make TCR and CD8 αβ co-expression. Signal 2: join CD28 to CD3ζ combined with 4-1BB or OX40 to enhance activation signals, construct the chimeric switch receptor (e. g., CD28 linked to PD-1,CTLA-4 and TIGHT) to reverse inhibitory signals. Signal 3: modify cytokine receptors (e. g., IL-2 orthogonal receptor) and increase the expression of autologous or heterologous cytokines or chemokines. B Produce multiple-function T cells: optimize CARs, secrete cytokines and enzymes, release extracellular vesicles containing RNAs, express multiple chemokine receptors, and modify immunosuppressive signal receptors. C Universal strategies to restore and increase the expression of MHC (inducing IFNγ production, using epigenetic silence or autophagy inhibitors (a)) and MICA/B (anti-MICA/B antibody (b))
Universal strategies for NRT cell therapeutic enhancement
MHC-TCR recognition pattern is the primary mechanism of NRT cell therapy. Under certain circumstances, however, tumor escape will occur when classical MHC molecules are downregulated or lose expression, which may be caused by gene reconstruction or mutation and deletion of functional components, loss of transcription factor, epigenetic silence, and pre-or post-transcriptional inhibitory regulation [114, 115]. Deficient expression of MHC also leads to dysfunction in the neoantigen presentation process [116]. This phenomenon will eventually affect the ability of T cells to recognize tumor cells. Since traditional TCR engineered T cells have MHC -restriction, modification of autologous T cells can be cumbersome and expensive. It is a tendency to use universal methods to enhance the antitumor function of T cells (Fig. 3C). These strategies can be used as auxiliary means to improve the efficacy of NRT cell therapy. The first strategy is to restore and increase expression of MHC molecules (Fig. 3C a). Previous researches have shown that IFN-γ can increase MHC expression [117]. Adopting epigenetic silence inhibitors, such as DNA methyltransferase inhibitors and histone deacetylase inhibitors, also has a pronounced effect on restoring or increasing MHC molecular expression [118–121]. Besides, reduction of MHC expression caused by autophagy can be another common tumor escape mechanism in a solid tumor. Autophagy inhibition recovers the MHC level of the tumor cell surface and promotes T cell activation [122]. NK cells acquire disinhibition when tumor cells decrease MHC molecular expression, and become activated when detecting ligands of activating receptors. Thus, exploiting the NK-involved antitumor mechanism can be a feasible and reasonable strategy to counteract tumor escape. A vaccine designed to induce antibodies that anchor MICA/B has been demonstrated to prevent tumor escape and enhance the function of tumor-reactive T cells and APCs (Fig. 3C b) [123]. Notably, this vaccine targeting MHC-I expressing tumors can be applied in clinical ACT as an inexpensive and effective “off-the-shelf” drug.
Influence of T cell infiltration and tumor microenvironment on the efficacy of immunotherapy
Although adoptive therapy of engineered T cells shows remarkable efficacy in clinical therapy, the interaction among malignant cells, immune cells, and other stromal components also requires deep exploration, which will offer feasible approaches to improving the infiltration and potency of T cells, preferably NRT. In some recent reviews, turning “cold tumors (immune-excluded and deserted tumors)” into “hot tumors (immune-inflamed tumors)” has become a research hot spot to strengthen immunotherapy efficacy [124, 125]. Compelling evidence proves that infiltration of antigen-specific T cells within tumors promotes favorable clinical outcomes. However, tumor-infiltrating T lymphocytes expressing high-level inhibitory receptors trigger downregulation of antitumor response, and low-level expression of chemokine receptors leads to poor T cell infiltration [126]. Strategies to improve T cell infiltration have been proposed to overcome these challenges (Fig. 4A). With radiotherapy and chemotherapy, tumor cells undergo immunogenic cell death (ICD), and release multiple cytokines and chemokines [127, 128]. Radiotherapy and thermal ablation can directly kill tumor cells or induce their apoptosis, as well as increase expression of MHC on the surface of the antigen presenting cells. [129, 130]. Compared with monotherapy, combination of NRT cell adoptive therapy or neoantigen vaccine with ICIs (Fig. 5) has been proven to achieve impressive outcomes [22, 24, 26]. Suppressive tumor microenvironment (TME) is composed of fibroblasts, immunosuppressive cells, abnormal proliferating vasculature and extracellular matrix, which may negatively impact host immune cell infiltration. Eliminating the physical barrier effect of extracellular matrix (ECM) by using ECM targeting agents can promote T cell infiltration into tumors (Fig. 4B), which has been addressed in the reviews of [131, 132]. The most common strategy for depleting the stroma is to use albumin-bound paclitaxel to facilitate T cell infiltration [133, 134]. Research showed that combinational therapy with nab-paclitaxel and gemcitabine or nab-paclitaxel and atezolizumab significantly improves tumor control and patient survival [135]. Aberrant growth of tumor vasculature will result in the formation of hypoxia and immunosuppressive TME [136]. Thus, normalizing the originally abnormal tumor vasculature via antiangiogenic agents (such as VEGFR inhibitor) and ICIs treatment will reduce hypoxia and remodel TME for a more favorable treating condition and potentiate antitumor immune cell activation (Fig. 4C) [136, 137]. Immune suppressive cells, such as Tregs, myeloid-derived suppressor cells (MDSCs) and tumor-associated macrophages (TAMs), are important study subjects in researches on overcoming resistance from TME. Depletion is the most common strategy for decreasing the quantity of both Tregs and MDSCs (Fig. 4D). Using anti-CCR4-antibody can deplete suppressive Treg cells in TILs and enhance antitumor-specific function of T cells [138]. For MDSCs, gemtuzumab can deplete intratumoral MDSCs, and the CXCR2 antagonist can block migration of MDSCs into tumors [139, 140]. Besides, targeting signaling pathway can also inhibit the proliferation and function of Tregs or MDSCs. Tregs can be inhibited by deleting transcription factor Blimp1 [141], while MDSCs can be inhibited by upregulating LXR expression [142], downregulating PEKR expression [143] or blocking CaMKK2 signal pathway [144]. In addition, another research hot spot is re-polarization of tumor-associated macrophages (TAM) from an anti-inflammatory (M2) phenotype to a pro-inflammatory (M1) one (Fig. 4E). This can be realized by using CpG-ODN [145] or non-coding RNAs as the immune regulator [146], or inhibiting the metabolism of lipid [147]. Tertiary lymphoid structures (TLSs) have a significant association with immune cell infiltration and cancer prognosis (Fig. 4F). In the two reviews of TLSs, the authors believed that solid tumors with more TLSs present a large quantity of effector memory T cells and cytotoxic T cells [148, 149]. Activated B cells in TLSs, except Bregs, can present antigens, stimulate activating signals, and secret cytokines to activate T cells and augment their antitumor function. Researches have demonstrated that the synergy work of B cells and T cells, together with the cooperation of humoral and cellular immunity, impacts the immune response and survival of patients [150, 151]. The number of TLSs is positively related to the efficacy of immunotherapy. And the induction of TLSs formation can be realized by applying ICIs [149, 152], vaccines [9], lymphoid chemokines or stromal cells [153, 154]. The abovementioned strategies strive to facilitate T cell infiltration, reinvigorate and augment the function of effector T cells, induce memory T cell generation, and eventually remodel adverse TME. These strategies can be performed by inhibiting immunosuppressive signals, increasing chemokine expression, removing barriers from TME, depleting or remodeling immunosuppressive cells, re-directing TAMs toward antitumor phenotype, and inducing formation of TLSs within the tumor. These measures dedicated to overcoming extracellular resistance can also improve the efficacy of adoptive NRT cell therapy in solid tumor treatment.
Fig. 4.

Strategies for promoting NRT cell tumor infiltration and modifying suppressive TME. A Improve tumor reactive T cell infiltration through inhibiting immunosuppressive signals, and promoting the expression of chemokines and the release of tumor specific antigens. B Eliminate the physical barrier effect of extracellular matrix(ECM) using drugs such as nab-paclitaxel. C Normalize abnormal vasculature by using VEGFR inhibitor. D Deplete immunosuppressive cells(Treg and MDSCs) and inhibit their activation signals. E Repolarize tumor-associated macrophages(TAM) from M2 towards M1. F Induce the formation of tertiary lymphoid structures(TLSs)(chemotherapy, ICIs, vaccine and stromal cell)
Fig. 5.
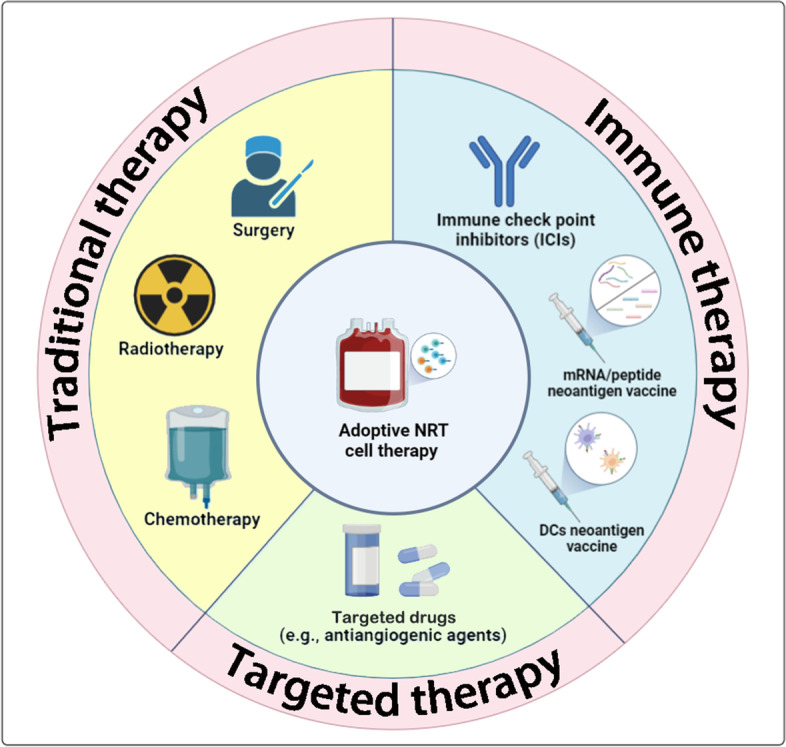
Feasible combinational therapy strategies of adoptive NRT cell therapy. Feasible combinational therapy strategies are shown in this figure. Adoptive NRT cell therapy can be combined with such strategies: immune therapy (immune check point inhibitors (ICIs), mRNA/peptide neoantigen vaccine, DC neoantigen vaccine), targeted drugs (e.g., antiangiogenic agents) and traditional therapy (surgery, radiotherapy and chemotherapy)
Clinical Trials for NRT cell therapy
We have summarized the clinical trials of NRT cell adoptive therapy and other NRT cell-related immunotherapies. Twenty-six eligible researches are incorporated, among which six are on NRT cell adoptive therapy, one on NRT cell adoptive therapy plus neoantigen vaccine (Neovax), one on neoantigen dendritic cell vaccine (Neo-DCVac) plus NRT cell adoptive therapy, seven on NeoVax, two on Neovax plus ICIs, three on tumor infiltrating lymphocytes (TILs), one on TIL plus ICIs, two on Neo-DCVac, one on Neo-DCVac plus ICIs and two on ICIs monotherapy. Almost all these researches are in phase I or II, and the majority are applied in melanoma due to its high mutant frequency. We have found that patients receiving NRT cell adoptive therapy or neoantigen vaccine therapy combined with ICIs outperform those who only receive monotherapy. In the following paragraphs we will mainly introduce clinical trials of NRT cell therapy. Other researches of NRT cell-related immunotherapy will be listed in Table 2. Schemes of feasible NRT cell combinational therapy are shown in Fig. 5.
Table 2.
Current NRT cell therapy and NRT cell relative clinical researches
| Identifier | Year | Cancer | Stage | Phase | Patients | Intervention | Name | Targets or formulation | Primary outcome | NRT cell induction or response | Inference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NCT01174121 | 2014 | cholangio-carcinoma | II | 1 | NRT | ERBB2IP-E805G CD4 + Th1 cell | SD | Vβ22 + ERBB2IP NRT cells exerted a major antitumor effect. | [155] | ||
| NCT03171220 | 2017 | solid tumors | advanced | 6 | NRT | CR: 1; PR: 1; SD: 4; PFS (median): 8.6 M | De novo approach: less than 30% of the peptides can induce NRT cells; Peptide library approach: nearly 50%. | [27] | |||
| NCT01174121 | 2017 | breast cancer | advanced (ER + HER2–) | II | 1 | NRT | mutSLC3A2, mutKIAA0368 | CR | SLC3A2 reactivity and KIAA0368 reactivity were mediated by CD4 + and CD8 + T cells, respectively. 11 TCR clonotypes recognized the four neoantigens (SLC3A2, KIAA0368, CADPS2 and CTSB). 72.7% of the TCR of NRT cells can be detected in patient’s peripheral blood, lasting at least 17 months. | [26] | |
| ChiCTR1800017836 | 2020 | collecting duct carcinoma | advanced | I | 1 | NRT + Neovax | 13 neoantigen peptides | SD | 12 of 13 peptides can induce NRT cells. The frequency of mutant allele decreased after three months. | [28] | |
| NCT03199807 | 2021 | HCC | IV | 1 | NRT | KRAS-G12A, KRAS-G13D, PIK3CA-H1047L, IDH1-R132H | CR | The proportion of NRT cells in vivo reached a maximum of 4.49% in four cycle reinfusions. | [25] | ||
| NCT03935893 | 2022 | solid tumor | advanced | II | 1 | NRT | KRAS-G12D TCR T cell | CR | Reinfused NRT cells represented approximately 2.4% of the total circulating T cells at 6 months. | [156] | |
| NCT01174121 and NCT03412877 | 2022 | solid tumor | advanced | II | 13 | NRT | TP53 TCR T cell | PR: 3; NR: 10 |
Percentage of NRT cells with a persistence of 6 weeks: 0.01%; NRT cells showed high level exhausted phenotype (PD-1:43%, TIM3: 33%, CD39: 93%) and low-level central memory phenotype (CD62L:5.02%). |
[157] | |
| NCT03067493 | 2022 | primary HCC | II | 23(10 vaccinated) | NRT + Neo-DCVac | peptide–loaded DC | SD: 5; PD: 5; DSF(median): 18.3 M | 42.5% of the neoantigen peptides induced NRT cell response; 70% of patients had neoantigen-specific immune response. | [158] | ||
| NCT02035956 | 2017 | melanoma | III-IV | I | 13 | NeoVax | IVAC MUTANOME | mRNA | SD: 8; PD: 5 | 60% of the neoantigen peptides induced NRT cell response; each patient developed NRT cells against at least 3 mutations; 57% of NRT cells were CD4 + T cell. | [159] |
| NCT02287428 | 2018 | MGMT-UG | I/Ib | I | 10 (2 withdrew) | NeoVax | peptides + poly-ICLC | PD: 8; PFS(median): 7.6 M; OR: 16.8 M | Two patients who did not receive dexamethasome generated NRT cell response towards neoantigens. | [160] | |
| NCT03480152 | 2019 | gastrointestinal cancer | advanced | I | 5(1 PD) | NeoVax | mRNA-4650 | mRNA | NR | 15.7% of the neoantigen peptides induced NRT cell response; 59% of NRT cells were CD4 + T cells and 41% were CD8 + T cells. | [161] |
| NCT03662815 | 2019 | solid tumors | advanced | 24 (22 vaccinated, 21 finish five) | NeoVax | iNeo-Vac-P01 | peptides + GM-CSF | SD: 15; PD: 6; PFS(median): 4.6 M; PFS(6): 27.3%; OS(12): 55.1%; | Nearly 80% of the peptides or peptide pools induced NRT cell response. | [162] | |
| NCT01461148 | 2020 | colorectal cancer | III/IV | I/IIa | 22(19 finished) | NeoVax | FSP (TAF1B (-1), HT001 (-1) and AIM2 (-1) | peptides | SD: 3 | 50% of the patients had neoantigen-specific immune response. Each patient developed NRT cells against at least 1 peptide. Some frameshift peptide neoantigens could not induce CD8 + NRT cells. | [163] |
| NCT02960230 | 2020 | diffuse midline glioma | HLA-A*02:01 + , H3.3K27 Mut | 19 | NeoVax | peptides + TT peptide + poly-ICLC | OS(median): 16.1 M, OS(12): 40% | Nearly 80% of the neoantigen peptides induced NRT cell response. Expansion of CD8 + NRT cells was associated with a better prognosis. | [164] | ||
| ChiCTR1900020990 | 2021 | HCC | II-III | I | 10 | NeoVax | peptides + poly-ICLC | CR: 2; PD: 8; median RFS: 7.4 M; | Nearly 70% of the neoantigen peptides induced NRT cell response. 50% of the patients had neoantigen-specific immune response. | [165] | |
| NCT01970358 | 2017 | melanoma | IIIB/C, IVM1a/b | I | 8 (6 vaccinated) | NeoVax + ICIs | ICI: pembrolizumab | peptides + poly-ICLC | CR: 2; SD: 6; PD: 2; PFS: 25 M; | 47% of the neoantigen peptide pools induced NRT cell response. 20% of the neoantigen peptides induced CD4 + T cell response. The proportion of neoantigen stimulation was higher in MHC class II than in MHC class I. | [22] |
| NCT02897765 | 2020 | melanoma, NSCLC, and TCC | III/IV | 82(60 vaccinated) | NeoVax + ICIs | NeoVax: Neo-PV-01,ICI: Nivolumab | peptides + poly-ICLC | PFS (median): 23.5 M (melanoma), 8.5 M (NSCLC); 5.8 M (TCC). OS(1Y): 96% (melanoma), 83% (NSCLC), 67% (TCC) | 52% of the neoantigen peptides induced NRT cell response in melanoma patients, 47% in NSCLC and 52% in bladder cancer patients. The average proportion of the immune response induced by neoantigen were 42% in CD4 + T cells and 24% in CD8 + T cells. | [24] | |
| NCT01807182 | 2018 | melanoma | III-IV | II | 1 | TIL | BRAFV600E | CR | Neoantigen peptides only stimulated CD4 + T cell response. | [166] | |
| NCT02278887 | 2020 | melanoma | IIIc/IV | I | 10 | TIL | CR: 2; PR: 3; SD: 1; PD: 4 | NRT cells can be detected in 66.7% of the patients with TIL infusion; The frequency of NRT cells responses peaked between 3 and 9 months. | [167] | ||
| NCT00937625 | 2021 | melanoma | advanced | I | 26 | TIL | CR: 5; PR: 6; SD: 10; PD: 5 | NRT cells can be detected in 69.2% of the patients with TIL infusion; 3.4% of the neoantigen peptides in TIL or peripheral blood induced NRT cell response. The median proportion of CD8 + T NRT cells was 0.63%. | [168] | ||
| NCT03215810 | 2020 | NSCLC | IV | I | 20(16 vaccinated) | TIL + ICIs | ICI: nivolumab | CR: 2; PR: 2; SD: 2; PD: 1 |
72.2% of the patients had neoantigen-specific immune response. The majority of TILs were terminally differentiated and only small subsets were in the stem-like state. |
[169] | |
| NCT00683670 | 2013 | melanoma | IV | I | 7 | Neo-DCVac | CR: 1; PR: 2; PD: 4 | DC vaccine augmented NRT cell response and broadened neoantigen-reactive TCR repertoire. Only three neoantigens with the highest binding affinity can induce NRT cell response in each patient. | [170, 171] | ||
| NCT02956551 | 2020 | lung cancer | IIIc/IV | I | 18(12 vaccinated) | Neo-DCVac | SD: 9; PD: 3; PFS(median): 5.5 M; OS(median):7.9 M | Nearly 62% of the neoantigen peptides induced NRT cell response on average. The percentage of CD8 + NRT cells was 21.69% and that of CD4 + NRT cells was 43.72% on average. | [172] | ||
| NCT01132014 | 2017 | ovarian cancer | IIIb-IV | I | 25 | Neo-DCVac + ICIs | ICI: bevacizumab | DC pulsed with oxidized autologous whole-tumor cell lysate | SD: 16; PD: 9; PFS(24 months): 25; OS(2Y, responder): 100%, OS(2Y,no responder): 25%; | Six patients who received DC vaccine generated NRT cells targeting at least one neoantigen. CD8 + NRT cells induced by DC vaccine were polyfunctional. | [173] |
| NCT02108652 | 2014 | UBC | IV | II | 24 | ICI | Atezolizumab | Early-stage NART expansion and activation are associated with response to ICB. | Increased NRT cell response can be observed between pre-treatment to three weeks post-treatment of ICI. The phenotypes of NRT cells with PD1 + Ki67 + effector and increased CD39 levels showed positively relevant to clinical response. | [174] | |
| NCT01903993 | 2019 | NSCLC | IIIb-IV | II | 14 | ICI | Atezolizumab | Neoantigen-specific T cells phenotype change: differentiated effector phenotype, memory-like phenotypic (PD). | Patients who responded to ICI therapy showed increased frequency of NRT cells and differentiated effector phenotype of T cells. | [175] |
UBC Urothelial bladder cancer, MGMT-UG MGMT-unmethylated glioblastoma, NSCLC Non-small cell lung cancer, TCC Urothelial carcinoma, HCC Hepatocellular carcinoma, NRT Neoantigen reactive T cell, NeoVax Neoantigen vaccine, Neo-DCVac Neoantigen dendritic cells vaccine, ICI Immune checkpoint inhibitor, TIL tumor infiltrating lymphocyte, FSP Frameshift peptide, GM-CSF Granulocyte–macrophage colony stimulating factor, CR complete response, PR partial response, SD Stable disease, PD Progressive disease, NR No response, OS Overall survival, PFS Progression-free survival, DFS Disease-free survival, RFS Relapse free progression, Y year (s), M Month (s))
The majority of traditional engineered TCR T cells are designed to target tumor-associated antigens (TAAs) while relatively few teams have conducted researches on neoantigens [89, 176]. It is difficult for these T cells to eliminate tumor cells thoroughly due to their heterogeneity, and patients also show limited clinical responses or even suffer autoimmune diseases caused by the “off-target” effect [177–180]. Rosenberg’s team is devoted to exploring NRT cell populations targeting shared antigens and developing engineered neoantigen-targeted TCR T cells as “off-the-shelf” products. Their first NRT cell therapy case was a metastatic cholangiocarcinoma patient who received ERBB2IP-targeted CD4 + T cell therapy and achieved disease stability for more than one year after twice reinfusions [155]. KRAS-G12D-targeted NRT cells in gastrointestinal cancers have also been screened [181, 182]. Recently, a case report showed that a pancreatic cancer patient who received KRAS-G12D NRT cell therapy obtained a 6-month partial objective response accompanied by long-term existence of effector T cells [156]. Besides, twelve patients in two clinical trials who harbored TP53 mutation received NRT cell therapy. Among them, two patients exhibited a partial response, and another patient with chemo-refractory breast cancer realized tumor regression that lasted for at least six months [157]. Another research team conducted two pioneering clinical trials, showing the value of transferring NRT cells in treating advanced and refractory solid tumors. In the first study, researchers compared the therapeutic effects of two sources of neoantigens-de novo and shared library. In three patients treated with NRT cells manufactured by the de novo pattern, the overall response rate of T cells to neoantigens was lower than 34%. However, using a shared neoantigen library could significantly increase the efficiency and accuracy of hot spot mutation identification. Six patients using NRT cells made by this pattern achieved one CR, one PR, and four SD [27]. In the second study, a patient with advanced hepatocellular carcinoma (HCC) received NRT cell therapy combined with radiotherapy and ICI therapy, and realized partial response and complete regression in the new lesion [25]. This study completed the first comprehensive NRT cell therapy in an advanced HCC patient who benefited from prolonged survival without severe side effects. In addition, other studies have also shown favorable clinical results of NRT cell therapy. Nikolaos Zacharakis et al. presented a case of a breast cancer patient with complete regression after reinfusion of NRT cells targeting four individual somatic mutations combined with ICIs [26]. Our team also reported a case of a collecting duct carcinoma (CDC) patient who obtained SD with decreased tumor loads and regression of metastatic lesions after administration of NRT immunotherapy [28]. More than 92% of the neoantigens in this research could fully stimulate reactive T cells in PBMC. The activation proportion of NRT elevated from 1.92% to 7.92%. The latest phase II clinical trial used NRT cell therapy combined with DC neoantigen vaccine, chemotherapy, radiofrequency ablation, and ICIs to treat hepatocellular carcinoma [158]. Fifty percent of the patients obtained disease stability without relapse for two years. Other patients failed to respond due to depletion of tumor neoantigen and generation of new neoantigen epitope. The overall safety of adoptive NRT cell therapy is good and no prominently serious adverse events have been observed. Only two among the seven studies of NRT cell therapy reported minor adverse reactions of grade 1–2 [27, 157]. These results demonstrate that this therapy is feasible and safe for the activation of autologous NRT cells to eliminate tumor cells.
Furthermore, some recruiting studies on NRT cell therapy and engineering TCR neoantigen T cells are listed in Table 3. All these researches aim at solid tumors, including three in phase I, six in phase I/II, and two in phase II clinical trials. Four studies use shared NRT cells, while two use de novo NRT cells. Six studies apply NRT cells combined with ICIs, one of which also adds CDX-1140, a monoclonal antibody targeting CD40. The feasibility and safety of these researches need to be confirmed by the publication of the latest results.
Table 3.
Recruiting researches of NRT cell and engineering TCR T cell therapy
| Identifier | Posted Year | Cancer | Intervention | Target | Phase | Combination | Country | Status |
|---|---|---|---|---|---|---|---|---|
| NCT03190941 | 2017 | gastrointestinal and pancreatic cancer | engineering TCR-T cell | HLA-A*11:01, KRAS G12V | I/II | none | United States | recruiting |
| NCT03745326 | 2018 | gastrointestinal and pancreatic cancer | engineering TCR-T cell | HLA-A*02:01,KRAS-G12D | I/II | none | United States | recruiting |
| NCT03412877 | 2018 | solid tumor | engineering TCR-T cell | unknown | II | Pembrolizumab | United States | recruiting |
| NCT04102436 | 2019 | solid tumor | engineering TCR-T cell | unknown | II | none | recruiting | |
| NCT03970382 | 2019 | solid tumor | engineering TCR-T cell | unknown | I | Nivolumab | United States | Suspended |
| NCT04032847 | 2019 | NSCLC | NRT cells(ATL001) | mutiple neoantigens | I/II | Pembrolizumab | United Kingdom | recruiting |
| NCT03997474 | 2019 | melanoma | NRT cells(ATL001) | mutiple neoantigens | I/II | Nivolumab | United Kingdom | recruiting |
| NCT04146298 | 2019 | pancreatic cancer | NRT cells | HLA-A*11:01,KRAS G12V | I/II | Anti-PD-1 monoclonal antibody | China | recruiting |
| NCT04520711 | 2020 | malignant epithelial cancer | engineering TCR-T cell | unknown | I | CDX-1140 + Pembrolizumab | United States | recruiting |
| NCT05194735 | 2022 | solid tumor | engineering TCR-T cell | unknown | I/II | none | United States | recruiting |
| NCT05478837 | 2022 | diffuse midline glioma | engineering TCR-T cell | HLA-A*0201, H3.3K27M | I | none | United States | not yet recruiting |
The studies above show that NRT cell therapy realizes favorable tumor regression and long-term antitumor effect, especially for patients of end-stage melanoma or refractory solid tumors, in a more individualized or “off-the-shelf” way. However, due to the inaccuracy of prediction algorithms or suppression of TME, the overall response to NRT cell therapy is limited. In some cases, shared antigens are not included in the top alternative neoantigens, which means driver gene peptides are not the optimal targets in some cancer patients. The efficacy of adoptive NRT cell therapy will be improved by both traditional therapy and other immunotherapies, which can broaden the repertoire and augment the function of autologous NRT cells.
Limitations of NRT cell therapy
Although adoptive NRT cell therapy has superiority in effectiveness and safety in advanced tumor treatment, it still has some limitations. Under the pressure of immune editing, the consequence of the tumor cell evolution is that the quantity of cancer neoantigens originating from driver mutation will decrease while the number of those deriving from passenger mutation will increase. Thus, NRT cells designed to target single driver gene mutations (e.g., KRAS, TP53) fail to achieve complete regression of primary tumors. Moreover, unlike driver mutation-derived neoantigens, passenger mutation-derived neoantigens are different in each patient, suggesting that the cell products for each patient need to be tailored. Besides, the deficiency of predicting platforms leads to the deviation of neoantigen prediction and dissatisfactory treatment efficacy. Compared with the neoantigen vaccine, adoptive NRT cell therapy targets fewer neoantigens and induces a narrower breadth of the immune response [5, 183], and the process of NRT cell manufacturing is complicated, time-consuming and costly. Existent evidence has shown that ex vivo cultivation will increase the proportion of the terminal differential phenotype of T cells and reduce activity and proliferation of NRT cells [184–186], whereas neoantigen vaccine will not result in these problems due to it induces NRT cells activation directly in vivo. Furthermore, potential cytokine release syndrome requires additional attention in NRT cell-based therapy, and IL-1 and IL-6 receptor antagonists or blockades are needed to cope with this problem [187, 188].
Conclusion
The discovery of neoantigens boosts the development of individually-tailored immunotherapy, including adoptive therapy with T cells. With more efficient and precise therapeutic potency, T cells stimulated by neoantigens exhibit powerful antitumor capability. Based on bio-information technology, T cell screening and engineering techniques, modified NRT cells can be implemented more economically and conveniently. Although the feasibility and safety of NRT-related immunotherapy have been verified, the majority of researches are still in the initial stage, and the overall treatment results are unsatisfactory. Improved methods have been proposed to meet the urgent demand for improvement of therapy effectiveness and development of novel platforms as well as of multiple-drug combinatorial therapy. Current challenges to adoptive NRT cell therapy are the high cost and difficulty in realizing individualization, which renders industrial mass production unlikely and needs to be solved by future technological innovation. However, generally speaking, we are convinced that NRT cell-based immunotherapy has the effectiveness and safety to realize enduring tumor elimination and significantly prolonged survival that benefit patients with advanced solid tumors.
Acknowledgements
Not applicable.
Abbreviation
- ACT
Adoptive cell therapy
- AICD
Activation-induced T cell death
- APC
Antigen presenting cell
- CAR-T
Chimeric antigen receptor-T cell
- CDC
Collecting duct carcinoma
- CR
Complete response
- DC
Dendritic cell
- ECM
Extracellular matrix
- HCC
Hepatocellular carcinoma
- ICD
Immunogenic cell death
- ICI
Immune checkpoint inhibitor
- MDSC
Myeloid-derived suppressor cell
- MHC
Major histocompatibility complex
- MICA/B
MHC class I polypeptide–related sequence A/B
- Neo-DCVac
Neoantigen dendritic cell vaccine
- NeoVax
Neoantigen vaccine
- NRT cells
Neoantigen-reactive T cells
- PBMC
Peripheral blood mononuclear cell
- PR
Partial response
- REP
Rapid expansion protocol
- SD
Stable disease
- TAA
Tumor associated antigen
- TAM
Tumor-associated macrophages
- TCR
T cell receptor
- TIL
Tumor infiltrating lymphocyte
- TLS
Tertiary lymphoid structure
- TME
Tumor microenvironment
- TSA
Tumor specific antigen
- VEGFR
Vascular endothelial growth factor receptor
- WES
Whole exome sequencing
Authors’ contributions
RH and BZ contributed equally to this review. RH, BZ, XS and WZ conceived the framework and content of this manuscript. RH and BZ collected the published literature and researches data and drafted the manuscript. SH, QZ, XS and WZ provided critical comments and edited the manuscript. All the authors read and approved the final manuscript.
Funding
The Basic Medical Research Project of the First Affiliated Hospital of Second Military Medical University (Grant No. 2021JCMS16), The San Hang Program of the Second Military Medical University, The Project of Disciplines of Excellence, Shanghai Municipal Health Commission (Grant No. 20224Z0037).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Ruichen Huang and Bi Zhao contributed equally to this work.
Contributor Information
Xiaoping Su, Email: bloooge@163.com.
Wei Zhang, Email: zhangweismmu@126.com.
References
- 1.Kennedy LB, Salama AKS. A review of cancer immunotherapy toxicity. CA Cancer J Clin. 2020;70(2):86–104. doi: 10.3322/caac.21596. [DOI] [PubMed] [Google Scholar]
- 2.Rosenberg SA, Restifo NP. Adoptive cell transfer as personalized immunotherapy for human cancer.pdf. Science. 2015;348(6230):62–68. doi: 10.1126/science.aaa4967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.McGranahan N, Swanton C. Clonal heterogeneity and tumor evolution: past, present, and the future. Cell. 2017;168(4):613–628. doi: 10.1016/j.cell.2017.01.018. [DOI] [PubMed] [Google Scholar]
- 4.Kast FKC, Umaña P, Gros A, Gasser S. Advances in identification and selection of personalized neoantigen/T-cell pairs for autologous adoptive T cell therapies. Oncoimmunology. 2021;10(1):1869389. doi: 10.1080/2162402X.2020.1869389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Schumacher TN, Scheper W, Kvistborg P. Cancer neoantigens. Annu Rev Immunol. 2019;37:173–200. doi: 10.1146/annurev-immunol-042617-053402. [DOI] [PubMed] [Google Scholar]
- 6.Hu Z, Ott PA, Wu CJ. Towards personalized, tumour-specific, therapeutic vaccines for cancer. Nat Rev Immunol. 2018;18(3):168–182. doi: 10.1038/nri.2017.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Leko V, Rosenberg SA. Identifying and targeting human tumor antigens for T cell-based immunotherapy of solid tumors. Cancer Cell. 2020;38(4):454–472. doi: 10.1016/j.ccell.2020.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Davis L, Tarduno A, Lu YC. Neoantigen-reactive T cells: the driving force behind successful melanoma immunotherapy. Cancers (Basel) 2021;13(23):6061. doi: 10.3390/cancers13236061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lutz ER, Wu AA, Bigelow E, Sharma R, Mo G, Soares K, Solt S, Dorman A, Wamwea A, Yager A, et al. Immunotherapy converts nonimmunogenic pancreatic tumors into immunogenic foci of immune regulation. Cancer Immunol Res. 2014;2(7):616–631. doi: 10.1158/2326-6066.CIR-14-0027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Restifo NP, Dudley ME, Rosenberg SA. Adoptive immunotherapy for cancer: harnessing the T cell response. Nat Rev Immunol. 2012;12(4):269–281. doi: 10.1038/nri3191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.De Plaen E, Lurquin C, Van Pel A, Mariamé B, Szikora JP, Wölfel T, Sibille C, Chomez P, Boon T. Immunogenic (tum-) variants of mouse tumor P815: cloning of the gene of tum-antigen P91A and identification of the tum- mutation. Proc Natl Acad Sci U S A. 1988;85(7):2274–2278. doi: 10.1073/pnas.85.7.2274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Brichard V, Van Pel A, Wölfel T, Wölfel C, De Plaen E, Lethé B, Coulie P, Boon T. The tyrosinase gene codes for an antigen recognized by autologous cytolytic T lymphocytes on HLA-A2 melanomas. J Exp Med. 1993;178(2):489–495. doi: 10.1084/jem.178.2.489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gaugler B, Van den Eynde B, van der Bruggen P, Romero P, Gaforio JJ, De Plaen E, Lethé B, Brasseur F, Boon T. Human gene MAGE-3 codes for an antigen recognized on a melanoma by autologous cytolytic T lymphocytes. J Exp Med. 1994;179(3):921–930. doi: 10.1084/jem.179.3.921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Boël P, Wildmann C, Sensi ML, Brasseur R, Renauld JC, Coulie P, Boon T, van der Bruggen P. BAGE: a new gene encoding an antigen recognized on human melanomas by cytolytic T lymphocytes. Immunity. 1995;2(2):167–175. doi: 10.1016/S1074-7613(95)80053-0. [DOI] [PubMed] [Google Scholar]
- 15.Wang RF, Robbins PF, Kawakami Y, Kang XQ, Rosenberg SA. Identification of a gene encoding a melanoma tumor antigen recognized by HLA-A31-restricted tumor-infiltrating lymphocytes. J Exp Med. 1995;181(3):1261. doi: 10.1084/jem.181.2.799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kawakami Y, Wang X, Shofuda T, Sumimoto H, Tupesis J, Fitzgerald E, Rosenberg S. Isolation of a new melanoma antigen, MART-2, containing a mutated epitope recognized by autologous tumor-infiltrating T lymphocytes. J Immunol. 2001;166(4):2871–2877. doi: 10.4049/jimmunol.166.4.2871. [DOI] [PubMed] [Google Scholar]
- 17.Lang F, Schrors B, Lower M, Tureci O, Sahin U. Identification of neoantigens for individualized therapeutic cancer vaccines. Nat Rev Drug Discov. 2022;21(4):261–282. doi: 10.1038/s41573-021-00387-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Huang J, El-Gamil M, Dudley ME, Li YF, Rosenberg SA, Robbins PF. T cells associated with tumor regression recognize frameshifted products of the CDKN2A tumor suppressor gene locus and a mutated HLA class I gene product. J Immunol. 2004;172(10):6057–6064. doi: 10.4049/jimmunol.172.10.6057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhou J, Dudley ME, Rosenberg SA, Robbins PF. Persistence of multiple tumor-specific T-cell clones is associated with complete tumor regression in a melanoma patient receiving adoptive cell transfer therapy. J Immunother. 2005;28(1):53–62. doi: 10.1097/00002371-200501000-00007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hoof I, Peters B, Sidney J, Pedersen LE, Sette A, Lund O, Buus S, Nielsen M. NetMHCpan, a method for MHC class I binding prediction beyond humans. Immunogenetics. 2009;61(1):1–13. doi: 10.1007/s00251-008-0341-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Stranzl T, Larsen MV, Lundegaard C, Nielsen M. NetCTLpan: pan-specific MHC class I pathway epitope predictions. Immunogenetics. 2010;62(6):357–368. doi: 10.1007/s00251-010-0441-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ott PA, Hu Z, Keskin DB, Shukla SA, Sun J, Bozym DJ, Zhang W, Luoma A, Giobbie-Hurder A, Peter L, et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature. 2017;547(7662):217–221. doi: 10.1038/nature22991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yang JC, Rosenberg SA. Adoptive T-cell therapy for cancer. Adv Immunol. 2016;130:279–294. doi: 10.1016/bs.ai.2015.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ott PA, Hu-Lieskovan S, Chmielowski B, Govindan R, Naing A, Bhardwaj N, Margolin K, Awad MM, Hellmann MD, Lin JJ, et al. A phase Ib trial of personalized neoantigen therapy plus anti-PD-1 in patients with advanced melanoma, non-small cell lung cancer, or bladder cancer. Cell. 2020;183(2):347–362.e24. doi: 10.1016/j.cell.2020.08.053. [DOI] [PubMed] [Google Scholar]
- 25.Liu C, Shao J, Dong Y, Xu Q, Zou Z, Chen F, Yan J, Liu J, Li S, Liu B, et al. Advanced HCC patient benefit from neoantigen reactive T cells based immunotherapy: a case report. Front Immunol. 2021;12:685126. doi: 10.3389/fimmu.2021.685126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zacharakis N, Chinnasamy H, Black M, Xu H, Lu YC, Zheng Z, Pasetto A, Langhan M, Shelton T, Prickett T, et al. Immune recognition of somatic mutations leading to complete durable regression in metastatic breast cancer. Nat Med. 2018;24(6):724–730. doi: 10.1038/s41591-018-0040-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chen F, Zou Z, Du J, Su S, Shao J, Meng F, Yang J, Xu Q, Ding N, Yang Y, et al. Neoantigen identification strategies enable personalized immunotherapy in refractory solid tumors. J Clin Invest. 2019;129(5):2056–2070. doi: 10.1172/JCI99538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zeng Y, Zhang W, Li Z, Zheng Y, Wang Y, Chen G, Qiu L, Ke K, Su X, Cai Z, et al. Personalized neoantigen-based immunotherapy for advanced collecting duct carcinoma: case report. J Immunother Cancer. 2020;8(1):e000217. doi: 10.1136/jitc-2019-000217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Parkhurst MR, Robbins PF, Tran E, Prickett TD, Gartner JJ, Jia L, Ivey G, Li YF, El-Gamil M, Lalani A, et al. Unique neoantigens arise from somatic mutations in patients with gastrointestinal cancers. Cancer Discov. 2019;9(8):1022–1035. doi: 10.1158/2159-8290.CD-18-1494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Schumacher TN, Schreiber RD. Neoantigens in cancer immunotherapy. Science. 2015;348(6230):69–74. doi: 10.1126/science.aaa4971. [DOI] [PubMed] [Google Scholar]
- 31.Blaha DT, Anderson SD, Yoakum DM, Hager MV, Zha Y, Gajewski TF, Kranz DM. High-throughput stability screening of neoantigen/HLA complexes improves immunogenicity predictions. Cancer Immunol Res. 2019;7(1):50–61. doi: 10.1158/2326-6066.CIR-18-0395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ng SB, Turner EH, Robertson PD, Flygare SD, Bigham AW, Lee C, Shaffer T, Wong M, Bhattacharjee A, Eichler EE, et al. Targeted capture and massively parallel sequencing of 12 human exomes. Nature. 2009;461(7261):272–276. doi: 10.1038/nature08250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Purcell AW, Ramarathinam SH, Ternette N. Mass spectrometry-based identification of MHC-bound peptides for immunopeptidomics. Nat Protoc. 2019;14(6):1687–1707. doi: 10.1038/s41596-019-0133-y. [DOI] [PubMed] [Google Scholar]
- 34.Li H, Durbin R. Fast and accurate short read alignment with burrows-wheeler transform. Bioinformatics. 2009;25(14):1754–1760. doi: 10.1093/bioinformatics/btp324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang K, Li M, Hakonarson H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010;38(16):e164. doi: 10.1093/nar/gkq603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cibulskis K, Lawrence MS, Carter SL, Sivachenko A, Jaffe D, Sougnez C, Gabriel S, Meyerson M, Lander ES, Getz G. Sensitive detection of somatic point mutations in impure and heterogeneous cancer samples. Nat Biotechnol. 2013;31(3):213–219. doi: 10.1038/nbt.2514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Larson DE, Harris CC, Chen K, Koboldt DC, Abbott TE, Dooling DJ, Ley TJ, Mardis ER, Wilson RK, Ding L. SomaticSniper: identification of somatic point mutations in whole genome sequencing data. Bioinformatics. 2012;28(3):311–317. doi: 10.1093/bioinformatics/btr665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Koboldt DC, Zhang Q, Larson DE, Shen D, McLellan MD, Lin L, Miller CA, Mardis ER, Ding L, Wilson RK. VarScan 2: somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012;22(3):568–576. doi: 10.1101/gr.129684.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Duan F, Duitama J, Al Seesi S, Ayres CM, Corcelli SA, Pawashe AP, Blanchard T, McMahon D, Sidney J, Sette A, et al. Genomic and bioinformatic profiling of mutational neoepitopes reveals new rules to predict anticancer immunogenicity. J Exp Med. 2014;211(11):2231–2248. doi: 10.1084/jem.20141308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhang H, Lundegaard C, Nielsen M. Pan-specific MHC class I predictors: a benchmark of HLA class I pan-specific prediction methods. Bioinformatics. 2009;25(1):83–89. doi: 10.1093/bioinformatics/btn579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.O'Donnell TJ, Rubinsteyn A, Bonsack M, Riemer AB, Laserson U, Hammerbacher J. MHCflurry: open-source class I MHC binding affinity prediction. Cell Syst. 2018;7(1):129–132.e4. doi: 10.1016/j.cels.2018.05.014. [DOI] [PubMed] [Google Scholar]
- 42.Sarkizova S, Klaeger S, Le PM, Li LW, Oliveira G, Keshishian H, Hartigan CR, Zhang W, Braun DA, Ligon KL, et al. A large peptidome dataset improves HLA class I epitope prediction across most of the human population. Nat Biotechnol. 2020;38(2):199–209. doi: 10.1038/s41587-019-0322-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Shao XM, Bhattacharya R, Huang J, Sivakumar IKA, Tokheim C, Zheng L, Hirsch D, Kaminow B, Omdahl A, Bonsack M, et al. High-throughput prediction of MHC class I and II neoantigens with MHCnuggets. Cancer Immunol Res. 2020;8(3):396–408. doi: 10.1158/2326-6066.CIR-19-0464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Liu C, Zhang Y, Jian X, Tan X, Lu M, Ouyang J, Liu Z, Li Y, Xu L, Chen L, et al. ProGeo-neo v2.0: a one-stop software for neoantigen prediction and filtering based on the proteogenomics strategy. Genes (Basel) 2022;13(5):783. doi: 10.3390/genes13050783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Vita R, Mahajan S, Overton JA, Dhanda SK, Martini S, Cantrell JR, Wheeler DK, Sette A, Peters B. The Immune epitope database (IEDB): 2018 update. Nucleic Acids Res. 2019;47(D1):D339–D343. doi: 10.1093/nar/gky1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chronister WD, Crinklaw A, Mahajan S, Vita R, Kosaloglu-Yalcin Z, Yan Z, et al. TCRMatch: predicting T-cell receptor specificity based on sequence similarity to previously characterized receptors. Front Immunol. 2021;12:640725. [DOI] [PMC free article] [PubMed]
- 47.Wells DK, van Buuren MM, Dang KK, et al. Key parameters of tumor epitope immunogenicity revealed through a consortium approach improve neoantigen prediction. Cell. 2020;183(3):818–834. doi: 10.1016/j.cell.2020.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Jurtz VPS, Andreatta M, Marcatili P, Peters B, Nielsen M. NetMHCpan 4.0 Improved peptide-MHC class I interaction predictions integrating eluted ligand and peptide binding affinity data1. J Immunol. 2017;199(9):3360–3368. doi: 10.4049/jimmunol.1700893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Jensen KK, Andreatta M, Marcatili P, Buus S, Greenbaum JA, Yan Z, Sette A, Peters B, Nielsen M. Improved methods for predicting peptide binding affinity to MHC class II molecules. Immunology. 2018;154(3):394–406. doi: 10.1111/imm.12889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.O'Donnell TJ, Rubinsteyn A, Laserson U. MHCflurry 2.0: improved pan-allele prediction of MHC class I-presented peptides by incorporating antigen processing. Cell Syst. 2020;11(1):42–48. doi: 10.1016/j.cels.2020.06.010. [DOI] [PubMed] [Google Scholar]
- 51.Bassani-Sternberg M, Chong C, Guillaume P, Solleder M, Pak H, Gannon PO, Kandalaft LE, Coukos G, Gfeller D. Deciphering HLA-I motifs across HLA peptidomes improves neo-antigen predictions and identifies allostery regulating HLA specificity. PLoS Comput Biol. 2017;13(8):e1005725. doi: 10.1371/journal.pcbi.1005725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Larsen MV, Lundegaard C, Lamberth K, Buus S, Lund O, Nielsen M. Large-scale validation of methods for cytotoxic T-lymphocyte epitope prediction. BMC Bioinformatics. 2007;8:424. doi: 10.1186/1471-2105-8-424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Roth A, Khattra J, Yap D, Wan A, Laks E, Biele J, Ha G, Aparicio S, Bouchard-Cote A, Shah SP. PyClone: statistical inference of clonal population structure in cancer. Nat Methods. 2014;11(4):396–398. doi: 10.1038/nmeth.2883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bjerregaard AM, Nielsen M, Hadrup SR, Szallasi Z, Eklund AC. MuPeXI: prediction of neo-epitopes from tumor sequencing data. Cancer Immunol Immunother. 2017;66(9):1123–1130. doi: 10.1007/s00262-017-2001-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ali M, Foldvari Z, Giannakopoulou E, et al. Induction of neoantigen-reactive T cells from healthy donors. Nat Protoc. 2019;14(6):1926–1943. doi: 10.1038/s41596-019-0170-6. [DOI] [PubMed] [Google Scholar]
- 56.Dudley ME, Wunderlich JR, Shelton TE, Even J, Rosenberg SA. Generation of tumor-infiltrating lymphocyte cultures for use in adoptive transfer therapy for melanoma patients. J Immunother. 2003;26(4):332–342. doi: 10.1097/00002371-200307000-00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Chow YT, Chen S, Wang R, Liu C, Kong CW, Li RA, Cheng SH, Sun D. Single cell transfection through precise microinjection with quantitatively controlled injection volumes. Sci Rep. 2016;6:24127. doi: 10.1038/srep24127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kumar ARK, Shou Y, Chan B, Krishaa L, Tay A. Materials for Improving immune cell transfection. Adv Mater. 2021;33(21):e2007421. doi: 10.1002/adma.202007421. [DOI] [PubMed] [Google Scholar]
- 59.Zappasodi R, Di Nicola M, Carlo-Stella C, Mortarini R, Molla A, Vegetti C, Albani S, Anichini A, Gianni AM. The effect of artificial antigen-presenting cells with preclustered anti-CD28/-CD3/-LFA-1 monoclonal antibodies on the induction of ex vivo expansion of functional human antitumor T cells. Haematologica. 2008;93(10):1523–1534. doi: 10.3324/haematol.12521. [DOI] [PubMed] [Google Scholar]
- 60.Turtle CJRS. Artificial antigen-presenting cells for use in adoptive immunotherapy. Cancer J. 2010;16(4):374–381. doi: 10.1097/PPO.0b013e3181eb33a6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ichikawa J, Yoshida T, Isser A, Laino AS, Vassallo M, Woods D, Kim S, Oelke M, Jones K, Schneck JP, et al. Rapid expansion of highly functional antigen-specific T cells from patients with melanoma by nanoscale artificial antigen-presenting cells. Clin Cancer Res. 2020;26(13):3384–3396. doi: 10.1158/1078-0432.CCR-19-3487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Xiao P, Wang J, Zhao Z, Liu X, Sun X, Wang D, Li Y. Engineering nanoscale artificial antigen-presenting cells by metabolic dendritic cell labeling to potentiate cancer immunotherapy. Nano Lett. 2021;21(5):2094–2103. doi: 10.1021/acs.nanolett.0c04783. [DOI] [PubMed] [Google Scholar]
- 63.Song S, Jin X, Zhang L, Zhao C, Ding Y, Ang Q, Khaidav O, Shen C. PEGylated and CD47-conjugated nanoellipsoidal artificial antigen-presenting cells minimize phagocytosis and augment anti-tumor T-cell responses. Int J Nanomedicine. 2019;14:2465–2483. doi: 10.2147/IJN.S195828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Meyer RA, Sunshine JC, Perica K, Kosmides AK, Aje K, Schneck JP, Green JJ. Biodegradable nanoellipsoidal artificial antigen presenting cells for antigen specific T-cell activation. Small. 2015;11(13):1519–1525. doi: 10.1002/smll.201402369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kato T, Matsuda T, Ikeda Y, Park JH, Leisegang M, Yoshimura S, Hikichi T, Harada M, Zewde M, Sato S, Hasegawa K, Kiyotani K, Nakamura Y. Effective screening of T cells recognizing neoantigens and construction of T-cell receptor-engineered T cells. Oncotarget. 2018;9(13):11009–11019. doi: 10.18632/oncotarget.24232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Chan JD, Lai J, Slaney CY, Kallies A, Beavis PA, Darcy PK. Cellular networks controlling T cell persistence in adoptive cell therapy. Nat Rev Immunol. 2021;21(12):769–784. doi: 10.1038/s41577-021-00539-6. [DOI] [PubMed] [Google Scholar]
- 67.Sakellariou-Thompson D, Forget MA, Creasy C, Bernard V, Zhao L, Kim YU, Hurd MW, Uraoka N, Parra ER, Kang Y, et al. 4–1BB agonist focuses CD8(+) tumor-infiltrating T-cell growth into a distinct repertoire capable of tumor recognition in pancreatic cancer. Clin Cancer Res. 2017;23(23):7263–7275. doi: 10.1158/1078-0432.CCR-17-0831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kwakkenbos MJ, Bakker AQ, van Helden PM, Wagner K, Yasuda E, Spits H, Beaumont T. Genetic manipulation of B cells for the isolation of rare therapeutic antibodies from the human repertoire. Methods. 2014;65(1):38–43. doi: 10.1016/j.ymeth.2013.07.002. [DOI] [PubMed] [Google Scholar]
- 69.Forget MA, Malu S, Liu H, Toth C, Maiti S, Kale C, Haymaker C, Bernatchez C, Huls H, Wang E, Marincola FM, Hwu P, Cooper LJ, Radvanyi LG. Activation and propagation of tumor-infiltrating lymphocytes on clinical-grade designer artificial antigen-presenting cells for adoptive immunotherapy of melanoma. J Immunother. 2014;37(9):448–469. doi: 10.1097/CJI.0000000000000056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Scheffel MJ, Scurti G, Simms P, Garrett-Mayer E, Mehrotra S, Nishimura MI, Voelkel-Johnson C. Efficacy of adoptive T-cell therapy is improved by treatment with the antioxidant n-acetyl cysteine, which limits activation-induced T-cell death. Cancer Res. 2016;76(20):6006–6016. doi: 10.1158/0008-5472.CAN-16-0587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Tran KQ, Zhou J, Durflinger KH, Langhan MM, Shelton TE, Wunderlich JR, Robbins PF, Rosenberg SA, Dudley ME. Minimally cultured tumor-infiltrating lymphocytes display optimal characteristics for adoptive cell therapy. J Immunother. 2008;31(8):742–751. doi: 10.1097/CJI.0b013e31818403d5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Dijkstra KK, Cattaneo CM, Weeber F, Chalabi M, van de Haar J, Fanchi LF, Slagter M, van der Velden DL, Kaing S, Kelderman S, et al. Generation of tumor-reactive T cells by co-culture of peripheral blood lymphocytes and tumor organoids. Cell. 2018;174(6):1586–1598.e12. doi: 10.1016/j.cell.2018.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Wang W, Yuan T, Ma L, Zhu Y, Bao J, Zhao X, Zhao Y, Zong Y, Zhang Y, Yang S, et al. Hepatobiliary tumor organoids reveal HLA class I neoantigen landscape and antitumoral activity of neoantigen peptide enhanced with immune checkpoint inhibitors. Adv Sci (Weinh) 2022;9:e2105810. doi: 10.1002/advs.202105810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Prakadan SM, Shalek AK, Weitz DA. Scaling by shrinking: empowering single-cell 'omics' with microfluidic devices. Nat Rev Genet. 2017;18(6):345–361. doi: 10.1038/nrg.2017.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Papalexi E, Satija R. Single-cell RNA sequencing to explore immune cell heterogeneity. Nat Rev Immunol. 2018;18(1):35–45. doi: 10.1038/nri.2017.76. [DOI] [PubMed] [Google Scholar]
- 76.Pai JA, Satpathy AT. High-throughput and single-cell T cell receptor sequencing technologies. Nat Methods. 2021;18(8):881–892. doi: 10.1038/s41592-021-01201-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Pasetto A, Gros A, Robbins PF, Deniger DC, Prickett TD, Matus-Nicodemos R, Douek DC, Howie B, Robins H, Parkhurst MR, et al. Tumor- and neoantigen-reactive T-cell receptors can be identified based on their frequency in fresh tumor. Cancer Immunol Res. 2016;4(9):734–743. doi: 10.1158/2326-6066.CIR-16-0001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Bobisse S, Genolet R, Roberti A, Tanyi JL, Racle J, Stevenson BJ, Iseli C, Michel A, Le Bitoux MA, Guillaume P, et al. Sensitive and frequent identification of high avidity neo-epitope specific CD8 (+) T cells in immunotherapy-naive ovarian cancer. Nat Commun. 2018;9(1):1092. doi: 10.1038/s41467-018-03301-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Lowery FJ, Krishna S, Yossef R, et al. Molecular signatures of antitumor neoantigen-reactive T cells from metastatic human cancers. Science. 2022;375(6583):877–884. doi: 10.1126/science.abl5447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Chester C, Sanmamed MF, Wang J, Melero I. Immunotherapy targeting 4–1BB: mechanistic rationale, clinical results, and future strategies. Blood. 2018;131(1):49–57. doi: 10.1182/blood-2017-06-741041. [DOI] [PubMed] [Google Scholar]
- 81.Parkhurst M, Gros A, Pasetto A, Prickett T, Crystal JS, Robbins P, Rosenberg SA. Isolation of T-cell receptors specifically reactive with mutated tumor-associated antigens from tumor-infiltrating lymphocytes based on CD137 expression. Clin Cancer Res. 2017;23(10):2491–2505. doi: 10.1158/1078-0432.CCR-16-2680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Duhen T, Duhen R, Montler R, Moses J, Moudgil T, de Miranda NF, Goodall CP, Blair TC, Fox BA, McDermott JE, et al. Co-expression of CD39 and CD103 identifies tumor-reactive CD8 T cells in human solid tumors. Nat Commun. 2018;9(1):2724. doi: 10.1038/s41467-018-05072-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Krishna S, Lowery FJ, Copeland AR, Bahadiroglu E, Mukherjee R, Jia L, Anibal JT, Sachs A, Adebola SO, Gurusamy D, Yu Z, Hill V, Gartner JJ, Li YF, Parkhurst M, Paria B, Kvistborg P, Kelly MC, Goff SL, Altan-Bonnet G, Robbins PF, Rosenberg SA. Stem-like CD8 T cells mediate response of adoptive cell immunotherapy against human cancer. Science. 2020;370(6522):1328–1334. doi: 10.1126/science.abb9847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Hanada KI, Zhao C, Gil-Hoyos R, Gartner JJ, Chow-Parmer C, Lowery FJ, Krishna S, Prickett TD, Kivitz S, Parkhurst MR, et al. A phenotypic signature that identifies neoantigen-reactive T cells in fresh human lung cancers. Cancer Cell. 2022;40(5):479–493.e6. doi: 10.1016/j.ccell.2022.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.He J, Xiong X, Yang H, Li D, Liu X, Li S, Liao S, Chen S, Wen X, Yu K, Fu L, Dong X, Zhu K, Xia X, Kang T, Bian C, Li X, Liu H, Ding P, Zhang X, Liu Z, Li W, Zuo Z, Zhou P. Defined tumor antigen-specific T cells potentiate personalized TCR-T cell therapy and prediction of immunotherapy response. Cell Res. 2022;32(6):530–542. doi: 10.1038/s41422-022-00627-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Veatch JR, Lee SM, Shasha C, Singhi N, Szeto JL, Moshiri AS, Kim TS, Smythe K, Kong P, Fitzgibbon M, et al. Neoantigen-specific CD4+ T cells in human melanoma have diverse differentiation states and correlate with CD8+ T cell, macrophage, and B cell function. Cancer Cell. 2022;40(4):393–409.e9. doi: 10.1016/j.ccell.2022.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Bajwa G, Lanz I, Cardenas M, Brenner MK, Arber C. Transgenic CD8alphabeta co-receptor rescues endogenous TCR function in TCR-transgenic virus-specific T cells. J Immunother Cancer. 2020;8(2):e001487. [DOI] [PMC free article] [PubMed]
- 88.Shenderov E, Kandasamy M, Gileadi U, Chen J, Shepherd D, Gibbs J, Prota G, Silk JD, Yewdell JW, Cerundolo V. Generation and characterization of HLA-A2 transgenic mice expressing the human TCR 1G4 specific for the HLA-A2 restricted NY-ESO-1157-165 tumor-specific peptide. J Immunother Cancer. 2021;9(6):e002544. doi: 10.1136/jitc-2021-002544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Rath JA, Arber C. Engineering strategies to enhance TCR-based adoptive T cell therapy. Cell. 2020;9(6):1485. doi: 10.3390/cells9061485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Halim L, Das KK, Larcombe-Young D, Ajina A, Candelli A, Benjamin R, Dillon R, Davies DM, Maher J. Engineering of an avidity-optimized CD19-specific parallel chimeric antigen receptor that delivers dual CD28 and 4–1BB Co-stimulation. Front Immunol. 2022;13:836549. doi: 10.3389/fimmu.2022.836549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Roselli E, Boucher JC, Li G, Kotani H, Spitler K, Reid K, et al. 4-1BB and optimized CD28 co-stimulation enhances function of human mono-specific and bi-specific third-generation CAR T cells. J Immunother Cancer. 2021;9(10):e003354. [DOI] [PMC free article] [PubMed]
- 92.Guercio M, Orlando D, Di Cecca S, Sinibaldi M, Boffa I, Caruso S, Abbaszadeh Z, Camera A, Cembrola B, Bovetti K, et al. CD28. OX40 co-stimulatory combination is associated with long in vivo persistence and high activity of CAR CD30 T-cells. Haematologica. 2021;106(4):987–999. doi: 10.3324/haematol.2019.231183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Fedorov VD, Themeli M, Sadelain M. PD-1- and CTLA-4-based inhibitory chimeric antigen receptors (iCARs) divert off-target immunotherapy responses. Sci Transl Med. 2013;5(215):215ra172. doi: 10.1126/scitranslmed.3006597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Liu H, Lei W, Zhang C, Yang C, Wei J, Guo Q, Guo X, Chen Z, Lu Y, Young KH, et al. CD19-specific CAR T cells that express a PD-1/CD28 chimeric switch-receptor are effective in patients with PD-L1-positive B-cell lymphoma. Clin Cancer Res. 2021;27(2):473–484. doi: 10.1158/1078-0432.CCR-20-1457. [DOI] [PubMed] [Google Scholar]
- 95.Hoogi S, Eisenberg V, Mayer S, Shamul A, Barliya T, Cohen CJ. A TIGIT-based chimeric co-stimulatory switch receptor improves T-cell anti-tumor function. J Immunother Cancer. 2019;7(1):243. doi: 10.1186/s40425-019-0721-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Zhang Q, Hresko ME, Picton LK, Su L, Hollander MJ, Nunez-Cruz S, Zhang Z, Assenmacher CA, Sockolosky JT, Garcia KC, et al. A human orthogonal IL-2 and IL-2Rbeta system enhances CAR T cell expansion and antitumor activity in a murine model of leukemia. Sci Transl Med. 2021;13(625):eabg6986. doi: 10.1126/scitranslmed.abg6986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kalbasi A, Siurala M, Su LL, Tariveranmoshabad M, Picton LK, Ravikumar P, Li P, Lin JX, Escuin-Ordinas H, Da T, et al. Potentiating adoptive cell therapy using synthetic IL-9 receptors. Nature. 2022;612(7938):E10. doi: 10.1038/s41586-022-05548-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Kagoya Y, Tanaka S, Guo T, Anczurowski M, Wang C-H, Saso K, Butler MO, Minden MD, Hirano N. A novel chimeric antigen receptor containing a JAK–STAT signaling domain mediates superior antitumor effects. Nat Med. 2018;24(3):352–359. doi: 10.1038/nm.4478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Luo H, Su J, Sun R, Sun Y, Wang Y, Dong Y, Shi B, Jiang H, Li Z. Coexpression of IL7 and CCL21 increases efficacy of CAR-T cells in solid tumors without requiring preconditioned lymphodepletion. Clin Cancer Res. 2020;26(20):5494–5505. doi: 10.1158/1078-0432.CCR-20-0777. [DOI] [PubMed] [Google Scholar]
- 100.Kerkar SP, Muranski P, Kaiser A, Boni A, Sanchez-Perez L, Yu Z, Palmer DC, Reger RN, Borman ZA, Zhang L, et al. Tumor-specific CD8+ T cells expressing interleukin-12 eradicate established cancers in lymphodepleted hosts. Cancer Res. 2010;70(17):6725–6734. doi: 10.1158/0008-5472.CAN-10-0735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Zhang L, Morgan RA, Beane JD, Zheng Z, Dudley ME, Kassim SH, Nahvi AV, Ngo LT, Sherry RM, Phan GQ, et al. Tumor-infiltrating lymphocytes genetically engineered with an inducible gene encoding interleukin-12 for the immunotherapy of metastatic melanoma. Clin Cancer Res. 2015;21(10):2278–2288. doi: 10.1158/1078-0432.CCR-14-2085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Krenciute G, Prinzing BL, Yi Z, Wu MF, Liu H, Dotti G, Balyasnikova IV, Gottschalk S. Transgenic expression of IL15 improves antiglioma activity of IL13Ralpha2-CAR T cells but results in antigen loss variants. Cancer Immunol Res. 2017;5(7):571–581. doi: 10.1158/2326-6066.CIR-16-0376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Drakes DJ, Rafiq S, Purdon TJ, Lopez AV, Chandran SS, Klebanoff CA, Brentjens RJ. Optimization of T-cell receptor-modified T cells for cancer therapy. Cancer Immunol Res. 2020;8(6):743–755. doi: 10.1158/2326-6066.CIR-19-0910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Ma X, Shou P, Smith C, Chen Y, Du H, Sun C, Porterfield Kren N, Michaud D, Ahn S, Vincent B, et al. Interleukin-23 engineering improves CAR T cell function in solid tumors. Nat Biotechnol. 2020;38(4):448–459. doi: 10.1038/s41587-019-0398-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Adachi K, Kano Y, Nagai T, Okuyama N, Sakoda Y, Tamada K. IL-7 and CCL19 expression in CAR-T cells improves immune cell infiltration and CAR-T cell survival in the tumor. Nat Biotechnol. 2018;36(4):346–351. doi: 10.1038/nbt.4086. [DOI] [PubMed] [Google Scholar]
- 106.Lai J, Mardiana S, House IG, Sek K, Henderson MA, Giuffrida L, Chen AXY, Todd KL, Petley EV, Chan JD, et al. Adoptive cellular therapy with T cells expressing the dendritic cell growth factor Flt3L drives epitope spreading and antitumor immunity. Nat Immunol. 2020;21(8):914–926. doi: 10.1038/s41590-020-0676-7. [DOI] [PubMed] [Google Scholar]
- 107.Wang G, Zhang Z, Zhong K, Wang Z, Yang N, Tang X, Li H, Lu Q, Wu Z, Yuan B, et al. CXCL11-armed oncolytic adenoviruses enhance CAR-T cell therapeutic efficacy and reprogram tumor microenvironment in glioblastoma. Mol Ther. 2023;31(1):134–153. doi: 10.1016/j.ymthe.2022.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Li X, Lu M, Yuan M, Ye J, Zhang W, Xu L, Wu X, Hui B, Yang Y, Wei B, et al. CXCL10-armed oncolytic adenovirus promotes tumor-infiltrating T-cell chemotaxis to enhance anti-PD-1 therapy. Oncoimmunology. 2022;11(1):2118210. doi: 10.1080/2162402X.2022.2118210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Tian Y, Wen C, Zhang Z, Liu Y, Li F, Zhao Q, Yao C, Ni K, Yang S, Zhang Y. CXCL9-modified CAR T cells improve immune cell infiltration and antitumor efficacy. Cancer Immunol Immunother. 2022;71(11):2663–2675. doi: 10.1007/s00262-022-03193-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Depil S, Duchateau P, Grupp SA, Mufti G, Poirot L. ‘Off-the-shelf’ allogeneic CAR T cells: development and challenges. Nat Rev Drug Discovery. 2020;19(3):185–199. doi: 10.1038/s41573-019-0051-2. [DOI] [PubMed] [Google Scholar]
- 111.Johnson LR, Lee DY, Eacret JS, Ye D, June CH, Minn AJ. The immunostimulatory RNA RN7SL1 enables CAR-T cells to enhance autonomous and endogenous immune function. Cell. 2021;184(19):4981–4995.e14. doi: 10.1016/j.cell.2021.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Rafiq S, Hackett CS, Brentjens RJ. Engineering strategies to overcome the current roadblocks in CAR T cell therapy. Nat Rev Clin Oncol. 2020;17(3):147–167. doi: 10.1038/s41571-019-0297-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Tokarew N, Ogonek J, Endres S, von Bergwelt-Baildon M, Kobold S. Teaching an old dog new tricks: next-generation CAR T cells. Br J Cancer. 2019;120(1):26–37. doi: 10.1038/s41416-018-0325-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Dhatchinamoorthy K, Colbert JD, Rock KL. Cancer immune evasion through loss of MHC class I antigen presentation. Front Immunol. 2021;12:636568. [DOI] [PMC free article] [PubMed]
- 115.Shklovskaya E, Rizos H. MHC class I deficiency in solid tumors and therapeutic strategies to overcome it. Int J Mol Sci. 2021;22(13):6741. doi: 10.3390/ijms22136741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Rosenthal R, Cadieux EL, Salgado R, Bakir MA, Moore DA, Hiley CT, Lund T, Tanic M, Reading JL, Joshi K, et al. Neoantigen-directed immune escape in lung cancer evolution. Nature. 2019;567(7749):479–485. doi: 10.1038/s41586-019-1032-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Zhou F. Molecular mechanisms of IFN-gamma to up-regulate MHC class I antigen processing and presentation. Int Rev Immunol. 2009;28(3–4):239–260. doi: 10.1080/08830180902978120. [DOI] [PubMed] [Google Scholar]
- 118.Ugurel S, Spassova I, Wohlfarth J, Drusio C, Cherouny A, Melior A, Sucker A, Zimmer L, Ritter C, Schadendorf D, et al. MHC class-I downregulation in PD-1/PD-L1 inhibitor refractory Merkel cell carcinoma and its potential reversal by histone deacetylase inhibition: a case series. Cancer Immunol Immunother. 2019;68(6):983–990. doi: 10.1007/s00262-019-02341-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Luo N, Nixon MJ, Gonzalez-Ericsson PI, Sanchez V, Opalenik SR, Li H, Zahnow CA, Nickels ML, Liu F, Tantawy MN, et al. DNA methyltransferase inhibition upregulates MHC-I to potentiate cytotoxic T lymphocyte responses in breast cancer. Nat Commun. 2018;9(1):248. doi: 10.1038/s41467-017-02630-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Ennishi D, Takata K, Beguelin W, Duns G, Mottok A, Farinha P, Bashashati A, Saberi S, Boyle M, Meissner B, et al. Molecular and genetic characterization of MHC deficiency identifies EZH2 as therapeutic target for enhancing immune recognition. Cancer Discov. 2019;9(4):546–563. doi: 10.1158/2159-8290.CD-18-1090. [DOI] [PubMed] [Google Scholar]
- 121.Briere D, Sudhakar N, Woods DM, Hallin J, Engstrom LD, Aranda R, Chiang H, Sodre AL, Olson P, Weber JS, et al. The class I/IV HDAC inhibitor mocetinostat increases tumor antigen presentation, decreases immune suppressive cell types and augments checkpoint inhibitor therapy. Cancer Immunol Immunother. 2018;67(3):381–392. doi: 10.1007/s00262-017-2091-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Yamamoto K, Venida A, Yano J, Biancur DE, Kakiuchi M, Gupta S, Sohn ASW, Mukhopadhyay S, Lin EY, Parker SJ, et al. Autophagy promotes immune evasion of pancreatic cancer by degrading MHC-I. Nature. 2020;581(7806):100–105. doi: 10.1038/s41586-020-2229-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Badrinath S, Dellacherie MO, Li A, Zheng S, Zhang X, Sobral M, Pyrdol JW, Smith KL, Lu Y, Haag S, et al. A vaccine targeting resistant tumours by dual T cell plus NK cell attack. Nature. 2022;606(7916):992–998. doi: 10.1038/s41586-022-04772-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Hegde PS, Chen DS. Top 10 challenges in cancer immunotherapy. Immunity. 2020;52(1):17–35. doi: 10.1016/j.immuni.2019.12.011. [DOI] [PubMed] [Google Scholar]
- 125.Liu YT, Sun ZJ. Turning cold tumors into hot tumors by improving T-cell infiltration. Theranostics. 2021;11(11):5365–5386. doi: 10.7150/thno.58390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Ahmadzadeh M, Johnson LA, Heemskerk B, Wunderlich JR, Dudley ME, White DE, Rosenberg SA. Tumor antigen-specific CD8 T cells infiltrating the tumor express high levels of PD-1 and are functionally impaired. Blood. 2009;114(8):1537–1544. doi: 10.1182/blood-2008-12-195792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Grassberger C, Ellsworth SG, Wilks MQ, Keane FK, Loeffler JS. Assessing the interactions between radiotherapy and antitumour immunity. Nat Rev Clin Oncol. 2019;16(12):729–745. doi: 10.1038/s41571-019-0238-9. [DOI] [PubMed] [Google Scholar]
- 128.Galluzzi L, Humeau J, Buque A, Zitvogel L, Kroemer G. Immunostimulation with chemotherapy in the era of immune checkpoint inhibitors. Nat Rev Clin Oncol. 2020;17(12):725–741. doi: 10.1038/s41571-020-0413-z. [DOI] [PubMed] [Google Scholar]
- 129.McLaughlin M, Patin EC, Pedersen M, Wilkins A, Dillon MT, Melcher AA, Harrington KJ. Inflammatory microenvironment remodelling by tumour cells after radiotherapy. Nat Rev Cancer. 2020;20(4):203–217. doi: 10.1038/s41568-020-0246-1. [DOI] [PubMed] [Google Scholar]
- 130.Chu KF, Dupuy DE. Thermal ablation of tumours: biological mechanisms and advances in therapy. Nat Rev Cancer. 2014;14(3):199–208. doi: 10.1038/nrc3672. [DOI] [PubMed] [Google Scholar]
- 131.Roma-Rodrigues C, Mendes R, Baptista PV, Fernandes AR. Targeting tumor microenvironment for cancer therapy. Int J Mol Sci. 2019;20(4):840. doi: 10.3390/ijms20040840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Zhu Y, Qian Y, Li Z, Li Y, Li B. Neoantigen-reactive T cell: An emerging role in adoptive cellular immunotherapy. MedComm (2020) 2021;2(2):207–220. doi: 10.1002/mco2.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Frese KK, Neesse A, Cook N, Bapiro TE, Lolkema MP, Jodrell DI, Tuveson DA. nab-Paclitaxel potentiates gemcitabine activity by reducing cytidine deaminase levels in a mouse model of pancreatic cancer. Cancer Discov. 2012;2(3):260–269. doi: 10.1158/2159-8290.CD-11-0242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Yardley DA. nab-Paclitaxel mechanisms of action and delivery. J Control Release. 2013;170(3):365–372. doi: 10.1016/j.jconrel.2013.05.041. [DOI] [PubMed] [Google Scholar]
- 135.Von Hoff DD, Ervin T, Arena FP, Chiorean EG, Infante J, Moore M, Seay T, Tjulandin SA, Ma WW, Saleh MN, et al. Increased survival in pancreatic cancer with nab-paclitaxel plus gemcitabine. N Engl J Med. 2013;369(18):1691–1703. doi: 10.1056/NEJMoa1304369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Huang Y, Kim BYS, Chan CK, Hahn SM, Weissman IL, Jiang W. Improving immune-vascular crosstalk for cancer immunotherapy. Nat Rev Immunol. 2018;18(3):195–203. doi: 10.1038/nri.2017.145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Tian L, Goldstein A, Wang H, Ching Lo H, Sun Kim I, Welte T, Sheng K, Dobrolecki LE, Zhang X, Putluri N, et al. Mutual regulation of tumour vessel normalization and immunostimulatory reprogramming. Nature. 2017;544(7649):250–254. doi: 10.1038/nature21724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Sugiyama D, Nishikawa H, Maeda Y, Nishioka M, Tanemura A, Katayama I, Ezoe S, Kanakura Y, Sato E, Fukumori Y, et al. Anti-CCR4 mAb selectively depletes effector-type FoxP3+CD4+ regulatory T cells, evoking antitumor immune responses in humans. Proc Natl Acad Sci U S A. 2013;110(44):17945–17950. doi: 10.1073/pnas.1316796110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Fultang L, Panetti S, Ng M, Collins P, Graef S, Rizkalla N, Booth S, Lenton R, Noyvert B, Shannon-Lowe C, et al. MDSC targeting with Gemtuzumab ozogamicin restores T cell immunity and immunotherapy against cancers. EBioMedicine. 2019;47:235–246. doi: 10.1016/j.ebiom.2019.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Highfill SL, Cui Y, Giles AJ, Smith JP, Zhang H, Morse E, Kaplan RN, Mackall CL. Disruption of CXCR2-mediated MDSC tumor trafficking enhances anti-PD1 efficacy. Sci Transl Med. 2014;6(237):237ra267. doi: 10.1126/scitranslmed.3007974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Dixon ML, Luo L, Ghosh S, Grimes JM, Leavenworth JD, Leavenworth JW. Remodeling of the tumor microenvironment via disrupting Blimp1(+) effector Treg activity augments response to anti-PD-1 blockade. Mol Cancer. 2021;20(1):150. doi: 10.1186/s12943-021-01450-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Tavazoie MF, Pollack I, Tanqueco R, Ostendorf BN, Reis BS, Gonsalves FC, Kurth I, Andreu-Agullo C, Derbyshire ML, Posada J, et al. LXR/ApoE activation restricts innate immune suppression in cancer. Cell. 2018;172(4):825–840.e18. doi: 10.1016/j.cell.2017.12.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Mohamed E, Sierra RA, Trillo-Tinoco J, Cao Y, Innamarato P, Payne KK, de Mingo PA, Mandula J, Zhang S, Thevenot P, Biswas S, Abdalla SK, Costich TL, Hänggi K, Anadon CM, Flores ER, Haura EB, Mehrotra S, Pilon-Thomas S, Ruffell B, Munn DH, Cubillos-Ruiz JR, Conejo-Garcia JR, Rodriguez PC. The unfolded protein response mediator PERK governs myeloid cell-driven immunosuppression in tumors through inhibition of STING signaling. Immunity. 2020;52(4):668–682.e7. doi: 10.1016/j.immuni.2020.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Huang W, Liu Y, Luz A, Berrong M, Meyer JN, Zou Y, et al. Calcium/Calmodulin dependent protein Kinase Kinase 2 regulates the expansion of tumor-induced Myeloid-derived suppressor cells. Front Immunol. 2021;12:754083. [DOI] [PMC free article] [PubMed]
- 145.Han S, Wang W, Wang S, Yang T, Zhang G, Wang D, Ju R, Lu Y, Wang H, Wang L. Tumor microenvironment remodeling and tumor therapy based on M2-like tumor associated macrophage-targeting nano-complexes. Theranostics. 2021;11(6):2892–2916. doi: 10.7150/thno.50928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Mohapatra S, Pioppini C, Ozpolat B, Calin GA. Non-coding RNAs regulation of macrophage polarization in cancer. Mol Cancer. 2021;20(1):24. doi: 10.1186/s12943-021-01313-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Liu C, Chikina M, Deshpande R, Menk AV, Wang T, Tabib T, Brunazzi EA, Vignali KM, Sun M, Stolz DB, et al. Treg cells promote the SREBP1-dependent metabolic fitness of tumor-promoting macrophages via repression of CD8+ T cell-derived interferon-γ. Immunity. 2019;51(2):381–397.e6. doi: 10.1016/j.immuni.2019.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Sautes-Fridman C, Petitprez F, Calderaro J, Fridman WH. Tertiary lymphoid structures in the era of cancer immunotherapy. Nat Rev Cancer. 2019;19(6):307–325. doi: 10.1038/s41568-019-0144-6. [DOI] [PubMed] [Google Scholar]
- 149.Cabrita R, Lauss M, Sanna A, Donia M, Skaarup Larsen M, Mitra S, Johansson I, Phung B, Harbst K, Vallon-Christersson J, et al. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature. 2020;577(7791):561–565. doi: 10.1038/s41586-019-1914-8. [DOI] [PubMed] [Google Scholar]
- 150.Wouters MCA, Nelson BH. Prognostic significance of tumor-infiltrating B cells and plasma cells in human cancer. Clin Cancer Res. 2018;24(24):6125–6135. doi: 10.1158/1078-0432.CCR-18-1481. [DOI] [PubMed] [Google Scholar]
- 151.Kroeger DR, Milne K, Nelson BH. Tumor-infiltrating plasma cells are associated with tertiary lymphoid structures, cytolytic t-cell responses, and superior prognosis in ovarian cancer. Clin Cancer Res. 2016;22(12):3005–3015. doi: 10.1158/1078-0432.CCR-15-2762. [DOI] [PubMed] [Google Scholar]
- 152.Meylan M, Petitprez F, Becht E, Bougouin A, Pupier G, Calvez A, Giglioli I, Verkarre V, Lacroix G, Verneau J, et al. Tertiary lymphoid structures generate and propagate anti-tumor antibody-producing plasma cells in renal cell cancer. Immunity. 2022;55(3):527–541.e5. doi: 10.1016/j.immuni.2022.02.001. [DOI] [PubMed] [Google Scholar]
- 153.Delvecchio FR, Fincham REA, Spear S, Clear A, Roy-Luzarraga M, Balkwill FR, Gribben JG, Bombardieri M, Hodivala-Dilke K, Capasso M, et al. Pancreatic cancer chemotherapy is potentiated by induction of tertiary lymphoid structures in mice. Cell Mol Gastroenterol Hepatol. 2021;12(5):1543–1565. doi: 10.1016/j.jcmgh.2021.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Zhu G, Nemoto S, Mailloux AW, Perez-Villarroel P, Nakagawa R, Falahat R, Berglund AE, Mule JJ. Induction of tertiary lymphoid structures with antitumor function by a lymph node-derived stromal cell line. Front Immunol. 2018;9:1609. doi: 10.3389/fimmu.2018.01609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Tran E, Turcotte S, Gros A, Robbins PF, Lu YC, Dudley ME, Wunderlich JR, Somerville RP, Hogan K, Hinrichs CS, Parkhurst MR, Yang JC, Rosenberg SA. Cancer immunotherapy based on mutation-specific CD4+ T cells in a patient with epithelial cancer. Science. 2014;344(6184):641–645. doi: 10.1126/science.1251102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Leidner R, Sanjuan Silva N, Huang H, Sprott D, Zheng C, Shih YP, Leung A, Payne R, Sutcliffe K, Cramer J, et al. Neoantigen T-cell receptor gene therapy in pancreatic cancer. N Engl J Med. 2022;386(22):2112–2119. doi: 10.1056/NEJMoa2119662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Kim SP, Vale NR, Zacharakis N, Krishna S, Yu Z, Gasmi B, Gartner JJ, Sindiri S, Malekzadeh P, Deniger DC, et al. Adoptive cellular therapy with autologous tumor-infiltrating lymphocytes and T-cell receptor-engineered t cells targeting common p53 neoantigens in human solid tumors. Cancer Immunol Res. 2022;10(8):932–946. doi: 10.1158/2326-6066.CIR-22-0040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Peng S, Chen S, Hu W, Mei J, Zeng X, Su T, Wang W, Chen Z, Xiao H, Zhou Q, et al. Combination neoantigen-based dendritic cell vaccination and adoptive T-cell transfer induces antitumor responses against recurrence of hepatocellular carcinoma. Cancer Immunol Res. 2022;10(6):728–744. doi: 10.1158/2326-6066.CIR-21-0931. [DOI] [PubMed] [Google Scholar]
- 159.Sahin U, Derhovanessian E, Miller M, Kloke BP, Simon P, Lower M, Bukur V, Tadmor AD, Luxemburger U, Schrors B, et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature. 2017;547(7662):222–226. doi: 10.1038/nature23003. [DOI] [PubMed] [Google Scholar]
- 160.Keskin DB, Anandappa AJ, Sun J, Tirosh I, Mathewson ND, Li S, Oliveira G, Giobbie-Hurder A, Felt K, Gjini E, et al. Neoantigen vaccine generates intratumoral T cell responses in phase Ib glioblastoma trial. Nature. 2019;565(7738):234–239. doi: 10.1038/s41586-018-0792-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Cafri G, Gartner JJ, Zaks T, Hopson K, Levin N, Paria BC, Parkhurst MR, Yossef R, Lowery FJ, Jafferji MS, et al. mRNA vaccine-induced neoantigen-specific T cell immunity in patients with gastrointestinal cancer. J Clin Invest. 2020;130(11):5976–5988. doi: 10.1172/JCI134915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Fang Y, Mo F, Shou J, Wang H, Luo K, Zhang S, Han N, Li H, Ye S, Zhou Z, et al. A pan-cancer clinical study of personalized neoantigen vaccine monotherapy in treating patients with various types of advanced solid tumors. Clin Cancer Res. 2020;26(17):4511–4520. doi: 10.1158/1078-0432.CCR-19-2881. [DOI] [PubMed] [Google Scholar]
- 163.Kloor M, Reuschenbach M, Pauligk C, Karbach J, Rafiyan MR, Al-Batran SE, Tariverdian M, Jager E, von Knebel DM. A frameshift peptide neoantigen-based vaccine for mismatch repair-deficient cancers: a phase I/IIa clinical trial. Clin Cancer Res. 2020;26(17):4503–4510. doi: 10.1158/1078-0432.CCR-19-3517. [DOI] [PubMed] [Google Scholar]
- 164.Mueller S, Taitt JM, Villanueva-Meyer JE, Bonner ER, Nejo T, Lulla RR, Goldman S, Banerjee A, Chi SN, Whipple NS, et al. Mass cytometry detects H3 .3K27M-specific vaccine responses in diffuse midline glioma. J Clin Invest. 2020;130(12):6325–6337. doi: 10.1172/JCI140378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Cai Z, Su X, Qiu L, Li Z, Li X, Dong X, Wei F, Zhou Y, Luo L, Chen G, et al. Personalized neoantigen vaccine prevents postoperative recurrence in hepatocellular carcinoma patients with vascular invasion. Mol Cancer. 2021;20(1):164. doi: 10.1186/s12943-021-01467-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Veatch JR, Lee SM, Fitzgibbon M, Chow IT, Jesernig B, Schmitt T, Kong YY, Kargl J, Houghton AM, Thompson JA, et al. Tumor-infiltrating BRAFV600E-specific CD4+ T cells correlated with complete clinical response in melanoma. J Clin Invest. 2018;128(4):1563–1568. doi: 10.1172/JCI98689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.van den Berg JH, Heemskerk B, van Rooij N, Gomez-Eerland R, Michels S, van Zon M, et al. Tumor Infiltrating Lymphocytes (TIL) therapy in metastatic melanoma: boosting of neoantigen-specific T cell reactivity and long-term follow-up. J Immunother Cancer. 2020;8(2):e000848. [DOI] [PMC free article] [PubMed]
- 168.Kristensen NP, Heeke C, Tvingsholm SA, Borch A, Draghi A, Crowther MD, Carri I, Munk KK, Holm JS, Bjerregaard AM, et al. Neoantigen-reactive CD8+ T cells affect clinical outcome of adoptive cell therapy with tumor-infiltrating lymphocytes in melanoma. J Clin Invest. 2022;132(2):e150535. doi: 10.1172/JCI150535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Creelan BC, Wang C, Teer JK, Toloza EM, Yao J, Kim S, Landin AM, Mullinax JE, Saller JJ, Saltos AN, et al. Tumor-infiltrating lymphocyte treatment for anti-PD-1-resistant metastatic lung cancer: a phase 1 trial. Nat Med. 2021;27(8):1410–1418. doi: 10.1038/s41591-021-01462-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Carreno BMMV, Becker-Hapak M, Kaabinejadian S, Hundal J, Petti AA, Ly A, Lie WR, Hildebrand WH, Mardis ER, Linette GP. Cancer immunotherapy: A dendritic cell vaccine increases the breadth and diversity of melanoma neoantigen-specific T cells. Science. 2015;348(6236):803–808. doi: 10.1126/science.aaa3828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Carreno BM, Becker-Hapak M, Huang A, Chan M, Alyasiry A, Lie WR, Aft RL, Cornelius LA, Trinkaus KM, Linette GP. IL-12p70-producing patient DC vaccine elicits Tc1-polarized immunity. J Clin Invest. 2013;123(8):3383–3394. doi: 10.1172/JCI68395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Ding Z, Li Q, Zhang R, Xie L, Shu Y, Gao S, Wang P, Su X, Qin Y, Wang Y, et al. Personalized neoantigen pulsed dendritic cell vaccine for advanced lung cancer. Signal Transduct Target Ther. 2021;6(1):26. doi: 10.1038/s41392-020-00448-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Tanyi JLBS, Ophir E, Tuyaerts S, Roberti A, Genolet R, Baumgartner P, Stevenson BJ, Iseli C, Dangaj D, Czerniecki B, Semilietof A, Racle J, Michel A, Xenarios I, Chiang C, Monos DS, Torigian DA, Nisenbaum HL, Michielin O, June CH, Levine BL, Powell DJ, Jr, Gfeller D, Mick R, Dafni U, Zoete V, Harari A, Coukos G, Kandalaft LE. Personalized cancer vaccine effectively mobilizes antitumor T cell immunity in ovarian cancer. Sci Transl Med. 2018;10(436):eaao5931. doi: 10.1126/scitranslmed.aao5931. [DOI] [PubMed] [Google Scholar]
- 174.Holm JS, Funt SA, Borch A, Munk KK, Bjerregaard AM, Reading JL, Maher C, Regazzi A, Wong P, Al-Ahmadie H, et al. Neoantigen-specific CD8 T cell responses in the peripheral blood following PD-L1 blockade might predict therapy outcome in metastatic urothelial carcinoma. Nat Commun. 2022;13(1):1935. doi: 10.1038/s41467-022-29342-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Fehlings M, Jhunjhunwala S, Kowanetz M, O'Gorman WE, Hegde PS, Sumatoh H, Lee BH, Nardin A, Becht E, Flynn S, et al. Late-differentiated effector neoantigen-specific CD8+ T cells are enriched in peripheral blood of non-small cell lung carcinoma patients responding to atezolizumab treatment. J Immunother Cancer. 2019;7(1):249. doi: 10.1186/s40425-019-0695-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Zhang J, Wang L. The emerging world of TCR-T cell trials against cancer: a systematic review. Technol Cancer Res Treat. 2019;18:1533033819831068. doi: 10.1177/1533033819831068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Jamal-Hanjani M, Quezada SA, Larkin J, Swanton C. Translational implications of tumor heterogeneity. Clin Cancer Res. 2015;21(6):1258–1266. doi: 10.1158/1078-0432.CCR-14-1429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Ping Y, Liu C, Zhang Y. T-cell receptor-engineered T cells for cancer treatment: current status and future directions. Protein Cell. 2018;9(3):254–266. doi: 10.1007/s13238-016-0367-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Huang J, Khong HT, Dudley ME, El-Gamil M, Li YF, Rosenberg SA, Robbins PF. Survival, persistence, and progressive differentiation of adoptively transferred tumor-reactive T cells associated with tumor regression. J Immunother. 2005;28(3):258–267. doi: 10.1097/01.cji.0000158855.92792.7a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Chandran SS, Paria BC, Srivastava AK, Rothermel LD, Stephens DJ, Dudley ME, Somerville R, Wunderlich JR, Sherry RM, Yang JC, et al. Persistence of CTL clones targeting melanocyte differentiation antigens was insufficient to mediate significant melanoma regression in humans. Clin Cancer Res. 2015;21(3):534–543. doi: 10.1158/1078-0432.CCR-14-2208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Tran E, Ahmadzadeh M, Lu YC, Gros A, Turcotte S, Robbins PF, Gartner JJ, Zheng Z, Li YF, Ray S, Wunderlich JR, Somerville RP, Rosenberg SA. Immunogenicity of somatic mutations in human gastrointestinal cancers. Science. 2015;350(6266):1387–1390. doi: 10.1126/science.aad1253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Tran E, Robbins PF, Lu YC, Prickett TD, Gartner JJ, Jia L, Pasetto A, Zheng Z, Ray S, Groh EM, et al. T-cell transfer therapy targeting mutant KRAS in cancer. N Engl J Med. 2016;375(23):2255–2262. doi: 10.1056/NEJMoa1609279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Zhao X, Pan X, Wang Y, Zhang Y. Targeting neoantigens for cancer immunotherapy. Biomark Res. 2021;9(1):61. doi: 10.1186/s40364-021-00315-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Gattinoni L, Klebanoff CA, Palmer DC, Wrzesinski C, Kerstann K, Yu Z, Finkelstein SE, Theoret MR, Rosenberg SA, Restifo NP. Acquisition of full effector function in vitro paradoxically impairs the in vivo antitumor efficacy of adoptively transferred CD8+ T cells. J Clin Invest. 2005;115(6):1616–1626. doi: 10.1172/JCI24480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Hinrichs CS, Borman ZA, Gattinoni L, Yu Z, Burns WR, Huang J, Klebanoff CA, Johnson LA, Kerkar SP, Yang S, Muranski P, Palmer DC, Scott CD, Morgan RA, Robbins PF, Rosenberg SA, Restifo NP. Human effector CD8+ T cells derived from naive rather than memory subsets possess superior traits for adoptive immunotherapy. Blood. 2011;117(3):808–814. doi: 10.1182/blood-2010-05-286286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Crompton JG, Sukumar M, Restifo NP. Uncoupling T-cell expansion from effector differentiation in cell-based immunotherapy. Immunol Rev. 2014;257(1):264–276. doi: 10.1111/imr.12135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Giavridis T, van der Stegen SJC, Eyquem J, Hamieh M, Piersigilli A, Sadelain M. CAR T cell-induced cytokine release syndrome is mediated by macrophages and abated by IL-1 blockade. Nat Med. 2018;24(6):731–738. doi: 10.1038/s41591-018-0041-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Norelli M, Camisa B, Barbiera G, Falcone L, Purevdorj A, Genua M, Sanvito F, Ponzoni M, Doglioni C, Cristofori P, et al. Monocyte-derived IL-1 and IL-6 are differentially required for cytokine-release syndrome and neurotoxicity due to CAR T cells. Nat Med. 2018;24(6):739–748. doi: 10.1038/s41591-018-0036-4. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.