

Abstract
Previous research has suggested that infants may have more robust memory for past experiences relative to memory for locations that have been encountered previously. This assertion, however, primarily results from the comparison of data that were collected using different experimental procedures. In the present study, we examined memory for events and memory for locations in the context of elicited imitation. Specifically, 13-, 16-, and 20-month-old infants were tested for long-term memory for events and locations after between-subjects delays of 1, 3, 6, 9, and 12 months. The results indicated that the event memory was retained over lengthier delays relative to the location memory, despite superior encoding of location information. The possible adaptive significance of long-term term memory for events ontogenetically preceding long-term memory for locations is discussed.
Keywords: Memory, events, locations, infancy, elicited imitation
One would experience great difficulty deciding whether the ability to remember past events or the ability to remember previously-encountered locations is the more valuable cognitive commodity, as both of these skills are required for adaptive functioning. Although both of these fundamental cognitive abilities undergo significant development during the first years of life, researchers have largely studied them using different experimental techniques. The present study was designed to determine whether memories for events and memories for locations are similarly maintained over the long term when assessed within the same experimental context, namely elicited imitation.
The research that has been conducted to date has established that the ability to remember events and locations develop concurrently, although memory for events may be better retained over the long term relative to memory for locations (Sluzenski, Newcombe, & Satlow, 2004). Direct comparisons of these two aspects of memory are complicated, however, by the fact that they have traditionally been investigated using different experimental procedures. During infancy and early childhood, memory for events is commonly assessed using the elicited imitation procedure, in which a researcher demonstrates a sequence of actions to an infant (e.g., Bauer & Mandler, 1989). Either immediately after the demonstration or after the imposition of a delay, the participant has the opportunity to interact with the stimulus materials; a researcher assesses whether the infant completes the same actions in the same order (Bauer, Wenner, Dropik, & Wewerka, 2000, for an example). Memory for locations, on the other hand, has been commonly assessed using various procedures including violation of expectation paradigms (Newcombe, Huttenlocher, & Learmonth, 1999) and locating hidden objects in laboratory testing space (Learmonth, Newcombe, & Huttenlocher, 2001), depending on the research question and the age of the participants being studied.
In addition to using dissimilar experimental methodologies, researchers have largely addressed different questions when studying memory for events and memory for locations. Whereas researchers studying memory for events have devoted significant attention to establishing the long-term time course of memory in infancy (e.g., Bauer et al., 2000), individuals examining memory for locations have primarily investigated infants’ abilities to successfully locate hidden items over short delays (Learmonth et al., 2001; Learmonth, Newcombe, Sheridan, &Jones, 2008). The only data to our knowledge to address long-term memory for locations included study of 18- to 42-month-old participants who searched for a hidden item in a sandbox after a filled two-minute delay (Sluzenski et al., 2004). The results indicated that the youngest infants were somewhat less successful at remembering the location of the hidden item after the delay relative to the older infants. In addition, the correlation between hiding and search locations was weaker and non-significant for the 18-month-old infants relative to the older age groups. The relatively poor performance of the youngest infants on this location memory task stands in stark contrast to what is known about long-term memory for events. At 6 months, infants remember one step of a three-step action sequence after a 24-hour delay (Collie & Hayne, 1996). When infants are 9 months of age, they remember the individual target actions that comprise a two-step sequence for delays of one month, but do not show reliable memory for temporal order information at that time (Bauer et al., 2001; Bauer et al., 2003; Carver & Bauer, 1999). By 10 months, infants remember the individual actions of a two-step sequence after delays of six months and show memory for temporal order information up to delays of three months (Carver & Bauer, 2001). Memory for events is further refined during the second year of life, such that 20-month-olds remember temporal order information for delays up to – and perhaps extending beyond – twelve months (Bauer et al., 2000).
As noted by Sluzenski and her colleagues (2004), the research conducted to date suggests that development of the ability to remember location-based information over the long term may be somewhat protracted relative to the ability to remember past experiences. An important limitation of the work that has been conducted to date, however, is that memory for these two types of information have not been examined in the same experimental context, thereby precluding the possibility for direct comparison of the data. An additional limitation concerns the duration of time over which infants might remember location-based information relative to memories of past events, given that previous studies have considered delays of seconds to minutes but rarely longer. The present study was designed to determine whether memories for events and memories for locations are similarly maintained over delays of months when assessed using the elicited imitation paradigm.
Method
Participants
Ninety 13-month-old infants (mean age = 13 months, 8 days; range from 12 months, 16 days to 13 months, 21 days), 89 16-month-old infants (mean age = 16 months, 6 days; range from 15 months, 21 days to 16 months, 21 days), and 90 20-month-old infants (mean age = 20 months, 9 days, range from 19 months, 17 days to 20 months, 24 days) were recruited from a participant pool maintained at the University of Minnesota. The pool includes parents who were contacted by mail shortly after the birth of their infants and subsequently returned informational cards indicating their willingness to be contacted about participating in research. All of the participants in the present study were born at term (40 ± 2 weeks) and were experiencing an apparently normal course of development. Infants were primarily of Caucasian descent and were born into families of middle- to upper-middle class socioeconomic status.
Procedure
Infants were scheduled to participate in a large cross-sectional study concerning remembering and forgetting in the second year of life (see Bauer et al., 2000, for details). In addition to participating in the experimental protocol described in Bauer et al., infants completed one one-step elicited imitation task designed to assess memory for events and one one-step elicited imitation task designed to assess memory for locations. The event memory task was completed before the location memory task at the exposure sessions and at delayed recall. The fixed order was required to accommodate the goals of the larger study.
Exposure sessions
A schematic of the testing space is shown in Figure 1. In the event memory task, infants were presented with commercially-available stacking rings and a stuffed rabbit. During the baseline period at the first session, the infants were allowed to interact with the stacking rings and the rabbit until they engaged in repetitive exploratory or off-task behavior (Bauer et al., 2000). The researcher then demonstrated how to make a “silly bunny” by placing the stacking rings on the ears of the stuffed rabbit. The action was modeled one time with narration, such that the researcher told the infant that she could use the provided materials “to make a silly bunny.” Immediately after the demonstration, the infants were encouraged to make a silly bunny as well. The researcher modeled the action at each of the three exposure sessions and the children were allowed the opportunity for immediate imitation each time (mean delay between the first and second sessions = 7 days, range from 4 days to 11 days; mean delay between the second and third sessions = 7 days, range from 3 days to 12 days).
Figure 1.
Schematic of the testing space.
In the location memory task, the infants were seated at a table next to a curtain that obscured a large giraffe decal. During the baseline period at the first session, the researcher observed whether the infants spontaneously looked at or pointed to the curtain under which the giraffe was hidden while seated at the table. The researcher then demonstrated how to find the giraffe decal by sliding the curtain to one side of the rod. The action was modeled one time with narration, such that the researcher told the infant “to find Mr. Giraffe and say hello” as she moved the curtain to the side. The infants were encouraged to look at or point to the decal. The researcher modeled the action at each of the three exposure sessions and the infants were allowed the opportunity for immediate imitation each time.
Delayed recall session
The infants returned to the laboratory for a delayed recall test after between-subjects delays of 1 (mean delay = 31 days, range from 23 to 40 days), 3 (mean delay = 91 days; range from 76 to 121 days), 6 (mean delay = 181 days; range from 162 to 200 days), 9 (mean delay = 272 days; range from 254 to 291 days), or 12 months (mean delay = 366 days; range from 354 to 385 days). Seventeen to 19 participants of each age were tested at each delay. In the event memory task, the infants were given the stacking rings and the stuffed rabbit and were provided with a verbal reminder of the actions that could be completed with them (e.g., “Last time you were here, we used this stuff to make a silly bunny. Show me how you make a silly bunny with this stuff”). The infants were allowed to interact with the stimulus materials until they engaged in repetitive or off-task behavior (see Bauer et al., 2000). After the infants completed the event memory task, they were seated at the table in the presence of the curtain hiding the large giraffe decal and were provided with a verbal reminder of the location memory task (e.g., “Last time you were here, we found Mr. Giraffe and said hello. Show me how you find Mr. Giraffe and say hello”).
Data Reduction
For the location memory task, the first exposure session and the fourth delayed recall session were videotaped for analysis. For the event memory task, the fourth session was videotaped for all participants; the first session was videotaped for 43% of the sample (n = 109 infants), approximately evenly divided among the age groups. Working from the videotapes, two experienced behavioral observers recorded whether each infant completed the target action for the event and location memory tasks at baseline and immediate imitation (Session 1) and at delayed recall (Session 4). Infants received credit for completing the target action for the event memory task if they placed or clearly attempted to place at least one stacking ring on the ears of the stuffed rabbit. They received credit for completing the target action in the location memory task if they looked or pointed towards the curtain hiding the giraffe decal. A third coder independently recoded the tapes for 69 (approximately 27%) of the 251 infants. Mean percent agreement was 90% for the event memory task and 97% for the location memory task. When disagreements occurred, the codes of the primary coders were used for analysis.
Visual inspection of the data revealed that some cells of the design were devoid of variability in the percent of participants who successfully completed each task. For the infants on whom we had baseline data on the event memory task (109 participants), 0% performed the target action. The same pattern was observed on the location memory task at baseline for all of the tested infants. That is 0% of the participants looked towards or pointed at the curtain hiding the giraffe decal. In contrast, at immediate imitation, 100% of the participants performed the target action on the location memory task; possible reasons for the observed high level of performance on this task at immediate imitation are provided in the Discussion. Because of the observed lack of variability in the data and because the dependent measures under consideration were dichotomous categorical variables, the data were analyzed using binomial distributions, which allow both for the inclusion of count data as well as cells that do not contain any variability in the scores.
Results
Four sets of mixed analyses with binomial distributions were conducted using SAS PROC GENMOD to assess the robustness of memory for events and locations both immediately and over the long term. Due to the categorical nature of the dependent variables, the data were transformed to a logit scale for analysis. The values have been re-transformed into the percent of participants who completed each action for ease of interpretation in the figures.
Analyses were conducted to directly compare performance on the event and location memory tasks at immediate imitation and delayed recall; these analyses included between-subjects factors of age and delay as well as one within-subjects factor of condition (event, location). Subsequent analyses were conducted to determine whether the infants successfully completed each target action at immediate imitation relative to baseline and whether their memories for the actions were differentially retained after delays of 1, 3, 6, 9, and 12 months. The original analysis for each type of memory included between-subjects factors of age (13-month-olds, 16-month-olds, 20-month-olds) and delay (1 month, 3 months, 6 months, 9 months, 12 months) as well as one within-subjects factor of phase (baseline, immediate imitation, delayed recall). In order to create the most parsimonious statistical model for each analysis, non-significant interaction terms were successively eliminated from the model until all remaining interactions were significant at p ≤ .15. Significant and marginal main effects, interactions, and follow-up analyses are reported when p ≤ .10. The proportion of infants who successfully completed the target actions as a function of age, phase, delay, and condition are shown in Table 1.
Table 1.
Percent of Infants who Successfully Completed the Target Actions as a Function of Age and Phase of Testing
| Phase | 13-Month-Olds | 16-Month-Olds | 20-Month-Olds | |||
|---|---|---|---|---|---|---|
|
| ||||||
| Event | Location | Event | Location | Event | Location | |
| Baseline | 0% | 0% | 0% | 0% | 0% | 0% |
| Immediate imitation | 60% | 100% | 88% | 100% | 90% | 100% |
| Delayed recall | ||||||
| One month | 65% | 22% | 94% | 61% | 88% | 88% |
| Three months | 31% | 11% | 56% | 29% | 71% | 67% |
| Six months | 29% | 6% | 33% | 6% | 61% | 33% |
| Nine months | 33% | 6% | 20% | 0% | 50% | 22% |
| Twelve months | 33% | 0% | 0% | 6% | 44% | 17% |
Between-Task Analyses
Immediate imitation
The most parsimonious statistical model for performance at immediate imitation included factors of age, delay, and condition. The main effect of age, χ2(2, N = 30) = 6.62, p < .04, indicated that the 13-month-old infants performed the target actions fewer times relative to the 16- and 20-month-olds; performance differences were not apparent between the two older groups of infants (percent successful = 88% of 13-month-olds, 97% of 16-month-olds, and 97% of 20-month-olds). The main effect of condition, χ2(1, N = 30) = 31.40, p < .0001, indicated that a greater number of the infants performed the target action on the location memory task than on the event memory task (percent successful = 100% on the location memory task and 80% on the event memory task). The main effect of delay was not significant or marginal.
Delayed recall
The most parsimonious statistical model for performance at delayed recall included factors of age, delay, condition, and the interaction between age and condition. The main effect of age, χ2(1, N = 30) = 47.87, p < .0001, indicated that a smaller proportion of the 13-month-old infants performed the target actions relative to the 16- and 20-month-old infants and that the 16-month-olds also performed the target action less frequently than the 20-month-olds (percent successful = 24% of 13-month-olds, 34% of 16-month-olds, and 53% of 20-month-olds). The main effect of delay, χ2(4, N = 30) = 82.83, p < .0001, indicated that a greater proportion of the infants successfully performed the target actions after one month than at 3, 6, 9, or 12 months; performance after 3 months exceeded that found at 6, 9, and 12 months. Performance differences were not apparent among any of the lengthier delays (6, 9, or 12 months) (percent successful = 69% at the 1 month delay, 44% at the 3 month delay, 29% at the 6 month delay, 22% at the 9 month delay, and 24% at the 12 month delay). The main effect of condition, χ2(1, N = 30) = 48.78, p < .0001, indicated that a greater proportion of infants performed the target action on the event memory task than on the location memory task (percent successful = 50% on the event memory task and 24% on the location memory task).
In addition to these main effects, a significant Age × Condition interaction also was apparent, χ2(2, N = 30) = 5.95, p = .05; these data are shown in Figure 2. Examination of the interaction by age indicated that a larger percentage of infants of each age performed the target action on the event memory task relative to the location memory task. Analysis of the interaction by condition indicated that on the event memory task a smaller proportion of the 13- and 16-month-old infants performed the target action than the 20-month-olds; differences in the percent of participants who successfully completed the task were not found between the two youngest age groups. On the location memory task, fewer 13-month-old infants performed the target action relative to the 16- and 20-month-olds; a smaller proportion of the 16-month-olds performed the target action than the 20-month-olds.
Figure 2.
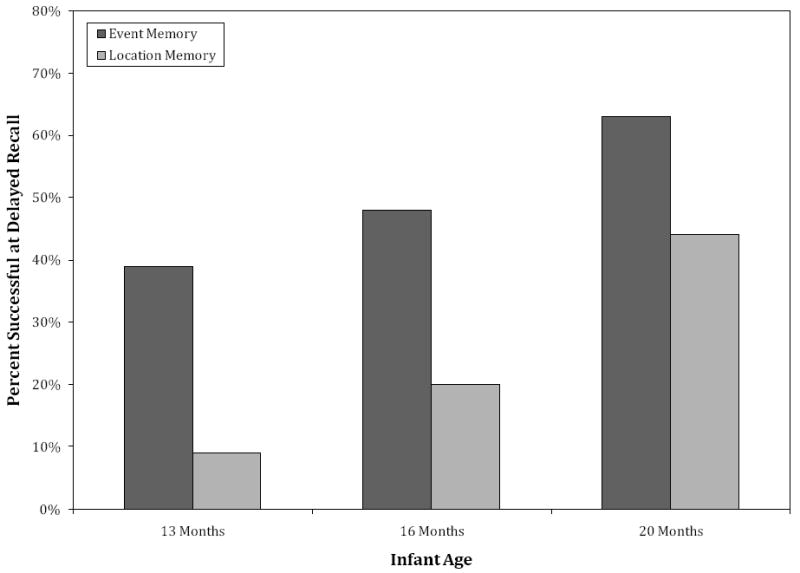
Successful performance on the event and location memory tasks at delayed recall as a function of age and condition.
Within-Task Analyses
Location memory task
The most parsimonious statistical model for performance on the location memory task included factors of age, delay, and phase as well as the interaction between delay and phase and the interaction between age and phase. The main effect of phase, χ2(2, N = 45) = 640.43, p < .0001, indicated that infants performed the target action more often at immediate imitation and delayed recall relative to baseline; significant forgetting was observed from immediate imitation to delayed recall (percent successful = 0% at baseline, 100% at immediate imitation, and 25% at delayed recall). The main effects of age and delay were not significant or marginal.
A significant Age × Phase interaction also was apparent, χ2(4, N = 45) = 11.27, p < .02; these data are shown in Table 2. Examination of the interaction by age indicated that infants of each age performed the target action more often at immediate imitation relative to baseline, but only the 16- and 20-month-old infants exhibited better performance at delayed recall relative to baseline. Analysis of the interaction by phase indicated that age-related differences were found at delayed recall only, such that the 13-month-old infants performed the target action less frequently relative to the 16- and 20-month-olds; performance differences were also apparent between the two older groups, such that 16-month-olds performed the target action less often than 20-month-olds.
Table 2.
Successful Performance on the Location Memory Task as a Function of Age and Phase of Testing
| Phase of Testing | |||
|---|---|---|---|
|
| |||
| Age | Baseline | Immediate Imitation | Delayed Recall |
| 13 months | 0% | 100% | 9% |
| 16 months | 0% | 100% | 20% |
| 20 months | 0% | 100% | 44% |
In addition to the main effects, a marginal Delay × Phase interaction also was apparent, χ2(8, N = 45) = 14.46, p = .07; these data are shown in Table 3. Examination of the interaction by delay indicated that infants more often performed the target action at immediate imitation relative to baseline at all delays and that infants tested after delays of 1 or 3 months also showed better performance at delayed recall relative to baseline. Evidence of forgetting from immediate imitation to delayed recall was apparent at all delays. Analysis of the interaction by phase indicated that performance differences were not apparent at baseline or at immediate imitation as a function of delay. Analyses of performance at delayed recall indicated that infants more often completed the target action after 1 month than at any of the other delays, and that infants more often completed the target action after 3 months than after delays of 6, 9, or 12 months; performance differences were not apparent at any of the lengthier delays.
Table 3.
Successful Performance on the Location Memory Task as a Function of Delay and Phase of Testing
| Phase of Testing | |||
|---|---|---|---|
|
| |||
| Delay | Baseline | Immediate Imitation | Delayed Recall |
| One month | 0% | 100% | 57% |
| Three months | 0% | 100% | 36% |
| Six months | 0% | 100% | 15% |
| Nine months | 0% | 100% | 9% |
| Twelve months | 0% | 100% | 7% |
Event memory task
The most parsimonious statistical model for the event memory task included factors of age, delay, phase, and the interaction between delay and phase. The significant main effect of age, χ2(2, N = 45) = 16.89, p < .0002, indicated that the 13-month-old infants performed the target action less frequently relative to the 16- and 20-month-old infants; performance differences were not apparent between the two older groups (percent successful = 35% of 13-month-olds, 46% of 16-month-olds, and 54% of 20-month-olds). The main effect of phase, χ2(2, N = 45) = 129.37, p < .0001, indicated that infants performed the target action more often at immediate imitation and delayed recall relative to baseline; significant forgetting was also apparent from immediate imitation to delayed recall (percent successful = 0% at baseline, 80% at immediate imitation, and 50% at delayed recall). The main effect of delay was not significant or marginal.
In addition to the main effects, a significant Delay × Phase interaction also was apparent, χ2(8, N = 45) = 16.32, p < .04; these data are shown in Table 4. Examination of the interaction by delay indicated that differences in performance were not found at baseline or immediate imitation. Analyses of performance at delayed recall indicated that infants more often completed the target action after 1 month than at any of the other delays; performance after 3 months also marginally exceeded that found at 9 months. Analysis of the interaction by phase indicated that infants performed the target action more often at immediate imitation and delayed recall relative to baseline at all tested delays, with evidence of forgetting from immediate imitation to delayed recall at all of the delays exceeding 1 month.
Table 4.
Successful Performance on the Event Memory Task as a Function of Delay and Phase of Testing
| Phase of Testing | |||
|---|---|---|---|
|
| |||
| Delay | Baseline | Immediate Imitation | Delayed Recall |
| 1 month | 0% | 76% | 82% |
| 3 months | 0% | 78% | 53% |
| 6 months | 0% | 72% | 43% |
| 9 months | 0% | 89% | 35% |
| 12 months | 0% | 87% | 40% |
Discussion
The present research addressed the question of whether memories for events and locations are similarly maintained over long delays when assessed in the experimental context of elicited imitation. Although there are extensive literatures on both event and location memory in the first years of life and beyond (see Bauer, 2006, for a review of event memory and Newcombe &Huttenlocher, 2006, for a review of location memory), the presented data are the first to our knowledge to compare the retention of both types of information over the long term using the same experimental paradigm. The results indicated greater delay- and age-related variability in the location memory task relative to the event memory task. Specifically, infants remembered the target action for the event memory task at each tested delay relative to baseline, whereas infants showed memory for the location relative to baseline only at the 1- and 3-month delays. The location memory task also revealed the most pronounced age differences at delayed recall, such that 13-month-olds performed less well than 16-month-olds who in turn performed less well than 20-month-olds. Additional analyses of performance at immediate imitation indicated that better retention of the event memory at delayed recall was not due to superior encoding: infants more often performed the target action on the location memory task at immediate imitation relative to the event memory task yet demonstrated superior recall on the event memory task when the analyses were collapsed across delay.
Taken together, these findings provide empirical support for the conclusion that long-term memory for events is more robust than long-term memory for locations in the second year of life. This effect was documented despite allowing all of the infants the opportunity to imitate the actions immediately after their presentation, an experimental manipulation that has been shown to facilitate delayed recall in younger infants (Lukowski et al., 2005). In addition, the differential findings at delayed recall were obtained despite ceiling levels of performance on the location memory task at immediate imitation. The high rate of success on this task likely resulted from assessing location memory using a looking or pointing response, whereas event memory was measured through a specific behavioral response. Despite including this easier response requirement on the location memory task, infants of each age performed better on the event memory task relative to the location memory task at delayed recall. Future research should address the impact of different responses on event and location memories by requiring the same behavioral response for each task.
In addition to equating the response demands for event and location memories, future researchers might also consider comparing performance at delayed recall to novel events tested at the same session. In the present research, we compared the percentage of infants who successfully completed the target actions to the percentage who performed the same actions at baseline.1 Although this procedure is commonly used when the delay between encoding and delayed recall approximates days rather than months (e.g., Wiebe et al., 2006), researchers often compare recall performance after months-long delays to novel events tested at the same session (e.g., Bauer et al., 2000). Use of novel control events as the comparison for familiar sequences tested at delayed recall equates trial-and-error problem solving abilities that may have developed over time, thereby resulting in a more stringent assessment of memory. Although significant maturation certainly occurred over the lengthy delays imposed in the present research, we argue that the inclusion of novel events at delayed recall was not critically necessary due to our primary goal of comparing long-term memory for events and locations. In addition, the forgetting function for the event memory task resembles that observed for the lengthier sequences in Bauer et al. (2000), in which novel control sequences were used as the comparison. As a result, we have comparison of the event memory data from an analogous task conducted with the same infants at the same sessions.
Although the present research was not designed to address the mechanism responsible for the obtained pattern of effects, one possibility is that the neural structures that support long-term memory of locations may develop somewhat more slowly relative to those subserving memory for events (Sluzenski et al., 2004). The ability to remember past experiences over the long term can be expected to facilitate learning new conceptual information, particularly vocabulary words or the functions of novel items encountered in the environment. We may speculate that months-long memory for locations would become necessary later in ontogeny, when infants gain greater independence and are more likely to explore the environment outside of the watchful eye of their parents. Nevertheless, additional research is needed to determine whether differences in neurological development are the primary mechanism responsible for the observed differential encoding and long-term retention of information related to events and locations as well as to identify the functional implications of better long-term memory for events relative to locations during the second year of life.
Acknowledgments
Data collection for this research was completed while the third author was at the Institute of Child Development, University of Minnesota. Data collection was supported by grants from the NICHD (HD28425, HD42483) to Patricia J. Bauer. Preparation for the manuscript was supported by the University of California, Irvine and Emory University. We are grateful to the participants and their families who gave so generously of their time to participate in this research and to members of the Cognition in the Transition Laboratory for their assistance with data collection and coding.
Footnotes
As mentioned in the Methods section, the baseline data were missing for 57% of the sample on the event memory task. The data were recorded for 109 infants, however, and these data were approximately evenly distributed among the three age groups. Given that baseline performance was uniform for the infants on whom data were available, we are confident that the findings would not have been materially different had the full complement of data been available.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Barr R, Dowden A, Hayne H. Developmental changes in deferred imitation by 6- to 24-month-olds. Infant Behavior and Development. 1996;19:159–170. doi: 10.1016/s0163-6383(96)90015-6. [DOI] [Google Scholar]
- Bauer PJ. Event memory. In: Damon W, Lerner RM, Kuhn D, Siegler RS, editors. Handbook of child psychology: Cognition, perception, and language. 6. Vol. 2. New York: Wiley; 2006. pp. 373–425. [Google Scholar]
- Bauer PJ, Mandler JM. One thing follows another: Effects of temporal structure on one- to two-year-olds’ recall of events. Developmental Psychology. 1989;25:197–206. [Google Scholar]
- Bauer PJ, Wiebe SA, Waters JM, Bangston SK. Reexposure breeds recall: Effects of experience on 9-month-olds’ ordered recall. Journal of Experimental Child Psychology. 2001;80:174–200. doi: 10.1006/jecp.2000.2628. [DOI] [PubMed] [Google Scholar]
- Bauer PJ, Wiebe SA, Carver LJ, Waters JM, Nelson CA. Developments in long-term explicit memory late in the first year of life: Behavioral and eletrophysiological indices. Psychological Science. 2003;14:629–635. doi: 10.1046/j.0956-7976.2003.psci_1476.x. [DOI] [PubMed] [Google Scholar]
- Bauer PJ, Wenner JA, Dropik PL, Wewerka SS. Parameters of remembering and forgetting in the transition from infancy to early childhood. Monographs of the Society for Research in Child Development. 2000;65:v–204. [PubMed] [Google Scholar]
- Carver LJ, Bauer PJ. When the event is more than the sum of its parts: 9-month-olds’ long-term ordered recall. Memory. 1999;7:147–174. doi: 10.1080/741944070. [DOI] [PubMed] [Google Scholar]
- Carver LJ, Bauer PJ. The dawning of a past: The emergence of long-term explicit memory in infancy. Journal of Experimental Child Psychology: General. 2001:726–745. doi: 10.1037/0096-3445/130/4/726. [DOI] [PubMed] [Google Scholar]
- Learmonth AE, Newcombe NS, Huttenlocher J. Toddlers’ use of metric information and landmarks to reorient. Journal of Experimental Child Psychology. 2001;80:225–244. doi: 10.1006/jecp.2001.2635. [DOI] [PubMed] [Google Scholar]
- Learmonth AE, Newcombe NS, Sheridan N, Jones M. Why size counts: Children’s spatial reorientation in large and small enclosures. Developmental Science. 2008;11:414–426. doi: 10.1111/j.1467-7687.2008.00686.x. [DOI] [PubMed] [Google Scholar]
- Newcombe N, Huttenlocher J. Development of spatial cognition. In: Damon W, Lerner RM, Kuhn D, Siegler RS, editors. Handbook of child psychology: Cognition, perception, and language. 6. Vol. 2. New York: Wiley; 2006. pp. 734–776. [Google Scholar]
- Newcombe N, Huttenlocher J, Learmonth E. Infants’ coding of location in continuous space. Infant Behavior and Development. 1999;22:483–510. doi: 10.1016/s0163-6383(00)000114. [DOI] [Google Scholar]
- Sluzenski J, Newcombe NS, Satlow E. Knowing where things are in the second year of life: Implications for hippocampal development. Journal of Cognitive Neuroscience. 2004;16:1443–1451. doi: 10.1162/0898929042304804. [DOI] [PubMed] [Google Scholar]
- Wiebe SA, Cheatham CL, Lukowski AF, Haight JC, Muehleck AM, Bauer PJ. Infants’ ERP responses to novel and familiar stimuli change over time: Implications for novelty detection and memory. Infancy. 2006;9:21–44. doi: 10.1207/s15327078in0901_2. [DOI] [Google Scholar]