

Abstract
Colorectal cancer (CRC) is one of the most prevalent malignant cancer types worldwide. Although the purine metabolism pathway is vital for cancer cell survival, little is known about the role of equilibrative nucleoside transporter 2 (ENT2) in CRC development and its association with purine metabolites. The aim of the present study was to evaluate the levels of hypoxanthine phosphoribosyl transferase (HPRT), hypoxanthine and uric acid (UA), as well as xanthine oxidase (XO) activity, and investigate their association with ENT2 expression levels in a normal human colon cell line and CRC cell lines derived from different stages of CRC. These analyses were performed using the normal colon CCD-841CoN cell line and a panel of human CRC cell lines comprising SW480, HCT15 and HCT116, which represent Dukes' B, C and D stages, respectively. Reverse transcription-quantitative PCR was performed to determine the level of ENT2 mRNA expression. In cells of all CRC stages, the levels of HPRT and hypoxanthine were significantly higher (P<0.05), while XO activity and UA levels were significantly decreased (P<0.05), compared with those in the CCD-841CoN cell line. ENT2 expression was found to be elevated in cells derived from all stages of CRC. The Dukes' D stage cell line had higher levels of HPRT and hypoxanthine, although its ENT2 level was not significantly lower than that of the Dukes' B and C stage cell lines. Increased levels of HPRT and hypoxanthine in various stages of CRC may indicate an increase in the activity of the salvage pathway. The increased expression of ENT2 implies the importance of the ENT2 protein in facilitating hypoxanthine transport, which is required for enhanced DNA synthesis via hypoxanthine recycling. In conclusion, ENT2 may have potential as a target in the development of CRC therapeutics.
Keywords: CRC, ENT2, hypoxanthine, hypoxanthine phosphoribosyl transferase, XO
Introduction
Globally, colorectal cancer (CRC) is the third most common malignant cancer in terms of incidence and the second in terms of mortality (1). In 2018, ~10% of all new cases of cancer and cancer-associated mortalities were recorded as being due to CRC (2). The Dukes' staging system classifies CRC into four groups known as Dukes' A, B, C and D, which represent stages I, II, III and IV, respectively (3). CRC develops through a multistep process, including morphological, histological and genetic alterations that accumulate over time (4). Furthermore, metabolic reprogramming allows cancer cells to maintain redox homeostasis while increasing ATP production to meet their high energy demand (5).
Recently, purine metabolism has become a topic of interest in CRC research, and alteration of the purine metabolism pathway, including the modification of enzyme activities, has been shown to be associated with disease progression (6). Purine metabolites supply cells with vital energy and cofactors to enhance cell survival and proliferation (7). Purine nucleosides (purine metabolites), such as adenosine, guanine and inosine, function as signaling molecules (8) and precursors to ATP and GTP nucleotides during DNA and RNA synthesis (7). An insufficient supply of nucleotides during DNA replication may reduce the rate of cell replication, eventually triggering DNA damage and genomic instability (9).
Purine levels are maintained by the de novo and salvage biosynthetic pathways, whereby under normal physiological conditions, degraded bases are recycled via a salvage pathway to sustain the cellular purine pool (10). Moreover, existing bases in the extracellular matrix can be transported into the cells to generate nucleotides (7). Conversely, high purine levels are required in the environment of cancer cells, which necessitates upregulation of the de novo biosynthetic pathway (7). Previous research has indicated that the treatment of cancer cells with purine anti-metabolites blocks DNA synthesis in the cells, thereby inhibiting their growth (11,12).
Hydrophilic purine nucleosides require specific membrane transport proteins known as nucleoside transporters (NTs) to transport them across plasma membranes. NTs are classified according to their transport mechanisms as Na+-dependent concentrative transporters, which are encoded by the solute carrier family 28 (SLC28) gene, and equilibrative NTs (ENTs), which are encoded by the SLC29 gene (13). The ENTs are bidirectional transporter proteins that facilitate the movement of nucleosides across the cell membrane driven solely via the concentration gradient between nucleoside permeants inside and outside the cell (14). The ENT2 protein is able to transport nucleosides and a wide range of purine and pyrimidine nucleobases. Although the exact function of ENT2 is unclear, its capacity and affinity for the translocation of hypoxanthine has been reported to be higher than that of ENT1 (15,16).
Hypoxanthine is a purine substrate in the salvage pathway for nucleotide synthesis, as well as the first metabolite in the purine catabolism process (17). Consequently, any interruption in the expression or activity of ENT2 could be deleterious to the cells, as the intracellular accumulation of reactive oxygen species (ROS), a by-product of the purine catabolism pathway, might disrupt the redox balance and ultimately kill the cancer cells (18). Furthermore, ENT2 and other NTs play crucial roles in the clearance from the body of certain anticancer nucleoside analogs, such as 5-fluorouracil, which is widely used as a chemotherapeutic drug for the treatment of gastrointestinal cancer, particularly CRC (19), and antiviral drugs, such as remdesivir, which is used as a treatment for COVID-19(20). The current study therefore aimed to compare the levels of hypoxanthine phosphoribosyl transferase (HPRT), hypoxanthine/xanthine and uric acid (UA), as well as the activity of xanthine oxidase (XO) in CRC cell lines and a normal colorectal cell line. It may be noted that enzyme activity measurements are typically used to estimate the concentration of enzyme present in a sample under certain conditions (21). In addition, the levels of ENT2 mRNA expression were evaluated and compared in cells derived from CRC of different stages.
Materials and methods
Cell culture
All cell lines were purchased from the American Type Culture Collection (ATCC). A panel of human CRC cell lines was used, where each cell line represented a different CRC stage, as well as a normal colon epithelial cell line. CCD-841CoN (cat. no. CRL-1790) was used as the normal colon epithelial cell line, and SW480 (cat. no. CCL-228; Dukes' B), HCT15 (cat. no. CCL-225; Dukes' C) and HCT116 (cat. no. CCL-247; Dukes' D) were used as the CRC cell lines. These CRC stages were classified according to the tumor stage following Dukes' classification criteria by the ATCC and previous studies (22,23). All cells were cultured in Dulbecco's modified Eagle's medium (high glucose; Nacalai Tesque, Inc.) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1% penicillin-streptomycin (MilliporeSigma). The cells were incubated at 37˚C in a humidified incubator (BINDER GmbH) with 5% CO2.
Cell lysate preparation
Following cell quantification, 1x106 cells were resuspended in 100 µl appropriate buffer as provided with the kit used for each assay. The cells were hen homogenized using an Ultrasonic Vibra-Cell™ homogenizer (Sonics & Materials, Inc.) with pulses of 5 sec on and 25 sec off for 10 cycles. The tube containing the cell lysate was homogenized in ice to reduce the heat produced by the ultrasonic treatment. Tubes containing cell lysate and debris were then centrifuged at 24,104 x g for 10 min at 4˚C. The supernatants were transferred to new tubes and kept at -20˚C until required for further use.
HPRT enzyme-linked immunosorbent assay (ELISA) assay
The HPRT level in the cells was measured using a sandwich ELISA kit for HPRT (cat. no. SEA717Hu; Cloud-Clone Corp.) according to the manufacturer's instructions. The cells were lysed as aforementioned. The detection method is based on the colorimetric reaction of horseradish peroxidase following the addition of a tetramethylbenzidine substrate. The absorbance was measured immediately at 450 nm using a microplate reader (BMG Labtech GmbH).
XO assay
The activity of XO was quantified spectrophotometrically using a Xanthine Oxidase Activity Colorimetric/Fluorimetric Assay Kit (cat. no. K710; BioVision, Inc.; Abcam). In this assay, XO in the sample oxidizes xanthine to hydrogen peroxide (H2O2), which reacts stoichiometrically with OxiRed Probe to generate a color that can be detected at 570 nm. Briefly, 44 µl assay buffer, 2 µl enzyme mix, 2 µl substrate mix and 2 µl OxiRed probe were mixed and added to the designated wells. The reactions were initiated by the addition of 50 µl prepared standard or cell lysate into each well. Absorbance (A) measurements A1 and A2 were taken after 20 min incubation in the dark at room temperature using a microplate reader (BMG Labtech GmbH) at a wavelength of 570 nm. A standard curve was plotted for several dilutions of H2O2 standard and the volume was adjusted to 50 µl/well using distilled water. The XO activity of each sample was calculated using the following equation: XO activity (mU/ml)=(B x dilution factor)/(T2-T1) x V, in which B is the amount of H2O2 generated by XO according to the standard curve (nmol), T1 is the time of the first reading (A1, min), T2 is the time of the second reading (A2, min) and V is the pre-treated sample volume added to the reaction well (ml).
Hypoxanthine/xanthine assay
The hypoxanthine/xanthine level in all cell lines was detected using a Xanthine/Hypoxanthine Colorimetric/Fluorimetric Assay Kit (cat. no. K685; BioVision, Inc.; Abcam) according to the manufacturer's instructions. In brief, 1x106 cells were incubated on ice with 100 µl ice-cold xanthine assay buffer for 10 min. After centrifugation at 24,104 x g for 5 min, the supernatant was collected and incubated with the reaction mix for 30 min at room temperature. A standard curve was plotted using several dilutions of xanthine standard and the volume was adjusted to 50 µl/well with xanthine assay buffer. The absorbance was measured at a wavelength of 570 nm.
UA assay
UA quantification was performed using a Uric Acid Colorimetric/Fluorometric Assay Kit (cat. no. K608; BioVision, Inc. Abcam) according to the manufacturer's instructions. Briefly, 46 µl UA assay buffer, 2 µl UA enzyme mix and 2 µl UA probe were mixed and added to the designated wells on the plate. The reactions were initiated by the addition of 50 µl prepared UA standard or cell lysate to each well. The plates were then incubated for 30 min at 37˚C and protected from light. A standard curve was plotted using several dilutions of UA standard and the volume was adjusted to 50 µl/well with UA assay buffer. The absorbance was measured at a wavelength of 570 nm.
RNA isolation and cDNA synthesis
Total RNA from all cell lines was extracted using a NucleoSpin RNA mini kit for RNA purification (cat. no. 740955; Macherey-Nagel, GmbH & Co. KG) according to the manufacturer's instructions. The total RNA concentration and the A230/208 and A260/280 nm ratios of the initial RNA were measured to assess the RNA quality using a SpectraMax® QuickDrop™ UV-Vis Spectrophotometer (Molecular Devices, LLC). RNA integrity was assessed via a gel electrophoresis assay. Next, 200 ng total RNA was used for reverse transcription (RT) using a SensiFAST™ cDNA Synthesis kit (cat. no. BIO-65053l Bioline; Meridian Bioscience, Inc.) according to the manufacturer's instructions.
Quantitative PCR (qPCR) of ENT2
Following RT, qPCR was performed using a CFX96™ Touch Real-Time PCR detection system (Bio-Rad Laboratories, Inc.). The reactions were carried out in triplicate using a SensiFAST™ SYBR® & Fluorescein Kit (cat. no. BIO96005; Bioline; Meridian Bioscience, Inc.) following the manufacturer's instructions. The sequences of the oligonucleotide primers that were used for qPCR amplification are listed in Table I. Each reaction was performed in a final volume of 10 µl and the primer concentration was 400 nM. Thermocycling was conducted with an initial start cycle at 95˚C for 2 min, followed by 39 cycles at 95˚C for 5 sec and 60˚C for 30 sec. To confirm product specificity, melting curve analysis was performed after each amplification. As normalization against a single reference gene may not be considered adequate (24), the mRNA level of ENT2 was normalized by the geometric averaging of two reference genes, GAPDH and HPRT, to obtain the most reliable results in the gene transcription analysis. The stability and reliability of GAPDH and HPRT1 as reference genes in the CRC and normal colon cell lines were determined via measurement of their quantification cycle (Cq) value and calculating the coefficient of variation using CFX96 software (Bio-Rad Laboratories, Inc.). The fold-change in ENT2 expression of the human CRC cell lines relative to the normal colon cell line was determined using the 2-ΔΔCq method (25). The geometric average (GeoMean) of the relative quantities [(EREF)∆Cq REF] of the two reference genes was utilized to determine the expression of the gene of interest (GOI) as shown in the following equation: Relative gene expression=[(EGOI)ΔCq GOI]/GeoMean[(EREF)ΔCq REF].
Table I.
Primer sequences used for quantitative PCR.
| Gene | Forward (5'-3') | Reverse (5'-3') |
|---|---|---|
| ENT2 | CCACTCTCTCACCGAAGCCTAA | GCAGGAAGAACAGCACCAACA |
| GAPDH | GCATCCTGGGCTACACTGAG | TCCTCTTGTGCTCTTGCTGG |
| HPRT1 | GAGTCCTATTGACATCGCCAGT | TCCGCCCAAAGGGAACTGAT |
ENT2, equilibrative nucleoside transporter 2; HPRT, hypoxanthine phosphoribosyl transferase.
Statistical analysis
All data were statistically analyzed using IBM SPSS Statistical Software version 27 (IBM Corp.). The results are expressed as the mean ± SD obtained from three independent experiments where each assay was performed in triplicate. The significant differences between multiple groups were assessed by one-way analysis of variance (ANOVA) followed by Tukey's post hoc multiple comparisons tests. Statistical evaluation of ENT2 gene expression was performed using CFX96 software followed by one-way ANOVA with Tukey's multiple comparisons tests. The correlations between ENT2 expression and the different stages of CRC were determined by Spearman's correlation test. P<0.05 was considered to indicate a statistically significant difference.
Results
Determination of HPRT levels
HPRT levels in the normal colon and CRC cell lines were determined by the binding of HPRT to an antibody in an ELISA experiment. When compared with normal colon cells, the levels of HPRT in the SW480 (P=0.0001), HCT15 (P=0.013) and HCT116 (P<0.0001) cell lines were significantly higher. HPRT levels in the cell lines representing Dukes' stages B and D CRC were significantly higher compared with that in the cell line representing Dukes' stage C (P=0.0079 and P=0.0003, respectively; Fig. 1).
Figure 1.
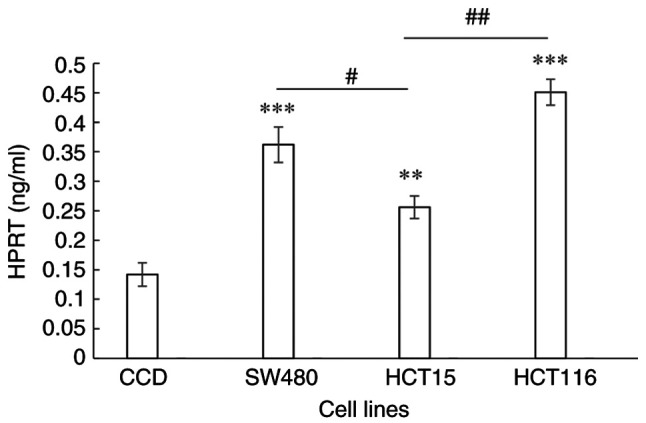
HPRT level in a normal colon epithelial cell line and the colorectal cancer SW480, HCT15 and HCT116 cell lines, which are derived from Dukes' B, C and D stage cancer, respectively. Data are presented as the mean ± SD (n=9). Statistical analysis was performed using one-way analysis of variance followed by Tukey's post hoc tests. **P≤0.001, ***P≤0.0001 vs. CCD; #P≤0.05, ##P≤0.001. HPRT, hypoxanthine phosphoribosyl transferase; CCD, CCD-841CoN normal colon epithelial cells.
Determination of XO activity
The XO enzymatic activity in the CRC cell lines representing the different stages of CRC were all significantly lower (all P<0.0001) than those in the normal colon cells (Fig. 2).
Figure 2.
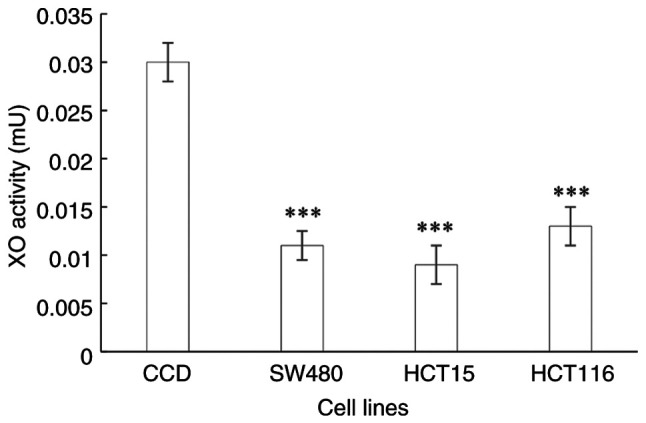
XO activity in a normal colon epithelial cell line and the colorectal cancer SW480, HCT15 and HCT116 cell lines, which are derived from Dukes' B, C and D stage cancer, respectively. Data are presented as the mean ± SD (n=9). Statistical analysis was performed using one-way analysis of variance followed by Tukey's post hoc test. ***P≤0.0001 vs. CCD. XO, xanthine oxidase; CCD, CCD-841CoN normal colon epithelial cells.
Determination of hypoxanthine/xanthine level
The hypoxanthine/xanthine levels of the Dukes' B, C and D stage CRC cell lines were significantly higher compared with those in the normal colon cells (P=0.007, P=0.004 and P<0.0001, respectively; Fig. 3). The hypoxanthine/xanthine levels ranged from 2.86 to 5.08 µM in the CRC cell lines compared with 0.92 µM in the normal colon cells. The hypoxanthine/xanthine levels were observed to increase as the CRC stage advanced, with 3.1-, 4.2- and 5.5-fold increases in Dukes' B, C and D stages, respectively, compared with the normal colon control. The hypoxanthine/xanthine level was significantly higher in the Dukes' D CRC cell line than that in the Dukes' B cell line (P=0.003).
Figure 3.
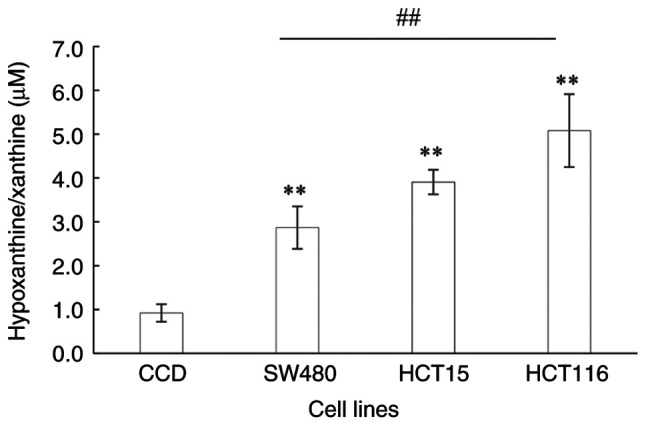
Level of hypoxanthine/xanthine in a normal colon epithelial cell line and the colorectal cancer SW480, HCT15 and HCT116 cell lines, which are derived from Dukes' B, C and D stage cancer, respectively. Data are presented as the mean ± SD (n=9). Statistical analysis was performed using one-way analysis of variance followed by Tukey's post hoc test. **P≤0.001; ##P≤0.001. CCD, CCD-841CoN normal colon epithelial cells.
Determination of UA level
The end product of purine catabolism is UA. When compared with that in the normal colon cells, the UA level in the HCT15 and HT116 cell lines was significantly downregulated (P<0.0001 and P=0.0002, respectively; Fig. 4). Furthermore, UA levels were significantly decreased in the CRC cell lines representing Dukes' C and D stages compared with those in the Dukes' B CRC cell line (P<0.0001 and P=0.0008, respectively).
Figure 4.
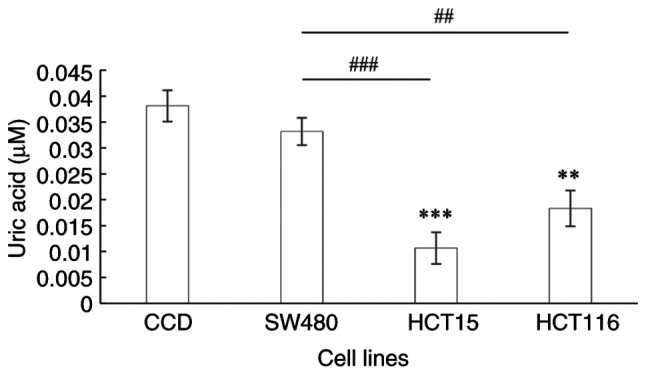
Uric acid level in a normal colon epithelial cell line and the colorectal cancer SW480, HCT15 and HCT116 cell lines, which are derived from Dukes' B, C and D stage cancer, respectively. Data are presented as the mean ± SD (n=9). Statistical analysis was performed using one-way analysis of variance followed by Tukey's post hoc test. **P≤0.001, ***P≤0.0001 vs. CCD; ##P≤0.001, ###P≤0.0001 between CRC stages . CCD, CCD-841CoN normal colon epithelial cells.
ENT2 gene expression
ENT2 was significantly upregulated in all CRC cell lines when compared with that in the normal colon cells (P<0.0001; Fig. 5). The ENT2 expression level was found to be 186-, 471- and 123-fold higher in cells from the Dukes' B, C and D stages of CRC, respectively, compared with that in the normal colon cell line. The ENT2 expression level was the highest in the Dukes' C stage CRC cells. Notably, ENT2 expression was lower in the cells of Dukes' stage D compared with those of Dukes' stage B, however this was insignificant. However, Spearman's correlation analysis for ENT2 expression and CRC stage did not reveal a significant correlation (ρ=0.475; P=0.128; data not shown).
Figure 5.
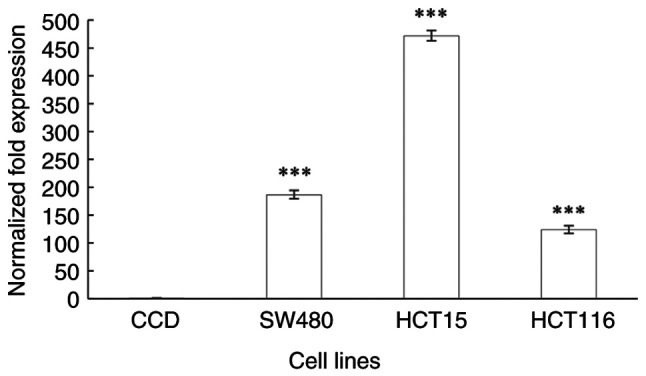
Differential expression of the equilibrative nucleoside transporter 2 gene (NM_001300868.1) in a normal colon epithelial cell line and the colorectal cancer SW480, HCT15 and HCT116 cell lines, which are derived from Dukes' B, C and D stage cancer, respectively. GAPDH and HPRT1 were used as the reference genes. Data are presented as the mean ± SD (n=9). Statistical differences were examined using one-way analysis of variance followed by Tukey's post hoc test. ***P≤0.0001 vs. CCD. CCD, CCD-841CoN normal colon epithelial cells.
Discussion
Purine metabolism has been observed to be one of the most affected pathways in CRC (26,27) and other cancer types, including breast cancer (28) and urothelial carcinoma (29). Moreover, alterations in its enzymatic pattern (30) and purine metabolism dysfunction have been found to be associated with cancer progression. For example, several cancer types have been demonstrated to express a high level of HPRT, the key enzyme in the salvage pathway, thus promoting metabolic flux to produce purine mononucleotides, which are precursors in nucleic acid synthesis (31). While the hypoxanthine concentration gradient alone may not provide a compelling argument for the involvement of ENT2 in hypoxanthine transport, as hypoxanthine influx/efflux was not observed in the present study, the molecules and substrates involved in the intracellular purine metabolic pathway, including HPRT level, XO activity and UA level, were examined. The present study recorded a significant upregulation of HPRT and hypoxanthine/xanthine levels in different stages of CRC, notably in advanced stages, whereas the XO activities and UA levels in the CRC cell lines were markedly decreased compared with those in the normal colon cells.
The upregulation of HPRT in several solid cancer types, including breast, lung, colon and prostate cancer (32), the significantly higher HPRT activities in colorectal (33) and breast (34) cancer, and a notable increase in HPRT activity with the clinical progression of human colon carcinoma (35) have been documented, which support the findings of the current study. A metabolomic study reported elevated hypoxanthine levels in lymphoblastic leukemia and lung and prostate cancer (36), whereas under normal physiological conditions, hypoxanthine levels are very low (37). Similarly, another study recorded elevated hypoxanthine and decreased UA levels in CRC tissues (38). The data in the present study suggest that increased hypoxanthine levels are associated with CRC progression. However, further investigation into the association between hypoxanthine elevation and cancer progression in vivo is required to verify this.
Consistent with the findings of the present study, several investigations have reported diminished XO activity in cancer types such as colon (39), kidney and bladder (40) cancer. Low XO expression has been shown to be associated with more aggressive ovarian cancer (41), indicating that it plays a vital role in cancer progression and could be a risk factor (42). Furthermore, the reduction in XO activity may benefit cancer cells by lowering the generation of ROS, which can trigger apoptosis and cell death. It has been demonstrated that reduced XO activity is advantageous to cancer cells, as it attenuates the detrimental effect of ROS generation on hypoxanthine degradation and catabolic pathway activation (43). Another parameter examined in the present study is UA levels. Previous studies have recorded elevated UA levels in advanced stages of rectal cancer (44) and head and neck carcinoma (45). Conversely, the current study recorded significantly decreased UA levels in the CRC cells of different stages compared with those in normal colon cells. The decrease in UA levels may be due to the low XO activity in all CRC stages evaluated.
The role of UA in tumorigenesis has not yet been determined. Several investigations have suggested that an elevated serum level of UA (hyperuricemia) is associated with an increased cancer risk due to its pro-inflammatory properties (46). It has also been suggested that UA plays an essential role as a potent antioxidant (31). Moreover, elevated UA levels might serve as a tumor defense, by protecting the tumor cells against free-radical oxidative damage by acting as antioxidant and by stimulating the immune system (47). Elevated UA levels have been shown to be associated with an increased survival time in patients with CRC (48). It is notable that the investigated metabolites and enzyme levels in the present study did not exhibit a strict association with Dukes' stage. This may be due to different mutational statuses in the different stages (49). Therefore, the rate of purine metabolism may not increase with higher staging. Advanced stages of cancer may have alternative cancer-protective/resistance mechanisms that are not limited to the purine metabolism pathway. Cells are particularly adaptable to intrinsic oxidative stress in the advanced stages of cancer, as they develop a strong ability to maintain the homeostatic balance between the generation and elimination of ROS (50-52).
The current study found that ENT2 expression was significantly higher in cells from all stages of CRC than that in normal colon cells. Similarly, in previous studies, the level of ENT2 was observed to be 2- to 5.5-fold higher in breast, kidney and prostate cancer cells than in corresponding normal cells (18,53), with the highest expression of ENT2 detected in cancer derived from digestive organ tissues (54). ENT2 has been reported to be highly expressed in colorectal biopsy samples (55), and to be highly expressed in four metastatic cell lines, namely LoVo, Colo205, SK-CO-1 and T84, and four primary CRC cell lines, namely HT29, Caco2, Colo320 and HCT116(56). Furthermore, high levels of ENT2 gene expression have been associated with the advanced stages of a variety of malignancies, including hepatocellular carcinoma, mantle cell lymphoma and ovarian carcinoma (57).
In the present study, ENT2 expression was significantly higher in CRC cell lines that representing Dukes' B, C, and D compared with that in normal colon cells, but was not significantly lower in Dukes' D stage cell line (stage IV) than in the Dukes' B and C stage cell lines (stage II and III)This suggests that the expression of ENT2 is not associated with CRC stage. This may imply that cancer cells, particularly in advanced stages, regulate ENT2 expression for protection and survival. However, the mechanism of action of ENT2, specifically on nucleoside permeant influx/efflux, requires further exploration via in vivo studies. ENT2 is a bidirectional transporter that has the ability to translocate a wide range of nucleosides into and out of cells; it has been documented that ENT2 is the main transporter responsible for cellular hypoxanthine efflux (58). In the current study, the low expression of ENT2 in Dukes' D stage cells corresponded to the highest level of hypoxanthine, an endogenous substrate of ENT2 (31). Increased ENT2 expression may allow cancer cells to escape from the damaging effects of ROS on hypoxanthine degradation, while lowered ENT2 expression may limit hypoxanthine efflux, in a process whereby hypoxanthine recycling supplies nucleotides to cancer cells to meet the demand for rapid cell proliferation.
During salvage purine metabolism, hypoxanthine is recycled to inosine monophosphate (IMP) by HPRT or synthesized through the deamination of adenosine by adenosine deaminase to produce inosine, which is converted to hypoxanthine by purine nucleoside phosphorylase (33). Subsequently, hypoxanthine is oxidized to produce xanthine via the purine catabolism pathway and then converted to UA via XO. During the catabolism pathway, ROS are produced as a by-product (42).
Studies have shown that intrinsic oxidative stress, such as ROS-induced stress, can effectively kill tumor cells via the activation of apoptotic processes (59-62). It has been observed that when ROS accumulation reaches a specific threshold that is incompatible with cell survival, cytotoxic effects are exerted, which induce apoptosis and slow cancer growth (63). ROS generation mediated by XO or NADH dehydrogenase has been linked with mitochondrial malfunction, which is considered to be the first indication of apoptotic intrinsic pathway activation (64). Excess hypoxanthine in cancer cells is then transported out of the cancer cells via the ENT2 transporter (58) to maintain low to average levels of ROS, or recycled to generate new nucleotides for DNA synthesis in order to meet the demand for rapid cell proliferation. Cancer cells may alter their metabolic regulation processes, in order to avoid the harmful effects of ROS and allow the cells to adapt and thrive in a hostile environment (38). These findings may explain the contradictory data observed for hypoxanthine levels in the serum of patients with CRC (6) and in CRC cells (65), particularly in advanced stages (27).
The upregulated HPRT and hypoxanthine levels reported in the CRC cells in the present study may imply that the activity of the salvage pathway is increased to meet the demand for rapid cell proliferation via the generation of nucleotides for DNA synthesis by hypoxanthine recycling, as previously proposed (31). Furthermore, the activity of the catabolism pathway may be reduced, as suggested by the decreased XO and UA levels, in order to preserve the antioxidant balance and avoid ROS-induced damage, which would lead to apoptosis (Fig. 6). Changes in the levels of enzymes and metabolites associated with the purine salvage and catabolism pathways may have diagnostic and prognostic importance for CRC progression. Furthermore, ENT2 may play a crucial role in the regulation of metabolite transport to enhance cancer cell survival and proliferation.
Figure 6.
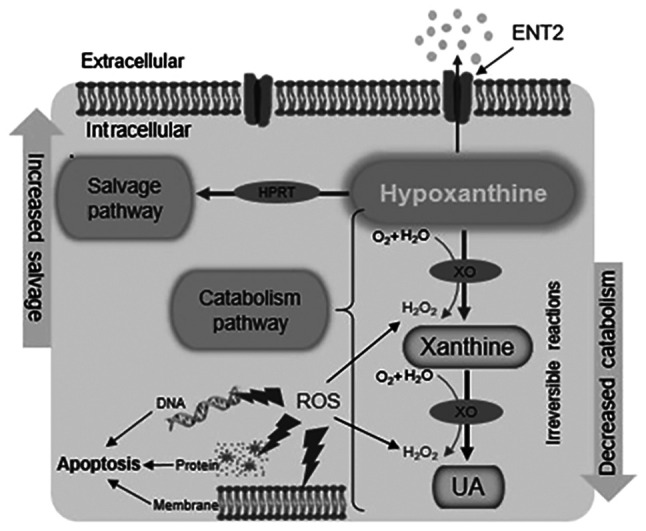
Schematic diagram of purine salvage and catabolism pathways in CRC cells. Modification of purine metabolism in CRC cells affects two processes: The activity of the salvage pathway is increased to meet the demands of rapid cell proliferation by the generation of new nucleotides for DNA synthesis via hypoxanthine recycling, and the activity of the catabolism pathway is decreased in order to avoid the detrimental effects of ROS. CRC, colorectal cancer; ROS, reactive oxygen species; ENT2, equilibrative nucleoside transporter 2; HPRT, hypoxanthine phosphoribosyl transferase; UA, uric acid; XO, xanthine oxidase.
Since hypoxanthine influx/efflux was not observed in the present study, the role of ENT2 in hypoxanthine transport and the hypoxanthine concentration gradient has not been clearly demonstrated. However, various molecules and substrates involved in the purine metabolic pathway were examined, including XO activity, and HPRT and UA levels, which act intracellularly.
Increased ENT2 expression in cells of higher Dukes' stages suggests that hypoxanthine may be being transported out of the cells as a form of defense, resulting in fewer substrates being available for the purine metabolism pathway. Evidence for this is provided by the inverse correlation of hypoxanthine levels with XO activity and UA levels. In this case, the degradation pathway is unfavorable to cells, and extracellular transport avoids the generation of high levels of ROS, which are toxic to cancer cells, and the antioxidant UA.
Notably, the level of HPRT was found to be upregulated in the CRC cell lines of the present study, which could imply that the hypoxanthine available inside the cells is converted to IMP via the salvage pathway to produce the nucleosides GMP and AMP for conversion to the nucleotides required for DNA synthesis and cell growth. However, further investigation is required for a definitive conclusion on the latter process. An in vivo experiment, which would give tangible support for the findings, is lacking and is a limitation of the present study; therefore, it could be the subject of future research.
In conclusion, CRC cells may utilize ENT2 in dual roles for protection and survival, that is, for avoiding the detrimental effects of ROS accumulation via purine catabolic metabolism, as demonstrated by decreased XO enzyme and UA production, and for utilizing hypoxanthine to meet the energy demands of rapid cell proliferation via the purine salvage pathway, as indicated by increased HPRT levels. Accordingly, the purine metabolism pathway appears to be an important metabolic pathway in the progression of CRC. However, the mechanism of action, in particular, whether additional pathways are affected by aberrant ENT2 expression, requires further investigation.
In the current study, HPRT, XO, hypoxanthine and UA levels, and ENT2 expression, were detected in a panel of CRC cell lines. However, further investigations such as measuring hypoxanthine influx/efflux and its association with low and high staging, assessment of protein expression levels by western blotting, performing ENT2 gene silencing and detecting the ROS level in different stages of colorectal carcinogenesis are recommended to determine the relevance and role of ENT2, and its association with purine metabolism in colorectal carcinogenesis. However, although the metabolomic analysis of cell lines has been extensively used in disease research due to certain advantages, including lower costs, ease of control and simpler data interpretation than the analysis of in vivo models using human tissues and bio-fluids (66), in vivo experiments are necessary to provide concrete support to the findings and a solid conclusion. Despite these limitations, the findings from the present study suggest that targeting ENT2 could potentially be a therapeutic strategy for the enhancement of CRC treatment efficacy.
Acknowledgements
Not applicable.
Funding Statement
Funding: The present study was supported by the Ministry of Higher Education Government of Malaysia under a Long-Term Research Grant (grant no. LRGS/2014/UKM-UiTM/K/03) and the Fundamental Research Grant Scheme (no. FRGS/1/2019/SKK08/UITM/02/11). It was funded by the National Cancer Council [Majlis Kanser Nasional; no. 100-IRMI/16/6/2 (011/2019)].
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
SMN drafted the manuscript. SMN and NAAH conducted the experiments, and analyzed and interpreted the data. SAR, MM and AAR designed the study, discussed experimental approaches and outcomes, and critically revised the manuscript. MM and AAR obtained funding. AAR, SAR and MM confirm the authenticity of all the raw data. All authors read and approved the final version of the manuscript.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Wong MC, Huang J, Lok V, Wang J, Fung F, Ding H, Zheng ZJ. Differences in incidence and mortality trends of colorectal cancer worldwide based on sex, age, and anatomic location. Clin Gastroenterol Hepatol. 2021;19:955–966.e61. doi: 10.1016/j.cgh.2020.02.026. [DOI] [PubMed] [Google Scholar]
- 2.Xie YH, Chen YX, Fang JY. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct Target Ther. 2020;5:1–30. doi: 10.1038/s41392-020-0116-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Peravali R, Hall N. Colorectal cancer: Features and investigation. Medicine. 2015;43:299–302. [Google Scholar]
- 4.Simon K. Colorectal cancer development and advances in screening. Clin Interv Aging. 2016;11:967–976. doi: 10.2147/CIA.S109285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Martinez-Outschoorn UE, Peiris-Pagés M, Pestell RG, Sotgia F, Lisanti MP. Cancer metabolism: A therapeutic perspective. Nat Rev Clin Oncol. 2017;14:11–31. doi: 10.1038/nrclinonc.2017.1. [DOI] [PubMed] [Google Scholar]
- 6.Hashim NAA, Ab-Rahim S, Ngah WZW, Nathan S, Ab Mutalib NS, Sagap I, Jamal ARA, Mazlan M. Global metabolomics profiling of colorectal cancer in Malaysian patients. Bioimpacts. 2021;11:33–43. doi: 10.34172/bi.2021.05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Pedley AM, Benkovic SJ. A new view into the regulation of purine metabolism: The purinosome. Trends Biochem Sci. 2017;42:141–154. doi: 10.1016/j.tibs.2016.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Di Virgilio F, Adinolfi E. Extracellular purines, purinergic receptors and tumor growth. Oncogene. 2017;36:293–303. doi: 10.1038/onc.2016.206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bester AC, Roniger M, Oren YS, Im MM, Sarni D, Chaoat M, Bensimon A, Zamir G, Shewach DS, Kerem B. Nucleotide deficiency promotes genomic instability in early stages of cancer development. Cell. 2011;145:435–446. doi: 10.1016/j.cell.2011.03.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yamaoka T, Kondo M, Honda S, Iwahana H, Moritani M, Ii S, Yoshimoto K, Itakura M. Amidophosphoribosyltransferase limits the rate of cell growth-linked de novo purine biosynthesis in the presence of constant capacity of salvage purine biosynthesis. J Biol Chem. 1997;272:17719–17725. doi: 10.1074/jbc.272.28.17719. [DOI] [PubMed] [Google Scholar]
- 11.Yin J, Ren W, Huang X, Deng J, Li T, Yin Y. Potential mechanisms connecting purine metabolism and cancer therapy. Front Immunol. 2018;9(1697) doi: 10.3389/fimmu.2018.01697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rosenthal EL, Chung TK, Parker WB, Allan PW, Clemons LL, Lowman D, Hong J, Hunt FR, Richman J, Conry RM, et al. Phase I dose-escalating trial of Escherichia coli purine nucleoside phosphorylase and fludarabine gene therapy for advanced solid tumors. Ann Oncol. 2015;26:1481–1487. doi: 10.1093/annonc/mdv196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.dos Santos-Rodrigues A, Grañé-Boladeras N, Bicket A, Coe IR. Nucleoside transporters in the purinome. Neurochem Int. 2014;73:229–237. doi: 10.1016/j.neuint.2014.03.014. [DOI] [PubMed] [Google Scholar]
- 14.Cass CE, Young JD, Baldwin SA, Cabrita MA, Graham KA, Griffiths M, Jennings LL, Mackey JR, Ng AM, Ritzel MW, et al. Nucleoside transporters of mammalian cells. Pharm Biotechnol. 2002;12:313–352. doi: 10.1007/0-306-46812-3_12. [DOI] [PubMed] [Google Scholar]
- 15.Altaweraqi RA, Yao SY, Smith KM, Cass CE, Young JD. HPLC reveals novel features of nucleoside and nucleobase homeostasis, nucleoside metabolism and nucleoside transport. Biochim Biophys Acta Biomembr. 2020;1862(183247) doi: 10.1016/j.bbamem.2020.183247. [DOI] [PubMed] [Google Scholar]
- 16.Naes SM, Ab-Rahim S, Mazlan M, Rahman AA. Equilibrative nucleoside transporter 2: Properties and physiological roles. Biomed Res Int. 2020;3(5197626) doi: 10.1155/2020/5197626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Young JD, Yao SY, Baldwin JM, Cass CE, Baldwin SA. The human concentrative and equilibrative nucleoside transporter families, SLC28 and SLC29. Mol Aspects Med. 2013;34:529–547. doi: 10.1016/j.mam.2012.05.007. [DOI] [PubMed] [Google Scholar]
- 18.Tang PC, Yang C, Li RWS, Lee SMY, Hoi MPM, Chan SW, Kwan YW, Tse CM, Leung GPH. Inhibition of human equilibrative nucleoside transporters by 4-((4-(2-fluorophenyl) piperazin-1-yl) methyl)-6-imino-N-(naphthalen-2-yl)-1,3,5-triazin-2-amine. Eur J Pharmacol. 2016;791:544–551. doi: 10.1016/j.ejphar.2016.07.002. [DOI] [PubMed] [Google Scholar]
- 19.Phua LC, Mal M, Koh PK, Cheah PY, Chan ECY, Ho HK. Investigating the role of nucleoside transporters in the resistance of colorectal cancer to 5-fluorouracil therapy. Cancer Chemother Pharmacol. 2013;71:817–823. doi: 10.1007/s00280-012-2054-0. [DOI] [PubMed] [Google Scholar]
- 20.Al-Tawfiq JA, Al-Homoud AH, Memish ZA. Remdesivir as a possible therapeutic option for the COVID-19. Travel Med Infect Dis. 2020;34(101615) doi: 10.1016/j.tmaid.2020.101615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Scopes RK. Enzyme activity and assays. e LS. 2001. [Google Scholar]
- 22.Gatzidou E, Mantzourani M, Giaginis C, Giagini A, Patsouris E, Kouraklis G, Theocharis S. Augmenter of liver regeneration gene expression in human colon cancer cell lines and clinical tissue samples. J BUON. 2015;20:84–91. [PubMed] [Google Scholar]
- 23.Yusof HM, Ab-Rahim S, Ngah WZW, Nathan S, Jamal ARA, Mazlan M. Metabolomic characterization of colorectal cancer cell lines highlighting stage-specific alterations during cancer progression. Bioimpacts. 2021;11:147–156. doi: 10.34172/bi.2021.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL, et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin Chem. 2009;55:611–622. doi: 10.1373/clinchem.2008.112797. [DOI] [PubMed] [Google Scholar]
- 25.Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative CT method. Nat Protoc. 2008;3:1101–1108. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- 26.Brown DG, Rao S, Weir TL, O'Malia J, Bazan M, Brown RJ, Ryan EP. Metabolomics and metabolic pathway networks from human colorectal cancers, adjacent mucosa, and stool. Cancer Metab. 2016;4(11) doi: 10.1186/s40170-016-0151-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Long Y, Sanchez-Espiridion B, Lin M, White L, Mishra L, Raju GS, Kopetz S, Eng C, Hildebrandt MAT, Chang DW, et al. Global and targeted serum metabolic profiling of colorectal cancer progression. Cancer. 2017;123:4066–4074. doi: 10.1002/cncr.30829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Luo X, Yu H, Song Y, Sun T. Integration of metabolomic and transcriptomic data reveals metabolic pathway alteration in breast cancer and impact of related signature on survival. J Cell Physiol. 2019;234:13021–13031. doi: 10.1002/jcp.27973. [DOI] [PubMed] [Google Scholar]
- 29.Sahu D, Lotan Y, Wittmann B, Neri B, Hansel DE. Metabolomics analysis reveals distinct profiles of nonmuscle-invasive and muscle-invasive bladder cancer. Cancer Med. 2017;6:2106–2120. doi: 10.1002/cam4.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zhu J, Djukovic D, Deng L, Gu H, Himmati F, Chiorean EG, Raftery D. Colorectal cancer detection using targeted serum metabolic profiling. J Proteome Res. 2014;13:4120–4130. doi: 10.1021/pr500494u. [DOI] [PubMed] [Google Scholar]
- 31.Garcia-Gil M, Camici M, Allegrini S, Pesi R, Petrotto E, Tozzi M. Emerging role of purine metabolizing enzymes in brain function and tumors. Int J Mol Sci. 2018;19(3598) doi: 10.3390/ijms19113598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Townsend MH, Felsted AM, Ence ZE, Piccolo SR, Robison RA, O'Neill K. Elevated expression of hypoxanthine guanine phosphoribosyltransferase within malignant tissue. Cancer Clin Oncol. 2017;6:19–34. [Google Scholar]
- 33.Sedano MJ, Ramos EI, Choudhari R, Harrison AL, Subramani R, Lakshmanaswamy R, Zilaie M, Gadad SS. Hypoxanthine phosphoribosyl transferase 1 is upregulated, predicts clinical outcome and controls gene expression in breast cancer. Cancers (Basel) 2020;12(1522) doi: 10.3390/cancers12061522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Camici M, Tozzi MG, Allegrini S, Del Corso A, Sanfilippo O, Daidone MG, De Marco C, Ipata PL. Purine salvage enzyme activities in normal and neoplastic human tissues. Cancer Biochem Biophys. 1990;11:201–209. [PubMed] [Google Scholar]
- 35.Sanfilippo O, Camici M, Tozzi MG, Turriani M, Faranda A, Ipata P, Silvestrini R. Relationship between the levels of purine salvage pathway enzymes and clinical/biological aggressiveness of human colon carcinoma. Cancer Biochem Biophys. 1994;14:57–66. [PubMed] [Google Scholar]
- 36.Kami K, Fujimori T, Sato H, Sato M, Yamamoto H, Ohashi Y, Sugiyama N, Ishihama Y, Onozuka H, Ochiai A, et al. Metabolomic profiling of lung and prostate tumor tissues by capillary electrophoresis time-of-flight mass spectrometry. Metabolomics. 2013;9:444–453. doi: 10.1007/s11306-012-0452-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nemkov T, Sun K, Reisz JA, Song A, Yoshida T, Dunham A, Wither MJ, Francis RO, Roach RC, Dzieciatkowska M, et al. Hypoxia modulates the purine salvage pathway and decreases red blood cell and supernatant levels of hypoxanthine during refrigerated storage. Haematologica. 2018;103:361–372. doi: 10.3324/haematol.2017.178608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ong ES, Zou L, Li S, Cheah PY, Eu KW, Ong CN. doi: 10.1074/mcp.M900551-MCP200. Metabolic profiling in colorectal cancer reveals signature metabolic shifts during tumorigenesis. Mol Cell Proteomics 10: doi: 10.1074, 2010. [DOI] [PubMed] [Google Scholar]
- 39.Durak İ, Cetin R, Devrim E, Ergüder İB. Effects of black grape extract on activities of DNA turn-over enzymes in cancerous and non cancerous human colon tissues. Life Sci. 2005;76:2995–3000. doi: 10.1016/j.lfs.2004.11.018. [DOI] [PubMed] [Google Scholar]
- 40.Battelli MG, Polito L, Bortolotti M, Bolognesi A. Xanthine oxidoreductase in cancer: More than a differentiation marker. Cancer Med. 2016;5:546–557. doi: 10.1002/cam4.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Linder N, Bützow R, Lassus H, Lundin M, Lundin J. Decreased xanthine oxidoreductase (XOR) is associated with a worse prognosis in patients with serous ovarian carcinoma. Gynecol Oncol. 2012;124:311–318. doi: 10.1016/j.ygyno.2011.10.026. [DOI] [PubMed] [Google Scholar]
- 42.Wang Y, Liu S, Tian S, Du R, Lin T, Xiao X, Wang R, Chen R, Geng H, Subramanian S, et al. C1QBP regulates apoptosis of renal cell carcinoma via modulating xanthine dehydrogenase (XDH) mediated ROS generation. Int J Med Sci. 2020;19:842–857. doi: 10.7150/ijms.71703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Linder N, Martelin E, Lundin M, Louhimo J, Nordling S, Haglund C, Lundin J. Xanthine oxidoreductase-Clinical significance in colorectal cancer and in vitro expression of the protein in human colon cancer cells. Eur J Cancer. 2009;45:648–655. doi: 10.1016/j.ejca.2008.10.036. [DOI] [PubMed] [Google Scholar]
- 44.Yuan C, Xu XH, Wang XL, Xu L, Chen Z, Li YQ. Relationship between serum uric acid and metastatic and nonmetastatic rectal cancer patients with undergoing no chemotherapy. Medicine (Baltimore) 2016;95(e5463) doi: 10.1097/MD.0000000000005463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Dhankhar R, Dahiya K, Sharma TK, Ghalaut VS, Atri R, Kaushal V. Diagnostic significance of adenosine deaminase, uric acid and C-reactive protein levels in patients of head and neck carcinoma. Clin Lab. 2011;57:795–798. [PubMed] [Google Scholar]
- 46.Fini MA, Elias A, Johnson RJ, Wright RM. Contribution of uric acid to cancer risk, recurrence, and mortality. Clin Transl Med. 2012;1:1–15. doi: 10.1186/2001-1326-1-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Shi L, Chen S, Yang L, Li Y. The role of PD-1 and PD-L1 in T-cell immune suppression in patients with hematological malignancies. J Hematol Oncol. 2013;6:1–6. doi: 10.1186/1756-8722-6-74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dziaman T, Banaszkiewicz Z, Roszkowski K, Gackowski D, Wisniewska E, Rozalski R, Foksinski M, Siomek A, Speina E, Winczura A, et al. 8-Oxo-7, 8-dihydroguanine and uric acid as efficient predictors of survival in colon cancer patients. Int J Cancer. 2014;134:376–383. doi: 10.1002/ijc.28374. [DOI] [PubMed] [Google Scholar]
- 49.Gamage CD, Park SY, Yang Y, Zhou R, Taş İ, Bae WK, Kim KK, Shim JH, Kim E, Yoon G, Kim H. Deoxypodophyllotoxin exerts anti-cancer effects on colorectal cancer cells through induction of apoptosis and suppression of tumorigenesis. Int J Mol Sci. 2019;20(2612) doi: 10.3390/ijms20112612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Trachootham D, Alexandre J, Huang P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat Rev Drug Dis. 2009;8:579–591. doi: 10.1038/nrd2803. [DOI] [PubMed] [Google Scholar]
- 51.Lu H, Li X, Lu Y, Qiu S, Fan Z. ASCT2 (SLC1A5) is an EGFR-associated protein that can be co-targeted by cetuximab to sensitize cancer cells to ROS-induced apoptosis. Cancer Lett. 2016;381:23–30. doi: 10.1016/j.canlet.2016.07.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ray PD, Huang BW, Tsuji Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012;24:981–990. doi: 10.1016/j.cellsig.2012.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Dueregger A, Guggenberger F, Barthelmes J, Stecher G, Schuh M, Intelmann D, Abel G, Haunschild J, Klocker H, Ramoner R, Sampson N. Attenuation of nucleoside and anti-cancer nucleoside analog drug uptake in prostate cancer cells by Cimicifuga racemosa extract BNO-1055. Phytomedicine. 2013;20:1306–1314. doi: 10.1016/j.phymed.2013.07.009. [DOI] [PubMed] [Google Scholar]
- 54.Al-Abdulla R, Perez-Silva L, Abete L, Romero MR, Briz O, Marin JJ. Unraveling ‘The Cancer Genome Atlas’ information on the role of SLC transporters in anticancer drug uptake. Expert Rev Clin Pharmacol. 2019;12:329–341. doi: 10.1080/17512433.2019.1581605. [DOI] [PubMed] [Google Scholar]
- 55.Mukhopadhya I, Murray GI, Berry S, Thomson J, Frank B, Gwozdz G, Ekeruche-Makinde J, Shattock R, Kelly C, Iannelli F, et al. Drug transporter gene expression in human colorectal tissue and cell lines: Modulation with antiretrovirals for microbicide optimization. J Antimicrob Chemother. 2015;71:372–386. doi: 10.1093/jac/dkv335. [DOI] [PubMed] [Google Scholar]
- 56.Liu Y, Zuo T, Zhu X, Ahuja N, Fu T. doi: 10.4238/gmr16019549. Differential expression of hENT1 and hENT2 in colon cancer cell lines. Genet Mol Res 16: doi: 10.4238, 2017. [DOI] [PubMed] [Google Scholar]
- 57.Pastor-Anglada M, Pérez-Torras S. Emerging roles of nucleoside transporters. Front Pharmacol. 2018;9(606) doi: 10.3389/fphar.2018.00606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Senyavina N, Tonevitskaya S. Effect of hypoxanthine on functional activity of nucleoside transporters ENT1 and ENT2 in caco-2 polar epithelial intestinal cells. Bull Exp Biol Med. 2015;160:160–164. doi: 10.1007/s10517-015-3118-z. [DOI] [PubMed] [Google Scholar]
- 59.Cho HD, Lee JH, Moon KD, Park KH, Lee MK, Seo KI. Auriculasin-induced ROS causes prostate cancer cell death via induction of apoptosis. Food Chem Toxicol. 2018;111:660–669. doi: 10.1016/j.fct.2017.12.007. [DOI] [PubMed] [Google Scholar]
- 60.Pan H, Wang BH, Lv W, Jiang Y, He L. Esculetin induces apoptosis in human gastric cancer cells through a cyclophilin D-mediated mitochondrial permeability transition pore associated with ROS. Chem Biol Interact. 2015;242:51–60. doi: 10.1016/j.cbi.2015.09.015. [DOI] [PubMed] [Google Scholar]
- 61.García V, Lara-Chica M, Cantarero I, Sterner O, Calzado MA, Muñoz E. Galiellalactone induces cell cycle arrest and apoptosis through the ATM/ATR pathway in prostate cancer cells. Oncotarget. 2016;7:4490–4506. doi: 10.18632/oncotarget.6606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Jung SN, Shin DS, Kim HN, Jeon YJ, Yun J, Lee YJ, Kang JS, Han DC, Kwon BM. Sugiol inhibits STAT3 activity via regulation of transketolase and ROS-mediated ERK activation in DU145 prostate carcinoma cells. Biochem Pharmacol. 2015;97:38–50. doi: 10.1016/j.bcp.2015.06.033. [DOI] [PubMed] [Google Scholar]
- 63.Fruehauf JP, Meyskens FL Jr. Reactive oxygen species: A breath of life or death? Clin Cancer Res. 2007;13:789–794. doi: 10.1158/1078-0432.CCR-06-2082. [DOI] [PubMed] [Google Scholar]
- 64.Pelin M, Fusco L, Martín C, Sosa S, Frontiñán-Rubio J, González-Domínguez JM, Vázquez E, Prato M, Tubaro A. Graphene and graphene oxide induce ROS production in human HaCaT skin keratinocytes: The role of xanthine oxidase and NADH dehydrogenase. Nanoscale. 2018;10:11820–11830. doi: 10.1039/c8nr02933d. [DOI] [PubMed] [Google Scholar]
- 65.Kim K, Yeo SG, Yoo BC. Identification of hypoxanthine and phosphoenolpyruvic Acid as serum markers of chemoradiotherapy response in locally advanced rectal cancer. Cancer Res Treat. 2015;47:78–89. doi: 10.4143/crt.2013.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Zhang A, Sun H, Xu H, Qiu S, Wang X. Cell metabolomics. OMICS. 2013;17:495–501. doi: 10.1089/omi.2012.0090. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.