

Abstract
Background Evidence indicates a strong link between Zika virus (ZikV) and neurological complications. Acute myelitis, optic neuritis, polyneuropathy, and encephalomyelitis that mimic inflammatory idiopathic demyelination disorders (IIDD) after ZikV infection have been reported in Brazil.
Objective The present study aims to investigate the possible occurrence of molecular mimicry between ZikV antigens and Multiple Sclerosis (MS) autoantigens, the most frequent IIDD of the central nervous system (CNS).
Methods A retrospective cohort study with 305 patients admitted due to suspected arbovirus infection in Rio de Janeiro was performed, all subjects were submitted to neurological examination, and a biological sample was collected for serologic and molecular diagnostic. Bioinformatics tools were used to analyze the peptides shared between ZikV antigens and MS autoantigens.
Results Of 305 patients, twenty-six were positive for ZikV and 4 presented IDD patterns found in MS cases. Sequence homology comparisons by bioinformatics approach between NS5 ZikV and PLP MS protein revealed a homology of 5/6 consecutive amino acids (CSSVPV/CSAVPV) with 83% identity, deducing a molecular mimicry. Analysis of the 3D structures revealed a similar conformation with alpha helix presentation.
Conclusions Molecular mimicry between NS5 Zika virus antigen and PLP MS autoantigens emerge as a possible mechanism for IDD spectrum in genetically susceptible individuals.
Keyword: Zika Virus, Demyelinating Diseases, Molecular Mimicry, Viral Nonstructural Proteins, Multiple Sclerosis
Resumo
Antecedentes Evidências indicam uma forte ligação entre o vírus Zika (ZikV) e complicações neurológicas. Mielite aguda, neurite óptica, polineuropatia e encefalomielite que mimetizam distúrbios inflamatórios de desmielinização idiopáticos (DDII) após infecção por ZikV têm sido relatadas no Brasil.
Obejtivo O presente estudo tem como objetivo investigar a possível ocorrência de mimetismo molecular entre antígenos do ZikV e autoantígenos da Esclerose Múltipla (EM), a DDII mais frequente do sistema nervoso central (SNC).
Métodos Foi realizado um estudo de coorte retrospectivo com 305 pacientes internados por suspeita de infecção por arbovírus no Rio de Janeiro, todos os indivíduos foram submetidos a exame neurológico e coleta de amostra biológica para diagnóstico sorológico e molecular. Ferramentas de bioinformática foram usadas para analisar os peptídeos compartilhados entre antígenos do ZikV e autoantígenos da EM.
Resultados Dos 305 pacientes, vinte e seis foram positivos para ZikV e 4 apresentaram padrão IDD encontrado em casos de EM. As comparações de homologia de sequência por abordagem de bioinformática entre a proteína NS5 ZikV e PLP EM revelaram uma homologia de 5/6 aminoácidos consecutivos (CSSVPV/CSAVPV) com 83% de identidade, deduzindo um mimetismo molecular. A análise das estruturas 3D revelou uma conformação semelhante com apresentação em alfa-hélice.
Conclusões O mimetismo molecular entre o antígeno NS5 do vírus Zika e o autoantígeno PLP da EM surge como um possível mecanismo para o espectro IDD em indivíduos geneticamente suscetíveis.
Palavras-chave: Zika Virus, Doenças Desmielinizantes, Mimetismo Molecular, Proteínas não Estruturais Virais, Esclerose Múltipla
INTRODUCTION
Over 80% of Zika Virus (ZikV) infections in humans are asymptomatic. Typical symptoms can include rash, fever, joint pain, and conjunctivitis for a period of 7 days. The outbreak of ZikV has increased the occurrence of long term neurological complications, such as Guillain-Barré syndrome, acute flaccid paralysis, and meningoencephalitis. 1 In addition, ZikV was detected by serology in cerebrospinal fluid (CSF), molecular and histopathological analysis of the brain, and amniotic fluid of microcephalic fetuses. 2 3 ZikV has also been associated with central nervous system (CNS) inflammatory demyelinating disorders (IDD) including optic neuritis, 4 neuromyelitis optica spectrum disorders (NMOSD), 5 transverse myelitis and acute disseminated encephalomyelitis (ADEM). 2 Our group has recently published a case in which the coexistence of the virus in the CNS of an MS patient led to the development of an ADEM-like episode. 6
Besides its direct neurotropic effect, 7 it is believed that ZikV may function as a trigger leading to the development of an immune-mediated injury against many parts of the CNS. 8 ZikV has already been related to the development of several autoimmune conditions. 9 In Guillain-Barre syndrome (GBS), for example, the molecular mimicry between glycolipids and surface molecules of the virus has explained the majority of cases. 10 Interestingly, ZikV is commonly associated with magnetic resonance imaging (MRI) lesions distributed in space and time, regarding heterogeneous gadolinium enhancement, as seen in the MS criteria. 11 Moreover, serum positivity for autoantibodies against myelin oligodendrocyte glycoprotein (MOG), a specific antibody against the myelin sheath was recently associated with ZikV. 12 As many radiological and clinical aspects of ZikV infection may mimic IIDD, patients can be misdiagnosed. MS is the most frequent IIDD of the CNS, 13 and several evidences have shown that molecular mimicry is a possible epigenetic mechanism in genetically susceptible individuals. 14
To investigate the mechanisms of ZikV induced neurological manifestations, it is essential to use various reproducible in vitro models and bioinformatics tools capable of recapitulating complex neurodevelopmental disorders, in an attempt to find specific targets. The molecular mechanisms underlying these conditions in adults are not clear. Focusing on the MS-like pattern, the present study investigated the possible occurrence of molecular mimicry between ZikV antigens and MS autoantigens. The underlying rationale is that shared peptides between pathogen and human host may lead to a break in immune tolerance through a cross-reactivity phenomenon. 15
METHODS
Study population and biological samples
A retrospective cohort study was performed with patients admitted in neurology service of three university hospitals and referred by Laboratório Central Noel Nutels (LACEN) in Rio de Janeiro. This work was approved by the National Council for Ethics in Research (CAAE 69411317.6.0000.5258). All subjects signed an informed consent agreeing to participate in this research. From 2016 to 2019, 305 patients with suspected arbovirus infection were evaluated by a multidisciplinary team. Complete physical and neurological examination was performed and, when necessary, MRI was requested. Biological sample (blood, urine, and CSF) was collected on admission and, according to clinical indication, tested by serology and/or ZikV molecular diagnostic.
Sequence analysis
Peptide sharing between ZikV antigens and MS autoantigens was analyzed as follows: A viral polyprotein library was constructed using the major viral antigens reported in the literature and protein sequences available in NCBI Protein Reference Sequences ( https://www.ncbi.nlm.nih.gov/protein ). An MS autoantigen library was constructed at random through UniProtKB Database ( www.uniprot.org/ ) using ‘Multiple Sclerosis’ as a keyword. The result was filtered and only the proteins confirmed as autoantigens were collected. ZikV polyproteins and MS autoantigens identified are outlined in Table 1 . The two libraries were analyzed for matches using BLASTP ( https://blast.ncbi.nlm.nih.gov/Blast.cgi ) and sequence alignment was done using EMBOSS ( https://www.ebi.ac.uk/Tools/psa/emboss_water/ ).
Table 1. ZikV polyproteins and human MS autoantigens-related proteins.
| ZikV polyproteins | Multiple sclerosis autoantigens |
|---|---|
| Chain B, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain PDB: 5GS6_B |
myelin oligodendrocyte glycoprotein (MOG) UniProtKB- Q16653 |
| Chain A, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain PDB: 5GS6_A |
myelin basic protein (MBP) UniProtKB- P02686 |
| Chain A, Zika Virus Non-structural Protein 1 (ns1) PDB: 5K6K_A |
Myelin associated glycoprotein (MAG) UniProtKB- P20916 |
| Chain B, Zika Virus Non-structural Protein 1 (ns1) PDB: 5K6K_B |
Myelin proteolipid protein (PLP) UniProtKB- P60201 |
| Chain A, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro PDB: 5 × 8Y_A |
|
| Chain B, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro PDB: 5 × 8Y_B |
|
| NS1 [Zika virus] NCBI Reference Sequence: YP_009430301.1 |
|
| NS2a NCBI Reference Sequence: YP_009430302.1 |
|
| NS2b NCBI Reference Sequence: YP_009430303.1 |
|
| NS3 NCBI Reference Sequence: YP_009430304.1 |
|
| NS4a NCBI Reference Sequence: YP_009430305.1 |
|
| NS4b Chain A, Structure of Zika Virus Ns5 NCBI Reference Sequence: YP_009430307.1 |
|
| Chain B, Structure of Zika Virus Ns5 PDB: 5TMH_A |
|
| NS5 protein, partial [Zika virus] GenBank: AJD79051.1, AMP44573.1, AMX81921.1, AMX81921.1 |
|
| envelope protein E, partial [Zika virus] GenBank: AOX24134.1 |
|
| capsid protein C [Zika virus] NCBI Reference Sequence: YP_009430296.1 |
Antigenic prediction
To confirm whether the NS5 ZikV sequence studied has antigenic properties, VaxiJen version 2.0 ( http://www.ddg-pharmfac.net/vaxijen/VaxiJen/VaxiJen.html ) was used. A threshold antigenic score of 0.5 was defined in order to filter probable non-antigenic sequences. Vaxijen server performs alignment-independent prediction, which is based on auto cross covariance (ACC) transformation of protein sequences into uniform vectors of principal amino acid properties.
3D comparative modelling
The 3D models were built using the Swiss-Model, an online modeling server ( https://swissmodel.expasy.org/ ). The template modeling scores (TM-scores) and root mean square deviations (RMSDs) of the NS5 ZikV and PLP MS three-dimensional overlap were calculated using TM-Align.
RESULTS
Inflammatory demyelinating disorder phenotypes in patients with ZikV infection
A total of 305 patients were evaluated. 26 were positive for ZikV and the remaining were diagnosed with either Dengue or Chikungunya. Out of the ZikV positive patients, 4 were classified as having IDD of the CNS requiring differential diagnosis with MS. Clinical examination, imaging, electrophysiologic, and laboratory findings of these patients are exposed in Table 2 and Figures 1 - 4 .
Table 2. Clinical and laboratory findings in patients with Zika virus–associated Multiple Sclerosis-like manifestations.
| Clinical presentation | Patient 1 | Patient 2 | Patient 3 | Patient 4 |
|---|---|---|---|---|
| Age | 30 | 51 | 48 | 57 |
| Sex | Male | Male | Male | Female |
| Medical history | EM | None | None | None |
| Viral prodrome | Fever and myalgia | Acute fever and rash | Acute fever and rash | Fever, intense myalgia, and skin rash |
| Neurologic symptoms | Acute encephalomyelitis with drowsiness, mental confusion, locomotor disorders and diplopia. ADEM |
Paraparesis, that evolved into tetraparesis. ADEM |
Agitation and disorientation preceded, acute encephalomyelitis | Visual loss and walking impairment |
| Time from viral prodrome to neurologic symptoms | 5 days | 11 days | 10 days | 11 days |
| Neurologic examination | Hypoesthesia right upper limb and papillitis, | Dysarthria, tetraparesis and drowsiness | Tetraparesis, disorientation and decreased consciousness level | Spastic symmetric crural paraplegia, papilemema, left visual loss |
| Diagnostic studies | ||||
| ZikV RT-PCR | Negative | Negative | Negative | Positive |
| Igm ZikV | Positive in the serum and urine | Positive in the serum | Positive in the serum | Positive in the serum |
| Igg ZikV | Positive in the serum, urine and LCR | Positive in the serum | Positive in the serum | Negative in the serum |
| CSF | 8 leukocyte/mm 3 , 27 mg/dl protein, OCB positive, IgG index 0,88 | 0 leukocytes/mm 3 , 27 mg/dl protein. | 15 leukocytes/mm 3 , 71 mg/dL protein, and 55 mg/dL glucose | 9 leukocyte/mm 3 , 64 mg/dl protein, OCB negative, AQP4 Ab Negative |
Figure 1.
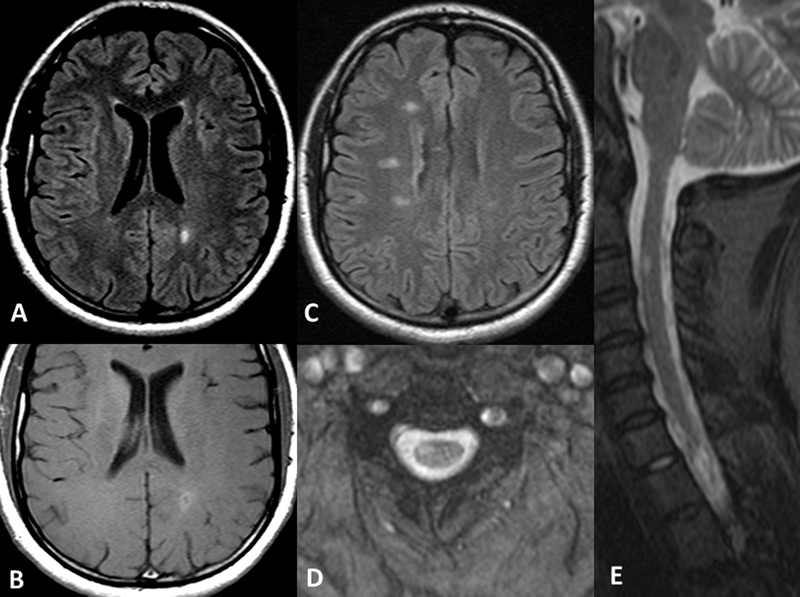
A focal subcortical hyperintense FLAIR lesion ( A ) with contrast enhancement ( B ) is observed in conjunction with other periventricular and pericalosal bright lesions ( C ), similar to Dawson's fingers described for MS disease. Cervical lesions follow the same pattern, eccentrically located in the T2* axial plane ( D ) and extending for one vertebral body dimension on the sagittal STIR cervical image ( E ).
Figure 4.
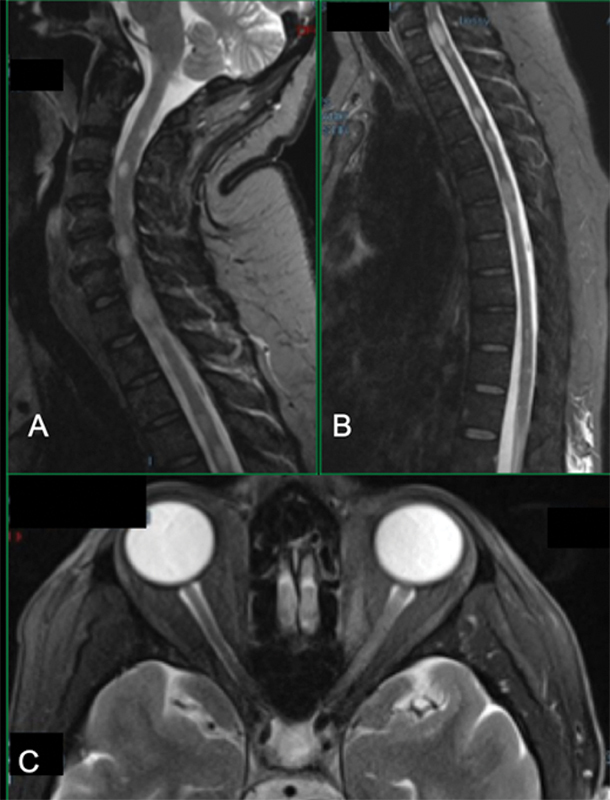
Sagittal STIR cervical ( A ) and dorsal ( B ) spinal cords have multiple small tumefactive bright lesions, randomly affecting all main cords tracts, diffusively distributed. Axial T2-fat suppressed at the orbital area shows bright thickening of the intra-orbital extent of the left optic nerve, reflecting extensive optic neuritis.
Patient 1 presented with headache, optical neuritis, and cervical myelitis associated with a cervical lesion ( Figure 1 E ) and asymptomatic multifocal brain lesions on MRI, one of which had gadolinium enhancement. This distribution of brain lesions, paired with positive oligoclonal bands (OCB) found on CSF analysis, resembles the pattern usually found in MS ( Figure 1 A-D and Table 1 ).
Patient 2 had a diagnosis of acute flaccid paraplegia 11 days after a viral prodrome, and 3 months later developed tetraparesis associated with longitudinal extensive transverse myelitis ( Figure 2C and 2D ), centrally located ( Figure 2 C3, D3 ), with focal tapering of the cervical/dorsal transition on sagittal STIR ( Figure 2 C1 ), resembling the extension and sequelae areas usually seen in NMOSD. Furthermore, the lesion had anterior horn involvement ( Figure 2 D4 ).
Figure 2.
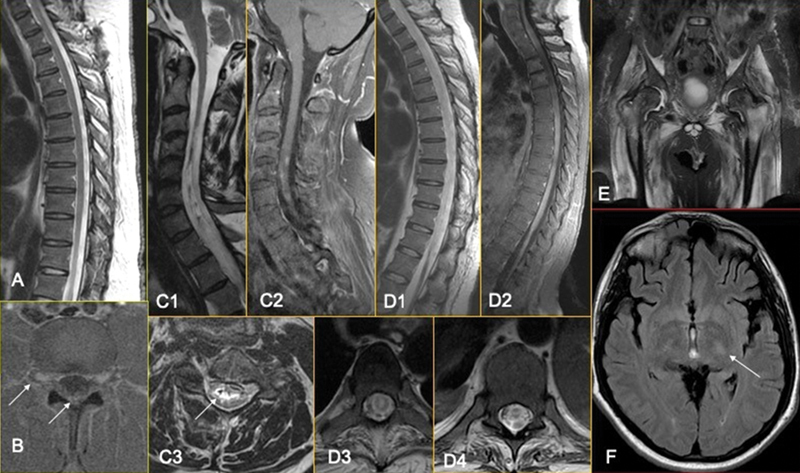
Initial dorsal MRI ( A ) was normal, and a significant contrast enhancement was observed in the axial T1 fat-sat image of the lumbar spine, involving the dorsal ganglia and the lumbo-sacral plexus inside the spine canal ( B ). After 3 months of evolution, the patient developed longitudinal extensive transverse myelitis ( C, D ), already with focal tapering of the cervical/dorsal transition on sagittal STIR (C1), remembering a sequel area. The lesion was centrally located (C3, D3), with anterior horn involvement (D4) and signals of previous bleeding inside the central canal (C3). A patch and irregular contrast enhancement were noticed along the sequel area (C2) and along the entire dorsal spinal cord (D2). Consequent muscle denervation was observed in the coronal STIR of the pelvic girdle muscles ( E ) and ascendant cortical-spinal tract degeneration consequent to the spinal cord damage on FLAIR axial images ( F ).
Patient 3 presented with tetra paresis and ataxia associated with brain lesions mainly affecting the brainstem on axial T2 images, including the posterior aspect of the mesencephalon ( Figure 3E ), pons ( Figure 3F ) and the medial cerebellar peduncle ( Figure 3G ).
Figure 3.
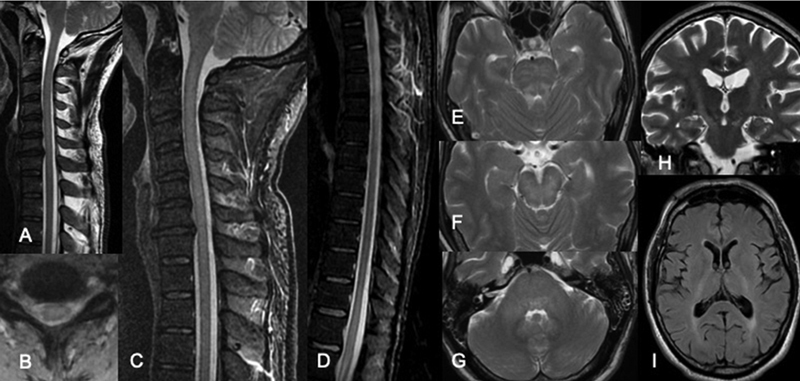
Cervical spinal cord sagittal T2 ( A ) shows extensive continuous high signal intensity lesion affecting the hole diameter of the spinal cord on axial T2* images ( B ), best identified on sagittal STIR ( C ). The extent of more than 3 vertebral bodies was confirmed, as well as the involvement of the medullary cone on sagittal STIR ( D ). Brain lesions were mainly detected affecting the brain stem on axial T2 images, including the posterior aspect of the mesencephalon ( E ), pons ( F ), and the medial cerebellar peduncle ( G ). Two years follow up brain images show hypersignal intensity on coronal T2 ( H ) and axial FLAIR ( I ) images located in the cortical-spinal tract, mostly associated with retrograde degeneration within the spinal cord lesions.
Patient 4 presented with optic neuritis and multifocal myelitis with cervical and dorsal lesions, as usually found in a first manifestation of MS ( Figures 4A and 4B ).
Sequence sharing between ZikV polyproteins and MS autoantigens
The bioinformatics approach identified an 83% identity between the NS5 antigen of ZikV and PLP MS autoantigen, deducing the molecular mimicry among them. Although statistically non-significant, it was also possible to observe a 67% identity between NS3 antigen of ZikV and MOG MS autoantigen. The identity results between all sequences are depicted in Table 3 . In addition, sequence analysis of NS5 using VaxiJen version 2.0 resulted in a score of 0.5091, confirming the antigenicity of the sequence studied.
Table 3. Results of alignment between ZikV polyproteins and MS autoantigens.
| Autoantigens | Polyproteins | Identity |
|---|---|---|
| MOG | NS1 | 50% |
| MOG | Chain B, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain | 43% |
| MOG | Chain A, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain | 43% |
| MOG | Chain A, Zika Virus Non-structural Protein 1 (ns1) | 43% |
| MOG | Chain B, Zika Virus Non-structural Protein 1 (ns1) | 43% |
| MOG | Chain A, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro | 36% |
| MOG | Chain B, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro | 36% |
| MOG | NS2a | 0% |
| MOG | NS2b | 0% |
| MOG | NS3 | 67% |
| MOG | NS4a | 50% |
| MOG | NS4b | 29% |
| MOG | Chain A, Structure of Zika Virus Ns5 | 50% |
| MOG | Chain B, Structure of Zika Virus Ns5 | 50% |
| MOG | NS5 protein, partial [Zika virus] | 50% |
| MOG | envelope protein E, partial | 50% |
| MOG | capsid protein C | 35% |
| PBM | Chain B, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain | 0% |
| PBM | Chain A, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain | 0% |
| PBM | Chain A, Zika Virus Non-structural Protein 1 (ns1) | 0% |
| PBM | Chain B, Zika Virus Non-structural Protein 1 (ns1) | 0% |
| PBM | Chain A, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro | 0% |
| PBM | Chain B, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro | 0% |
| PBM | NS1 | 0% |
| PBM | NS2a | 50% |
| PBM | NS2b | 21% |
| PBM | NS3 | 33% |
| PBM | NS4a | 33% |
| PBM | NS4b | 40% |
| PBM | Chain A, Structure of Zika Virus Ns5 | 32% |
| PBM | Chain B, Structure of Zika Virus Ns5 | 32% |
| PBM | NS5 protein, partial | 30% |
| PBM | envelope protein E, partia | 40% |
| PBM | capsid protein C | 0% |
| MAG | Chain B, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain | 0% |
| MAG | Chain A, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain | 0% |
| MAG | Chain A, Zika Virus Non-structural Protein 1 (ns1) | 0% |
| MAG | Chain B, Zika Virus Non-structural Protein 1 (ns1) | 0% |
| MAG | Chain A, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro | 0% |
| MAG | Chain B, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro | 0% |
| MAG | NS1 | 0% |
| MAG | NS2a | 50% |
| MAG | NS2b | 21% |
| MAG | NS3 | 33% |
| MAG | NS4a | 33% |
| MAG | NS4b | 40% |
| MAG | Chain A, Structure of Zika Virus Ns5 | 32% |
| MAG | Chain B, Structure of Zika Virus Ns5 | 32% |
| MAG | NS5 protein, partial | 30% |
| MAG | envelope protein E, partial | 40% |
| MAG | capsid protein C | 0% |
| PLP | Chain B, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain | 56% |
| PLP | Chain A, Full-length Ns1 Structure of Zika Virus From 2015 Brazil Strain | 56% |
| PLP | Chain A, Zika Virus Non-structural Protein 1 (ns1) | 56% |
| PLP | Chain B, Zika Virus Non-structural Protein 1 (ns1) | 56% |
| PLP | Chain A, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro | 0% |
| PLP | Chain B, A Mutation Identified in Neonatal Microcephaly Destabilizes Zika Virus Ns1 Assembly In Vitro | 0% |
| PLP | NS1 | 56% |
| PLP | NS2a | 24% |
| PLP | NS2b | 38% |
| PLP | NS3 | 46% |
| PLP | NS4a | 0% |
| PLP | NS4b | 50% |
| PLP | Chain A, Structure of Zika Virus Ns5 | 30% |
| PLP | Chain B, Structure of Zika Virus Ns5 | 30% |
| PLP | NS5 protein, partial | 83% |
| PLP | envelope protein E, partial | 42% |
| PLP | capsid protein C | 20% |
Structural conformation between NS5 ZikV and PLP MS
In order to predict the 3D structures conformation of the two proteins, TM-Align was used to align them. As Blast P showed us a high identity between a particular region of PLP and NS5, a structural conformation was performed only with that region where the corresponding high identity was obtained PLP 131-198 and NS5 281-325 ( Figure 5B ). The CSAVPV sequence which is 83% identity by BlastP, obtained a TM-score of 0.47071 and RMDS of 2.39 and is in the alpha helix structure of both proteins.
Figure 5.

Results of alignment between NS5 antigen of ZikV and PLP MS autoantigens. Alignment of amino acid using EMBOSS needle. Note the motif highlighted has 83% identity (A). Structural alignment between PLP 131-198 and NS5 281-325 proteins. In red the PLP 131-198 is represented and in blue the NS5 281-325 (B).
DISCUSSION
Several studies have shown that IDD in CNS can be triggered by viral infection or immunizations. After a variable period of incubation, myelin destruction undergoes courses of remission and exacerbation. MS is a most common disease that compromises CNS myelin sheath. 16
Viral infection can trigger autoimmune diseases through different mechanisms: molecular mimicry, epitope spreading, bystander activation, superantigen production, and inadequate activation of an immune response. 17 Molecular mimicry can be defined as similar structures shared by a host epitope and microorganism or environmental proteins. 17 Using bioinformatics tools, common sequences and structural homology between Chikungunya virus (ChikV) E1 glycoprotein and human HLA-B27 molecule were identified. In addition, the peptides derived from ChikV glycoprotein E1 induced significant inflammation in C57BL/6J mice. 18 Based on proteomic studies and sequence analysis, some evidence has also shown that Dengue Hemorrhagic Fever may be caused by molecular mimicry between different coagulation molecules with prM, E, and NS1 viral proteins. 19 Furthermore, it is already widely proposed that cross-recognition of common viral peptides with myelin antigens induces a molecular mimicry involved in MS development, especially in genetically susceptible individuals. 14
Zika Virus has 7 non structural proteins (NS1, NS2a, NS2b, NS3, NS4a, NS4b, and NS5). Each of them has a paper on the antagonism to the innate immunity. However, the NS5 protein stands out here, as it is the most conserved protein in the flavivirus proteome and it can modulate the host immune response during ZikV infection. 20 NS5 is an antagonist of the interferon response in the host human system and promotes the degradation of STAT2 in ZikV. This same mechanism is well reported in DenV. The importance of NS5 in host immune response modulation and viral replication makes it an attractive target for developing broadly acting antiviral inhibitors. 21
Zika fever is a self-limited disease, still, less than 5% of symptomatic patients may develop neurological manifestations. 22 23
Although patient 1 developed neurological manifestations five days after the first symptoms of viral infection, it was only possible to make the ZikV diagnosis after sixty days, thus explaining the IgM negativity in serum.
A recent case report identified concurrent GBS and ADEM in a 24-year-old woman who developed acute ZikV infection. The authors postulate this case was para-infectious, induced by neurotropism and activation of an immune response against ZikV. 24 This same mechanism is probably involved in the development of this NMOSD phenotype in our patient 2.
Patient 3 could be classified as having a clinical isolated syndrome (CIS) with a high risk of conversion to MS due to the distribution and number of T2 white matter lesions. Although the optical neuritis pattern resembles the one of NMOSD, the spinal cord lesions are MS-like.
Lucchese et al . 2016, observed that ZikV antigens are commonly involved in microcephaly and GBS. 129 immunopositive epitopes are reported as having peptide overlap with human proteins that may relate to demyelination and axonal neuropathies. This indicates that cross-reactivity with human proteins might contribute to the mechanisms linking ZikV infection to GBS. 10 The IDD phenotype attributed to ZikV infection seems to mimic MS manifestations. Molecular mimicry is assessed in this study by investigating homologous regions between ZikV antigens and human MS autoantigens using bioinformatics tools. Sequence homology comparisons between NS5 ZikV and PLP MS protein revealed a homology of 5/6 consecutive amino acids CSSVPV/CSAVPV ( Figure 5A ). A study that performed antigenic B-cell epitopes prediction found an antigenic peptide from position 528 to 539 (NAI CSSVPV DWV) of ZikV NS5, which had the maximum residual score of 1.203 and might present a preliminary set of peptides for future vaccine development against ZikV. 25 Calculating the TM-score of NS5 ZikV and PLP MS 3D structures demonstrated that both proteins are in almost the same fold, both are in alpha helix and they have topological similarity ( Figure 5B ). 26
Interestingly, ZikV African (MR766) lineage strain, revealed exactly the same Human PLP sequence (CSAVPV), and recombinant NS5 proteins from Africa and from Brazil revealed similar levels of RNA synthesis. 27 It is already known that the MR766 strain is more virulent and causes more severe brain damage than the current Asian lineage and dengue virus. 28 When inoculated subcutaneously in adult transgenic mice (knockout) C57BL/6 Stat2 -/- MR766 strain induces short episodes of severe neurological symptoms, followed by lethality. Furthermore, this strain was able to induce higher levels of inflammatory cytokines and markers associated with cellular infiltration into the brain of infected mice. 29 Li et al. 2019, observed that MR766 strain and epidemic Brazilian (BR15 and ICD) ZikV strains are different in viral attachment to host neuronal cells, viral permissiveness, and replication, as well as in the induction of cytopathic effects. 30
Autoreactivity to PLP in patients with MS has been investigated in human and animal model by various groups worldwide. 31 A recent study involving PLP's Epitopes involved in MS, found CSAVPV (in PLP 161-177 residues) among the most immunogenic regions of PLP. 32 In addition, the crystal structure of the NS5 ZikV protein reveals a conserved domain conformation of Flaviviruses, a genus that includes a variety of human pathogens such as dengue virus, yellow fever virus, WNV, Spondweni virus, and the Japanese encephalitis virus. 33 So, the presence of high identity between NS5 ZikV and PLP, an autoantigen widely implicated in the pathogenesis of MS, 34 leads us to postulate that molecular mimicry may have a role in the development of inflammatory demyelinating damage, a hallmark of the IDD produced by this genus of virus.
Both genetic and environmental factors have been shown to contribute to the pathogenesis of autoimmune diseases. It is well-established that the HLA-DR15 haplotype bears the strongest association with MS. 35 In a Brazilian study, it was observed that the presence of HLA-DRB1*1501 allele confers an ethnicity-dependent MS susceptibility in Caucasian patients and that the HLA-DQB1*0602 allele confers an ethnicity independent susceptibility. 36 Using HLA class II transgenic (Tg) mice, several studies have demonstrated HLA-DR-dependent disease following immunization by MBP, PLP, or MOG. 37 38 However, it was observed that HLA-DRB1*1501 Tg mice were refractory to disease induction by overlapping PLP peptides, while HLA-DQB1*0602 Tg mice were susceptible to disease induction by PLP 139-151 and PLP 175-194 peptides. 39 It has been seen that Both PLP 139-151 and PLP 178-191 epitopes are key targets of T-cells, and are increased in MS patients versus healthy controls. 40 However, this does not mean that PLP 161-177 residues are not encephalitogenic-related, but that they need further animal and human model studies. Therefore, PLP autoimmunity and HLA haplotype have been strongly associated with lesion localization, as well as remission and relapse rates in MS. 41
In conclusion, the concept of molecular mimicry remains a viable hypothesis for understanding the genetics, epigenetics, and environmental involvement in the pathogenic mechanisms of IDD. Studies using bioinformatics tools further encourage the identification of molecules that could be used in the development of either diagnostic or prognostic biomarkers. We found that NS5 ZikV presented a high identity with PLP MS autoantigen, and both are structurally similar to alpha helix chains. These findings may justify IDD CNS manifestations following ZikV infection, as in the 4 cases here reported. Further investigation is required to understand whether PLP 161-177 residues are encephalitogenic and how the recognition of NS5 epitopes by HLA molecules drives the pathogenic T-cell autoimmune response in vivo.
Acknowledgements
We appreciate the support of Secretaria de Saúde do Estado do Rio de Janeiro and Laboratório Central Noel Nutels (LACEN) and the Laboratorio de Virologia Molecular of Professor Amilcar Tanuri.
Conflict of Interest There is no conflict of interest to declare.
Authors' Contributions
SVAL, FLFD: conceived and designed the experiments; SVAL, FLFD, LCF, DGG, ADA, JPCG, CCSR, EVS, OJMN, FCRL, FFAF, JPBMS: subject recruitment and collection of the samples; ALH, RSA, OCFJ: serology and molecular diagnostic; FLFD, LCF, JFM: performing bioinformatics analyzes; LCF, FLFD, SVAL: analyzed the data and drafted the manuscript; LCF, FLFD: contributed equally to this work. All authors read and approved the final version.
Support
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Brazilian National Council for Scientific and Technological Development (CNPq Number 440779/2016-2), Coordination for the Improvement of Higher Education Personnel (CAPES Number 88887.130752/2016-00), Department of Science and Technology (DECIT No. 14/2016), Rede Nacional de Especialistas em ZIKA e Doenças Correlatas (RENEZIKA) and Foundation for Rio de Janeiro State Research (FAPERJ).
References
- 1.Baud D, Gubler D J, Schaub B, Lanteri M C, Musso D.An update on Zika virus infectionLancet 2017;390(10107):2099–2109. Doi: 10.1016/S0140-6736(17)31450-2 [Internet] [DOI] [PubMed]
- 2.Galliez R M, Spitz M, Rafful P P. Zika virus causing encephalomyelitis associated with immunoactivation. Open Forum Infect Dis. 2016;3(04):ofw203. doi: 10.1093/ofid/ofw203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mlakar J, Korva M, Tul N.Zika Virus Associated with Microcephaly N Engl J Med 201637410951–958. 10.1056/NEJMoa1600651[Internet] [DOI] [PubMed] [Google Scholar]
- 4.Singh S, Kumar A. Ocular Manifestations of Emerging Flaviviruses and the Blood-Retinal Barrier. Viruses. 2018;10(10):1–20. doi: 10.3390/v10100530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Aspahan M C, Leonhard S E, Gomez R S, Rocha E da S, Vilela M da S, Alvarenga P PM https://doi.org/10.1212/CPJ.0000000000000546. https://doi.org/10.1212/CPJ.0000000000000546 [DOI] [PMC free article] [PubMed]
- 6.Alves-Leon S V, Lima M DR, Nunes P CG. Zika virus found in brain tissue of a multiple sclerosis patient undergoing an acute disseminated encephalomyelitis-like episode. Mult Scler. 2019;25(03):427–430. doi: 10.1177/1352458518781992. [DOI] [PubMed] [Google Scholar]
- 7.Platt D J, Smith A M, Arora N, Diamond M S, Coyne C B, Miner J J.Zika virus-related neurotropic flaviviruses infect human placental explants and cause fetal demise in mice Sci Transl Med 201810426eaao7090. 10.1126/scitranslmed.aao7090[Internet] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wen J, Shresta S. T Cell Immunity to Zika and Dengue Viral Infections. J Interferon Cytokine Res. 2017;37(11):475–479. doi: 10.1089/jir.2017.0106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Monsalve D M, Pacheco Y, Acosta-ampudia Y, Rodríguez Y, Ramírez-santana C.Zika virus and autoimmunity. One-step forward. Autoimmun Rev [Internet]. 2017;17(S1568-9972):30258–6 Available from:https://doi.org/10.1016/j.autrev.2017.10.008 [DOI] [PubMed]
- 10.Lucchese G, Kanduc D.Zika virus and autoimmunity: From microcephaly to Guillain-Barré syndrome, and beyond Autoimmun Rev 20161508801–808. 10.1016/j.autrev.2016.03.020[Internet] [DOI] [PubMed] [Google Scholar]
- 11.Zare Mehrjardi M, Keshavarz E, Poretti A, Hazin A N. Neuroimaging findings of Zika virus infection: a review article. Jpn J Radiol. 2016;34(12):765–770. doi: 10.1007/s11604-016-0588-5. [DOI] [PubMed] [Google Scholar]
- 12.Neri V C, Xavier M F, Barros P O, Melo Bento C, Marignier R, Papais Alvarenga R. Case Report: Acute Transverse Myelitis after Zika Virus Infection. Am J Trop Med Hyg. 2018;99(06):1419–1421. doi: 10.4269/ajtmh.17-0938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Thompson A J, Baranzini S E, Geurts J, Hemmer B, Ciccarelli O.Multiple sclerosisLancet 2018;391(10130):1622–1636. Doi: 10.1016/S0140-6736(18)30481-1 [Internet] [DOI] [PubMed]
- 14.Huynh J L, Casaccia P. Epigenetic mechanisms in multiple sclerosis: implications for pathogenesis and treatment. Lancet Neurol. 2013;12(02):195–206. doi: 10.1016/S1474-4422(12)70309-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Robinson A P, Harp C T, Noronha A, Miller S D. The experimental autoimmune encephalomyelitis (EAE) model of MS: utility for understanding disease pathophysiology and treatment. Handb Clin Neurol. 2014;122:173–189. doi: 10.1016/B978-0-444-52001-2.00008-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Waubant E, Ponsonby A L, Pugliatti M, Hanwell H, Mowry E M, Hintzen R Q.Environmental and genetic factors in pediatric inflammatory demyelinating diseases Neurology 201687(9, Suppl 2)S20–S27. 10.1212/WNL.0000000000003029 [DOI] [PubMed] [Google Scholar]
- 17.Oldstone M B. Molecular mimicry and immune-mediated diseases. FASEB J. 1998;12(13):1255–1265. doi: 10.1096/fasebj.12.13.1255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Reddy V, Desai A, Krishna S S, Vasanthapuram R. Molecular Mimicry between Chikungunya Virus and Host Components: A Possible Mechanism for the Arthritic Manifestations. PLoS Negl Trop Dis. 2017;11(01):e0005238. doi: 10.1371/journal.pntd.0005238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lin Y-S, Yeh T-M, Lin C-F. Molecular mimicry between virus and host and its implications for dengue disease pathogenesis. Exp Biol Med (Maywood) 2011;236(05):515–523. doi: 10.1258/ebm.2011.010339. [DOI] [PubMed] [Google Scholar]
- 20.NG https://doi.org/10.1021/acsinfecdis.8b00373 https://doi.org/10.1021/acsinfecdis.8b00373
- 21.Shi Y, Gao G F. Structural Biology of the Zika Virus. Trends Biochem Sci. 2017;42(06):443–456. doi: 10.1016/j.tibs.2017.02.009. [DOI] [PubMed] [Google Scholar]
- 22.Meltzer E, Leshem E, Lustig Y, Gottesman G, Schwartz E.The Clinical Spectrum of Zika Virus in Returning Travelers Am J Med 2016129101126–1130. 10.1016/j.amjmed.2016.04.034[Internet] [DOI] [PubMed] [Google Scholar]
- 23.Beckham J D, Pastula D M, Massey A, Tyler K L. Zika virus as an emerging global pathogen: Neurological complications of zika virus. JAMA Neurol. 2016;73(07):875–879. doi: 10.1001/jamaneurol.2016.0800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Román G C, Anaya J-M, Mancera-Páez Ó, Pardo-Turriago R, Rodríguez Y.Concurrent Guillain-Barré syndrome, transverse myelitis and encephalitis post-Zika: A case report and review of the pathogenic role of multiple arboviral immunity J Neurol Sci 201939684–85. 10.1016/j.jns.2018.10.032[Internet] [DOI] [PubMed] [Google Scholar]
- 25.Mirza M U, Rafique S, Ali A.Towards peptide vaccines against Zika virus: Immunoinformatics combined with molecular dynamics simulations to predict antigenic epitopes of Zika viral proteins Sci Rep 20166(July):37313. 10.1038/srep37313 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhang Y, Skolnick J. TM-align: a protein structure alignment algorithm based on the TM-score. Nucleic Acids Res. 2005;33(07):2302–2309. doi: 10.1093/nar/gki524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhao B, Yi G, Du F.Structure and function of the Zika virus full-length NS5 protein Nat Commun 2017814762. 10.1038/ncomms14762[Internet] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Shao Q, Herrlinger S, Zhu Y-N, Yang M, Goodfellow F, Stice S L https://doi.org/10.1242/dev.156752. https://doi.org/10.1242/dev.156752 [DOI] [PMC free article] [PubMed]
- 29.Tripathi S, Balasubramaniam V RMT, Brown J A, Mena I, Grant A https://doi.org/10.1371/journal.ppat.1006258 https://doi.org/10.1371/journal.ppat.1006258
- 30.EZV https://doi.org/10.3390/v11020157 https://doi.org/10.3390/v11020157
- 31.Greer J M. Autoimmune T-cell reactivity to myelin proteolipids and glycolipids in multiple sclerosis. Mult Scler Int. 2013;2013:151427. doi: 10.1155/2013/151427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zamanzadeh Z, Ataei M, Nabavi S M, Ahangari G, Sadeghi M.In Silico Perspectives on the Prediction of the PLP ' s Epitopes involved in Multiple Sclerosis. Natl Inst Genet Eng Biotechnol [Internet]. 2017;15(1):10–21. Available from:http://dx.doi.org/10.15171/ijb.1356 [DOI] [PMC free article] [PubMed]
- 33.Wang B, Tan X F, Thurmond S.The structure of Zika virus NS5 reveals a conserved domain conformation Nat Commun 2017814763. 10.1038/ncomms14763[Internet] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kuhlmann T, Ludwin S, Prat A, Antel J, Brück W, Lassmann H. An updated histological classification system for multiple sclerosis lesions. Acta Neuropathol. 2017;133(01):13–24. doi: 10.1007/s00401-016-1653-y. [DOI] [PubMed] [Google Scholar]
- 35.Svejgaard A. The immunogenetics of multiple sclerosis. Immunogenetics. 2008;60(06):275–286. doi: 10.1007/s00251-008-0295-1. [DOI] [PubMed] [Google Scholar]
- 36.Alves-Leon S V, Papais-Alvarenga R, Magalhães M, Alvarenga M, Thuler L CS, Fernández y Fernandez O. Ethnicity-dependent association of HLA DRB1-DQA1-DQB1 alleles in Brazilian multiple sclerosis patients. Acta Neurol Scand. 2007;115(05):306–311. doi: 10.1111/j.1600-0404.2006.00750.x. [DOI] [PubMed] [Google Scholar]
- 37.Mangalam A K, Khare M, Krco C, Rodriguez M, David C. Identification of T cell epitopes on human proteolipid protein and induction of experimental autoimmune encephalomyelitis in HLA class II-transgenic mice. Eur J Immunol. 2004;34(01):280–290. doi: 10.1002/eji.200324597. [DOI] [PubMed] [Google Scholar]
- 38.Rich C, Link J M, Zamora A. Myelin oligodendrocyte glycoprotein-35-55 peptide induces severe chronic experimental autoimmune encephalomyelitis in HLA-DR2-transgenic mice. Eur J Immunol. 2004;34(05):1251–1261. doi: 10.1002/eji.200324354. [DOI] [PubMed] [Google Scholar]
- 39.Kaushansky N, Altmann D M, David C S, Lassmann H, Ben-Nun A. DQB1*0602 rather than DRB1*1501 confers susceptibility to multiple sclerosis-like disease induced by proteolipid protein (PLP) J Neuroinflammation. 2012;9(01):29. doi: 10.1186/1742-2094-9-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bielekova B, Sung M-H, Kadom N, Simon R, McFarland H, Martin R. Expansion and functional relevance of high-avidity myelin-specific CD4+ T cells in multiple sclerosis. J Immunol. 2004;172(06):3893–3904. doi: 10.4049/jimmunol.172.6.3893. [DOI] [PubMed] [Google Scholar]
- 41.Greer J M, Csurhes P A, Muller D M, Pender M P. Correlation of blood T cell and antibody reactivity to myelin proteins with HLA type and lesion localization in multiple sclerosis. J Immunol. 2008;180(09):6402–6410. doi: 10.4049/jimmunol.180.9.6402. [DOI] [PubMed] [Google Scholar]