

Abstract
Human-associated streptococci have not been viewed as productive sources of natural products. Against expectation, bioinformatic searches recently revealed a large collection of diverse biosynthetic gene clusters coding for ribosomally synthesized and post-translationally modified peptides (RiPPs) in streptococcal genomes. The most abundant of these, the tqq gene cluster, is specific to Streptococcus suis, a burdensome agricultural pathogen and zoonotic agent. Herein, we used high-throughput elicitor screening to identify both small molecule elicitors and products of the tqq cluster. We show that the B3 vitamin niacin effectively elicits the tqq cluster leading to the biosynthesis of a family of RiPP natural products, which we termed threoglucins and characterized structurally. The defining feature of threoglucins is an aliphatic ether bond giving rise to a substituted 1,3-oxazinane heterocycle in the peptide backbone. Isolation of 22 congeners of threoglucins facilitated structure activity relationship studies, demonstrating the requirement for the oxazinane substructure and a Trp-Tyr C-terminal dyad for biological activity, namely antibiotic persistence and allolysis at low and high doses, respectively. Potential therapeutic applications of threoglucins are discussed.
Microbial natural products account for the majority of clinically used antibiotics.1,2 Even after decades of mining through “grind and find” approaches, genome sequencing and bioinformatic analyses show that most natural products have yet to be discovered as evidenced by the large number of orphan biosynthetic gene clusters (BGCs).3 A significant challenge is that many microbial BGCs are transcriptionally silent or sparingly expressed when the producer is cultured under standard laboratory conditions.4 Strategies have been developed to activate silent BGCs, including high-throughput elicitor screening (HiTES), wherein inducers are identified from a library of small molecules in a forward chemical genetic screen.5–8 HiTES and other approaches have been successful in identifying novel metabolites from biosynthetically gifted organisms, but they are rarely applied to human or mammalian microbiota. These bacteria typically carry small, host-adapted genomes with fewer BGCs and are generally overlooked as sources of natural products.9–11 We recently identified a large network of BGCs coding for ribosomally synthesized and post-translationally modified peptides (RiPPs) from human-commensal and pathogenic streptococci (Figure 1A).12 The products from three of the ∼600 clusters have revealed new subclasses of RiPPs, including tryglysin A,13 streptide,14 and streptosactin,15 with unusual carbon–carbon or carbon–heteroatom linkages installed by radical S-adenosylmethionine (RaS) enzymes (Figure 1B). However, products from the majority of these subfamilies remain to be identified. The largest subfamily in the streptococcal RiPP network, termed the TQQ family based on a conserved amino acid motif in the precursor, encodes a 37mer peptide (TqqA), a RaS enzyme (TqqB), and a protease-transporter (TqqC) (Figure 1C). Previous studies have shown that TqqB catalyzes the formation of an ether cross-link by joining the threonine side chain oxygen to the α-carbon of the adjacent glutamine residue in TqqA.16 The tqq BGC is exclusively found in Streptococcus suis, a taxing agricultural pathogen and zoonotic agent.17 Interestingly, nearly half of the roughly 1620 unique S. suis strains within the NCBI database contain a tqq BGC, indicating it is prevalent and likely important for the overall fitness of this organism. Herein, we applied HiTES to S. suis using a human endogenous metabolite library and report both the product of the tqq cluster, a novel 1,3-oxazinane heterocycle-containing family of peptides termed threoglucins, as well as B3 vitamins as effective elicitors. We show, somewhat paradoxically, that threoglucins act as strong and highly selective antibiotics against S. suis with potential therapeutic applications.
Figure 1.
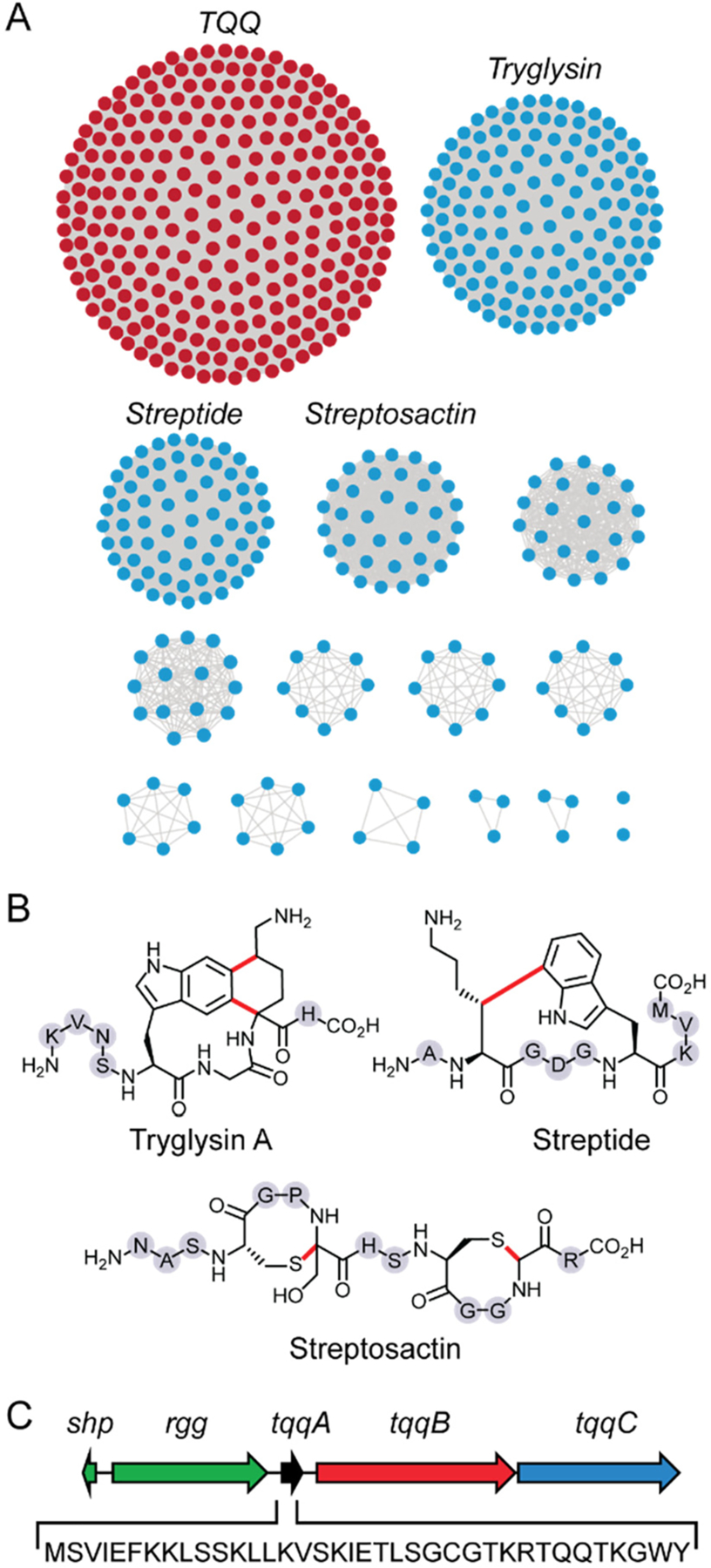
Streptococcal RaS-RiPP network and the tqq cluster. (A) RaS-RiPP network with each node representing a unique RiPP BGC. Subfamilies with known products are labeled. (B) Structures of mature RiPPs from the network. Gray spheres represent unmodified amino acids with the indicated one-letter code. (C) The tqq cluster is adjacent to an shp/rgg QS operon (green) and codes for a precursor peptide (TqqA, sequence shown), a RaS enzyme (TqqB), and a combined peptidase-transporters (TqqC).
The tqq cluster is downstream of an shp/rgg locus, which is thought to act as a quorum sensing (QS) regulatory element.18–20 We cultured S. suis to stationary phase in a chemically defined medium13,15 but were unable to confirm tqq-derived products under these conditions. We therefore attempted to elicit production using HiTES.5 Given the native context of S. suis, we chose a ∼442-member human endogenous metabolite library as possible elicitors. As readout, we resorted to UPLC-Qtof-MS, which allowed us to scan the ∼442 differentially elicited cultures in 36 h with a protocol optimized for detection of polar peptides. The features (ions with a specific m/z, abundance, and retention time) were extracted from each well and aligned in XCMS.21 Induced features were visualized through a 3D difference map generated by subtracting five-times the average of the vehicle-control feature abundance from the elicited conditions using MetEx.22 Positive values of the resulting difference matrix were displayed in a 3D plot, which shows the m/z and intensity for all cryptic metabolites as a function of elicitor (Figure 2A).
Figure 2.
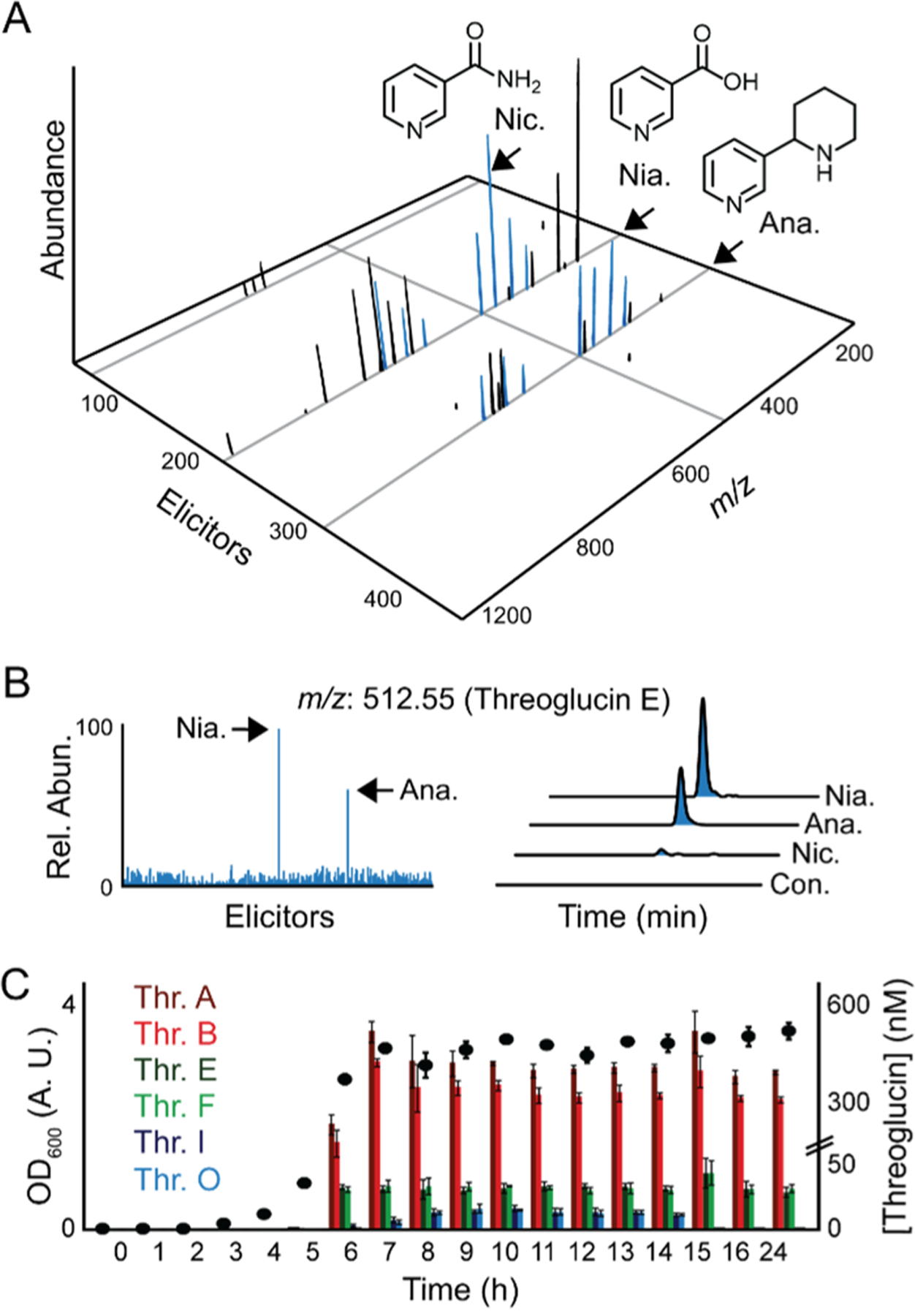
Activation of threoglucin production by HiTES. (A) 3D difference map showing residual features (m/z and abundance) as a function of elicitor. Peaks corresponding to threoglucins are colored blue; structures are shown for the top elicitors. (B, left) 2D slice from the 3D map in panel (A) showing abundance of threoglucin E across all 442 elicitors. Niacin and anabasine-treated samples are indicated. (B, right) Extracted ion chromatograms show relative abundance of threoglucin E from S. suis cultures treated with vehicle-control (Con.), nicotinamide (Nic.), anabasine (Ana.), and niacin (Nia.). (C) Quantification of threoglucins A, B, E, F, I, and O in niacin-treated S. suis cultures as a function of OD600.
The 3D map revealed a series of elicited peaks upon treatment with the B3 vitamins niacin and nicotinamide as well as anabasine, a compound containing the pyridine substructure. Several features, colored blue in Figure 2A, had accurate m/z values consistent with possible products from the tqq BGC, based on the precursor peptide sequence and the characterized reaction of TqqB.16 We focused on the most abundant of these from the network, which we later termed threoglucin E. A 2D slice from the 3D map shows that niacin and anabasine are the best elicitors for this compound (Figure 2B). Flask cultures validated induction by the pyridine-based elicitors and also revealed a larger family of related peptides, six of which we quantified in the presence of niacin as a function of optical density at 600 nm (OD600, Figure 2B,C). In flask cultures, two variants termed threoglucin A and B, which were significantly more nonpolar and therefore not captured in the 3D metabolite map, accumulated to 0.5 μM in stationary phase. Others, threoglucin C–R, reached ∼15–50 nM in stationary phase. The HiTES assay thus identified elicitors of possible tqq products and suggested that the biosynthetic pathway is highly divergent.
To determine the structures of threoglucins, production cultures (10 L) of S. suis were generated in the presence of niacin allowing isolation of 3.5 mg of a mixture of threoglucin A and B (threoglucin A/B), which was subjected to tandem high resolution mass spectrometry (HR-MS) and 1D/2D nuclear magnetic resonance (NMR) spectral analysis (Figure S1, Table S1–2). The former showed that threoglucin A is a 15mer peptide and located the modification to Thr8-Gln9, as all b ions downstream of b7 and all y ions upstream of y7 were –2 Da relative to the unmodified linear peptide (Figure 3A). Tandem HR-MS analysis showed that mature threoglucin A was trimmed only N-terminally relative to the full-length precursor. Total correlation spectroscopy (TOCSY) and heteronuclear multiple bond correlation (HMBC) spectra revealed key correlations, notably from the Gln9 β–1H to the Thr8 β-carbon that, together with 1H/13C chemical shift analyses, were consistent with an aliphatic ether linking the Thr8 side chain oxygen to the Gln9 α-carbon. This linkage gives rise to a substituted 1,3-oxazinane modification in the peptide backbone (Figure 3A). Threoglucin A is the first RiPP to contain this feature.
Figure 3.
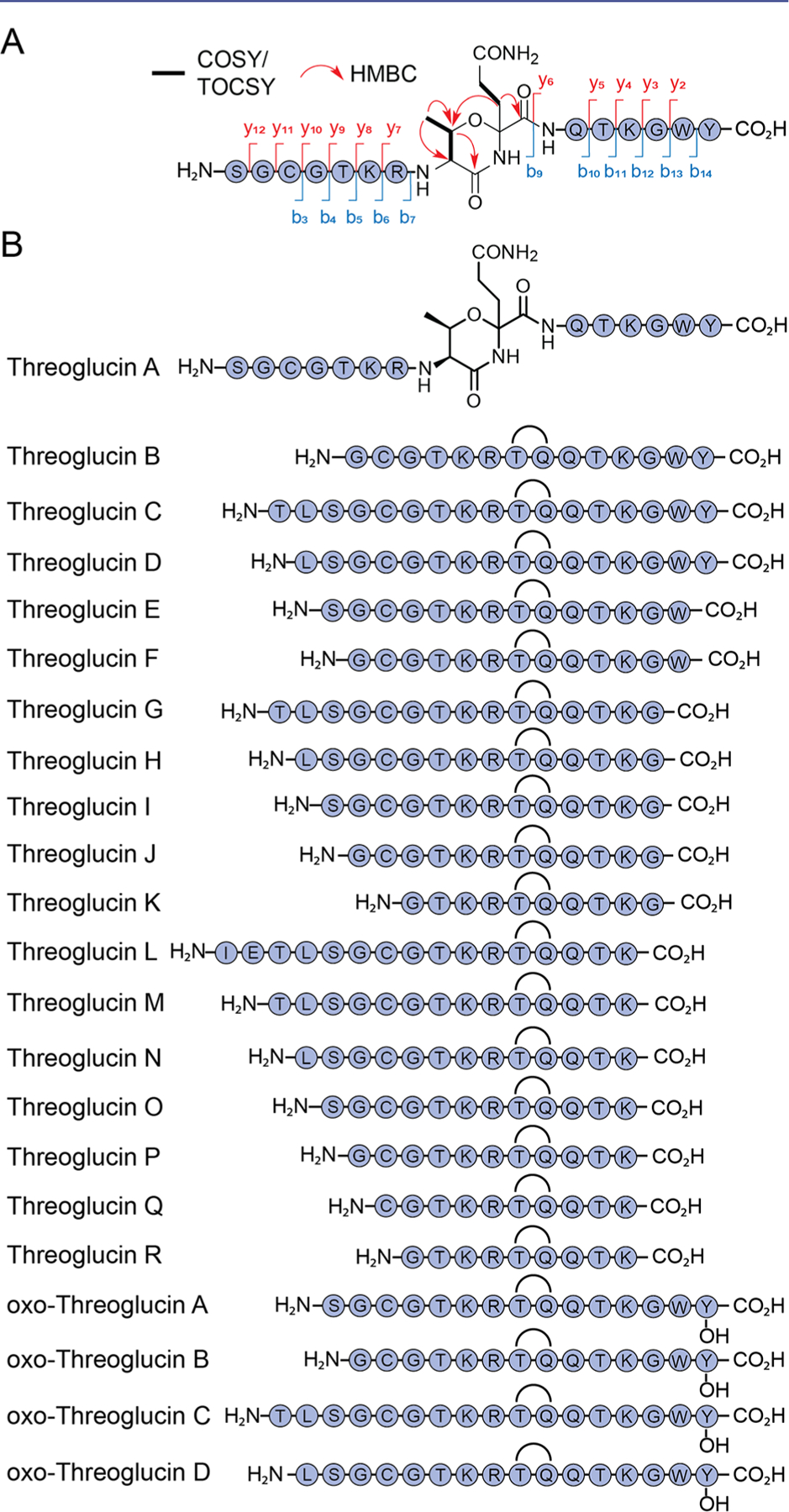
Structures of mature threoglucins. (A) Shown are relevant NMR correlations around the threonine-glutamine α-ether cross-link and observed MS/MS b- and y-ion fragments. (B) Structures of 22 threoglucin congeners. Gray spheres represent unmodified amino acids. Curved lines between threonine and glutamine represent the oxazinane heterocycle.
With the structure of threoglucin A at hand, we characterized 21 additional congeners by NMR spectroscopy and/or tandem HR-MS (Tables S3–S23). Seventeen of these (threoglucins B–R) contain the 1,3-oxazinane modification, but are trimmed further either N- or C-terminally (Figure 3B). Four additional variants, the oxo-threoglucins, contained a hydroxyl group on the C-terminal Tyr, though the small yields did not permit NMR-based structure elucidation. Interestingly, the timing of production of threoglucins is similar, suggesting that the tqq pathway generates these concurrently, rather than via proteolytic degradation from an initial product. Together, the results show that tqq directs the biogenesis of a divergent family of RiPPs containing an unusual, substituted heterocycle at the core with varying N- and C-terminal truncations. Threoglucins are the first structurally characterized RiPPs from S. suis.
Other RiPPs from the network have shown potent, narrow-spectrum inhibitory activities. Tryglysin A, for example, is a nM inhibitor of Streptococcus pneumoniae.13 Fratricidal activities have been attributed to streptosactin also at nM concentrations.15 With these in mind, we examined the effect of threoglucins against a panel of 19 different bacteria, including S. suis and 11 other streptococci. Threoglucin A/B exhibited an apparent half-maximal inhibitory concentration (IC50) of 0.5 μM only against S. suis but did not affect any other strain tested, even at a final concentration of 30 μM (Table S24). We conducted SAR studies with threoglucins A/B, C, E/F, and I/ O, and the linear 15mer peptide of threoglucin A that lacked the 1,3-oxazinane modification, which was prepared via solid-phase peptide synthesis. The results showed that both the C-terminal Trp-Tyr dyad and the 1,3-oxazinane were required for activity. The linear peptide with the Trp-Tyr dyad and the oxazinane-bearing peptide lacking the dyad or the terminal Tyr residue showed significantly reduced or no activity. Interestingly, threoglucin C showed no effects on S. suis, indicating the overall peptide length is also important for activity (Table S25).
The bioactivity results suggest S. suis is the target of threoglucins. Further examination revealed concentration-dependent effects with significant growth delays at 0.5–4 μM threoglucin A/B (Figure 4A); optical densities fully recovered, suggesting the peptide is bacteriostatic at these titers. Similarly, application of threoglucin A/B at early exponential phase resulted in rapid growth arrest, from which the bacteria recovered at low doses (Figure 4B). Treatment with high titers resulted in loss of viability; after 6 h exposure to 30 μM threoglucin A/B, <1% of the culture remained viable based on analysis of colony forming units, suggesting the RiPP is bactericidal at these concentrations (Figure 4C). No effects were observed against five different human cell lines at 60 μM (Table S24). Given the remarkable narrow-spectrum activity and the lack of effect on human cells in vitro, threoglucin A/B could be explored further as an S. suis antibiotic.
Figure 4.
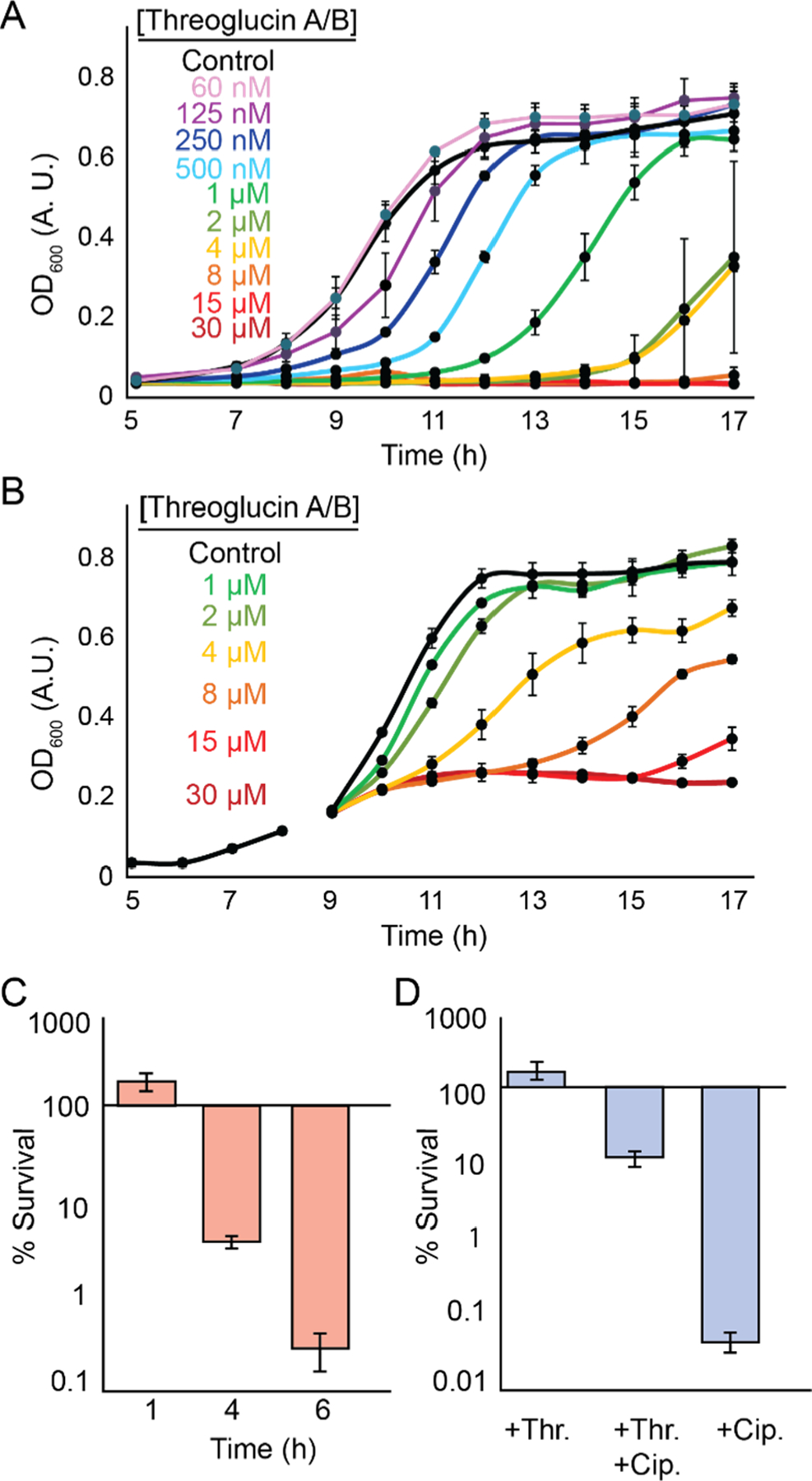
Biological activity of threoglucin A/B. (A) Growth curve for untreated (black) and threoglucin A/B-treated S. suis with concentrations ranging from 60 nM to 30 μM as shown. (B) Effect of threoglucin A/B addition at OD600 of 0.2 on the growth kinetics of S. suis. Traces are color-coded as shown. (C) Survival of S. suis, as measured by CFUs, after treatment of cultures with 30 μM threoglucin A/B at OD600 of 0.2. (D) Survival of S. suis after treatment during exponential phase (OD600 of 0.2) with 2 μM threoglucin A/B (+Thr.), 2 μM threoglucin A/B and 200 μM ciprofloxacin (+Thr. + Cip.) or 200 μM ciprofloxacin (+Cip.), as measured by CFU analysis. The results are scaled to growth of untreated cultures.
The recovery from low doses suggests threoglucin A/B can act as a growth-curbing or dormancy signal (Figure 4A), which can render cells tolerant to antibiotic treatment.23–25 We therefore tested the ability of S. suis to survive ciprofloxacin treatment in the absence and presence of threoglucin A/B. Remarkably, the presence of the RiPP at 2 μM resulted in ∼350-fold greater viability upon treatment with 200 μM ciprofloxacin (Figure 4D). These results suggest that threoglucin is a growth-curbing signal that can increase the tolerance of S. suis to antibiotics or other toxins, thereby providing an advantage only to the producing host, thus explaining its narrow-spectrum activity. Unnaturally high threoglucin concentrations act as selective bactericidal agents.
In summary, we describe the discovery, structural characterization, and initial biological activity of threoglucins, a family of RiPPs synthesized by a RaS enzyme in the opportunistic pathogen S. suis. Threoglucins are formed by installation of a simple aliphatic C–O bond in the peptide backbone, which transforms an otherwise unremarkable linear peptide into a highly potent, specific toxin (at high doses) or signal (at low doses). Given the production of this natural product by S. suis, susceptibility of only this species among those tested, and the conservation of the tqq cluster in S. suis genomes, threoglucins must play roles that confer a competitive advantage. The biological activity results suggest that threoglucins are dormancy signals that allow S. suis to survive the challenges of stationary phase growth, including assault by exogenous cytotoxins. Additional work to test this model, the molecular target of threoglucins, application of genetic methods to better understand how threoglucin production is regulated, and whether dormancy signals can be used as a source of species-specific antibiotics will be the subject of future studies.
Supplementary Material
ACKNOWLEDGMENTS
We thank Esther J. Han for assistance in testing the effect of threoglucins on human cell lines and the National Institutes of Health (GM140034 to M.R.S.) for financial support.
Footnotes
ASSOCIATED CONTENT
Supporting Information
The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/jacs.2c05790.
Detailed materials and methods, HR-MS, tandem HR-MS, and NMR spectral data for threoglucins, bioactivity assay results and IC50 measurements (PDF)
Notes
The authors declare no competing financial interest.
Complete contact information is available at: https://pubs.acs.org/10.1021/jacs.2c05790
Contributor Information
Brett C. Covington, Department of Chemistry, Princeton University, Princeton, New Jersey 08544, United States
Mohammad R. Seyedsayamdost, Department of Chemistry, Princeton University, Princeton, New Jersey 08544, United States; Department of Molecular Biology, Princeton University, Princeton, New Jersey 08544, United States
REFERENCES
- (1).Newman DJ; Cragg GM Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod 2020, 83, 770–803. [DOI] [PubMed] [Google Scholar]
- (2).Cragg GM; Newman DJ Natural products: a continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Nett M; Ikeda H; Moore BS Genomic basis for natural product biosynthetic diversity in the actinomycetes. Nat. Prod. Rep 2009, 26, 1362–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (4).Amos GCA; Awakawa T; Tuttle RN; Letzel A-C; Kim MC; Kudo Y; Fenical W; Moore BS; Jensen PR Comparative transcriptomics as a guide to natural product discovery and biosynthetic gene cluster functionality. Proc. Natl. Acad. Sci. U.S.A 2017, 114, E11121–E11130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (5).Seyedsayamdost MR High-throughput platform for the discovery of elicitors of silent bacterial gene clusters. Proc. Natl. Acad. Sci. U.S.A 2014, 111, 7266–7271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Xu F; Wu Y; Zhang C; Davis KM; Moon K; Bushin LB; Seyedsayamdost MR A genetics-free method for high-throughput discovery of cryptic microbial metabolites. Nat. Chem. Biol 2019, 15, 161–168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (7).Moon K; Xu F; Zhang C; Seyedsayamdost MR Bioactivity-HiTES unveils cryptic antibiotics encoded in Actinomycete bacteria. ACS Chem. Biol 2019, 14, 767–774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (8).Moon K; Xu F; Seyedsayamdost MR Cebulantin, a cryptic lanthipeptide antibiotic uncovered using bioactivity-coupled HiTES. Angew. Chem., Int. Ed. Engl 2019, 58, 5973–5977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Cimermancic P; Medema MH; Claesen J; Kurita K; Wieland Brown LC; Mavrommatis K; Pati A; Godfrey PA; Koehrsen M; Clardy J; Birren BW; Takano E; Sali A; Linington RG; Fischbach MA Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Donadio S; Monciardini P; Sosio M Polyketide synthases and nonribosomal peptide synthetases: the emerging view from bacterial genomics. Nat. Prod. Rep 2007, 24, 1073–1109. [DOI] [PubMed] [Google Scholar]
- (11).Gavriilidou A; Kautsar SA; Zaburannyi N; Krug D; Müller R; Medema MH; Ziemert N Compendium of specialized metabolite biosynthetic diversity encoded in bacterial genomes. Nat. Microbiol 2022, 7, 726–735. [DOI] [PubMed] [Google Scholar]
- (12).Bushin LB; Clark KA; Pelczer I; Seyedsayamdost MR Charting an unexplored streptococcal biosynthetic landscape reveals a unique peptide cyclization motif. J. Am. Chem. Soc 2018, 140, 17674–17684. [DOI] [PubMed] [Google Scholar]
- (13).Rued BE; Covington BC; Bushin LB; Szewczyk G; Laczkovich I; Seyedsayamdost MR; Federle MJ Quorum sensing in Streptococcus mutans regulates production of tyglysin, a novel RaS-RiPP antimicrobial compound. mBio 2021, 12, No. e02688–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).Schramma KR; Bushin LB; Seyedsayamdost MR Structure and biosynthesis of a macrocyclic peptide containing an unprecedented lysine-to-tryptophan crosslink. Nat. Chem 2015, 7, 431–437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (15).Bushin LB; Covington BC; Rued BE; Federle MJ; Seyedsayamdost MR Discovery and biosynthesis of streptosactin, a sactipeptide with an alternative topology encoded by commensal bacteria in the human microbiome. J. Am. Chem. Soc 2020, 142, 16265–16275. [DOI] [PubMed] [Google Scholar]
- (16).Clark KA; Bushin LB; Seyedsayamdost MR Aliphatic ether bond formation expands the scope of radical SAM enzymes in natural product biosynthesis. J. Am. Chem. Soc 2019, 141, 10610–10615. [DOI] [PubMed] [Google Scholar]
- (17).Fittipaldi N; Segura M; Grenier D; Gottschalk M Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol 2012, 7, 259–79. [DOI] [PubMed] [Google Scholar]
- (18).Fleuchot B; Gitton C; Guillot A; Vidic J; Nicolas P; Besset C; Fontaine L; Hols P; Leblond-Bourget N; Monnet V; Gardan R Rgg proteins associated with internalized small hydrophobic peptides: a new quorum-sensing mechanism in streptococci. Mol. Microbiol 2011, 80, 1102–19. [DOI] [PubMed] [Google Scholar]
- (19).Junges R; Salvadori G; Shekhar S; Åmdal HA; Periselneris JN; Chen T; Brown JS; Petersen FC A quorum-sensing system that regulates Streptococcus pneumoniae biofilm formation and surface polysaccharide production. mSphere 2017, 2, No. e00324–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (20).Neiditch MB; Capodagli GC; Prehna G; Federle MJ Genetic and structural analyses of RRNPP intercellular peptide signaling of Gram-positive bacteria. Annu. Rev. Genet 2017, 51, 311–333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (21).Smith CA; Want EJ; O’Maille G; Abagyan R; Siuzdak G XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem 2006, 78, 779–787. [DOI] [PubMed] [Google Scholar]
- (22).Covington BC; Seyedsayamdost MR MetEx, a metabolomics explorer application for natural product discovery. ACS Chem. Biol 2021, 16, 2825–2833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (23).Lemma AS; Brynildsen MP Toxin induction or inhibition of transcription or translation posttreatment increases persistence to fluoroquinolones. mBio 2021, 12, No. e01983–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (24).Dorr T; Vulić, M.; Lewis, K. Ciprofloxacin causes persister formation by inducing the TisB toxin in Escherichia coli. PLoS Biol 2010, 8 (2), No. e1000317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (25).Rittershaus ESC; Baek S-H; Sassetti CM The normalcy of dormancy: common themes in microbial quiescence. Cell Host Microbe 2013, 13, 643–651. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.