

Abstract
Therapeutically popular controlled release-enabling technology has forayed into the nutrition sector. Polymer coated forms of L-methionine used in soy protein diets, and its intermediate metabolite, S-adenosyl-L-methionine, used in myriad of medical conditions have proved more efficacious over (highly catabolized) free forms. In this premier study, L-methionine-loaded chitosan nanoparticles (M-NPs) were synthesized using ionic gelation method and their efficacy was evaluated. Biophysical characterization of the NPs was done using a Nanopartica SZ 100 analyser, transmission electron microscopy, and Fourier transform infrared spectroscopy. The M-NPs were spherical and smooth and 218.9 ± 7.4 nm in size and in vitro testing confirmed the controlled release of methionine. A 60-days feeding trial in L. rohita fish fingerlings was conducted. A basal diet suboptimal (0.85%) in methionine was provided with one of the supplements as under: none (control), 0.8% chitosan NPs (0.8% NPs), 1.2% L-methionine (1.2% M) (crystalline free form), 0.6% M-NPs and 1.2% M-NPs. While the addition of 0.6% M-NPs to the basal diet complemented towards meeting the established dietary requirement and resulted in significantly highest (P < 0.05) growth and protein efficiency and sero-immunological test scores (serum total protein, serum globulin, serum albumin: globulin ratio, phagocytic respiratory burst/NBT reduction and lysozyme activity), 1.2% supplementation in either form (free or nano), for being 0.85% excess, was counterproductive. Liver transaminases and dehydrogenases corroborated enhanced growth. It was inferred that part of the methionine requirement in nano form (M-NPs) can confer intended performance and health benefits in animals relying on plant proteins-based diets limiting in this essential amino acid. The study also paves the way for exploring chitosan NPs-based sustained delivery of amino acids in human medical conditions.
Subject terms: Physiology, Nanoscience and technology
Introduction
Animal-based and plant-based proteins meet the global demand for proteins. Animal proteins have superior amino acids profile, but their cost prohibits usage in farm species diets, despite preference, and are anathema to vegetarian people. Fishmeal is the most preferred protein in the diets of farm animals, especially fish and shrimp, in semi-intensive and intensive aquaculture systems that rely on a large quantity of aquafeed for optimal performance. While it is added to feed to stimulate intake in young fish and or to balance protein content, the currently stagnated fish meal production clearly predicts an unsustainable scenario, wherein, proteins from marine sources will no longer meet the demands of the aquafeed industry. Globally, there is a paradigm shift in feed production practices, which have already switched from marine to plant ingredients1,2 with the usage of feeding stimulants. The feed industries are extensively reliant on proteins from legumes and oilseeds. However, these plant proteins are poorly utilised due to the presence of anti-nutritional factors and often are not sufficient in one or other essential amino acids. For example, despite the numero uno position of soybean meal for being the preferred and predominantly used source of plant protein in fish feeds, it is limiting in methionine (cells use only L-forms). Methionine is known to enhance feed intake and promote growth in fish3 and acts as a methyl donor for DNA methylation4–6. Deficiency of methionine, from exclusive or extensive use of soymeal (i.e., no or low fish meal) in diets is corrected by supplementation of a synthetic source of L-methionine.
Across species, the use of free crystalline amino acids helps to make up dietary deficits and devise optimum environmental friendly diets7,8, but poses limitations as they are absorbed and catabolised faster due to temporal mismatch in availability at the sites protein synthesis9. In addition, they are prone to leaching in water when used in aquatic feed and are potentially degraded in the rumen in herbivores. As such, dietary crystalline methionine in its free form is not very efficient10 and is economically expensive and requires 10% more than the NRC requirement to achieve growth in fish11. Secondly, increasing the amount of methionine in the diet may not be an alternative or good option, because the excess amount could turn toxic12 and increase the feed cost. Further, a low dose, long-term supplementation of free amino acids is better utilised than short-term high dose13, but can compromise dietary levels. Thus, the best strategy that emerges is the use of slow-release systems for free amino acids. While Guo et al.14 opined that supplementing methionine in coated or microform would improve its bioavailability, Yúfera et al.15 reported a 68% reduction in the leaching of free amino acids (FAA) into the water in 1 h with the use of the microencapsulated diet over a gelatin microbound diet.
In humans, S-adenosyl-L-methionine (SAMe) therapy has been suggested and positive results have been obtained in diseases such as depression, dementia, liver diseases (cholestasis, steatohepatitis, cirrhosis), pancreatitis, vacuolar myelopathy, osteoarthritis and type II diabetes16–18. Because only 1% of the administered dose reaches the circulation due to its degradation in the gastrointestinal tract, transit in the liver and low permeability, the recommended dose of 800–1600 mg a day for a variety of clinical conditions cannot be met. While SAMe is relatively expensive and unstable, its precursor, methionine can be a good choice for supplementation. However, there are only a few studies on improving its bioavailability. Controlled release systems over coated amino acids are also desired for people with inborn errors in amino acid metabolism e.g. phenylketonuria19–21.
Cellulose-acetate-phthalate, acrylic-resin, tripalmitin-polyvinyl alcohol22, or natural materials like alginate, carrageenan, agar, gum arabic and zein have been used for coating amino acids to improve bioavailability, but reports on nanocomplexes and their in vivo evaluation are scarce. The unique size-dependent property of nanoparticles (NPs) enables the delivery of virtually any small molecule for targeted disease and health outcomes. Classified by the FDA as generally regarded as safe (GRAS), biodegradable polymers are used as a promising carrier for the delivery of biomolecules and drugs23,24. Chitosan, a natural biopolymer, is considered to be the most suitable and sustainable nanocarrier of pharmaceutically and nutritionally active compounds for oral delivery because of its non-toxic, non-irritant nature25, biocompatibility and mucoadhesive property26–29. It shields the load from the adverse conditions of the gastrointestinal tract30. Chitosan is cationic in nature which makes it soluble across wide pH gradients in acidic and basic solutions. The addition of polyanionic compounds in chitosan solution under specific conditions would lead to the formation of NPs by ionic gelation or poly-electrolyte complexing31. Encapsulation of therapeutic drugs in chitosan NPs is known to not only protect them from leaching and physiological barriers of the gut32 but also enable their controlled release and absorption33 and cellular uptake34,35. We have synthesized chitosan nanoparticles and used them for delivery of proteolytic enzymes36 and RNA37 orally and hormone38 through injection with beneficial effects in fish. There are only a few studies on improving its bioavailability. Earlier, Ergin, et al.39 synthesized and evaluated in vitro chitosan NP-based systems for colon targeted delivery of S-adenosyl-L-methionine. In this pioneering study, methionine-loaded chitosan NPs were fabricated and tested in vitro for methionine release and in vivo for their ability to promote growth performance and immune profile of a model fish, L. rohita, which is one of the major cultivable species in freshwater aquaculture.
Materials and methods
Chemicals
Low molecular weight chitosan (MW < 200 kDa) with a deacetylation degree of 90% and the sodium salt of tripolyphosphate were purchased from Sigma-Aldrich (Saint Louis, MI, USA). Purified feed ingredients were procured from Hi-Media (Hi-Media Laboratories, Mumbai, India). Other chemicals and consumables used in the study were of analytical grade.
Preparation of chitosan nanoparticles
Chitosan nanoparticles were prepared by ionic gelation method31. Five different ratios of chitosan to L-methionine (1:0.5, 1:1, 1:1.5, 1:2, and 1:2.5) were employed to make L-methionine-loaded chitosan NPs. 0.2 g of purified chitosan flakes were dissolved in 10 ml of 10% acetic acid. A pH of 5.74 was reached by adding 0.2 M sodium hydroxide solution, and the total volume was made up to 100 ml bringing the final concentration of chitosan to 0.2%. The solution was sterile filtered using a 0.22 µm syringe filter. Sodium tripolyphosphate (TPP) (0.7 mg ml−1) solution was then added dropwise to the chitosan solution to allow the formation of nanoparticles. Loading of L-Methionine (referred to without rotational conformation hereafter) was done by dissolving it in the chitosan solution before adding TPP. The nanoparticles were freeze-dried for experimental diet preparation.
Characterisation of nanoparticles
Measurement of particle size
The hydrodynamic mean diameter of the nanoparticles and their size distribution was determined by a dynamic light scattering (DLS)- nanoparticle size analyser, Nanopartica SZ-100 (Horiba Ltd, Kyoto, Japan). All measurements were performed in a wavelength of 633 nm at 25 °C with an angle detection of 90°.
Determination of zeta potential
The zeta potential measurements were made using Nanopartica SZ-100 (Horiba Ltd, Kyoto, Japan) by applying the principle of Laser Doppler Electrophoresis. Lyophilised samples were diluted in deionised water and determined in triplicates.
Transmission electron microscopy (TEM)
Transmission Electron Microscopy (TEM) (FEI Tecnai G2 F30) at Sophisticated Analytical Instruments Facility (SAIF), Indian Institute of Technology (IIT), Mumbai, was used to observe the morphology of chitosan nanoparticles. The lyophilised nanoparticles were suspended in deionised water to 1/100 (v/v) and stained with 2% phosphotungstic acid (a negative stain) 1 h before observation.
FT-IR spectroscopy
A Fourier transform infrared spectrophotometer (FT-IR, Bruker, Germany) of the Sophisticated Analytical Instrument Facility (SAIF) at the Indian Institute of Technology (IIT), Mumbai, was used to investigate the interaction of functional groups between methionine and chitosan. The lyophilised nanoparticles were taken with KBr pellets and pressed to a plate for this reason.
Determination of encapsulation efficiency
The optimum loading capacity for methionine during the ionic gelation procedure was determined by calculating the encapsulation efficiency of chitosan nanoparticles- loaded with methionine in ratios of 1:0.5, 1:1, 1:1.5, 1:2 and 1:2.5. The encapsulation efficiency (EE) of the chitosan NPs was determined as the percentage ratio of the encapsulated methionine to the total methionine according to the equation; EE% = Encapsulated methionine/Total methionine × 100. The amount of the encapsulated methionine was indirectly determined as the difference between the total amount of methionine initially added and that of the free and unencapsulated methionine. For this, free methionine in the aqueous medium was separated from nano-encapsulated methionine by ultracentrifugation at 40000×g at 4 °C for 45 min and the free methionine in the supernatant was measured by the nitroprusside method40. Briefly, 1 ml of 5N NaOH and 0.5 ml of 1% sodium nitroprusside were added to 1 ml of the supernatant. Samples were subjected to photometric measurements at 520 nm in a UV–VIS spectrophotometer (Shimadzu UV 1601 spectrophotometer, Tokyo, Japan) after 10 min of incubation at room temperature.
In vitro release study
In vitro release of methionine was tested on nanoparticles formulated using a 1:1.5 chitosan to methionine ratio for their highest encapsulation efficiency (83.63 ± 1.67%) among combinations tried. The release rate of methionine from the chitosan NPs was measured in phosphate- buffered solution (pH 7.4) at 37 °C in triplicate. Briefly, 10 mg of freeze-dried nanoparticles having the desired load of methionine were taken in screw-capped tubes and dispersed in a 10 mL phosphate-buffered saline (PBS) solution. The tubes were placed in a shaking water bath (Julabo, SW3; Seelbach, Germany) maintained at 37 °C and shaken horizontally at 90 rpm. The tubes were taken out of the water bath at predetermined time intervals and centrifuged at 40000×g for 30 min at 25 °C. The supernatant was carefully collected and replaced with an equal volume of fresh buffer solution. The quantity of released methionine in the supernatant was analysed by the nitroprusside method.
Fish and feeding trial
Fingerlings of Labeo rohita species were brought from the local fish farm and acclimatized in a circular tank (2000 L) for 15-days and fed a diet with 30% crude protein. The feeding experiment was conducted in the wet laboratory for 60-days in FRP tanks having 150 L capacity. The tanks were arranged as per a completely randomised design and there were three for each treatment and four replicates for the control group. Fifteen acclimatized fingerlings of L. rohita (mean weight 2.8 ± 0.41 g) were stocked in each tank. The experimental feeding was carried out twice a day at 9:00 a.m. and 4:00 p.m. up to satiation level and uneaten feed was siphoned out. All fish from each experimental tank were weighed every 15 days to assess the biomass and adjust feeding rate calculation during the trial period. During the study period, the water quality parameters were in the optimal range required for the L. rohita fingerlings and are presented in Table S1. The research conducted was approved by the statutory authorities of the ICAR-Central Institute of Fisheries Education, Mumbai 400,061. Guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA, Ministry of Environment & Forests, Animal Welfare Division, Government of India) were followed. The provisions of the Wildlife Protection Act of 1972 are not applicable for experiments on L. rohita as it is not an endangered fish.
Experimental diets
The basal control diet containing about 31.2% crude protein and 3530 kcal/kg digestible energy (DE) was formulated using purified ingredients (Table 1). To make experimental diets, an equal quantity of a non-nutritive ingredient, cellulose, was replaced by the item that defined treatment (0.8% chitosan NPs, 1.2% free L-methionine, 0.6% M-NPs and 1.2% M-NPs. All the ingredients were weighed accurately as per the requirements based on their percent content in the diet. The first six ingredients (see Table 1) were appropriately mixed to make a dough and steam cooked for 30 min. The rest of the ingredients along with methionine, chitosan nanoparticles, or L-methionine-loaded chitosan nanoparticles were sprayed into the steam -cooked dough after cooling. Nanoparticles used in the experimental diet were synthesized using chitosan to methionine in the ratio of 1:1.5. The amount of methionine in the nanoparticles used for diet preparation was ensured by calculating the encapsulation efficiency. The composition of all five diets used in the study is given in Table 1. Two levels (0.6 and 1.2%) of sustained release nano-methionine were selected for supplementation of the basal diet that contained 0.85% indigenous protein-bound methionine estimated following standard methods, McCarthy and Sullivan40 and Kader et al.41. One level was set close to safely meeting the reported methionine requirement of 1.15% to 1.2%42–44 for carps on a similar type of diet. The dough was pressed through a pelletizer to get uniform-sized pellets, which were spread on a sheet and air-dried. After drying, the pellets were packed in cryo bags and stored in the deep freezer.
Table 1.
Ingredient composition and sulphur amino acid content (%) of the experimental diets.
| Ingredients | Control | 0.8% NPs | 1.2% M | 0.6% M-NPs | 1.2% M-NPs |
|---|---|---|---|---|---|
| Casein | 33.00 | 33.00 | 33.00 | 33.00 | 33.00 |
| Gelatin | 8.25 | 8.25 | 8.25 | 8.25 | 8.25 |
| Dextrin | 16.75 | 16.75 | 16.75 | 16.75 | 16.75 |
| Starch soluble | 20.38 | 20.38 | 20.38 | 20.38 | 20.38 |
| Cellulose | 10.07 | 9.27 | 8.87 | 8.67 | 7.91 |
| CMC | 1.50 | 1.50 | 1.50 | 1.50 | 1.50 |
| Cod liver oil | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| Sunflower oil | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| Ascorbic acid | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 |
| Vit.-Min. Premixa | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| BHT | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| Chitosan NPs (free) | 0.80 | 0.32 | |||
| L-methionine | 1.20 | ||||
| M-NPs complexb | |||||
| Methionine | 0.60 | 1.20 | |||
| Chitosan NPs | 0.48 | 0.96 | |||
| Total | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Total methionine (%) | 0.85 | 0.85 | 2.05 | 1.45c | 2.05d |
| Cystine (%) | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 |
| Proximate analysise and other LAAf | |||||
CMC carboxymethyl cellulose, BHT butylated hydroxytoluene.
aComposition of the standard premix in our lab is described in detail earlier47.
bM-NPs-L-Methionine loaded Chitosan NPs (Chitosan to methionine ratio of 1:1.5).
c, d Of the total percent content, 0.6% and 1.2% (nano) methionine is released slowly.
eAll the feeds were isonitrogenous and isocaloric and contained on DM basis: crude protein 31.19 ± 2.6, ether extract 8.83 ± 0.07, total CHO (TC) 51.03 ± 0.40, ash 8.14 ± 0.29, and digestible energy (DE) 353.10 ± 3.14 kcal.
DE (Kcal/100 g) = (% CP × 4) + (% EE × 9) + (TC × 4). (n = 3), (As per AOAC method).
fOther limiting amino acid contents: lysine: 2.66%, tryptophan: 0.019% and arginine: 2.02%.
Amino acid composition of experimental diets
The method of Kader et al. was followed. In brief, a 100 mg dry sample was weighed and placed into clean dry hydrolysis glass tubes (Thermo Fisher 29,571), which was then filled with 1 mL of 6N hydrochloric acid. After that, all of the air was evacuated from the tubes using a vacuum pump, and the tubes were filled with nitrogen before sealing tightly. The sealed hydrolyzing tubes were then heated for 24 h on a dry block heater at 110 °C. The tubes were then removed from the block heater and allowed to cool to room temperature. The 10 mL cooled digested sample was then transferred to a 15 mL centrifuge tube with the help of a 3 mL syringe fitted with a long stainless-steel needle, and the hydrolyzing tube was cleaned by washing out the particulate matter with 9 ml of BioChrom loading buffer. The pH of a 10 mL sample was then adjusted to between 2 and 2.2 using 7.5 M NaOH solution and filtered through a 25 mm syringe filter. Then, a 200 µL filtered sample was placed into a 1.5 mL sample tube along with 800 µL BioChrom loading buffer. This tube was then placed in the autosampler, and the amino acid concentrations of samples were determined using an automated amino acid analyzer (BioChrom 30 +, Biochrom Ltd, United Kingdom).
Sampling and analytical methods
At the end of the 60-days feeding trial, five fish from each tank were collected and anaesthetised with clove oil (50 ml/L) for the collection of blood samples. While quantification of respiratory burst activity/ nitroblue tetrazolium (NBT) reduction45 and blood glucose (Somogyi method, see46) was done immediately on some samples, serum was separated from the rest for estimation of lysozyme activity47, total serum myeloperoxidase content48 and serum complement-3 (C3) (ELISA kit) (CUSABIO, Houston, TX USA), serum total protein and albumin (Qualigens Diagnostics, Mumbai, India). Serum globulin was calculated by subtracting albumin from total serum protein. Fish were sacrificed and muscle and liver tissues from the same fish were aseptically dissected out and kept in an appropriate buffer with 0.25% sucrose as cryoprotectant and stored in a deep freezer. Tissue samples were homogenised using phosphate buffer (pH 7) containing 0.25% sucrose and 0.1 X Halt™ protease inhibitor and the supernatant was collected after centrifuging at 7000×g. Activities of aspartate aminotransaminase (AST) (E.C. 2.6.1.1) and alanine aminotransaminase (ALT) (E.C. 2.6.1.2), lactate dehydrogenase (LDH; L-lactate NAD+ oxidoreductase: E.C. 1.1.1.27) malate dehydrogenase (MDH; L-malate: NAD+ oxidoreductase: EC.1.1.1.37) were measured as described earlier45,49. Myotomal muscle glycogen content was determined following KOH (30%) digestion of weighed sample at 100 °C for 15 min. Ethanol was used to precipitate glycogen from an aliquot of the digest50, and the precipitated glycogen was assayed by the anthrone method51.
Data collection and growth efficiency study
Growth performance of fingerlings was evaluated in terms of weight gain (%), specific growth rate (SGR), food conversion ratio (FCR), and protein efficiency ratio (PER) based on the following standard formulae: Weight gain (%) = (Final weight − initial weight)/initial weight × 100; SGR (% day−1) = (Loge final weight − Loge initial weight)/number of culture days × 100; FCR = total dry feed intake(g)/wet weight gain(g) and PER = total weight gain(g)/crude protein fed (g).
Statistical analysis
Tanks (with 15 fish in each) constituted an experimental unit for data on growth, while pooled samples (e.g. serum) from fishes were a unit for the rest of the data such as biochemical and immuno-hematological measurements. The data were analysed using the IBM Statistical Package, SPSS, version 22.0 (SPSS Inc., Chicago, IL). The data tested for normality (Shapiro–Wilk’s test) and homogeneity of variance (Levene’s test) were subjected to one-way ANOVA. The significant levels of difference for all measured parameters were calculated and means were compared by Duncan’s Multiple Range Test at a 5% probability level. The p value < 0.05 was considered statistically significant.
Results
Synthesized L-methionine-loaded chitosan nanoparticles have desired physical characteristics
A comparison of the physical and chemical characteristics of the synthesized chitosan and methionine-loaded chitosan nanoparticles (NPs) is given in Table 2. During the process of making M-NPs, as the proportion of methionine increased from 0.5 to 2.5 in fixed 1 part of chitosan NPs, the size of chitosan M-NPs kept on increasing (P = 0.000), but zeta potential (P = 0.036) and encapsulation efficiency (EE%) (P = 0.000) increased until methionine proportion was 1.5, where optimum loading capacity was found. However, zeta potential and encapsulation efficiency were decreased when the methionine proportion was highest (NPs: Met ratio, 1:2.5). Following encapsulation of methionine in chitosan NPs the polydispersity index was reduced from 0.72 to 0.52 (Fig. 1A,B). Transmission electron spectroscopy (TEM) images of methionine-loaded chitosan nanoparticles (Fig. 1C,D) exhibited almost spherical and dispersed particles in the medium.
Table 2.
Characteristics of methionine-loaded chitosan nanoparticles.
| Chitosan TPP NP: methionine ratio | Size (nm) | Zeta potential | Encapsulation efficiency |
|---|---|---|---|
| Chitosan (NPs) blank (1:0) | 173.26a ± 12.1 | 28.6a ± 3.7 | – |
| NPs: Met (1:0.5) | 180.09a ± 3.67 | 27.77a ± 1.23 | 51.78a ± 1.92 |
| NPs: Met (1:1.0) | 208.9b ± 1.53 | 25.33a ± 1.76 | 65.86c ± 1.4 |
| NPs: Met (1:1.5) | 218.9bc ± 7.4 | 31.37b ± 1.6 | 83.63e ± 1.67 |
| NPs: Met (1:2.0) | 232.07cd ± 4.79 | 28.33ab ± 1.45 | 74.69d ± 2.06 |
| NPs: Met (1:2.5) | 252.33d ± 14.13 | 25.33a ± 1.76 | 57.78b ± 1.83 |
Chitosan NPs: Methionine ratio was varied by using fixed chitosan nanoparticle concentration and different methionine concentration (0, 0.4, 0.8, 1.2, 1.6 and 2%).
Values are represented as mean ± standard error (n = 3).
a,b,c,d,eMeans bearing different superscript letters in the same column differs significantly (P < 0.05).
Figure 1.

Particle size distribution. (A) Chitosan nanoparticles; (B) Methionine encapsulated chitosan nanoparticles; (C) Transmission electron microscopic images of the non-encapsulated form of chitosan nanoparticles; and (D) Chitosan nanoparticles after encapsulation of methionine.
FT-IR spectra confirm L-methionine loading onto chitosan NPs
Spectral analysis using FT-IR revealed potential interactions of L-methionine with chitosan nanoparticles. The spectral broadband wavelength for L-methionine, chitosan, chitosan NPs and L-methionine-loaded chitosan NPs (M-NPs) ranged from 3500 to 3300 cm−1 (See Fig. 2). A peak at 3454 cm−1 in chitosan was shifted to 3427 cm−1 in nanoparticulate forms. The peak of a broad band at 3150 cm−1 in the L-methionine spectrum disappeared and was not visible in M-NPs. The peak at 2914 cm−1 in L-methionine and 2968 in chitosan NPs was shifted to 2926 cm−1 in M-NPs. A small peak at 2600–2550 cm−1 was found only in L-methionine and L-methionine-loaded nanoparticles spectra. The peak at 1609 cm−1 in L-methionine and at 1643 cm−1 in chitosan NPs was found at 1641 in M-NPs. The peak at 1514 cm−1 in L-methionine is reflected in M-NPs as well. A peak found in L-methionine at 746 cm−1 disappeared in M-NPs. Peaks at 644, 660 and 764 cm−1 were visible in both L-methionine and M-NPs.
Figure 2.

FT-IR spectra of L-methionine, chitosan, chitosan nanoparticle (NPs) and L-methionine loaded chitosan nanoparticle (M-NPs).
Slow and controlled release of L-methionine from chitosan nanoparticle in vitro
The nanoparticles having desired characteristics showed a consistently slow-release rate with only 10% release until 12 h of incubation, which increased faster in the next 6 h reaching a peak (23%) at 18 h (Fig. 3) and then slowly decreasing thereafter until 42 h. Of the loaded amount, 16%, 40%, and 86.5% total methionine was released at the end of 12, 18, and 42 h, respectively.
Figure 3.
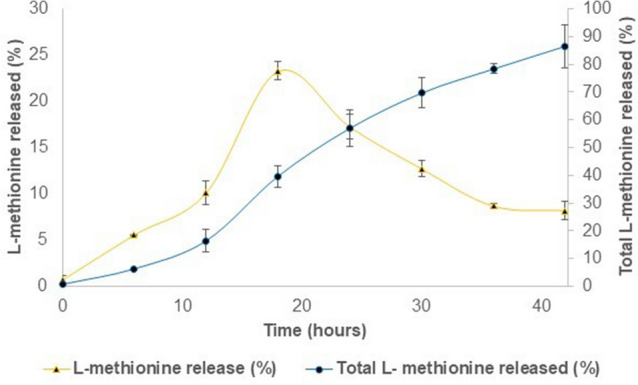
In vitro release profiles of L-methionine from chitosan nanoparticles. Chitosan to methionine ratio used for the study was 1:1.5. The proportion of L-methionine released to the initial L-methionine load is indicated in the secondary axis as total L-methionine released (%.) Proportion of L-methionine released to the Methionine content after each sampling is L-methionine release (%). Results were reported as mean ± standard error, n = 3.
Superior performance in fish when nano-methionine partly covers the total methionine requirement
Growth and performance responses of L. rohita fish given different dietary treatments are depicted in Fig. 4A–D. Compared to the control, the addition of chitosan NPs, 0.6% M-NPs and 1.2% methionine in free (1.2%M) or nano form (1.2%M-NPs) to the basal diet improved performance parameters such as percent weight gain (%WG), specific growth rate (SGR), feed conversion ratio (FCR) and protein efficiency ratio (PER) (P = 0.000 for %WG and SGR and P = 0.001 for FCR and PER). While chitosan NPs improved (P < 0.05) % WG, SGR and PER of fish, the effect was significantly potentiated (P < 0.05) on 0.6% methionine-loaded chitosan NPs (0.6% M-NPs), which together with casein-bound methionine helped fulfil the required dietary concentration, but the performance of 1.2% M-NPs was alike methionine in free form (1.2% M), the level that contributed 0.85% excess methionine. From the third week to the eighth week, the differences in per cent weight gain between 0.6% M-NPs supplemented methionine adequate and 1.2% M (free form) and 1.2% M-NPs supplemented methionine excess diet were significant and ranged to the tune of 30 to 40% and 10 to 20%, respectively (Fig. 5).
Figure 4.

Growth and performance of L. rohita fish fed diets containing methionine-loaded in chitosan nanoparticles for 60 days. (A) Weight Gain (%); (B) SGR: Specific Growth Rate; (C) Protein Efficiency Ratio (PER); and (D) Food Conversion Ratio (FCR). Abbreviations: 0.8% NPs 0.8% chitosan nanoparticles; 1.2% M methionine in free form (M); 0.6% M-NPs or 1.2% M-NPs methionine loaded in chitosan nanoparticles (M-NPs). Values are presented as means ± standard error (control, n = 4; treatments, n = 3). a,b,cMeans bearing different superscript letters differ significantly (P < 0.05).
Figure 5.
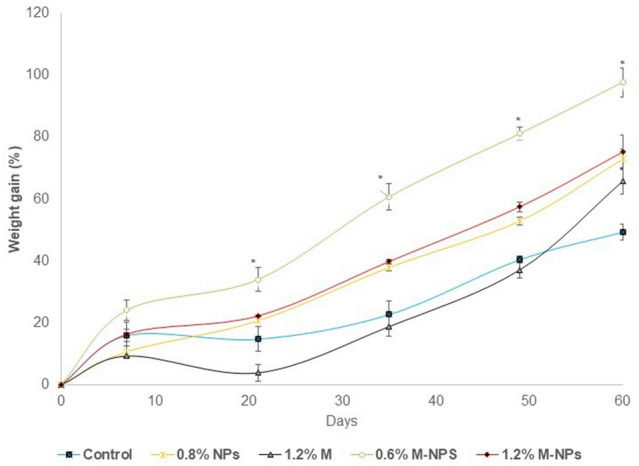
Time- interval growth pattern of L. rohita fish fed diets containing methionine loaded in chitosan nanoparticles for 60 days. Abbreviations: 0.8% NPs 0.8% chitosan nanoparticles; 1.2% M methionine in free form (M); 0.6% M-NPs or 1.2% M-NPs methionine loaded in chitosan nanoparticles(M-NPs). Values represented as means ± standard error (control, n = 4; treatments, n = 3). *Mean significantly (P < 0.05) different from the rest of means on the respective day.
Metabolic responses corresponded to growth and performance in fish
There were significant differences in hepatic AST (P = 0.000), ALT (P = 0.000), LDH (P = 0.003), and MDH (P = 0.015) in response to NPs. Liver ALT and AST activities of the L. rohita fingerlings were not influenced by plain chitosan NPs (P > 0.05). While activities of both transaminases in nano (0.6% M-NPs) supplemented-methionine-adequate diet were significantly higher than that of the free form 1.2% M diet, ALT activity in 1.2% methionine via either form (free or nano) was similar and AST in the 1.2% M-NPs group did not differ from that of the 0.6%M-NPs (Fig. 6A). Compared to the control, feeding 0.8% chitosan NPs and 1.2% methionine (1.2 M) decreased (P < 0.05) liver LDH. A comparison of liver dehydrogenases, LDH, and MDH activity, in both 0.6% and 1.2% M-NPs was found similar to the 1.2% M methionine-fed group (Fig. 6 B). The blood glucose level (P = 0.098) and muscle glycogen (P = 0.268) were similar among treatment groups (Table 3).
Figure 6.
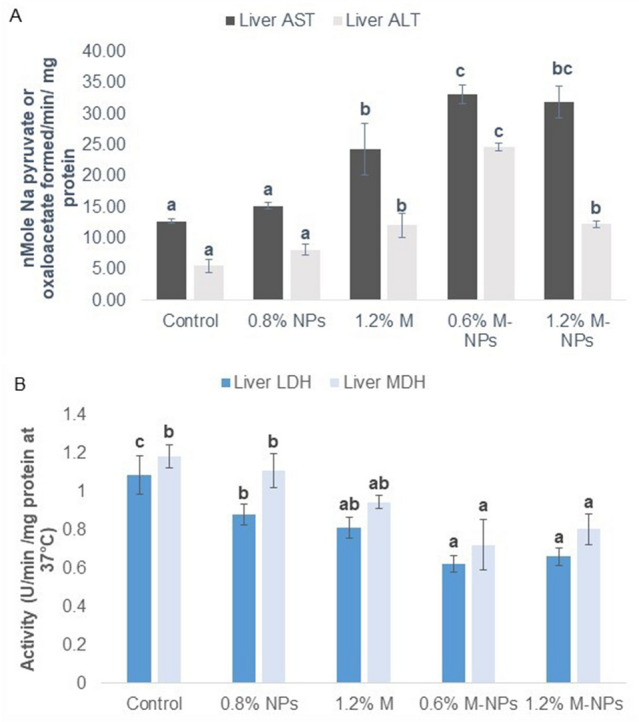
The activity of liver transaminases (A) and dehydrogenases (B) of L. rohita fish fed diets containing methionine loaded in chitosan nanoparticles for 60 days. Abbreviations: 0.8% NPs 0.8% chitosan nanoparticles; 1.2% M methionine in free form (M); 0.6% M-NPs or 1.2% M-NPs methionine loaded in chitosan nanoparticles (M-NPs). Values represented as means ± standard error (ALT and AST, n = 3 to 5; LDH and MDH, n = 3). a,b,cMeans bearing different superscript letters differ significantly (P < 0.05).
Table 3.
Blood glucose and muscle glycogen of L. rohita fish fed diets containing methionine loaded chitosan nanoparticles for 60 days.
| Treatments | Blood glucose (mg dl−1) | Muscle glycogen (mg/g) |
|---|---|---|
| Control | 91.89 ± 3.87 | 1.41 ± 0.06 |
| 0.8% NPs | 82.55 ± 4.84 | 1.55 ± 0.12 |
| 1.2% M | 88.21 ± 2.07 | 1.37 ± 0.09 |
| 0.6% M-NPs | 101.29 ± 9.93 | 1.54 ± 0.20 |
| 1.2% M-NPs | 92.07 ± 5.97 | 1.45 ± 0.04 |
0.8% NPs 0.8% chitosan nanoparticles; 1.2% M methionine in free form(M); 0.6% M-NPs or 1.2% M-NPs methionine loaded chitosan nanoparticles (M-NPs).
Values represented as means ± standard error (n = 3).
Improved sero-immunological profile in fish fed methionine-loaded chitosan nanoparticles
Dietary NPs had significant effects on fish’s serological attributes like total serum protein, albumin, and globulin (all P = 0.000) and immunological indicators such as albumin: globulin (A:G) ratio (P = 0.005), NBT score/respiratory burst (P = 0.001), myeloperoxidase (P = 0.018), serum complement, C3 (P = 0.018) and lysozyme (P = 0.002) (Fig. 7). Unaffected by chitosan NPs, serum total protein, albumin, and globulin were increased by feeding 1.2% M, but only total protein and globulin were further potentiated (P < 0.05) by feeding it in nanoform at 0.6% level (0.6% M-NPs), with 1.2% (1.2% M-NPs) retaining the effect on only globulin, while the form of methionine had no bearing on serum albumin of fish. As a result, the A:G ratio in fish fed on nanoform of methionine at both levels was improved (P < 0.05).
Figure 7.

Immunological parameters of L. rohita fish fed diets containing methionine loaded chitosan nanoparticles for 60 days. Abbreviations: 0.8% NPs 0.8% chitosan nanoparticles (NPs), 1.2% M methionine in free form (M); 0.6% M-NPs or 1.2% M-NPs methionine loaded chitosan nanoparticles (M-NPs). Values represented as means ± standard error (n = 6 for NBT and 3 for the rest). a,b,cMeans bearing different superscript letters differ significantly (P < 0.05).
Significantly highest (P < 0.05) respiratory burst activity and lysozyme activity was found on 0.6% M-NPs, a level that complemented the attainment of the required dietary concentration, as against 1.2% free (1.2% M) or nano form (1.2% M-NPs), which contributed 0.7% excess methionine and yielded lower scores in these tests. Compared to the basal diet subnormal for methionine, all five tests (respiratory burst, lysozyme, MPO activity, serum complement 3, and serum A: G) showed improved immune profile on nano-methionine-fed diets. Compared to the control, supplementation of 0.8% chitosan NPs increased (P < 0.05) respiratory burst activity in fish.
Discussion
Chitosan nanoparticle-based delivery system never tried before have been attempted in order to address various issues associated with methionine nutrition in aquatic species. Methionine and cysteine are sulphur containing amino acids, which are needed for the synthesis of sulphur-containing biochemicals. As the only variable in diets was methionine, cysteine (and other amino acids) being constant, only the former is the focus of discussion.
Chitosan NPs synthesised by ionic gelation method have similar size distribution31. Size of the NPs was slightly increased after loading methionine, which agrees with earlier reports52–54. The hydrated form of NPs before (173 nm) and after (218 nm) loading was 1.28 to 2.47 times larger than that of dehydrated nanoparticles (70–150 nm) (TEM images). The larger size of hydrated NPs is due to the extended and large size conformation of chitosan in aqueous solution55, and swelling or aggregation of single particles dispersed in water53,56. TEM images showed the spherical and smooth surfaces of chitosan NPs. Similarly, chitosan NPs57 without or with vitamin C28 or bovine serum albumin (BSA)-loading55 were found to exhibit a smooth spherical shape.
Chitosan has mucoadhesive characteristics due to the positive charge of protonated amine groups on it at pH less than 6.226,28. The NPs before and after loading of L-methionine had a surface charge density (zeta potential) around + 30 mV, which is the minimum for physically stable NPs in an aqueous solution58,59. The positive surface charge helps to exclude NPs from lysosomal reduction60 hence they could be targeted to the cytoplasm61.
Good encapsulation of methionine in chitosan NPs needs defined conditions. When chitosan and tripolyphosphate (TPP) are mixed, it forms a compact nano-complex by cross-linking the positively charged amide group on chitosan and the negatively charged phosphate group on TPP59. Loading of L-methionine on chitosan NPs at pH 6.0 (the isoelectric pH of L-methionine, pI is 5.7 and pI of chitosan is 6.2) was found to be most suitable. BSA- loaded chitosan NPs were in a range of 250–350 nm with 61–78%62 encapsulation efficiency. Relatively, high encapsulation in the present study is attributed to the hydrophobic interactions formed between chitosan and hydrophobic amino acids63.
Fourier transforms infrared (FT-IR) spectroscopy further ensured the encapsulation of L-methionine in chitosan NPs. A broad peak at 3500–3300 cm−1 is attributed to overlapped stretches of carboxyl O–H, N–H and aliphatic C-H bonds. A weak band between 3300 and 2950 cm−1 is attributed to C–H stretching vibrations in L-methionine. The peak shifting from 3454 cm−1 in chitosan NPs to 3427 cm−1 in L-methionine-loaded chitosan NPs is attributed to N–H and O–H stretching vibrations64. Similarly, the peak at 3150 cm-1 corresponding to N–H stretching vibrations of a primary amine in L-methionine65 disappeared after the formation of M-NPs. The peak at 2914 cm−1 in L-methionine and 2968 cm−1 in chitosan were attributed to CH2 stretching vibrations57, which shifted to 2926 cm−1 in M-NPs. The band from 2600 to 2550 cm−1 attributed to weak S–H stretching of the thiol group was found only in L-methionine, and L-methionine-loaded NPs spectra. Wave number assigned to asymmetric stretching of CH3-S of L-methionine at 746 cm−1 disappeared in L-methionine-loaded NPs, while highly variable CH2-S stretches of L-methionine at 644, 660 and 764 cm−1 were visible in L-methionine-loaded NPs as well66. All these findings further ensured that chitosan NPs could be used to encapsulate L-methionine.
In vitro test results of 40% methionine release in 18 h, or 86.5% release at the end of 42 h shows a slow, delayed, and sustained release of methionine from chitosan NPs. In one study39, 51.29% S-adenosyl-L-methionine (SAMe)-loaded onto nano-chitosan was released at the end of the first hour, while 70% of pure SAMe was dissolved. Using the same vehicle, chitosan NPs, 96% SAMe release in about 14 h67, and only 12% ciprofloxacin release in 96 h68 have also been reported. The release of methionine might occur by diffusion, which involves penetration of medium into the system, causing swelling and then release33,52,69. A drag observed in the earlier hours of the in vitro release test may indicate the time taken up for swelling of chitosan NPs in the medium. Such slow release of drug from NPs is also attributed to the decomposition of the carrier69. There is a caveat to bear in mind while extending results on the release of methionine at pH 7.4 at 37 °C during in vitro tests to the actual gut of animals. In a more realistic release kinetics study70, the chitosan nanoparticles system ensures a burst delivery efficiency of 50% in 6 h (and 70% in 12 h) of rifampicin at a simulated intestinal pH in comparison to the simulated gastric environment of less than 10%.
Chitosan NPs are well-proven active carriers of drugs used for effective oral delivery71, and hence supplementation of methionine by loading in chitosan nanoparticles enhanced the growth of fish in the current investigation. The positive charge of chitosan NPs interacts with the negatively charged epithelial membrane and induces structural reorganisation to enhance transport through the paracellular pathway72,73. It is also reported that the bi-adhesion nature of chitosan NPs helps in controlled release26,28 and its mucoadhesive property can extend the residence time of NPs in the gastrointestinal tract73. In the present study, the base level of 0.8% chitosan was ensured while preparing M-NPs to negate their effect on the parameters studied. By doing so, the significantly highest growth rate was found in the 0.6% M-NPs supplemented diet, where M-NPs contributed to gross methionine adequacy of diet through methionine’s slow release from chitosan NPs and its efficient absorption in the fish lumen.
Although reports on amino acids/M-NPs are not available, a few on coated forms of methionine or lysine showed encouraging results. When 0.3% acrylic resin coated methionine supplemented diet containing 46% CP was fed, 23.64% higher weight gain and 7.99% higher specific growth rate of the juvenile cobia, Rachycentron canadum (Linnaeus) was found22. Successful replacement of 21.2% fish meal by a mixture of protein with 0.5% lysine and (hydrogenated palm oil) coated 0.34% methionine in the diet of black sea bream, Acanthopagrus schlegelii has been attempted74. Compared to an optimal methionine diet containing 0.6% M-NPs, supplementation of 1.2% methionine in either form (nano or free) to the basal diet caused 0.7% methionine excess leading to decreased % weight gain, specific growth rate and protein efficiency ratio. A significant decrease in these parameters was also observed in another Indian major carp, Cirrhinus mrigala43 and in channel catfish, Ictalurus punctatus75 when fed 0.5 to 0.75% more methionine than requirement.
Increased fish growth on controlled release form 0.6 M-NPs can be attributed to the availability of methionine in plasma for extended hours. Studies in rats on a single oral dose of free vs chitosan-encapsulated chlorogenic acid (CGA), a phenolic antioxidant, clearly indicated that the level for free CGA peaks at 1.5 h and then reaches baseline at the end of 2 h, while for encapsulated CGA, plasma concentration reaches the peak at 4 h and then comes to baseline at 6 h. Such lower and sustained release over a longer duration (P < 0.01) of nano-CGA has more bioavailability upon oral administration76 which is also likely for methionine. Rolland et al.9 reported that supplementation with crystalline amino acids in a plant protein-based diet resulted in the rapid appearance of amino acids in the plasma, especially a greater and faster rise of methionine, followed by sharp decreases compared to fish meal. Such highest plasma availability of amino acids results in a temporal mismatch at the site of protein synthesis in rainbow trout (Oncorhynchus mykiss W.), leading to a decrease in growth and an increase in ammonia excretion77, which can be avoided with the usage of slow releasing nano-methionine, M-NPs, at an appropriate level, as used in this study. Studies78 in grass carp, Ctenopharyngodon idella, suggest significantly (P < 0.05) increased growth coupled with high activity of intestinal enzymes such as trypsin, lipase and amylase, Na+ /K+ -ATPase, alkaline phosphatase (AKP), γ-glutamyl transpeptidase (γ-GT) and creatine kinase (CK) (in one of the segments) in fish given methionine close to their dietary requirement, while decreased (P < 0.05) growth and compromised activities were noticed when dietary methionine level was 1.5 to 2 times higher than the requirement. This explains the good growth patterns on 0.6% M-NPs but not 1.2% in M or 1.2%M-NPs. Although, chitosan at 2 to 5% is known to display growth-enhancing effect in a variety of fish species79–81, this effect on 0.8% level of chitosan nanoparticles in this study suggests more effectiveness of chitosan NPs over plain chitosan.
Since amino acids are primary sources of energy in fish82, enzymes involved in their metabolism were studied. High energy demand for protein synthesis in fish is achieved by increasing the gluconeogeic pathway. Substrate for this pathway is provided by transaminases, especially AST and ALT82. Activities of these enzymes in the liver corresponded with energy-demanding protein synthesis processes such as growth and immuno-synthesis, which were highest in 0.6% M-NPs than 1.2% methionine (in free or nano form) indicating increased utilisation of dietary protein towards growth. Direct correlation between growth and transaminases activity has been shown in L. rohita36 and in Rachycentron canadum83. Our results agrees with Wu, et al.78 who reported decreased growth in grass carp (Ctenopharyngodon idella) along with decreased hepatopancreatic AST, but not ALT activity on either side (0.5 × less or 2 times more) of the required dietary level. Similarly, reduced lactate and malate dehydrogenase activity corresponds to increased growth in fish, which is attributed to reduced glycolytic breakdown during growth49,84. In this study, both LDH and MDH were comparatively lower in M-NPs fed groups than in control, but not in 1.2% free methionine-fed fish. Further, blood glucose and muscle glycogen levels showed no significant difference between treatment groups, which indicates the effective utilisation of glucose produced via gluconeogenesis36.
Immuno-serological tests employed in the study for fish fed with M-NPs are robust and well-validated tests. Macrophage exhibits respiratory burst activity for destroying invading pathogenic microbes, which can be determined by nitroblue-tetrazolium (NBT) reduction assay85. Myeloperoxidase enzyme uses H2O2 to produce HCl during the respiratory burst. Lysozymes acts as an opsonin which activates phagocytes and the complement system80. The activated complement system destructs foreign organisms by opsonizing phagocytes86. Further higher serum protein content, globulin content and low A:G ratio indicate the immune boosting activity of an immunostimulant87,88.
Methyl donors are well known immunomodulatory agents and our studies on betaine and choline, compounds involved in one-carbon metabolism (methionine cycle), have convincingly demonstrated these properties in fish45,47. Significantly improved scores of three (respiratory burst activity of neutrophils, lysozyme activity and serum albumin: globulin ratio) of five (myeloperoxidase and serum complement 3) immune profile parameters were found in the diet in which 0.6% nano-methionine (0.6% M-NPs) contributed to methionine adequacy of the basal diet. On the other hand, higher than required methionine through supplementation of 1.2% in either form (free or nano) to basal diet with 0.85% methionine in intact protein was of no use as fish were not challenged, a situation which may increase its requirement. In yellow catfish, Pelteobagrus fulvidraco89, dietary methionine close to requirement (0.97% and 1.17%) showed the highest respiratory burst activity, lysozyme and total globulin than fed with subnormal (lower) or higher levels up to 1.64%. Increased complement (C3) activity and lysozyme activity were found at 1–1.2% of free L-methionine supplied groups in juvenile Jian carp44,86 and in European seabass90. Similarly, Wu et al.78 convincingly demonstrated the importance of a normal level of methionine in the diet of sub-adult grass carp, Ctenopharyngodon idella, for the maintenance of levels of numerous antioxidant indicators such as malondialdehyde, protein carbonyl content, catalase, glutathione-S-transferase, anti-superoxide anion, anti-hydroxyl radical, superoxide dismutase, glutathione peroxidase, glutathione reductase and glutathione contents in hepatopancreas and intestine. In fish fed on either side of the requirement level (~ 0.62%) (0.5 × times less or 1.5 to 2 times higher) of this amino acid, the first four attributes were significantly increased, and the rest were decreased (P < 0.1) or remained unaffected and unchanged. The present study harnessed immune boosting79,91,92 and metabolism modulating- properties of chitosan79,81 to develop immune response in fish93. Chitosan NPs synergistically and effectively acted with L-methionine towards improving immune profile in addition to growth and performance when given as potent M-NPs at a level not to exceed the total (intact and free) dietary methionine level.
Conclusion
This is the pioneering report on the synthesis of L-methionine-loaded chitosan nanoparticles (M-NPs). These M-NPs during in vitro testing exhibited sustained and slow release of methionine over a prolonged period. Complementing diets limiting in methionine with M-NPs (nano methionine) to meet its dietary requirement was found to confer intended performance and health benefits in fish. Specific amino acid-loaded chitosan nanoparticles such as M-NPs in this study can find dietary applications to achieve desired goals in animal production and health practice, where a slow and controlled release of this amino acid is desired.
Supplementary Information
Acknowledgements
The authors are grateful to the Director/Vice Chancellor, ICAR-Central Institute of Fisheries Education for providing the necessary facilities for conducting this experiment. Research Grant from Consortium Research Platform on Nanotechnology, Indian Council of Agricultural Research is sincerely acknowledged. The authors are also thankful to the Head, SAIF, IIT, Mumbai, India for providing the facilities for nanoparticle characterization.
Author contributions
S.G. and S.B.J. conceptualised the project idea. SG acquired the funding, designed the study, and supervised the project. P.M.N. and S.G. worked in the lab and acquired the data, while S.G. provided technical and administrative support. P.M.N. and S.G. carried out animal experiments. P.M.N., S.G., and S.G. sketched the first draft. S.G. and S.B.J. did critical and extensive interpretation of the data and did major re-writing. P.M.N., S.G., S.G. and S.B.J. approved the manuscript.
Data availability
All data generated or analysed during this study are included in this published article.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Pallath Muhammed Nuzaiba and Subodh Gupta.
Contributor Information
Subodh Gupta, Email: sgupta@cife.edu.in.
Sanjay Balkrishna Jadhao, Email: sbjadhao@hotmail.com.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-023-34448-6.
References
- 1.Hardy RW. Utilization of plant proteins in fish diets: effects of global demand and supplies of fishmeal. Aquac. Res. 2010;41:770–776. doi: 10.1111/j.1365-2109.2009.02349.x. [DOI] [Google Scholar]
- 2.Li X, Zheng S, Cheng K, Ma X, Wu G. Use of alternative protein sources for fishmeal replacement in the diet of largemouth bass (Micropterus salmoides). Part II: Effects of supplementation with methionine or taurine on growth, feed utilization, and health. Amino Acids. 2021;53:49–62. doi: 10.1007/s00726-020-02922-4. [DOI] [PubMed] [Google Scholar]
- 3.Sanchez-Muros M, et al. Effect of feeding method and protein source on Sparus aurata feeding patterns. Aquaculture. 2003;224:89–103. doi: 10.1016/S0044-8486(03)00211-4. [DOI] [Google Scholar]
- 4.Niculescu MD, Zeisel SH. Diet, methyl donors and DNA methylation: interactions between dietary folate, methionine and choline. J. Nutr. 2002;132:2333S–2335S. doi: 10.1093/jn/132.8.2333S. [DOI] [PubMed] [Google Scholar]
- 5.Giroux I, Kurowska EM, Carroll KK. Role of dietary lysine, methionine, and arginine in the regulation of hypercholesterolemia in rabbits. J. Nutr. Biochem. 1999;10:166–171. doi: 10.1016/S0955-2863(98)00091-6. [DOI] [PubMed] [Google Scholar]
- 6.Froese DS, Fowler B, Baumgartner MR. Vitamin B12, folate, and the methionine remethylation cycle—Biochemistry, pathways, and regulation. J. Inherit. Metab. Dis. 2019;42:673–685. doi: 10.1002/jimd.12009. [DOI] [PubMed] [Google Scholar]
- 7.Cappelaere L, Le Cour Grandmaison J, Martin N, Lambert W. Amino acid supplementation to reduce environmental impacts of broiler and pig production: A review. Front. Vet. Sci. 2021;8:689259. doi: 10.3389/fvets.2021.689259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Furuya WM, Cruz TPD, Gatlin DM., III Amino acid requirements for nile tilapia: An update. Animals. 2023;13:900. doi: 10.3390/ani13050900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rolland M, Larsen BK, Holm J, Dalsgaard J, Skov PV. Effect of plant proteins and crystalline amino acid supplementation on postprandial plasma amino acid profiles and metabolic response in rainbow trout (Oncorhynchus mykiss) Aquac. Int. 2015;23:1071–1087. doi: 10.1007/s10499-014-9865-4. [DOI] [Google Scholar]
- 10.Lovell RT. Nutrition of aquaculture species. J. Anim. Sci. 1991;69:4193–4200. doi: 10.2527/1991.69104193x. [DOI] [PubMed] [Google Scholar]
- 11.Nguyen L, Salem SM, Davis DA. Indispensable and dispensable amino acid supplementation in diets offered to Nile tilapia, Oreochromis niloticus. Anim. Feed Sci. Technol. 2022;290:115361. doi: 10.1016/j.anifeedsci.2022.115361. [DOI] [Google Scholar]
- 12.Kim S-Y, Kim H, Min H. Effects of excessive dietary methionine on oxidative stress and dyslipidemia in chronic ethanol-treated rats. Nutr. Res. Pract. 2015;9:144–149. doi: 10.4162/nrp.2015.9.2.144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Plauth M, et al. Long-term treatment of latent portosystemic encephalopathy with branched-chain amino acids: A double-blind placebo-controlled crossover study. J. Hepatol. 1993;17:308–314. doi: 10.1016/S0168-8278(05)80210-7. [DOI] [PubMed] [Google Scholar]
- 14.Guo J, Zhou W, Liu S, Zhang W, Mai K. Efficacy of crystalline methionine and microencapsulation methionine in diets for Pacific white shrimp Litopenaeus vannamei. Aquac. Res. 2020;51:4206–4214. doi: 10.1111/are.14762. [DOI] [Google Scholar]
- 15.Yúfera M, Kolkovski S, Fernández-Dıaz C, Dabrowski K. Free amino acid leaching from a protein-walled microencapsulated diet for fish larvae. Aquaculture. 2002;214:273–287. doi: 10.1016/S0044-8486(01)00902-4. [DOI] [Google Scholar]
- 16.Bottiglieri T. S-Adenosyl-L-methionine (SAMe): From the bench to the bedside—Molecular basis of a pleiotrophic molecule. Am. J. Clin. Nutr. 2002;76:1151S–1157S. doi: 10.1093/ajcn/76.5.1151S. [DOI] [PubMed] [Google Scholar]
- 17.Lieber CS, Packer L. S-Adenosylmethionine: Molecular, biological, and clinical aspects—An introduction. Am. J. Clin. Nutr. 2002;76:1148S–1150S. doi: 10.1093/ajcn/76.5.1148S. [DOI] [PubMed] [Google Scholar]
- 18.Navik U, Sheth VG, Kabeer SW, Tikoo K. Dietary supplementation of methyl donor L-methionine alters epigenetic modification in type 2 diabetes. Mol. Nutr. Food Res. 2019;63:1801401. doi: 10.1002/mnfr.201801401. [DOI] [PubMed] [Google Scholar]
- 19.Aliu, E., Kanungo, S. & Arnold, G. L. Amino acid disorders. Ann. Transl. Med.6 (2018). [DOI] [PMC free article] [PubMed]
- 20.MacDonald A, Singh RH, Rocha JC, van Spronsen FJ. Optimising amino acid absorption: Essential to improve nitrogen balance and metabolic control in phenylketonuria. Nutr. Res. Rev. 2019;32:70–78. doi: 10.1017/S0954422418000173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Scala I, et al. Beneficial effects of slow-release large neutral amino acids after a phenylalanine oral load in patients with phenylketonuria. Nutrients. 2021;13:4012. doi: 10.3390/nu13114012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chi S, Tan B, Dong X, Yang Q, Liu H. Effects of supplemental coated or crystalline methionine in low-fishmeal diet on the growth performance and body composition of juvenile cobia Rachycentron canadum (Linnaeus) Chin. J. Oceanol. Limnol. 2014;32:1297–1306. doi: 10.1007/s00343-014-3276-2. [DOI] [Google Scholar]
- 23.Pridgen EM, Langer R, Farokhzad OC. Biodegradable, polymeric nanoparticle delivery systems for cancer therapy. Nanomedicine. 2007;2:669–680. doi: 10.2217/17435889.2.5.669. [DOI] [PubMed] [Google Scholar]
- 24.Prajapati SK, Jain A, Jain A, Jain S. Biodegradable polymers and constructs: A novel approach in drug delivery. Eur. Polym. J. 2019;120:109191. doi: 10.1016/j.eurpolymj.2019.08.018. [DOI] [Google Scholar]
- 25.Fàbregas A, et al. Impact of physical parameters on particle size and reaction yield when using the ionic gelation method to obtain cationic polymeric chitosan–tripolyphosphate nanoparticles. Int. J. Pharm. 2013;446:199–204. doi: 10.1016/j.ijpharm.2013.02.015. [DOI] [PubMed] [Google Scholar]
- 26.Dorkoosh F, Verhoef J, Rafiee-Tehrani M, Borchard G, Junginger H. Peroral drug delivery systems for peptides and proteins. STP Pharma Sci. 2002;12:213–221. [Google Scholar]
- 27.Muzzarelli R. Chitins and chitosans as immunoadjuvants and non-allergenic drug carriers. Mar. Drugs. 2010;8:292–312. doi: 10.3390/md8020292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Alishahi A, et al. Chitosan nanoparticle to carry vitamin C through the gastrointestinal tract and induce the non-specific immunity system of rainbow trout (Oncorhynchus mykiss) Carbohydr. Polym. 2011;86:142–146. doi: 10.1016/j.carbpol.2011.04.028. [DOI] [Google Scholar]
- 29.Liu M, et al. Efficient mucus permeation and tight junction opening by dissociable “mucus-inert” agent coated trimethyl chitosan nanoparticles for oral insulin delivery. J. Control. Release. 2016;222:67–77. doi: 10.1016/j.jconrel.2015.12.008. [DOI] [PubMed] [Google Scholar]
- 30.Aranaz I, et al. Functional characterization of chitin and chitosan. Curr. Chem. Biol. 2009;3:203–230. doi: 10.2174/187231309788166415. [DOI] [Google Scholar]
- 31.Calvo P, Remuñán-López C, Vila-Jato JL, Alonso M. Novel hydrophilic chitosan-polyethylene oxide nanoparticles as protein carriers. J. Appl. Polym. Sci. 1997;63:125–132. doi: 10.1002/(SICI)1097-4628(19970103)63:1<125::AID-APP13>3.0.CO;2-4. [DOI] [Google Scholar]
- 32.Sabbagh, F., Muhamad, I. I., Niazmand, R., Dikshit, P. K. & Kim, B. S. Recent progress in polymeric non-invasive insulin delivery. Int. J. Biol. Macromol. (2022). [DOI] [PubMed]
- 33.Agnihotri SA, Mallikarjuna NN, Aminabhavi TM. Recent advances on chitosan-based micro-and nanoparticles in drug delivery. J. Control. Release. 2004;100:5–28. doi: 10.1016/j.jconrel.2004.08.010. [DOI] [PubMed] [Google Scholar]
- 34.Huang M, Khor E, Lim L-Y. Uptake and cytotoxicity of chitosan molecules and nanoparticles: effects of molecular weight and degree of deacetylation. Pharm. Res. 2004;21:344–353. doi: 10.1023/B:PHAM.0000016249.52831.a5. [DOI] [PubMed] [Google Scholar]
- 35.He C, Hu Y, Yin L, Tang C, Yin C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials. 2010;31:3657–3666. doi: 10.1016/j.biomaterials.2010.01.065. [DOI] [PubMed] [Google Scholar]
- 36.Kumari R, et al. Chitosan nanoencapsulated exogenous trypsin biomimics zymogen-like enzyme in fish gastrointestinal tract. PLoS ONE. 2013;8:e74743. doi: 10.1371/journal.pone.0074743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ferosekhan S, et al. RNA-loaded chitosan nanoparticles for enhanced growth, immunostimulation and disease resistance in fish. Curr. Nanosci. 2014;10:453–464. doi: 10.2174/1573413710666140115220300. [DOI] [Google Scholar]
- 38.Rather MA, et al. Chitosan-nanoconjugated hormone nanoparticles for sustained surge of gonadotropins and enhanced reproductive output in female fish. PLoS ONE. 2013;8:e57094. doi: 10.1371/journal.pone.0057094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ergin, A. D., Bayindir, Z. S. & Yuksel, N. Characterization and optimization of colon targeted S-adenosyl-L-methionine loaded chitosan nanoparticles. Marmara Pharm. J.23 (2019).
- 40.McCarthy TE, Sullivan M. A new and highly specific colorimetric test for methionine. J. Biol. Chem. 1941;141:871–876. doi: 10.1016/S0021-9258(18)72759-2. [DOI] [Google Scholar]
- 41.Kader MA, Koshio S, Ishikawa M, Yokoyama S, Bulbul M. Supplemental effects of some crude ingredients in improving nutritive values of low fishmeal diets for red sea bream, Pagrus major. Aquaculture. 2010;308:136–144. doi: 10.1016/j.aquaculture.2010.07.037. [DOI] [Google Scholar]
- 42.Murthy H, Varghese T. Total sulphur amino acid requirement of the Indian major carp, Labeo rohita (Hamilton) Aquac. Nutr. 1998;4:61–66. doi: 10.1046/j.1365-2095.1998.00045.x. [DOI] [Google Scholar]
- 43.Ahmed I, Khan MA, Jafri A. Dietary methionine requirement of fingerling Indian major carp, Cirrhinus mrigala (Hamilton) Aquac. Int. 2003;11:449–462. doi: 10.1023/B:AQUI.0000004181.89420.a2. [DOI] [Google Scholar]
- 44.Tang L, et al. Effect of methionine on intestinal enzymes activities, microflora and humoral immune of juvenile Jian carp (Cyprinus carpio var. Jian) Aquac. Nutr. 2009;15:477–483. doi: 10.1111/j.1365-2095.2008.00613.x. [DOI] [Google Scholar]
- 45.Muthappa N, et al. Lipotropes promote immunobiochemical plasticity and protect fish against low-dose pesticide-induced oxidative stress. Cell Stress Chaperones. 2014;19:61–81. doi: 10.1007/s12192-013-0434-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kumar N, et al. Anti-oxidative and immuno-hematological status of Tilapia (Oreochromis mossambicus) during acute toxicity test of endosulfan. Pestic. Biochem. Physiol. 2011;99:45–52. doi: 10.1016/j.pestbp.2010.10.003. [DOI] [Google Scholar]
- 47.Kumar N, et al. Lipotropes protect against pathogen-aggravated stress and mortality in low dose pesticide-exposed fish. PLoS ONE. 2014;9:e93499. doi: 10.1371/journal.pone.0093499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sahoo P, Kumari J, Mishra B. Non-specific immune responses in juveniles of Indian major carps. J. Appl. Ichthyol. 2005;21:151–155. doi: 10.1111/j.1439-0426.2004.00606.x. [DOI] [Google Scholar]
- 49.Akhtar MS, et al. Stress mitigating and immunomodulatory effect of dietary pyridoxine in Labeo rohita (Hamilton) fingerlings. Aquac. Res. 2010;41:991–1002. doi: 10.1111/j.1365-2109.2009.02383.x. [DOI] [Google Scholar]
- 50.Seifter S, Dayton S, Novic B, Muntwyler E. The estimation of glycogen with the anthrone reagent. Arch. Biochem. 1950;25:191–200. [PubMed] [Google Scholar]
- 51.Carroll NV, Longley RW, Roe JH. The determination of glycogen in liver and muscle by use of anthrone reagent. J. Biol. Chem. 1956;220:583–593. doi: 10.1016/S0021-9258(18)65284-6. [DOI] [PubMed] [Google Scholar]
- 52.Keawchaoon L, Yoksan R. Preparation, characterization and in vitro release study of carvacrol-loaded chitosan nanoparticles. Colloids Surf. B. 2011;84:163–171. doi: 10.1016/j.colsurfb.2010.12.031. [DOI] [PubMed] [Google Scholar]
- 53.Hosseini SF, Zandi M, Rezaei M, Farahmandghavi F. Two-step method for encapsulation of oregano essential oil in chitosan nanoparticles: Preparation, characterization and in vitro release study. Carbohydr. Polym. 2013;95:50–56. doi: 10.1016/j.carbpol.2013.02.031. [DOI] [PubMed] [Google Scholar]
- 54.Sotelo-Boyás M, Correa-Pacheco Z, Bautista-Baños S, Corona-Rangel M. Physicochemical characterization of chitosan nanoparticles and nanocapsules incorporated with lime essential oil and their antibacterial activity against food-borne pathogens. LWT Food Sci. Technol. 2017;77:15–20. doi: 10.1016/j.lwt.2016.11.022. [DOI] [Google Scholar]
- 55.Gan Q, Wang T, Cochrane C, McCarron P. Modulation of surface charge, particle size and morphological properties of chitosan–TPP nanoparticles intended for gene delivery. Colloids Surf. B. 2005;44:65–73. doi: 10.1016/j.colsurfb.2005.06.001. [DOI] [PubMed] [Google Scholar]
- 56.Yoksan R, Jirawutthiwongchai J, Arpo K. Encapsulation of ascorbyl palmitate in chitosan nanoparticles by oil-in-water emulsion and ionic gelation processes. Colloids Surf. B. 2010;76:292–297. doi: 10.1016/j.colsurfb.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 57.Xu Y, Du Y. Effect of molecular structure of chitosan on protein delivery properties of chitosan nanoparticles. Int. J. Pharm. 2003;250:215–226. doi: 10.1016/S0378-5173(02)00548-3. [DOI] [PubMed] [Google Scholar]
- 58.Müller R, Jacobs C, Kayser O. Nanosuspensions as particulate drug formulations in therapy: Rationale for development and what we can expect for the future. Adv. Drug Deliv. Rev. 2001;47:3–19. doi: 10.1016/S0169-409X(00)00118-6. [DOI] [PubMed] [Google Scholar]
- 59.Du W-L, Niu S-S, Xu Y-L, Xu Z-R, Fan C-L. Antibacterial activity of chitosan tripolyphosphate nanoparticles loaded with various metal ions. Carbohydr. Polym. 2009;75:385–389. doi: 10.1016/j.carbpol.2008.07.039. [DOI] [Google Scholar]
- 60.Dodane V, Vilivalam VD. Pharmaceutical applications of chitosan. Pharm. Sci. Technol. Today. 1998;1:246–253. doi: 10.1016/S1461-5347(98)00059-5. [DOI] [Google Scholar]
- 61.Panyam J, Labhasetwar V. Biodegradable nanoparticles for drug and gene delivery to cells and tissue. Adv. Drug Deliv. Rev. 2003;55:329–347. doi: 10.1016/S0169-409X(02)00228-4. [DOI] [PubMed] [Google Scholar]
- 62.Gan Q, Wang T. Chitosan nanoparticle as protein delivery carrier—Systematic examination of fabrication conditions for efficient loading and release. Colloids Surf. B. 2007;59:24–34. doi: 10.1016/j.colsurfb.2007.04.009. [DOI] [PubMed] [Google Scholar]
- 63.Ma Z, Yeoh HH, Lim LY. Formulation pH modulates the interaction of insulin with chitosan nanoparticles. J. Pharm. Sci. 2002;91:1396–1404. doi: 10.1002/jps.10149. [DOI] [PubMed] [Google Scholar]
- 64.Rumengan I, et al. Structural characteristics of chitin and chitosan isolated from the biomass of cultivated rotifer, Brachionus rotundiformis. Int. J. Fish. Aquat. Sci. 2014;3:12–18. [Google Scholar]
- 65.Dhakshinamoorthy A, Sharmila A, Pitchumani K. Layered double hydroxide-supported L-methionine-catalyzed chemoselective O-methylation of phenols and esterification of carboxylic acids with dimethyl carbonate: A “green” protocol. Chem. Eur. J. 2010;16:1128–1132. doi: 10.1002/chem.200902626. [DOI] [PubMed] [Google Scholar]
- 66.Smith BC. Fundamentals of Fourier Transform Infrared Spectroscopy. 2. CRC Press; 2011. [Google Scholar]
- 67.Magesh B, Naidu P, Rajarajeswari G. S-adenosyl-l-methionine (SAMe)-loaded nanochitosan particles: Synthesis, characterisation and in vitro drug release studies. J. Exp. Nanosci. 2015;10:828–843. doi: 10.1080/17458080.2014.910616. [DOI] [Google Scholar]
- 68.Sobhani Z, Samani SM, Montaseri H, Khezri E. Nanoparticles of chitosan loaded ciprofloxacin: Fabrication and antimicrobial activity. Adv. Pharm. Bull. 2017;7:427–432. doi: 10.15171/apb.2017.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Yuan Q, Shah J, Hein S, Misra R. Controlled and extended drug release behavior of chitosan-based nanoparticle carrier. Acta Biomater. 2010;6:1140–1148. doi: 10.1016/j.actbio.2009.08.027. [DOI] [PubMed] [Google Scholar]
- 70.Ghosh R, et al. Oral drug delivery using a polymeric nanocarrier: Chitosan nanoparticles in the delivery of rifampicin. Mater. Adv. 2022;3:4622–4628. doi: 10.1039/D2MA00295G. [DOI] [Google Scholar]
- 71.Pan Y, et al. Bioadhesive polysaccharide in protein delivery system: Chitosan nanoparticles improve the intestinal absorption of insulin in vivo. Int. J. Pharm. 2002;249:139–147. doi: 10.1016/S0378-5173(02)00486-6. [DOI] [PubMed] [Google Scholar]
- 72.Borchard G, et al. The potential of mucoadhesive polymers in enhancing intestinal peptide drug absorption. III: Effects of chitosan-glutamate and carbomer on epithelial tight junctions in vitro. J. Control. Release. 1996;39:131–138. doi: 10.1016/0168-3659(95)00146-8. [DOI] [Google Scholar]
- 73.Bowman K, Leong KW. Chitosan nanoparticles for oral drug and gene delivery. Int. J. Nanomed. 2006;1:117–128. doi: 10.2147/nano.2006.1.2.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Lu J, et al. Effects of supplementation coated lysine and methionine in mixture protein diets on growth performance, digestibility and serum biochemical indices of juvenile Black Sea bream, Acanthopagrus schlegelii. Turk. J. Fish. Aquat. Sci. 2014;14:633–642. doi: 10.4194/1303-2712-v14_3_05. [DOI] [Google Scholar]
- 75.Cai Y, Burtle GJ. Methionine requirement of channel catfish fed soybean meal-corn-based diets. J. Anim. Sci. 1996;74:514–521. doi: 10.2527/1996.743514x. [DOI] [PubMed] [Google Scholar]
- 76.Nallamuthu I, Devi A, Khanum F. Chlorogenic acid loaded chitosan nanoparticles with sustained release property, retained antioxidant activity and enhanced bioavailability. Asian J. Pharm. Sci. 2015;10:203–211. doi: 10.1016/j.ajps.2014.09.005. [DOI] [Google Scholar]
- 77.Bodin N, et al. Effects of fish size and diet adaptation on growth performances and nitrogen utilization of rainbow trout (Oncorhynchus mykiss W.) juveniles given diets based on free and/or protein-bound amino acids. Aquaculture. 2012;356:105–115. doi: 10.1016/j.aquaculture.2012.05.030. [DOI] [Google Scholar]
- 78.Wu P, et al. The relationship between dietary methionine and growth, digestion, absorption, and antioxidant status in intestinal and hepatopancreatic tissues of sub-adult grass carp (Ctenopharyngodon idella) J. Anim. Sci. Biotechnol. 2017;8:63. doi: 10.1186/s40104-017-0194-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Cha S-H, Lee J-S, Song C-B, Lee K-J, Jeon Y-J. Effects of chitosan-coated diet on improving water quality and innate immunity in the olive flounder, Paralichthys olivaceus. Aquaculture. 2008;278:110–118. doi: 10.1016/j.aquaculture.2008.01.025. [DOI] [Google Scholar]
- 80.Geng X, et al. Effects of dietary chitosan and Bacillus subtilis on the growth performance, non-specific immunity and disease resistance of cobia, Rachycentron canadum. Fish Shellfish Immunol. 2011;31:400–406. doi: 10.1016/j.fsi.2011.06.006. [DOI] [PubMed] [Google Scholar]
- 81.Sajid M, Prabjeet S, Samoon MH, Balange AK. Effect of dietary chitosan on non-specific immune response and growth of Cyprinus carpio challenged with Aeromonas hydrophila. Int. Aquat. Res. 2010;2:77–85. [Google Scholar]
- 82.Knox WE, Greengard O. The regulation of some enzymes of nitrogen metabolism—An introduction to enzyme physiology. Adv. Enzyme. Regul. 1965;3:247–313. doi: 10.1016/0065-2571(65)90059-2. [DOI] [Google Scholar]
- 83.Luo Y, et al. Effects of dietary rapeseed meal on growth performance, digestion and protein metabolism in relation to gene expression of juvenile cobia (Rachycentron canadum) Aquaculture. 2012;368:109–116. doi: 10.1016/j.aquaculture.2012.09.013. [DOI] [Google Scholar]
- 84.Koedijk RM, et al. Ontogenetic effects of diet during early development on growth performance, myosin mRNA expression and metabolic enzyme activity in Atlantic cod juveniles reared at different salinities. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010;156:102–109. doi: 10.1016/j.cbpa.2010.01.007. [DOI] [PubMed] [Google Scholar]
- 85.Alföldy P, Lemmel E-M. Reduction of nitroblue tetrazolium for functional evaluation of activated macrophages in the cell-mediated immune reaction. Clin. Immunol. Immunopathol. 1979;12:263–270. doi: 10.1016/0090-1229(79)90029-1. [DOI] [PubMed] [Google Scholar]
- 86.Kuang S-Y, et al. Effects of graded levels of dietary methionine hydroxy analogue on immune response and antioxidant status of immune organs in juvenile Jian carp (Cyprinus carpio var. Jian) Fish Shellfish Immunol. 2012;32:629–636. doi: 10.1016/j.fsi.2011.12.012. [DOI] [PubMed] [Google Scholar]
- 87.Harikrishnan R, Rani MN, Balasundaram C. Hematological and biochemical parameters in common carp, Cyprinus carpio, following herbal treatment for Aeromonas hydrophila infection. Aquaculture. 2003;221:41–50. doi: 10.1016/S0044-8486(03)00023-1. [DOI] [Google Scholar]
- 88.Rao YV, Das B, Jyotyrmayee P, Chakrabarti R. Effect of Achyranthes aspera on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila. Fish Shellfish Immunol. 2006;20:263–273. doi: 10.1016/j.fsi.2005.04.006. [DOI] [PubMed] [Google Scholar]
- 89.Elmada C, et al. The effect of dietary methionine on growth, antioxidant capacity, innate immune response and disease resistance of juvenile yellow catfish (Pelteobagrus fulvidraco) Aquac. Nutr. 2016;22:1163–1173. doi: 10.1111/anu.12363. [DOI] [Google Scholar]
- 90.Machado M, et al. Dietary tryptophan and methionine as modulators of European seabass (Dicentrarchus labrax) immune status and inflammatory response. Fish Shellfish Immunol. 2015;42:353–362. doi: 10.1016/j.fsi.2014.11.024. [DOI] [PubMed] [Google Scholar]
- 91.Dobrovolskaia MA, Aggarwal P, Hall JB, McNeil SE. Preclinical studies to understand nanoparticle interaction with the immune system and its potential effects on nanoparticle biodistribution. Mol. Pharm. 2008;5:487–495. doi: 10.1021/mp800032f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Gopalakannan A, Arul V. Immunomodulatory effects of dietary intake of chitin, chitosan and levamisole on the immune system of Cyprinus carpio and control of Aeromonas hydrophila infection in ponds. Aquaculture. 2006;255:179–187. doi: 10.1016/j.aquaculture.2006.01.012. [DOI] [Google Scholar]
- 93.Métayer S, et al. Mechanisms through which sulfur amino acids control protein metabolism and oxidative status. J. Nutr. Biochem. 2008;19:207–215. doi: 10.1016/j.jnutbio.2007.05.006. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data generated or analysed during this study are included in this published article.